

Зоологический журнал, 2020, T. 99, № 12, стр. 1323-1344
Ревизия рода Mesocyclops Sars 1914 (Copepoda, Cyclopidae) мировой фауны
В. Р. Алексеев *
Зоологический институт РАН
199034 С. Петербург, Россия
* E-mail: alekseev@zin.ru
Поступила в редакцию 15.03.2020
После доработки 25.03.2020
Принята к публикации 10.04.2020
Аннотация
Весьма перегруженный видами род Mesocyclops Sars 1914 разделен на четыре подрода исходя из особенностей морфологии и ареалов. Подрод Neomesocyclops subgen. n. объединил виды, почти исключительно обитающие в Новом Свете. Единственным исключением является M. (N.) tenuisaccus (Sars 1927), описанный из Южной Африки, но, возможно, он является младшим синимом M. (N.) annulatus (Wierzejski 1892) из Южной Америки и появился в Африке в результате антропогенной инвазии из Нового Света. Подрод Tethymesocyclops subgen. n. включает виды, распространение которых совпадает с границами субконтинента Гондвана, а точнее – c побережьем моря Тетис периода отделения Гондваны от Лавразии. В состав этих двух подродов вошли виды с щетинкой на внутреннем выросте базального сегмента первой пары плавательных ног. Эта щетинка отсутствует у всех представителей номинативного подрода Mesocyclops s. str. с гладкими каудальными ветвями и последним торакальным сомитом, а также у нового подрода Pilosomesocyclops subgen. n., имеющего волоски на этих структурах. Большинство видов этих двух подродов обитают в водоемах Старого Света. Наряду с подробной характеристикой новых подродов приводится описание нового вида Mesocyclops (Neomesocyclops) frankfiersi sp. n. В рамках ревизии виды в двух наиболее крупных подродах Neomesocyclops subgen. n. и Pilosomesocyclops subgen. n. дополнительно разделяются на три секции по наличию у них волосообразных структур на последнем торакальном сомите и/или каудальных ветвях. Названия секциям даны по имени первого вида, описанного с этими признаками. Такое структурирование рода позволяет значительно упростить определение видов мировой фауны и в перспективе дополнять ключи при описании новых таксонов. По сравнению с последней фаунистической сводкой Рейд и Уэда приведенные ключи расширены на 12 таксонов, описанных в последовавшие за выходом этого определителя годы.
Род Mesocyclops Sars (Copepoda: Cyclopidae) насыщен видами, и процесс описания новых форм продолжается весьма интенсивно, что приводит к необходимости частых ревизий рода (Kiefer, 1981; Van de Velde, 1984; Alekseev, 2002; Holynska, 2003). В основе видового многообразия этого рода лежит, с одной стороны, его всесветное распространение. По мере замены парадигмы космополитизма на представления о видах-близнецах или супервидах, объединяющих близкородственные, но изолированные группировки морфологически близких форм (Kiefer, 1981) с вытекающими отсюда вариантами изоляции (горообразование, движение континентов, формирование ледников и их таяние), число описываемых видов существенно возросло.
С другой стороны, к этому роду в прошедшем веке было привлечено усиленное внимание экологического сообщества, благодаря заметной роли, которую он повсеместно играет в пелагических экосистемах континентальных водоемов (Gophen, 1984; Alekseev, 2002; Reid, Ueda, 2003; Meleg et al., 2012).
Последняя ревизия мировой фауны Mesocyclops, выполненная Марией Холинской (Holynska, 2003), свела воедино свыше 60 видов рода. В ходе исследования филогенетических связей число рассмотренных таксонов с учетом подвидов составило свыше 70 (Holynska, 2006). С таким массивом видов в рамках одного не поделенного рода весьма сложно работать, к тому же вследствие продолжающегося активного описания новых таксонов, прежде всего из мало изученных районов Юго-Восточной Азии, Южной и Северной Америк, последний ключ мировой фауны рода, составленный Марией Холинской (Holynska, 2003), устарел. Важные работы по филогенетической структуре рода Mesocyclops (Holynska, 2006; Wyngaard et al., 2010) подтвердили как валидность многих мало различимых форм, так и монофилию рода, но не решили вопроса об упрощения определительных ключей. В связи с участившимися случаями межконтинентальных биологических инвазий и наблюдающимся расселением тропических таксонов в водоемы более высоких широт, обусловленным климатическими изменениями последних лет (Alekseev et al., 2009; Fefilova et al., 2014; Montoliu et al., 2015; Alekseev et al., 2020), вопрос о проведении кардинальной ревизии данного рода стал весьма актуален.
Незадолго до выхода определителя под редакцией Дженет Рейд и Хироши Уэда (Reid, Ueda, 2003) мною был проведен предварительный анализ данного рода. Анализ показал сложную структуру этого рода и устойчивые различия в морфологии, достаточно хорошо связанные с современным распределением видов по континентам (Alekseev, 2002). Такое соотношение морфологической и географической дивергенции вполне можно было бы свести, по меньшей мере, к четырем подродам. За прошедшие годы подобного разделения так и не было сделано, что затрудняет дальнейший прогресс в изучении рода. Настоящая работа выполнена для решения этих и других накопившихся таксономических и систематических проблем рода Mesocyclops.
В ходе выполнения этой ревизии рода и разбиения его на подроды был изучен материал островной фауны. В этом материале, собранном мною ранее в районе Атлантического побережья Мексики, был обнаружен и описывается ниже новый для науки вид.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использован значительный материал, собранный лично автором во время экспедиций по России (от Калининграда до Камчатки и от дельты Лены до Тувы) и в следующих странах: Германия (1992 – Боденское озеро; 1996, 2000–2003 – озера Голштинии, реки Рейн, Одер и другие, впадающие в Северное море), Монголия (1995 – озера Гоби, 2000 – озеро Хубсугул, реки Тоуул, Селенга), Испания (2005 – р. Гвадалквивир, Севилья; 2006–2013, 2019 – водоемы Кастилии и Валенсии), Канада (1997 – водоемы пров. Торонто; 1998–2000, 2016–2017 – озера пров. Квебек), США (1996 – оз. Эри; 2016 – Мидоулэйк, Нью-Йорк, включая типовой материал для подрода Neomesocyclops subgen. n. из Великих озер), Мексика (2009 – водоемы в городах Мехико и Агуаскальентес; 2011 – п-ов Юкатан и о-в Косумель), Сингапур (1998), Эфиопия (2003 – водосборы и верховья Белого и Голубого Нила), Малайзия (2009 – Малаккский п-ов, 2011 – о-в Борнео), Египет (2003 – Нил в районе Каира), Тунис (2002 – приморские водоемы гор Атлас), Турция (2012–2019 – реки и озера Анатолийского побережья), Лаос (2013 – р. Меконг), Камбоджа (2013 – водосбор Меконга), Вьетнам (2011 – р. Сайгон; дельта р. Меконг; 2013 – городские пруды в Дананге и Ханое), Таиланд (2002 – г. Кхон Каен; 2013 – водосбор р. Меконг), Индонезия (2009 – Джакарта, Ява), Китай (2011 – пруды и реки в Пекине и Шанхае; 2013 – реки Хуанхэ, Жемчужная, о-в Хайнань), Тайвань (2005–2009 – оз. Луны и Солнца, водоемы по всему острову), Япония (2010 – пров. Киото, Хиросима, Токио). Весь материал передан на хранение в федеральную коллекцию Зоологического института РАН № 96-03-16.
В работе был также использован материал из коллекций тропического зоопланктона проф. Ф. Кифера (место хранения – Музей Естественной истории, Карлсруэ, Германия), С.Н. Фернандо (Музей Сингапурского Университета, Сингапур), Г. Дюмона (Кафедра гидробиологии Гентского Университета, Бельгия), Б. Дюссара (Музей Естественной Истории, Париж, Франция). В совокупности просмотренный материал охватывает все континенты и важнейшие водоемы большинства стран мира.
Для определения видов, описания новых подродов и типовых видов для них изготовляли постоянные препараты по методу проф. Кифера, для чего отпрепарированные в глицерине организмы накрывали покровными стеклами с восковыми ножками, глицериновую каплю, занимающую не более четверти общей площади, обводили канадским бальзамом с вытеснением воздуха со всех сторон. При соблюдении этих условий после затвердевания бальзама, которое занимает около двух недель, препарат может храниться неограниченное время. Фотографирование, зарисовку и исследование морфологии таксономически важных структур копепод проводили на микроскопах с использованием масляной и водной иммерсии при увеличении до ×1200, оптики Намарского (с увеличением до ×800) и лазерной конфокальной микроскопии с естественной люминесценцией хитина (увеличение до ×1000). Рисунки выполняли карандашом и обводили капиллярными фломастерами толщиной 0.1–0.15 мм, сканировали с разрешением не ниже 600 dpi, масштабировали и редактировали с использованием компьютерной программы Adobe Photoshop CS5.
Публикация зарегистрирована в базе данных Zoobank под номером LSID: urn:lsid:zoobank.org: pub:2235D007-2FCE-436C-A1B1-E33A2361B854.
Использованные признаки, термины и сокращения (рис. 1)
Рис. 1.
Признаки, имеющее наибольшее значение в систематике рода Mesocyclops, и их обозначения: a – соотношение каудальных щетинок; b – соотношение щетинок P5; c – строение P4; d – семяприемник, тип V; e, f – семяприемники типа Т с изогнутым и прямым поровым каналом; g – каудальная орнаментация A2; h – наличие латеральных волосков на Th4; i – наличие внутренней щетинки базиподита P1.
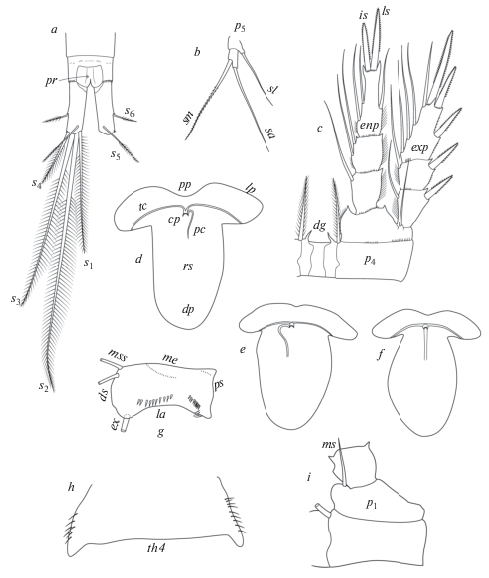
Наибольшее значение при определении и описании видов этого рода имеют следующие морфологические признаки, наблюдаемые у половозрелых самок (в скобках даны использованные сокращения):
Антеннулы (А1): длина относительно просомы, конструкция гиалиновой пластинки на дистальном сегменте;
Антенны (А2): орнамент из шипиков и волосков на базиподите (Bas) и число щетинок на втором сегменте эндоподита (Enp2); me – медиальный, la – латеральный, ds – дистальный, ps – проксимальный края; mss – медиальные щетинки, ex – экзоподитная щетинка;
Первая пара плавательных ног (Р1): наличие/ отсутствие щетинки на внутреннем выросте базиподита и его опушение волосками; ms – внутренняя щетинка базиподита;
Четвертая пара плавательных ног (Р4): длина и форма выростов (dg) по наружному краю соединительной пластинки коксы (CoxPl), соотношение длины и наибольшей ширины дистального членика эндоподита (Enp3P4), соотношение длин наружного (ls) и внутреннего (is) концевых шипов этого членика (lo/li), отношение длины наружного шипа к длине самого членика (lo/Ls), присутствие шипиков по наружным краям первого (проксимального) сегмента экзоподита (Exp1P4);
Рудиментная пятая пара ног (Р5): соотношение длин медиального шипа (sm) и концевой (апикальной) щетинки (sa) дистального членика, а также соотношение длин медиального шипа дистального членика и латеральной щетинки (sl) базального членика;
Последний торакальный сомит (Th4): наличие/отсутствие волосоподобных щетинок по наружному краю;
Генитальный двойной сомит уросомы (GDS): отношение длины к наибольшей ширине; форма семяприемника (RS); форма и направление изгиба порового канала (pc), рукавов (lp) и угол, под которым соединяются поперечные каналы(tc); pp – проксимальная часть, dp – дистальная часть, cp – копуляторная пора;
Каудальные ветви (CR): наличие/отсутствие волосков или шипиков по внутренней поверхности; отношение длины к ширине (L/W); соотношение длин внутренней и наружной щетинок; соотношение длин дорсальной и наружной щетинок; соотношение длин внутренней щетинки и ветви (li/Lcr); наличие шипиков в местах прикрепления боковой (s6) и наружной (s4) щетинок; орнаментация проктодеума (pr).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Род Mesocyclops Sars 1914
Анализ мировой фауны рода Mesocyclops
Представители рода Mesocyclops были отмечены повсеместно на всех континентах кроме Антарктиды (Dussart, Defaye, 2006). В свое время с этим было связано выделение вида-космополита M. leuckarti (Claus 1857), который в настоящее время распался более чем на 15 видов (Holynska, 2003). В последней ревизии рода число валидных видов было сведено к 66 (Holynska, 2003). Mesocyclops arakhlensis Alekseev 1993 Холинска (Holynska, 2003) свела в младшие синомимы M. leuckarti, однако по результатам проведенных гибридизационных экспериментов эти таксоны оказались генетически изолированы (собственные данные, неопубликованные). Найденные впоследствии мною особенности в строении эндоподитов четвертой пары плавательных ног позволяют их легко различить (см. ключ рода Mesocyclops) (Alekseev, 1993, 1998). Вместе с описанными за последние 17 лет 11 новыми видами, включая описанный ниже M. frankfiersi sp. n., количество видов этого рода в настоящее время составляет 78. Представляется вполне вероятным, что в ближайшее время следует ожидать дальнейшего расширения видового состава этого древнего (судя по всесветному распространению) рода, который и так явно перегружен. Анализ морфологии и распределения видов по странам света позволяет предложить вполне логичную группировку видов по некоторым устойчивым и легко находимым морфологическим признакам, что может быть использовано для разделения рода на подроды.
Первая такая группировка была произведена по наличию либо отсутствию щетинки на внутреннем выросте базального сегмента первой пары плавательных ног. По этому признаку представители рода распадаются на две группы, что было отмечено уже довольно давно (Alekseev, 2002). Дальнейшее деление внутри этих групп идет по отсутствию либо присутствию волосообразных структур на латеральной поверхности 4-го свободного торакального сомита и/или на внутренней поверхности каудальных ветвей.
В итоге в составе рода выделяются четыре следующих подрода:
1. Представители типового подрода Mesocyclops (Mesocyclops) – не имеют щетинки на базеподите Р1 и волосков на Th4 и CR.
2. Представители подрода Mesocyclops (Pilosomesocyclops) subgen. n. – не имеют щетинки на базеподите Р1, но есть волоски на Th4 и/или CR.
3. Представители подрода Mesocyclops (Neomesocyclops) subgen. n. – обладают щетинкой на базеподите Р1 и волосками на Th4 и/или CR.
4. Представители подрода Mesocyclops (Tethymesocyclops) subgen. n. – со щетинкой на базеподите Р1, но без волосков на Th4 и CR.
Наиболее крупные подрода (с волосками на Th4 и/или CR) можно разделить на три естественные морфологические группы или секции, основываясь на легко наблюдаемых признаках – присутствию волосков одновременно на Th4 и CR либо на одной из этих структур. Эти секции, названные по имени первого из описанного с такими признаками вида, были использованы при составлении ключей и позволяют не только упростить ключи для подрода, но и сделать некоторые предсказания о перспективе расширения их видового состава. В частности, такие секции, как reidae и edax во вновь выделенном подроде Mesocyclops (Neomesocyclops) subgen. n., а также pseudospinosus из подрода Mesocyclops (Pilosomesocyclops) subgen. n. содержат по одному-двум видам и, по-видимому, число таксонов в этих секциях сильно недооценено, прежде всего за счет тропической и экваториальной фаун.
Следует указать, что большинство видов, которые относятся к вновь выделяемому подроду Mesocyclops (Neomesocyclops) subgen. n., были описаны из Неотропиков, что также оправдывает его образование. Подрод Mesocyclops (Tethymesocyclops) subgen. n. (со щетинкой на базиподите P1 и без волосков на Th4 и каудальных ветвях) наибольшее количество представителей имеет в Старом Свете, а именно – Африке, Индии и Австралии, которые, в соответствии с теорией движения материков около 100 млн лет назад были частью единого материка Гондваны. Описанный недавно из подземных вод Вьетнама M. sondoongensis Tran et Holynska 2015 вполне возможно представляет собой реликтовый вид островной фауны моря Тетис, которая, вследствие сближения материковых плит азиатского и индийского континентов, оказалась в составе азиатской материковой платформы. Во всяком случае, существующие модели движения материковых плит это подтверждают (Besse, 1995).
Вероятным механизмом появления представителей неотропического подрода Neomesocyclops в Старом Свете могли стать перевозки воды между континентами, изначально питьевой, в периодически освежаемых бочках парусного флота, а позднее, с 1860 г., в подсланевых водах и в балластных отсеках морских судов. Описанный из Южной Африки M. tennuisaccus (Sars 1927) вплоть до мелких деталей похож на массовый южноамериканский вид и вполне мог попасть в Африку из Латинской Америки, которая на протяжении нескольких веков была основой работорговли и перевозки рабов на плантации Нового Света. Подобное предположение высказала также Холинска в результате проведенных филогенетических исследований (Holynska, 2006). Занос инвазийных видов подрода Mesocyclops во встречном направлении был отмечен в недавних работах – находки M. ogunnus в Бразилии (Peixoto et al., 2010) и M. pehpeiensis в Северной Америке (Montoliu et al., 2015; Connoly et al., 2019).
Ключ для определения подродов рода Mesocyclops
1. Базиподит P1 не имеет щетинки на внутреннем выросте....................................................... (2)
– Базиподит P1 несет щетинку на внутреннем выросте............................................................... (3)
2. Латеральная поверхность Th4 и медиальная поверхность CR не покрыты волосками...... Mesocyclops (Mesocyclops)
– Латеральная поверхность Th4 и/или медиальная поверхность CR покрыты волосками..... Mesocyclops (Pilosomesocyclops) subgen. n.
3. Латеральная поверхность Th4 и/или медиальная поверхность CR покрыты волосками..... Mesocyclops (Neomesocyclops) subgen. n.
– Латеральная поверхность Th4 и медиальная поверхность CR не покрыты волосками..... Mesocyclops (Tethymesocyclops) subgen. n.
Подрод Mesocyclops (Mesocyclops)
Диагноз. Подрод включает виды со следующей комбинацией признаков: базиподит P1 без щетинки на внутреннем выросте, волоски на латеральной поверхности 4 свободного сомита торакса и внутренней стороне каудальных ветвей отсутствуют, изредка вместо них встречаются короткие шипики, зубчики или ямки. Представитель подрода M. leuckarti – типовой вид также и для всего рода, который в недалеком прошлом считался видом-космополитом. В состав подрода входят 17 видов, описанных главным образом из водоемов Европы и Северной Азии (4 вида), Австралии(3 вида), Африки (2 вида), Юго-Восточной Азии, включая Китай и Японию (8 видов), Северной Америки (2 вида). Ни один вид этого подрода не показан для Южной Америки. Типовой вид M. leuckarti очень подробно переописан при последних ревизиях (Van de Velde, 1984; Dahms, Fernando, 1993). Ниже дана краткая его характеристика, поскольку это необходимо для выделения нового подрода. Типовой вид – M. (M.) leuckarti (Claus 1875).
Описание типового вида Mesocyclops (Mesocyclops) leuckarti (Claus 1857)
Самка (рис. 2). Длина тела без щетинок 700–1300 мкм. 4 свободный торакальный сомит лишен волосков. Длина генитального двойного сомита и его ширина соотносятся как 1.2–1.5 : 1. Он также лишен волосков. Семяприемник с широкими относительно короткими боковыми лопастями, слегка изогнутыми книзу, и широким нижним отделом. Семяпровод очень короткий, изогут внутрь. Каудальные ветви гладкие, лишены волосков, довольно длинные, отношение длины к ширине 3–4. У мест вставки боковой и наружной щетинок имеются шипики. Концевые щетинки крепкие и густо оперены волосками, соотношение их длин начиная от наружной: 1/2.5–2.9/3.1–3.8/2–2.6. Спинная щетинка короче наружной. Наибольшая из каудальных щетинок короче уросомы. Антеннула 17-сегментная, достигает 3 торакального сомита, ее концевой членик несет широкую пилообразную гиалиновую мембрану с 1 крупной зазубриной в середине. Антенны с 3-члениковым эндоподитом, базиподит по обеим сторонам несет лишь основные группы зубчиков. Второй членик эндоподита с 7 щетинками. Ротовые конечности типичного для циклопид строения. Плавательные ноги 1–4 пар двуветвистые и трехчлениковые. Базиподит 1 пары без щетинки на внутреннем выросте. Коксальная пластинка без орнаментации, у ног 4 пары по наружному краю с длинными заостренными палочковидными выростами. Дистальный членик эндоподита 4-й пары длинный и узкий с отношением длины к ширине около 3, на конце с двумя крепкими шипами, внутренний из которых слегка короче наружного. Рудиментная Р5 двучлениковая, базальный членик с короткой оперенной щетинкой, конечный членик удлиненный, на конце с оперенной апикальной щетинкой, которая несколько длиннее крепкой медиальной шипообразной щетинки.
Рис. 2.
Mesocyclops (Mesocyclops) leuckarti (Claus 1857): a – каудальные ветви, дорсально; b – генитальный сомит; c – P5; d – последние сегменты A1; e – базиподит А2, каудально; f – базиподит А2, фронтально; g – P1; h – Р5. Масштаб (мкм): a–c, g, h – 100; d – 70; e, f – 60 (по: Van de Velde, 1984).
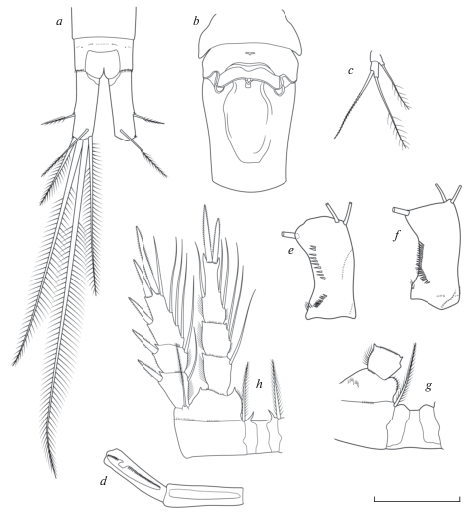
Самец. Длина тела без концевых щетинок 750–950 мкм. Сперматофоры окрашены в светло желтый цвет. Просома и уросома соотносятся как 1.8 : 1, отношение длины цефалосомы к ширине 1.25 : 1. Четвертый свободный торакальный сомит лишен волосков. Каудальные ветви короче, чем у самок, без волосков на внутренней поверхности. У мест вставки боковой и наружной щетинок имеются шипики. Геникулирующие антеннулы 14-члениковые хватательного типа. Второй членик эндоподита антенны с 7 щетинками. Конструкция плавательных ног такая же, как у самки. Рудиментная 5-я пара: апикальная щетинка длиннее, чем шипообразная медиальная щетинка, щетинка базального членика заметно короче обоих придатков дистального членика. Шестая рудиментная пара представлена широкой пластинкой с тремя придатками, из которых внутренний шип почти равен по длине средней и почти в два раза короче наружной щетинок.
Распространение. Палеарктика (?), восточнее Байкала возможно замещен близкородственными видами (Alekseev, 1998).
Ключ для определения видов подрода Mesocyclops (Mesocyclops)
1. Внутренняя щетинка каудальных ветвей (s1) очень короткая не более чем в 1.5 раза длиннее наружной (s4)..................................................... (2)
– Внутренняя щетинка длиннее................... (3)
2. Последний членик эндоподита Р4 очень короткий с L/W примерно 1.8; RS V-типа....M. (M.) brevisetosus Dussart et Sarnita 1987 (Индонезия, Калимантан)
– Этот членик длиннее (L/W 2.2–2.5); RS Т-типа.......M. (M.) yenae Holynska 1998 (Вьетнам)
3. Внутренний шип Р5 очень короткий, составляет менее половины длины апикальной щетинки, выросты пластинки коксы Р4 короткие и округлые.......................M. (M.) cuttacuttae Dumont et Maas 1985 (Австралия)
– Этот шип заметно длиннее, выросты заостренные................................................................ (4)
4. Медиальный вырост базиподита P4 без волосков................................................................. (5)
– Медиальный вырост базиподита Р4 с волосками.................................................................... (6)
5. Каудальная поверхность базиподита А2 с дистальным полем мелких шипиков у основания медиальных щетинок; за подковообразной копуляторной порой имеются 2 круглые поры...M. (M.) papuensis Van de Velde 1987 (Новая Гвинея, Австралия, Индонезия)
– Каудальная поверхность базиподита А2 без подобного поля шипиков; за подковообразной копуляторной порой находится 1 круглая пора или же последней вообще нет.....M. (M.) pehpeiensis Hu 1943 (Китай, Узбекистан)
6. Волоски на медиальном выросте Р4 расположены проксимально, дистально же имеется ряд мелких шипиков.........M. (M.) kieferi Van de Velde 1984 (Африка)
– Волоски на медиальном выросте Р4 расположены дистально............................................. (7)
7. Соединительная пластинка Р4 с волосками на каудальной поверхности..........M. (M.) salinus Onabamiro 1957 (Африка)
– Соединительная пластинка Р4 без волосков.................................................................(8)
8. Коксальный шип Р4 короткий и не достигает дистального конца внутреннего выроста базиподита...........M. (M.) restrictus Dussart et Fernando 1985 (Мньяма)
– Коксальный шип Р4 длинный, он заходит за дистальный край выроста базиподита...............(9)
9. Каудальная поверхность базиподита А2 с косым рядом шипиков, расположенных в середине у медиального края.......................................... (10)
– Каудальная поверхность базиподитов А2 без подобного ряда................................................. (15)
10. На каудальной поверхности А2 у основания медиальных щетинок имеется группа мелких шипиков и треугольное поле мелких шипиков проксимальнее него..........M. (M.) ferjemurami Holynska et Nam 2000 (колодцы во Вьетнаме)
– На каудальной поверхности А2 у основания медиальных щетинок шипиков нет..................(11)
11. Каудальные ветви относительно короткие, L/W < 3....................M. (M.) pescei Petkoski 1986 (Багамские о-ва)
– Каудальные ветви длиннее, L/W > 3........(12)
12. Поровый канал длинный и узкий, с боковым изгибом..............M. (M.) yesoensis Ishida 1999 (Япония, Малайзия)
– Этот канал очень короткий и широкий, прямой при просмотре препарата строго вентрально без бокового разворота.....................................(13)
13. Шип Р5 заметно короче апикальной щетинки и примерно равен по длине латеральной...................M. (M.) bodanicola Kiefer 1929 (Европа, крупные постледниковые озера)
– Этот шип равен апикальной щетинке и явственно длиннее латеральной щетинки...........(14)
14. Каудальные ветви относительно короткие, L/W < 4; все членики ExpР4 по наружному краю с волосками...............M. (M.) leuckarti (Claus 1857) (Палеарктика ?)
– Каудальные ветви длиннее, L/W > 4; первый (проксимальный) членик ExpР4 по наружному краю с крепкими шипиками.....M. (M.) arakhlensis Alekseev 1993 (Забайкалье)
15. A2 enp2 с 7 или 8 щетинками; размер рачка 0.7–0.8 мм; на каудальной поверхности А2 латеральный ряд сильно редуцирован и состоит из 2–7 мелких шипиков...........M. (M.) tobae Kiefer 1933 (Суматра, Индонезия)
– A2 enp2 с 6 щетинками; размер рачка 0.9–1.2 мм; на каудальной поверхности А2 латеральный ряд состоит примерно из 10 крупных шипов.....M. (M.) americanus Dussart 1985 (Канада)
Подрод Mesocyclops (Pilosomesocyclops) Alekseev subgen. n.
Zoobank LSID: urn:lsid:zoobank.org:act:4673BFF6-62CE-4219-B5E5-3074E60286AA
Диагноз. Наиболее богатый по числу таксонов подрод. Он включает 35 видов, у которых отсутствует щетинка на внутреннем выросте базиподита Р1, но имеются волоски на латеральной поверхности 4-го торакального сомита и/или на внутреннем крае каудальных ветвей, что и определило его название (pilosus (lat.) – волосатый). Ввиду большого числа описанных видов, этот подрод на основе легко наблюдаемых признаков подразделен на три секции: секция aspericornis (9 видов в Тропиках Старого Света), секция pseudospinosus (2 вида в Юго-Восточной Азии) и секция thermocyclopoides (24 вида, практически все из тропиков и субтропиков Старого Света). Последняя, самая крупная, секция также может быть разделена на три подсекции по 6–10 видов в каждой (обозначены в общем ключе). Получившаяся система ключей существенно облегчает определение уже известных видов и выявление новых для науки таксонов. Типовой вид для подрода М. (P.) aspericornis (Daday 1906) имеет широкое распространение практически во всех тропических и субтропических странах Старого Света. Он был весьма подробно переописан Изабелой Ван-де-Велде при ревизии рода (Van de Velde, 1984), и его характеристика приведена вкратце на основе ее описания, ибо в моей коллекции он отсутствует.
Описание типового для подрода вида Mesocyclops (Pilosomesocyclops) aspericornis (Daday 1906) comb. n.
Самка (рис. 3). Длина тела без щетинок 1100–1600 мкм. По внешнему краю 4-го свободного торакального сомита с группой волосков. Длина генитального двойного сомита и его ширина соотносятся как 1.1–1.3 : 1. Семяприемник с широкими и длинными боковыми лопастями, слегка изогнутыми книзу, и широким нижним отделом. Семяпровод изогнут вправо. Каудальные ветви по внутренней поверхности покрыты густыми волосками, отношение длины к ширине 2.7–3.6, боковая щетинка прикреплена почти на трети длины ветвей. У мест вставки боковой и наружной щетинок имеются шипики. Концевые щетинки крепкие и густо оперены волосками, соотношение их длин начиная от наружной: 1/2.5–3.8/5.5–6.8/2.6–3.4. Спинная щетинка равна наружной, наибольшая из каудальных щетинок равна или в 1.3 раза длиннее уросомы. Антеннула 17-сегментная, достигает 2 торакального сомита, ее концевой членик несет широкую пилообразную гиалиновую мембрану с 1 очень глубоким вырезом. Антенны с 3-члениковым эндоподитом, базиподит на каудальной стороне, помимо основных групп шипиков, имеет широкое поле мелких шипиков в месте вставки медиальных щетинок, а также группу крошечных шипиков между латеральным и медиальным рядами шипов. Второй членик эндоподита антенн у популяций из Нового и Старого Света с разным числом щетинок (от 7 до 9), что, возможно, указывает на сборный характер этого таксона. Ротовые конечности типичного для циклопид строения и, наряду с антеннами, нередко бывают покрыты мелкими ямочками. Плавательные ноги 1–4 пар двуветвистые и трехчлениковые. Базиподит 1 пары без щетинки на внутреннем выросте, но покрыт густыми длинными волосками. Коксальная пластинка у ног 4 пары без волосков, по наружному краю с коническими заостренными выростами. Дистальный членик эндоподита 4-й пары длинный и узкий (L/W 2.5–3), на конце с двумя крепкими шипами, внутренний в 1.1–1.5 раз длиннее наружного и заметно короче членика. Рудиментная Р5 двучлениковая, базальный членик с длинной оперенной щетинкой, конечный членик на конце с оперенной щетинкой, которая в 1.1–1.6 раз длиннее крепкой медиальной шипообразной щетинки, которая в свою очередь равна щетинке базального членика.
Рис. 3.
Mesocyclops (Pilosomesocyclops) aspericornis (Daday 1906). a – генитальный сомит; b – каудальная ветвь; c – P1; d – P4; e – P4 Enp3; f – A1, последний сегмент; g – базиподит А2, каудально; h – базиподит А2, фронтально; i – Р5 и Р6, самец. Масштаб (мкм): a, b – 100; c–e, g–i – 65; f – 40 (a–h – по: Van de Velde, 1984; i – по: Holynska, 2000).

Самец. Длина тела без концевых щетинок 750–950 мкм. По внешнему краю 4-го свободного торакального сомита волосков немного. Сперматофоры крупные, бобовидные. Каудальные ветви короче, чем у самок (L/W 2.6–3.4), без волосков на внутренней поверхности, у мест вставки боковой и наружной щетинок имеются шипики. Дорсальная щетинка примерно равна наружной. Геникулирующие антеннулы 14-члениковые хватательного типа. Антенны и другие ротовые конечности, как у самок, нередко с ямками, но второй членик эндоподита антенны с 6 щетинками. Первая пара плавательных ног на внутреннем выросте базиподита без щетинки. Конструкция плавательных ног такая же, как у самки. Рудиментная 5-я пара: апикальная щетинка в полтора раза длиннее, чем шипообразная медиальная щетинка, щетинка базального членика немного меньше медиальной щетинки дистального членика. Рудиментная 6-я пара представлена широкой пластинкой с крепким внутренним шипом, почти равной ему по длине средней щетинкой и вдвое более длинной наружной щетинкой.
Распространение. Вид описан из Индонезии (о-в Суматра) и позднее был обнаружен повсеместно в тропиках, что позволило авторам сводок обозначить его ареал как пантропический (Reid, Sanders, 1986; Suárez-Morales et al., 2011). Учитывая значительные расхождения в морфологии, особенно для популяций из Старого и Нового Света, это заключение представляется мне маловероятным и основанным либо на существовании куста сестринских видов-близнецов, либо на процессах биоинвазий, либо того и другого вместе. Для получения решительного ответа на эти вопросы необходимо проведение широкомасштабного молекулярно-генетического и морфологического исследований.
Ключ для разделения на секции подрода Mesocyclops (Pilosomesocyclops) subgen. n.
1. Волоски имеются как на латеральной поверхности четвертого торакального сомита, так и на внутренней поверхности каудальных ветвей.....секция aspericornis
– Волоски имеются лишь на одной из этих структур.............................................................. (2)
2. Волосками покрыты внутренние поверхности каудальных ветвей..........секция pseudospinosus
– Волоски находятся на боковых лопастях четвертого сомита.................секция thermocyclopoides
Ключ для определения видов секции aspericornis
1. Рачки довольно крупные, длина самок 1.1–1.6 мм.................................................................. (2)
– Рачки мелкие, самки менее 1.1 мм длиной....(6)
2. Генитальный сегмент с RS типа T.............(3)
– Форма RS иная.................M. (P.) aspericornis (Daday 1906) (пантропический)
3. У А2 базиподиты на каудальной стороне с группой крупных шипиков дистально ближе к основанию 2 медиальных щетинок..........M. (P.) kayi Holynska et Brown 2003 (Бирма)
– У А2 базиподиты на каудальной стороне без таких шипиков...................................................(4)
4. На каудальной поверхности соединительной пластинки P4 2 ряда волосков.....M. (P.) pilosus Kiefer 1930 (Юго-Восточная Азия)
– Соединительные пластинки P4 не имеют волосков.................................................................(5)
5. На каудальных ветвях у места прикрепления наружной щетинки есть шипики; CR L/W меньше 3..... M. (P.) spinosus Van de Velde 1984 (Африка)
– На каудальных ветвях у места прикрепления наружной щетинки шипики отсутствуют; CR L/W больше 3.......M. (P.) mariae Guo 2000 (Китай)
6. У А2 базиподиты на каудальной стороне с группой крупных шипиков дистально у медиальных щетинок.............M. (P.) geminus Holynska 2000 (Калимантан)
– Базиподиты А2 на каудальной стороне с группой мелких шипиков у медиальных щетинок.......(7)
7. Соединительные пластинки P4 не имеют волосков....M. (P.) microlasius Kiefer 1981 (Филиппины)
– Соединительные пластинки P4 с волосками на каудальной поверхности................................(8)
8. Выросты на соединительной пластинке P4 короткие; рачки средних размеров (0.9–1.1 мм)........M. (P.) dadayi Holynska 1997 (Индия)
– Выросты соединительной пластинки P4 длинные; рачки очень мелкие (0.7–0.8 мм).....M. (P.) insulensis Dussart 1982 (Мадагаскар)
Ключ для определения видов секции pseudospinosus
1. Поровый канал RS сильно изогнут; CR с волосками на верхней части внутренней поверхности и шипиками на нижней части; выросты на соединительной пластинке P4 широкие......M. (P.) pseudospinosus Dussart et Fernando 1988 (Индонезия)
– Поровый канал RS слабо изогнут; CR с волосками на верхней части внутренней поверхности и без шипиков на нижней части; выросты на соединительной пластинке P4 узкие...M. (P.) shenzhenensis Guo 2000 (Ю. Китай и Цент. Вьетнам)
Ключ для определения подсекций и видов секции thermocyclopoides
1. В семяприемнике поперечные каналы встречаются у копуляторной поры под острым углом (V-type).......................(2) (подсекция woutersi)
– Каналы встречаются под тупым или почти развернутым углом...........................................(14)
2. Дорсальная поверхность последнего торакального сомита не покрыта густыми волосками помимо нескольких сенсилл............................. (3)
– Дорсальная поверхность последнего торакального сомита покрыта волосками................ (6)
3. На CR у места прикрепления боковой и наружной щетинок нет шипиков.......M. (P.) woutersi Van de Velde 1987 (Новая Гвинея, Австралия, Индокитай)
– На CR у места вставки боковой и/или наружной щетинок имеются шипики...................(4)
4. На дорсальной поверхности сегментов А1 нет ямок; 8 пор позади P6........M. (P.) dissimilis Defaye et Kawabata 1993 (Япония, В. Сибирь, Китай)
– На дорсальной поверхности сегментов 1, 4 у А1 есть ямки; 6 пор позади P6..........................(5)
5. Внутренний шип P4 Enp3 короче или равен наружному; выросты на соединительной пластинке P4 мелкие, тупые или острые....M. (P.) brooksi Pesce, De Laurentiis et Humphreys 1996 (Австралия)
– Внутренний шип P4 Enp3 равен или длиннее наружного; выросты на соединительной пластинке P4 длинные и острые........M. (P.) acanthoramus Holynska et Brown 2003 (Австралия)
6. На CR у места вставки боковой и наружной щетинок нет шипиков........................................(7)
– На CR у места вставки боковой и/или наружной щетинок имеются шипики...................(8)
7. CR L/W 2.9–4.5; А2 каудально с латеральным рядом из 13–19 шипов, фронтально 21–28 шипиков......................M. (P.) granulatus Dussart et Fernando 1988 (Пакистан, Индия, Мьянма)
– CR L/W 3.0–3.5; между последним торакальным сегментом и генитальным сомитом имеется “псевдосомит”; А2 каудально с латеральным рядом из 10–14 шипов, фронтально 22–26 шипиков..........M. (P.) arcanus Defaye 1995 (Израиль)
8. Генитальный сомит дорсально покрыт волосками...............................................................(9)
– Генитальный сомит дорсально не покрыт волосками.........................................................(11)
9. Генитальный сомит вентрально покрыт волосками.........M. (P.) pubiventris Holynska et Brown 2003 (частично) (Австралия)
– Генитальный сомит вентрально не покрыт волосками.........................................................(10)
10. На каудальной поверхности А2 у места прикрепления медиальных щетинок ряд крупных шипов; P4 Enp3 L/W 3.0–3.9..........M. (P.) notius Kiefer 1981 (Австралия)
– На каудальной поверхности А2 у места прикрепления медиальных щетинок поле мелких шипиков; P4 Enp3 L/W 2.4–2.7.......M. (P.) medialis Defaye 2001 (Новая Каледония)
11. На вентральной поверхности CR в нижней четверти у медиального края ряд длинных шипов...M. (P.) francisci Holynska 2000 (Малайзия)
– Таких шипов на CR нет............................(12)
12. A2 Enp2 с 9 щетинками; выросты на соединительной пластинке P4 тупые; волоски на дорсальной поверхности последнего торакального сегмента короткие и расположены близ дистальной границы...........M. (P.) roberti Holynska et Stoch 2012 (Фиджи)
– A2 enp2 с 7 щетинками; выросты на соединительной пластинке P4 острые; дорсальная поверхность последнего торакального сегмента покрыта длинными волосками.............................(13)
13. На CR шипики имеются у места прикрепления как боковой, так и наружной щетинок; А2 каудально с латеральным рядом из 11–12 шипов..... M. (P.) friendorum Holynska 2000 (Индонезия)
– На CR шипики имеются только у наружной щетинки; А2 каудально с латеральным рядом из 13–16 шипов........M. (P.) parentium Holynska 1997 (Индия)
14. Поровый канал семяприемника прямой или слегка изогнут....(15) (подсекция aequatorialis)
– Этот канал сильно изогнут......(22) (подсекция ogunnus)
15. На CR у места прикрепления боковой и/или наружной щетинок имеются шипики...(16)
– На CR ни у латеральной, ни у наружной щетинок шипиков нет.......................................... (21)
16. На каудальной поверхности соединительной пластинки P4 есть ряды волосков............. (17)
– На соединительной пластинке P4 волосков нет..................................................................... (18)
17. На каудальной поверхности базиподита А2 у места прикрепления медиальных щетинок имеется группа крупных шипиков, у Exp – несколько шипиков, латеральный ряд состоит примерно из 18–20 шипиков; CR L/W 2.3–2.5; на CR дорсальная щетинка короче наружной; выросты на соединительной пластинке P4 мелкие.....M. (P.) jakartensis Alekseev 2013 (Индонезия)
– На каудальной поверхности базиподита А2 орнаментация другая; CR L/W 2.7–3.4; на CR дорсальная щетинка длинее наружной; выросты на соединительной пластинке P4 длинные...M. (P.) major Sars 1927 (Африка)
18. Дорсальная поверхность последнего торакального сомита покрыта волосками...M. (P.) dussarti Van de Velde 1984 (З. Африка)
– Она не покрыта волосками.......................(19)
19. На каудальной поверхности базиподита А2 у места прикрепления медиальных щетинок нет поля мелких шипиков; у P5 апикальная щетинка в 1.6–2.0 раза длиннее медиального шипа....M. (P.) aequatorialis aequatorialis Kiefer 1929 (Африка, Мадагаскар)
– На каудальной поверхности базиподита А2 в этом месте есть поле мелких шипиков; у P5 апикальная щетинка в 1.1 – 1.5 раза длиннее медиального шипа......................................................... (20)
20. На CR внутренняя средняя щетинка (s2) немного короче или равна длине уросомы (0.9–1.0 : 1); позади P6 имеются 6 или 8 пор; поровый канал семяприемника прямой и широкий....M. (P.) aequatorialis similis Van de Velde 1984 (Африка, Узбекистан)
– На CR эта щетинка немного длиннее уросомы (1.1–1.5 : 1); позади P6 имеются 6 пор; поровый канал семяприемника слегка изогнутый и тонкий...........M. (P.) affinis Van de Velde 1987 (Новая Гвинея, Индокитай)
21. На каудальной поверхности соединительной пластинки P4 есть ряды волосков; выросты соединительной пластинки P4 крупные и острые; на каудальной поверхности базиподита А2 у места прикрепления медиальных щетинок шипиков нет; CR L/W 2.6–2.8; L/W генитального сегмента примерно 1.1................M. (P.) bosumtwii Mirabdullayev, Sanful et Frempong 2007 (З. Африка)
– На соединительной пластинке P4 волосков нет; выросты соединительной пластинки P4 мелкие и тупые; на каудальной поверхности базиподита А2 у медиальных щетинок есть ряд крупных шипиков; CR L/W 2.9–3.6; L/W генитального сегмента 1.3–1.5.........M. (P.) thermocyclopoides Harada 1931 (частично)
22. На CR нет шипиков ни у места вставки боковой, ни у наружной щетинки........................(23)
– На CR у места вставки боковой и/или наружной щетинок имеются шипики................. (24)
23. На каудальной поверхности базиподита А2 у места прикрепления медиальных щетинок ряд крупных шипиков………………M. (P.) thermocyclopoides Harada 1931 (частично)
– На базиподите А2 в этом месте поле мелких шипиков...........................M. (P.) isabellae Dussart et Fernando 1988 (Индия)
24. Дорсальная поверхность последнего торакального сомита покрыта волосками.............. (25)
– Дорсальная поверхность последнего торакального сомита не покрыта волосками..........(26)
25. На каудальной поверхности базиподита А2 у места прикрепления медиальных щетинок поле мелких шипиков.........M. (P.) pubiventris Holynska et Brown, 2003 (частично) (Австралия)
– На каудальной поверхности базиподита А2 в этом месте ряд крупных шипиков...M. (P.) ogunnus Onabamiro 1957 (Африка, Ю-В Азия)
26. На проктодеуме имеется проксимально группа коротких шипиков и дистально ряд длинных шипиков; дорсальная поверхность генитального сомита не покрыта волосками...M. (P.) augusti Papa et Holynska 2013 (Филиппины)
– На проктодеуме подобных шипиков нет; дорсальная поверхность генитального сомита покрыта волосками................M. (P.) australiensis (Sars 1908) (Австралия)
Подрод Mesocyclops (Tethymesocyclops) Alekseev subgen. n.
Zoobank LSID: urn:lsid:zoobank.org:act:A56349FD-F455-4B50-85DE-69DB67A51803
Диагноз. Группа видов со щетинкой на выросте базиподита Р1 и не имеющих волосков на теле в районе латеральной поверхности четвертого свободного торакального сомита и медиальной поверхности каудальных ветвей. Этот подрод объединяет 11 видов, известных из Индии (+о-в Цейлон), Африки (+о-в Мадагаскар), Австралии (+Новая Зеландия и Новая Гвинея), южной Мексики и подземных вод Вьетнама. Большинство этих находок приходится на континенты, входившие в состав южного суперконтинента Гондваны, а точнее – на ту их часть, что составляла побережье моря Тетис периода отделения Гондваны от Лавразии. Возможно, что современное распределение по миру видов этