

Зоологический журнал, 2020, T. 99, № 1, стр. 45-56
Численность и сезонное распределение западной популяции гаги-гребенушки (Somateria spectabilis), организация мониторинга в северных морях России
Ю. В. Краснов a, *, А. В. Ежов a, **, К. В. Галактионов b, c, ***, А. А. Шавыкин a, ****
a Мурманский морской биологический институт КНЦ РАН
183010 Мурманск, Россия
b Зоологический институт РАН
199034 Санкт-Петербург, Россия
c Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Россия
* E-mail: kharlov51@mail.ru
** E-mail: mr.haliaeetus51@mail.ru
*** E-mail: kirill.galaktionov@zin.ru
**** E-mail: shavykin@mmbi.info
Поступила в редакцию 23.07.2018
После доработки 08.03.2019
Принята к публикации 15.05.2019
Аннотация
На основе собственных полевых наблюдений 1990–2018 гг. и литературных источников описаны ареалы линьки и зимовки западной популяции гаги-гребенушки (Somateria spectabilis) в Баренцевом, Белом и Карском морях; представлены картосхемы районов концентрации птиц в эти периоды. Приведены данные учетов зимующих и линяющих птиц, их половозрастного состава в российской части ареалов. Показано, что в Белом море и южной части Баренцева моря в период 1994–2009 гг. зимовало от 6 до 10 тыс. уток данного вида. Зимой 2009 г. в этом районе наблюдали явное преимущество самцов – 70.2%, количество неполовозрелых особей составило 2.6%. В период линьки наиболее массовую концентрацию птиц регистрировали в юго-восточной части Баренцева моря. Общее количество линяющих гаг-гребенушек в этом районе – 18–20 тыс. особей. Выполнена оценка общей численности западной популяции в Баренцевоморском регионе. Без учета птиц, зимующих у берегов Новой Земли, этот показатель может составлять от 80 до 110 тыс. особей. Выявлены и продемонстрированы факторы, влияющие на размещение птиц в конкретные сезоны и их перемещение в пределах ареала. Предложены районы акваторий, пригодные для дальнейшего мониторинга, и адекватные способы учета численности гаги-гребенушки.
Акватории Баренцева, Карского и Белого морей играют важную роль в жизненном цикле морских водоплавающих птиц. Здесь линяют, зимуют и останавливаются в ходе миграций сотни тысяч морских уток (Бианки и др., 1975; Scott, Rose, 1996; Краснов, 2011).
Три вида гаг – типичных представителей орнитофауны данных бассейнов – занимают разные экологические ниши и стоят на вершинах трофических пирамид прибрежных морских экосистем. Два из них, обыкновенная (Somateria mollissima (L. 1758)) и сибирская (Polysticta stelleri (Pallas 1769)) гаги, находятся в центре внимания исследователей и являются объектами регионального мониторинга (Aarvak et al., 2012; Краснов и др., 2015).
Особенности экологии третьего вида ‒ гаги-гребенушки (Somateria spectabilis (L. 1758)) – таковы, что определение общей численности популяции и обнаружение районов массовой концентрации на разных отрезках годового цикла затруднены. Как следствие, существуют лишь отдельные литературные источники, в той или иной степени посвященные изучению западной популяции гаги-гребенушки (Успенский, 1979; Бустнес, Бианки, 2003; Краснов, 2016).
Основные места размножения западной популяции гаги-гребенушки располагаются восточнее п-ова Канин – на островах и в тундрах юго-восточной части Баренцева и в материковых тундрах Карского морей (Бустнес, Бианки, 2003). На Новой Земле гнездящиеся птицы встречаются приблизительно до 77о с.ш. (Исаков, 1952). Граница между гнездовыми ареалами западной и восточной популяций данного вида проходит на Центральном Таймыре, аналогично границе между районами размножения западной и восточной популяций сибирской гаги (Petersen et al., 2006; Bustnes et al., 2010; Соловьева и др., 2016). Таким образом, область размножения западной популяции гаги-гребенушки охватывает обширные пространства между п-овом Канин и Центральным Таймыром. Обычно утки этого вида размножаются в приморских тундрах, порой на значительном удалении от морских побережий, нигде не создавая крупных поселений, характерных для обыкновенной гаги. Вследствие этого дать точную оценку размеров гнездовой части популяции практически невозможно.
В отличие от других видов гаг, обитающих в узкой полосе моря у самого берега, гаги-гребенушки свободно распределяются на акватории прибрежных районов и могут удаляться от кромки берега в море на несколько километров. Зимой они встречаются у побережий Кольского п-ова в Баренцевом и Белом морях, у норвежского побережья Финнмарка, в Норвежском море ‒ у берегов провинций Тромс и Сёр-Трёнделаг (Бустнес, Бианки, 2003; Аарвак и др., 2011). Установлено, что птицы, зимующие у баренцевоморских побережий, размножаются в российских тундрах вплоть до Центрального Таймыра (Бианки, 1989; Bustnes et al., 2010). Таким образом, они и гребенушки, зимующие в северных районах Белого моря, относятся к российской западной популяции. Статус гаг-гребенушек из Норвежского моря не ясен. Допускается их принадлежность к российской западной популяции, в то же время не исключается, что здесь зимуют гаги-гребенушки со Шпицбергена и Гренландии (Bustnes et al., 2010).
Представленная работа является итогом многолетнего изучения комплекса морских уток в Баренцевом, Белом и Карском морях и естественным образом продолжает предыдущую публикацию, посвященную организации мониторинга обыкновенной гаги (Краснов и др., 2015). При подготовке статьи мы использовали как собственные материалы экспедиционных исследований, так и имеющиеся в литературе данные. На этой основе нами предпринята попытка оценить общую численность западной популяции гаги-гребенушки, определить основные районы ее линьки и зимовки, а также выявить факторы, воздействующие на размещение птиц в конкретных сезонах. Нами предложены районы, время и оптимальные методы учетов, пригодные для мониторинга в европейском секторе российской Арктики.
МАТЕРИАЛЫ И МЕТОДЫ
В настоящем исследовании представлены оригинальные материалы по гаге-гребенушке, полученные нами в период 1990–2018 гг. в ходе наземных, судовых, авиационных наблюдений в различных районах Баренцева, Белого и Карского морей. Во время полевых работ основное внимание уделялось сбору информации о численности и характере размещения птиц на открытых и прибрежных акваториях этих морей. Кроме материалов собственных экспедиционных наблюдений нами использованы данные из отечественных и зарубежных литературных источников.
Круглогодичные наземные наблюдения, начиная с 1998–1999 гг., осуществляли в вершине Кандалакшского залива Белого моря и в Кольском заливе Баренцева моря (Краснов, Горяев, 2013). Одновременно с ними, в течение нескольких лет производили наблюдения за морскими утками на участке побережья Восточного Мурмана протяженностью 60 км на восток от губы Териберская. Методы наземных наблюдений описаны в работе, посвященной орнитофауне Кольского залива (Краснов, Горяев, 2009). Во время экспедиций летом 2003, 2004 и 2007 гг. выполнены наблюдения с суши и маломерных плавсредств за прибрежными акваториями на Терском берегу Белого моря (Краснов, Горяев, 2012). Летом 1992 и 2016–2018 гг. – на западном побережье южного острова Новой Земли, в 2000, 2002–2004, 2007 и 2010 гг. – на ряде островов Печорского моря: Долгий, Вайгач и Колгуев. Методы полевых исследований отражены в соответствующих публикациях (Краснов, 2004, 2014; Краснов, Горяев, 2012).
Орнитологические наблюдения с борта морских судов на постоянной основе осуществляли с начала 2000-х гг. (Краснов, 2013). В том числе с судов ледового класса, включая атомные ледоколы, следующие по трассе Северного морского пути. Это позволило нам собрать информацию об орнитофауне восточных районов Баренцева и западной части Карского морей на протяжении всего годового цикла (Краснов и др., 2007, 2015а). Более детальное обследование различных районов Баренцева, Белого и Карского морей проводили с борта яхт и научно-исследовательских судов. В ходе таких экспедиций наиболее полно изучена акватория Белого моря (Краснов, 2012). Всего за годы исследований мы располагаем данными 77 серий орнитологических наблюдений с судна в Баренцевом море, 39 серий – в Карском море и 13 серий ‒ в Белом море. Использованные нами методы и особенности орнитологических наблюдений с борта морских судов представлены в специальных публикациях (Краснов и др., 2004a; Краснов, 2007).
Изучение орнитофауны и ее пространственного размещения на акваториях морей с использованием авиационной техники осуществляли с 1994 г. В прибрежных районах Мурмана и на акватории Белого моря ‒ с вертолета Ми-8, в открытых районах Баренцева моря – с самолетов Ил-18Д и Ан-26БРЛ (Краснов и др., 2002, 2013). С самолета орнитологическая информация получена в Баренцевом море в летне-осенние периоды 1994, 1995 и 1997-1999 гг.; с вертолета ‒ в ходе совместных российско-норвежских исследований зимой 1994, 2003 и 2009 гг.; летом 1998, 2003 гг. и осенью 2000 г. (Nygård et al., 1995; Isaksen et al., 2000). При наблюдениях с борта вертолета через открытую дверь производилось фотографирование встреченных птиц. Впоследствии эти материалы позволяли уточнить численность птиц в крупных стаях и в зимний период определить половозрастной состав популяции.
Методы, использованные при изучении морских птиц в открытых районах северных морей и их критический анализ, приведены в ряде работ (Краснов и др., 2002; Краснов, 2007; Krasnov, Nikolaeva, 2013).
В ходе комплексных судовых и наземных экспедиций в местах концентрации гаг-гребенушек регистрировали формы кормовой активности, производили сбор проб пищи и экскрементов; в районах массового кормления уток данного вида изучали качественный и количественный состав бентосных сообществ (Краснов и др., 2008; Sukhotin et al., 2008; Denisenko et al., 2019). При этом использовали количественные сборы бентосных организмов дночерпателем ван-Вина (площадь захвата 0.1 м2) и качественные – тралом Сигсби-Агассица (ширина 1 м) (Denisenko et al., 2019).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Районы зимовки
Основные районы зимовки гаг-гребенушек в северных морях России находятся у побережий Кольского п-ова в Баренцевом и Белом морях. В то же время результаты многолетних наблюдений с борта морских судов в зимний период также свидетельствуют о наличии в восточной части Баренцева и в Карском морях некоторого количества гаг-гребенушек (рис. 1, Краснов и др., 2007; 2015а). Однако из-за методических ограничений наблюдений с судна в зимних условиях полных данных о характере пребывания и численности встреченных птиц до сих пор получить не удается. Впервые предположение о зимовке гаг-гребенушек в полыньях и разводьях у западного побережья архипелага Новая Земля появилось еще в начале 1930-х гг. (Горбунов, 1932). Спутниковое слежение за несколькими помеченными птицами в 2008 г. показало, что отдельные самцы гаги-гребенушки способны зимовать в юго-восточной части Баренцева моря (Bustnes et al., 2010). Демме11 (1946) указывала, что в полыньях у Новой Земли гаги-гребенушки появляются в марте. В этот период основная масса птиц все еще находится в местах зимовки.
Рис. 1.
Районы распределения гаги-гребенушки в Баренцевом, Белом и Карском морях в зимний период по данным авиационных и судовых наблюдений: 1 – район зимовки у берегов Кольского п-ова по данным авиационных наблюдений (район рекомендуемого мониторинга); 2 – район предполагаемой зимовки гаг-гребенушек (необходимо обследование с борта вертолета); 3 – районы позднезимних встреч гаг-гребенушек по данным судовых наблюдений.
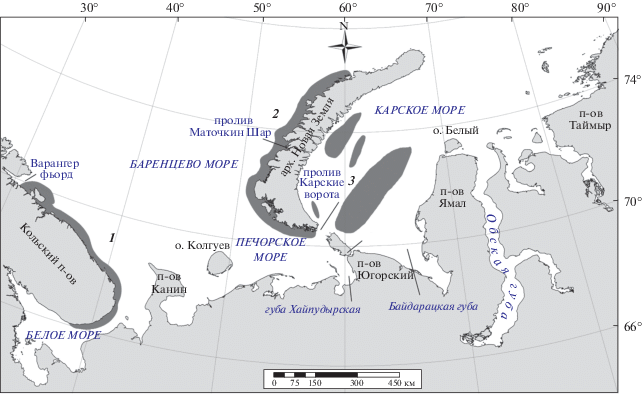
Это означает, что такие встречи в восточной части Баренцева и западной части Карского морей могут быть отражением локальных перемещений птиц в пределах зимовки (рис. 1). По всей видимости, эти перемещения обусловлены как состоянием окружающей среды (например, ледовой обстановки), так и поиском доступных калорийных пищевых ресурсов. Подобные перемещения мы неоднократно отмечали в различных районах общего зимовочного ареала. Передвижение птиц из открытых районов моря в укрытые губы из-за сильных штормов ‒ обычное явление.
В суровые зимы из районов Белого моря птицы могут перелетать в поисках свободных ото льда акваторий на большие расстояния. Например, в феврале 1979 г. группы неполовозрелых гаг-гребенушек в поисках открытой воды появились в окрестностях г. Кандалакша в вершине Кандалакшского залива, где и погибли от истощения. Позднее, в конце марта, кромка дрейфующих льдов необычно далеко продвинулась в западном направлении и достигла архипелага Семь островов на Восточном Мурмане. Массив битого льда, плотно заполнив все мелководье в восточных районах Кольского п-ова, вытеснил несколько тысяч гаг-гребенушек в район интенсивных приливно-отливных течений у острова Харлов. Ни в предыдущие, ни в последующие годы такого скопления птиц этого вида здесь не отмечали (данные Ю.В. Краснова).
При проведении наблюдений с вертолета у берегов Кольского п-ова и провинции Финнмарк в отдельные годы также наблюдали крупномасштабное перераспределение зимующих птиц. В некоторые зимы, например, в 1994 и 2009 гг., утки вытеснялись льдом из Белого моря в юго-западные районы Баренцева моря, где численность гаг-гребенушек, соответственно, возрастала (Nygård et al., 1995; Краснов и др., 2011).
В отдельные годы наблюдений во второй половине зимы при наличии массового нереста мойвы (Mallotus villosus (Müller 1776)) размеры отдельных стай гаг-гребенушек составляли 5–7 тыс. особей (Nygård et al., 1995; Бустнес, Бианки, 2003; Краснов и др., 2011). В северных районах Белого моря стай такой величины никогда не наблюдали, хотя зимующие здесь птицы нередко кормятся молодью сайки (Boreogadus saida (Lepechin 1774)) (Шкляревич, 1979). Этот вид рыбы в Белом море значимых концентраций не создает. Вероятно, по этой причине стаи зимующих на Белом море гаг-гребенушек никогда не превышали нескольких сотен особей (Краснов и др., 2006).
Половозрастной состав зимующих птиц западной популяции определяли дважды: в 2003 и 2009 гг. С борта вертолета Ми-8 у побережий Кольского п-ова в Белом и Баренцевом морях были получены серии фотографий, качество которых позволило определить пол и возраст гаг-гребенушек (Краснов и др., 2004; Ю.В. Краснов, М.В. Гаврило, неопубл. данные за 2009 г.).
В 2003 г. в северо-восточных и восточных районах прибрежья Кольского п-ова (от устья р. Стрельна до мыса Дворовой) половозрастной состав был определен у 1620 особей. В целом на учетном маршруте самцы составили 42.6% (690 особей), самки – 49.9% (812 особей) и 7.3% (118 особей) оказались неполовозрелыми. В 2009 г., при наиболее полном обследовании районов зимовки в Белом море и на Мурмане, из 1180 гаг-гребенушек с определенным возрастом и полом 2.6% (31 особь) были неполовозрелыми, среди взрослых птиц самки составили 27.2% (320 особей), самцы ‒ 70.2% (827 особей). Неполовозрелые гребенушки были встречены лишь на восточных участках учетного маршрута: в Онежском заливе и у Терского берега в Белом море, а также в самых восточных районах Мурманского побережья в Баренцевом море. Среди взрослых птиц доля самцов в российских районах зимовки постепенно снижалась в западном направлении от 75.0 до 41.0%, за исключением Кольского залива, где она неожиданно возросла до 73.4%.
Сравнение данных, полученных в 2003 и 2009 гг. на полностью совпадающем участке авиаучетов у северо-восточных и восточных берегов Кольского п-ова (устье р. Стрельна – мыс Дворовой), показало, что доля неполовозрелых птиц сократилась с 7.3 до 1.8%, а доля самцов, напротив, возросла с 42.6 до 62.7%.
Районы линьки
К местам линьки птицы мигрируют Восточно-Атлантическим пролетным путем (Scott, Rose, 1996). Для взрослых особей характерны летние послебрачные миграции. Наиболее четко они выражены у самцов по окончании половой активности и у холостых птиц. У неполовозрелых особей полная линька оперения со сменой маховых перьев может проходить либо в районах зимовки, либо, как и у взрослых птиц, на акваториях ближе к районам гнездования (Краснов, 2016). В районах зимовки в прибрежных водах у побережья Кольского п-ова на линьку остается лишь незначительное количество птиц (рис. 2, Краснов и др., 2002; Краснов и др., 2006). В отдельные годы в Кандалакшском заливе Белого моря могут формироваться сравнительно крупные линные скопления гребенушек общей численностью до одной тысячи особей (Бианки, 1989). Окольцованный в период линьки в Кандалакшском заливе взрослый самец был встречен в середине июня на северо-восточном побережье Ямала (Бианки, 1989). Одна из самок, помеченных спутниковым датчиком зимой 2008 г. у побережья Финнмарка, линяла у о-ва Вилькицкого между Обской губой и Енисейским заливом, другая самка – в Хатангском заливе (Bustnes et al., 2010).
Рис. 2.
Районы летне-осеннего размещения гаги-гребенушки: 1 – район линьки у берегов Кольского п-ова (данные авианаблюдений); 2 – район предполагаемой линьки (необходимо обследование с борта вертолета); 3, 4 – районы линьки в юго-восточной части Баренцева моря по данным авиационных наблюдений (район рекомендуемого мониторинга); 5 – районы встреч мигрирующих гаг-гребенушек; штриховая линия – граница между районами 3 и 4.
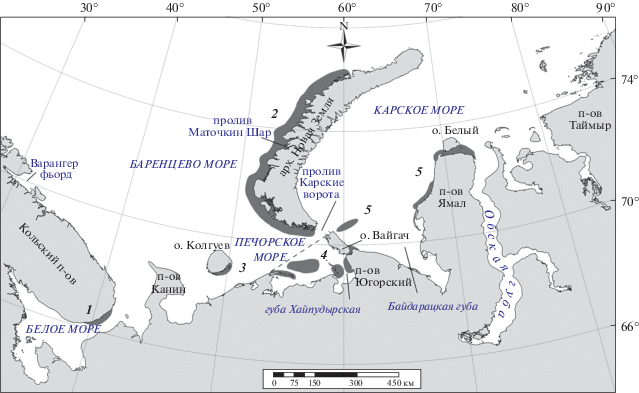
Районы линьки и постгнездовое распределение гаг-гребенушек у западного побережья Новой Земли почти не изучены. Линька взрослых особей в этом районе подтверждена нами в ходе визуальных наблюдений. Однако не ясно, линяют здесь только местные гаги-гребенушки или к ним присоединяются птицы из других районов гнездового ареала. Не известны характер распределения птиц, места их массовой концентрации и состав кормов линяющих птиц.
Наиболее известный район линьки западной популяции находится на мелководьях юго-восточной части Баренцева моря, где нами в 1997 г. обнаружены особенно крупные агрегации гаг-гребенушек (рис. 2). Основная их часть сосредоточена на мелководьях в районе южного побережья острова Долгий и Югорского п-ова (Краснов и др., 2004a). Именно здесь отдельные стаи в 1990-е гг. насчитывали до 16 тыс. особей (Isaksen et al., 2000; Краснов и др., 2002). Понятно, что столь крупные агрегации линяющих уток продолжительное время могут существовать только за счет значительных источников высококалорийного корма.
Специальные исследования трофических и паразитарных связей гаги-гребенушки в районе о-ва Долгий (Гаврило, Стрём, 2005; Краснов и др., 2008; Sukhotin et al., 2008; Galaktionov et al., 2019) показали, что основу существования скоплений морских уток обеспечивают сублиторальные поселения мидий (Mytilus edulis (L. 1758)) со своеобразной структурой и ограниченной продуктивностью. В этом районе мидии образуют на небольших глубинах (2‒11 м) разреженные поселения, которые характеризуются высокой агрегированностью (рис. 3) (Sukhotin et al., 2008; Denisenko et al., 2019). Биомасса мидий в агрегациях в 2007 г. достигала 4 кг/м2 (Sukhotin et al., 2008), и такие мидии более доступны для гаг, чем одиночные бентосные организмы. Этим и объясняется преобладание мидий в питании гаги-гребенушки в данном районе (Гаврило, Стрём, 2005; Краснов и др., 2008). Мидии в сублиторальных популяциях Печорского моря отличаются медленным ростом, успешное пополнение молодью происходит не каждый год (Sukhotin et al., 2008). Их поселения на распространенных здесь песчаных и галечных грунтах находятся под сильным воздействием факторов среды (Краснов и др., 2008; 2014; Denisenko et al., 2019).
Рис. 3.
Агрегации мидий, типичные для мелководий юго-восточной части Печорского моря (на фото моллюски, добытые при тралении в районе о-ва Долгий).

В последние годы в районе о-вов Долгий, Матвеев, Большой и Малый Зеленец наметилось снижение биомассы бентосных организмов-фильтраторов, включая мидий (Denisenko et al., 2019). Это связано с перераспределением донного осадка в результате волнового воздействия, вызванного усилением ветровой активности, а также под влиянием деятельности моржей, лежбище которых расположено на о-ве Матвеев и численность которых растет (Denisenko et al., 2019; Semenova et al., 2019). Резкая деградация поселений мидий в районе о-ва Долгого произошла в результате летних ураганов 2010 г. Это подорвало кормовую базу гаги-гребенушки и привело к отсутствию их крупных скоплений, что было отмечено в июле–августе 2014, 2016 и 2017 гг. (неопуб. данные А.А. Сухотина и К.В. Галактионова).
К 2016 г. поселения мидий частично восстановились, но прежних значений обилия так и не достигли (максимальная биомасса мидий отмечена на двух станциях и составила 86 и 68 г/м2) (Denisenko et al., 2019). По всей видимости, этим объясняется полное отсутствие гаги-гребенушки во второй половине лета 2017 г. в данном районе. Скопления птиц этого вида в то же время были отмечены в устьевой части Хайпудырской губы с борта научно-исследовательского судна, где при проведении тралений обнаружены плотные поселения мидии (неопуб. данные К.В. Галактионова, С.Г. Денисенко).
Пути миграций и миграционные стоянки
В конце зимы локальные перемещения гаг-гребенушек плавно перетекают в предбрачную миграцию. Крупные стаи половозрелых птиц в течение апреля постепенно продвигаются от мест зимовки в восточном направлении. У побережья Мурмана агрегации неполовозрелых особей встречаются вплоть до последней декады мая.
Визуально наблюдать весеннюю миграцию гаг-гребенушек удается очень редко. Движение групп этих уток регистрировали в апреле с борта судов у юго-восточного побережья Новой Земли, т.е. птицы проникали в Карское море через Карские Ворота. Но и в открытых районах Карского моря также в апреле регистрировали движение стай гаг-гребенушек. Вполне возможно, что в эти районы они могли попасть, проникая в Карское море через пролив Маточкин Шар. В это же время пролётные стаи размером до 200 особей встречали и вблизи о-ва Белый. Таким образом, доступная для визуальных наблюдений весенняя миграция гаг-гребенушек проходит через систему стационарных полыней в открытых районах Печорского и Карского морей. Стаи уток без остановок преодолевают сплошные ледовые пространства в промежутках между ними, при этом мигрирующие птицы огибают о-в Вайгач с севера. Птицы, помеченные спутниковыми передатчиками у побережья Финнмарка в 2008 г., приступили к миграции во второй декаде апреля (Bustnes et al., 2010). При этом некоторые меченые особи, двигаясь кратчайшим путем к местам размножения, совершали краткие остановки в открытых районах южной части Баренцева моря для отдыха. Продолжительные миграционные стоянки были зарегистрированы только в юго-восточной части Печорского моря (Bustnes et al., 2010). По данным спутникового слежения птицы проникли в Карское море через Югорский Шар.
Осенние перемещения гаг-гребенушек могут носить характер постепенного продвижения в сторону зимовок. В сентябре стаи уток, линявшие на мелководьях у о-ва Долгий, смещаются в западном направлении. Визуально начало миграции гаг-гребенушек наблюдали в конце сентября. В двадцатых числах сентября 1997 г. крупные скопления птиц этого вида обнаружены с самолета в открытых районах моря севернее Печорской губы и о-вов Гуляевские кошки (Краснов и др., 2002). Птицы отдыхали на поверхности моря. В начале октября 2018 г. с борта научно-исследовательского судна в Карском море наблюдали миграцию малой интенсивности. Утки небольшими группами двигались из района о-ва Белый генеральным курсом в западном направлении вдоль побережья над мелководьями. В районе центральной части западного побережья п-ова Ямал птицы разделялись. Часть групп двигалась в направлении проливов Карские ворота и Югорский Шар, огибая о-в Вайгач с севера и юга, а часть продолжала движение вдоль берега в южном направлении (рис. 2). В устье Югорского Шара были обнаружены небольшие группы гаг-гребенушек, состоящие из самок, молодых птиц и единичных самцов. Интенсивное движение птиц отмечали в первых числах октября 2000 г. с борта вертолета. Птицы, в подавляющем большинстве самцы, летели крупными стаями (до 1000 особей) от южного побережья Карского моря через пролив Карские Ворота в южную часть Печорского моря, где совершали промежуточные остановки, образуя крупные скопления (Краснов и др., 2002). К середине октября они достигают мест зимовки на Мурманском побережье Баренцева моря и районов зимовки в Белом море. По данным спутникового слежения осенняя миграция гребенушек, помеченных в период зимовки у берегов Финнмарка, шла двумя путями: птицы попадали в Баренцево море как через Карские Ворота, так и через Югорский Шар (Bustnes et al., 2010). Длительные остановки в ходе миграции они совершали в юго-восточной части Печорского моря, включая о-ва Долгий и Колгуев. Одна из птиц некоторое время кочевала у юго-западного побережья Новой Земли.
Численность
В пределах своего ареала западная популяция гаг-гребенушек наиболее компактно распределена в зимний период. Но даже и относительно ограниченная область зимовки включает в себя протяженные, с наличием сложного рельефа, побережья Норвежского, Баренцева и Белого морей. В таких условиях обследовать хотя бы половину зимовочного ареала затруднительно и порой просто невозможно. Как следствие, учетные работы проводили чаще всего на ограниченных, но доступных участках побережья в рамках национальных программ Норвегии и России. Многолетние наблюдения в отдельных губах и на отдельных участках Мурманского побережья показали, что результаты учетов с берега в большей степени отражают локальные перемещения птиц, а не изменения общей численности. Например, было зарегистрировано, что численность гаг (гребенушек и обыкновенных) в Кольском заливе Баренцева моря в течение одного месяца зимовки из-за погодных условий периодически менялась более чем в два раза (Краснов, Горяев, 2013). В итоге от трудоемких и малоинформативных учетов с берега мы вынуждены были отказаться в пользу наблюдений с борта вертолета.
В Норвегии усилиями многочисленных волонтерских групп подсчитывают зимующих уток с берега в большинстве доступных мест побережья (Бустнес, Бианки, 2003). Ясно, что при указанных выше различиях в методиках получения, данные национальных исследований мало сопоставимы между собой и в целом не позволяли корректно выявить существующие тенденции многолетней динамики численности западной популяции. Для определения количественных параметров большей части популяции было налажено тесное сотрудничество с норвежскими исследовательскими группами. В рамках совместных российско-норвежских экспедиций в 1994 и 2003 гг. осуществляли наблюдения, охватывавшие участок российского побережья Кольского п-ова, а в 2009 г. по унифицированной методике авиаучетов и норвежское побережье Финнмарка. В эти зимы количество птиц у берегов Кольского п-ова и прилегающих районов норвежского побережья Варангер-фьорда регистрировали на уровне 6–10 тыс. особей (Nygård et al., 1995; Краснов и др., 2011; наши неопубликованные. данные). В 1994 г. основная масса гаг-гребенушек держалась у северного побережья Кольского п-ова, в 2003 г. – у его восточного и северо-восточного побережья, а в 2009 г. – у норвежского побережья Финнмарка. Сравнение данных, полученных в 2003 и 2009 гг. на полностью совпадающем участке маршрутов у берегов Кольского п-ова (р. Стрельна – мыс Дворовой), показало, что количество зимующих гаг-гребенушек с 6540 сократилось до 2687 особей. С учетом полноты охвата наблюдениями участков побережий и точности самого метода было признано, что в 1994–2009 гг. общая численность гаги-гребенушки в районе зимовок у Кольского п-ова и Финмарка оставалась относительно стабильной.
В целом же, у норвежского побережья Финнмарка может зимовать до 30 тыс. птиц (Бустнес, Бианки, 2003). Общая численность гаг-гребенушек, зимующих у берегов Норвегии, оценивалась в 70–100 тыс. особей, при этом доля птиц со Шпицбергена и Гренландии не определена, хотя и считается незначительной (Бустнес, Бианки, 2003).
В летний период численность гаг-гребенушек наиболее высока в юго-восточной части Печорского моря. По данным авианаблюдений их общее количество в этом районе может составлять около 18–20 тыс. особей (Краснов и др., 2002, наши неопубликованные данные).
ЗАКЛЮЧЕНИЕ И РЕКОМЕНДАЦИИ
Из представленных выше результатов следует, что многотысячные скопления гаг-гребенушек традиционно зимуют у берегов Кольского п-ова, у норвежских побережий Баренцева и Норвежского морей, а в массе линяют они в юго-восточной части Баренцева моря. Наиболее компактно гаги-гребенушки располагаются в период зимовки в российских водах. Но и в российской части ареала до сих пор полностью отсутствуют количественные данные о численности птиц, зимующих и линяющих у западного побережья Новой Земли. Из-за логистических сложностей и суровых погодных условий возможности зимнего учета в этом районе сильно ограничены и, вероятно, в дальнейшем результатами этих учетов придется пренебречь. К настоящему времени установлено, что к западной популяции относятся птицы, зимующие в Баренцевом и Белом морях. Популяционная принадлежность гаг-гребенушек Норвежского моря до конца не выяснена (Bustnes et al., 2010). Эти пробелы в изученности вида осложняют оценку общей численности популяции и допускают высокую степень ее неопределенности. В настоящее время об общей численности популяции можно судить только на основании приблизительной оценки в 80–110 тыс. особей, возможно сильно заниженной, и не позволяющей корректно выявить многолетние тенденции ее изменений.
Как было показано нами выше, характерной особенностью гаг-гребенушек является подвижный образ жизни. В ходе многолетних наблюдений выявлены их перемещения в пределах ареала, определяемые не только штормовой активностью и ледовитостью, но и поиском доступных источников массовых кормов. Являясь типичными бентофагами, гаги-гребенушки активно используют и другие корма, нередко мелкую рыбу. Не очень ясно, каким образом они способны в короткие сроки обнаружить доступный косяк рыбы, чтобы сформировать стаю кормящихся птиц, состоящую из нескольких тысяч особей. Однако понятно, что пропуск такой стаи при количественном учете может кардинально изменить результаты всего исследования.
Обнаружение косяков рыбы морскими утками происходит, по-видимому, в период локальных перемещений, как мы неоднократно регистрировали это у обыкновенных гаг (Краснов и др., 2015). Стаи из нескольких тысяч уток формируются лишь в районах многолетнего активного нереста мойвы, например, в Варангер-фьорде, как было отмечено в марте 2009 г. (Аарвак и др., 2011). Иногда целенаправленное использование рыбы и ее икры гагами-гребенушками, как доступного и калорийного корма, ставится под сомнение. Тем не менее, явление это хорошо известно. Кроме наших визуальных наблюдений за поведением птиц на косяках мойвы, в литературе существует анализ кормов на основе данных вскрытия гаг-гребенушек. Продемонстрировано наличие в желудках как отходов рыболовного промысла (Bustnes, Galaktionov, 1999), так и нерестящейся мойвы и ее икры (Gjosaeter, Saetre, 1974; Nygård et al., 1988).
Гага-гребенушка включена в “Перечень видов флоры и фауны, являющихся индикаторами устойчивого состояния морских экосистем Арктической зоны РФ”, в связи с чем в нашей стране необходима организация ее мониторинга. Для корректного определения численности и ее долговременных изменений оптимальные условия существуют во время линьки и зимовки. Однако и в тот и другой период птицы держатся на обширных акваториях труднодоступных участков северных морей. В течение одного конкретного исследования недостаточность технического обеспечения не позволяет полностью охватить наблюдениями ареал зимовки или ареал линьки. В период наших наблюдений районы массового сосредоточения гаг-гребенушек регистрировали в разных частях ареалов зимовки и линьки. Например, выше нами было продемонстрировано, что в 2003 г. массовое скопление зимующих гаг-гребенушек (более 6 тыс. особей) существовало на Терском берегу Кольского п-ова. В 2009 г. численность птиц здесь сократилась до 2.7 тыс. особей, но одновременно их количество резко выросло у берегов Финнмарка в Норвегии. С общим падением численности у Терского берега в сильной степени изменился половозрастной состав гаг-гребенушек, зимующих в этом районе: резко сократилась доля неполовозрелых особей, а среди взрослых птиц увеличилась доля самцов.
При планировании мониторинга приходится учитывать вышеперечисленные факторы и поэтому основной проблемой становится выбор районов, доступных для регулярных наблюдений и достаточных, чтобы адекватно отражать изменения численности популяции в целом. Нами выделены и рекомендованы для дальнейшего мониторинга районы акваторий в южной части Баренцева моря, где традиционно существуют скопления линяющих и зимующих птиц (рис. 1, 2). Это побережья Кольского п-ова вплоть до государственной границы с Норвегией, а в оптимальном варианте, включая и побережье Финнмарка. Наиболее подходящее время для количественных учетов в этом районе – период зимовки. В юго-восточной части Печорского моря, где в районе Хайпудырской губы и у Югорского п-ова, на мелководьях западнее о-ва Долгий существует одно из основных мест массовой линьки гаг-гребенушек, целесообразно проводить учет во второй половине летнего периода.
При планировании мониторинга морских уток на акваториях морей неизбежно встает проблема выбора исследовательской платформы. Практика орнитологических исследований в море, особенно проводимых в интересах газо- нефтедобывающих компаний, в большинстве своем опирается на судовые наблюдения (Krasnov, Nikolaeva, 2013). Фактически при проведении подобных работ не учитываются поведенческие особенности морских уток и характер их распространения в море. С борта морского судна затруднительно корректно учитывать морских уток в обширных мелководных районах и в узкой прибрежной полосе. Даже если судно полностью работает в интересах орнитологических наблюдений с него проблематично осмотреть в полном объеме изрезанные губами и заливами побережья северных морей в ходе одной экспедиции. На открытых акваториях ряд видов морских уток, включая гагу-гребенушку, заблаговременно избегает судна, часто оставаясь за пределами зоны учета (Краснов, 2007). При таких наблюдениях трудно оценить численность крупных стай гаг-гребенушек с высокой плотностью распределения. Как правило, численность особей в таких стаях многократно недооценивается.
В ходе многолетних исследований (сухопутных, судовых и авиационных) и отработки методов наблюдений за птицами в море показано, что наиболее эффективным методом мониторинга гаги-гребенушки являются авиационные учеты (Краснов и др., 2002; Краснов, 2007; Krasnov, Nikolaeva, 2013). В рекомендованной для мониторинга гаги-гребенушки в юго-восточной части Баренцева моря в летний период оптимальной исследовательской платформой является самолет Ан-26 БРЛ или самолет подобного типа, а при учетах в зимний период у берегов Кольского п-ова и Финнмарка – вертолет. При наблюдении с борта вертолета необходимо использование фототехники высокого разрешения, что позволит получить данные о текущем половозрастном составе зимующих птиц.
Оптимальное решение задачи мониторинга западной популяции гаги-гребенушки может быть найдено в рамках тесной кооперации с норвежскими исследователями.
Список литературы
Аарвак Т., Краснов Ю.В., Гаврило М.В., Шавыкин А.А., 2011. Зимовки морских уток в Баренцевом и Белом морях: размещение и численность // Глобальные климатические процессы и их влияние на экосистемы арктических и субарктических регионов. Тез. докл. Междунар. науч. конф. (г. Мурманск, 9–11 ноября 2011 г.), Мурманский морской биол. ин-т Кольского науч. центра РАН. Апатиты: Изд-во Кольского научн. центра РАН. С. 7–8.
Бианки В.В., 1989. Гага-гребенушка // Миграции птиц Восточной Европы и Северной Азии. Пластинчатоклювые. М.: Наука. С. 213–215.
Бианки В.В., Коханов В.Д., Скокова Н.Н., 1975. Осенний пролет водоплавающих птиц на Белом море // Труды Кандалакшского государственного заповедника. Вып. IX. Мурманское кн. изд-во. С. 3–76.
Бустнес Я.У., Бианки В.В., 2003. Гага-гребенушка Somateria spectabilis // Состояние популяций морских птиц, гнездящихся в регионе Баренцева моря. Норвежский полярный институт. Отчет № 113. С. 51–53.
Гаврило М.В., Стрём Х., 2005. Рацион гаг-гребенушек (Somateria spectabilis), линяющих в Печорском море. // Гусеобразные птицы Северной Евразии. Тез. докладов Третьего междунар симпозиума (Санкт-Петербург, Россия. 6–10 октября 2005 г.). СПб. С. 72‒73.
Горбунов Г.П., 1932. Птицы Земли Франца-Иосифа // Труды Всесоюз. Арктич. ин-та. Т. 4. Л.: Гидрометеоиздат. 244 с.
Исаков Ю.А., 1952. Птицы Советского Союза (Гусеобразные). / Под ред. Дементьева Г.П., Гладкова Н.А. М.: Советская Наука. Т. 4. 640 с.
Краснов Ю.В., 2004. Позднелетние наблюдения 2002 г. за птицами прибрежных районов западного побережья о-ва Вайгач // Орнитология. М.: Изд-во Московского университета. Вып. 31. С. 228–231.
Краснов Ю.В., 2007. Методы исследования морских птиц в открытых районах моря // Методы и теоретические аспекты исследования морских птиц: Материалы V Всерос. шк. по морской биологии (25–27 октября 2006 г., г. Ростов-на-Дону). Ростов-на-Дону: Изд-во ЮНЦ РАН. С. 28–41.
Краснов Ю.В., 2011. Характеристика орнитофауны и исследование ее роли в экосистеме Баренцева моря на современном этапе // Комплексные исследования больших морских экосистем России. Апатиты: Изд-во Кольского научн. центра РАН. С. 258–277.
Краснов Ю.В., 2012. Морские птицы: современное состояние популяций, распределение и трофические связи // Биологические ресурсы Белого моря: изучение и использование. Исследования фауны морей. Т. 69 (77). СПб.: ЗИН РАН. С. 244–260.
Краснов Ю.В., 2013. История отечественных исследований морских птиц // Птицы северных и южных морей России: фауна, экология. Мурманский морской биол. ин-т КНЦ РАН. Апатиты: изд-во КНЦ РАН. С. 7–16.
Краснов Ю.В., 2014. Орнитофауна западного прибрежья о. Вайгач и прилегающих островов// Морские экосистемы и сообщества в условиях современных климатических изменений/ (отв. ред. Матишов Г.Г.). Мурманский морской биол. ин-т КНЦ РАН. СПб.: Реноме. С. 295–312.
Краснов Ю.В., 2016. Гага-гребенушка Somateria spectabilis // Миграции птиц Северо-Запада России. Неворобьиные / под ред. Носкова Г.А., Рымкевич Т.А., Гагинской А.Р. СПб.: Изд-во АНО ЛА “Профессионал”. С. 214–217.
Краснов Ю.В., Гаврило М.В., Аарвак. Т., 2011. Зимовки морских уток в Белом море и на Мурмане: характер размещения и адекватные методы учета // Гусеобразные Северной Евразии: география, динамика и управление популяциями. Тез докл. Междунар. конф. по гусеобразным Северной Евразии. Элиста. С. 46.
Краснов Ю.В., Гаврило М.В., Стрём Х., Шавыкин А.А., 2006. Численность и распределение птиц на прибрежных акваториях Кольского полуострова по данным авианаблюдений в позднелетний период 2003 года // Орнитология. М.: Изд-во Московского ун-та. Вып. 33. С. 125–137.
Краснов Ю.В., Гаврило М.В., Шавыкин А.А., 2015. Состояние, численность и организация мониторинга популяций обыкновенной гаги (Somateria mollissima) в Баренцевом и Белом морях // Зоологический журнал. Т. 94. № 1. С. 62–67.
Краснов Ю.В., Горяев Ю.И., 2009. Фауна морских и водоплавающих птиц // Кольский залив: освоение и рациональное природопользование. Мурманский морской биол. ин-т КНЦ РАН. М.: Наука. С. 264–284.
Краснов Ю.В., Горяев Ю.И., 2012. Итоги летних наблюдений за птицами на западном берегу Горла Белого моря // Экология морских птиц Белого моря. Апатиты: Изд-во КНЦ РАН. С. 105–128.
Краснов Ю.В., Горяев Ю.И., 2013. Основные тенденции развития авифауны Кольского залива и факторы ее определяющие // Птицы северных и южных морей России: фауна, экология. Мурманский морской биол. ин-т КНЦ РАН. Апатиты: Изд-во КНЦ РАН. С. 38–64.
Краснов Ю.В., Горяев Ю.И., Ежов А.В., 2007. Орнитологические исследования: ключевые районы и места концентрации морских птиц на акваториях Баренцева и Карского морей (по трассе Севморпути) // Биология и океанография Северного морского пути: Баренцево и Карское моря. М.: Наука. С. 124–129.
Краснов Ю.В., Горяев Ю.И., Ежов А.В., 2013. Характеристика орнитофауны на современном этапе // Птицы северных и южных морей России: фауна, экология. Мурманский морской биол. ин-т КНЦ РАН. Апатиты: изд-во КНЦ РАН. С. 16–38.
Краснов Ю.В., Горяев Ю.И., Шавыкин А.А., Николаева Н.Г., Гаврило М.В., Черноок В.И., 2002. Атлас птиц Печорского моря: распределение, численность, динамика, проблемы охраны. Апатиты: Изд-во Кольского научн. центра РАН. 164 с.
Краснов Ю.В., Ежов А.В., Горяев Ю.И., Баданин Ю.А., 2015а. Результаты наблюдений миграций и сезонных размещений морских уток с борта морских судов на трассе Севморпути // Гусеобразные Северной Евразии: изучение, сохранение и рациональное использование. Тез. Междунар. конф. (Салехард, Россия. 30 ноября–6 декабря 2015 г.). Салехард. С. 100–101.
Краснов Ю.В., Стрём Х., Гаврило М.В., Шавыкин А.А., 2004. Зимовка морских птиц в полыньях у Терского берега Белого моря и на Восточном Мурмане // Орнитология. М.: Изд-во Московского университета. Вып. 31. С. 51–57.
Краснов Ю.В., Черноок В.И, Гаврило М.В., Шавыкин А.А., Терещенко В.А., 2004а. Использование самолетов-лабораторий для мониторинга морских птиц и млекопитающих на акваториях арктических морей // Зоологический журнал. Т. 83. № 3. С. 330–341.
Краснов Ю.В., Шкляревич Г.А., Горяев Ю.И., 2008. Особенности питания гаги-гребенушки (Somateria spectabilis) на мелководьях у острова Долгий (юго-восток Баренцева моря) // Ученые записки Петрозаводского гос. ун-та. Изд.-во ПетрГУ. Петрозаводск. № 4. С. 37–40.
Краснов Ю.В., Шкляревич Г.А., Сухотин А.А., 2014. Особенности питания обыкновенной гаги Somateria mollissima на западом побережье о-ва Вайгач // Доклады Акад. наук. Т. 459. № 1, ноябрь. С. 125–127.
Соловьева Д.В., Краснов Ю.В., Контиокорпи Я., Антипин М.А., 2016. Сибирская гага Polysticta stelleri // Миграции птиц Северо-Запада России. Неворобьиные. Под ред. Носкова Г.А., Рымкевич Т.А., Гагинской А.Р. СПб.: Изд-во АНО ЛА “Профессионал”. С. 218–221.
Успенский С.М., 1979. Гаги СССР, состояние их изученности и некоторые задачи дальнейших исследований // Экология и морфология гаг в СССР. М.: Наука. С. 5–12.
Шкляревич Ф.Н., 1979. Зимовки обыкновенной гаги на Белом море // Экология и морфология гаг в СССР. М.: Наука. С. 61–67.
Aarvak T., Øien I.J., Krasnov Y.V., Gavrilo M.V., Shavykin A.A., 2012. The European wintering population of Steller’s eider Polysticta stelleri reassessed // Bird Conservation International. https://doi.org/10.1017/S0959270912000251
Bustnes J.O., Galaktionov K., 1999. Anthropogenic inXuences on the infestation of intertidal gastropods by seabird trematodes larvae in the southern Barents Sea coast // Mar Biol. 133:449–454
Bustnes J.O., Mosbech A., Sonne C., Systad G.H., 2010. Migration patterns, breeding and moulting locations of king eiders wintering in north-eastern Norway // Polar Biology V. 33. Issue 10. P. 1379–1385. https://doi.org/10.1007/s00300-010-0827-7
Denisenko S.G., Denisenko N.V., Chaban E.M., Gagaev S.Yu., Petryashov V.V., Zhuravleva N.E., Sukhotin·A.A., 2019. The current status of the macrozoobenthos around the Atlantic walrus haul-outs in the Pechora Sea (SE Barents Sea) // Polar Biology (published online) https://doi.org/10.1007/s00300-018-02455-3
Galaktionov K.V., Nikolaev K.E., Aristov D.A., Levakin I.A., Kozminsky E.V., 2018. Parasites on the edge: patterns of trematode transmission in the Arctic intertidal at the Pechora Sea (South‑Eastern Barents Sea) // Polar Biology (published online) https://doi.org/10.1007/s00300-018-2413-3
Gjosaeter J., Saetre R., 1974. Predation of eggs of Capelin (Mallotus villosus) by diving ducks // Astarte. V. 7. P. 83–89.
Isaksen K., Strøm H., Gavrilo M., Krasnov Yu.V., 2000. Distribution of seabirds and wildfowl in the Pechora Sea during August 1998 // Norwegian Ornithological Society. Report № 2. P. 7–38.
Krasnov Yu.V., Nikolaeva N.G., 2013. Ornithological Investigations and Estimation of Potential Threats in a Process of Off-Shore Oil and Gas Fields Exploitation in the Barents Sea (Russian). https://doi.org/10.2118/166958-RU
Nygård T., Larsen B.H., Follestad A., Strann K.-B., 1988. Numbers and distribution of wintering waterfowl in Norway. Wildfowl 39. P.164–176.
Nygård T., Jordhoy P., Kondakov A., Krasnov Yu.V., 1995. A survey of waterfowl and seal on the coast of southern Barents Sea in March 1994 // NINA Oppdragsmelding. № 361. Trondheim: NINA. 24 p.
Petersen M.R., Bustnes J.O., Systad G.H., 2006. Migration patterns, breeding- and moulting locations of Steller’s eiders wintering on the coast of Norway as determined from satellite telemetry // Journal of Avian Biology. V. 37. P. 58–68.
Scott D.A., Rose P.M., 1996. Atlas of Anatidae populations in Africa and Western Eurasia. Wetlands International Publication no. 41. Wageningen. The Netherlands. 336 pp.
Semenova V., Boltunov A., Nikiforov V., 2019. Key habitats and movement patterns of Pechora Sea walruses studied by satellite telemetry // Polar Biology (published online). https://doi.org/10.1007/s00300-018-02451-7
Sukhotin A.A., Krasnov Yu.V., Galaktionov K.V., 2008. Subtidal populations of the blue mussel Mytilus edulis as key determinants of waterfowl flocks in the southeastern Barents Sea // Polar Biology. V. 31. P. 1357–1363.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал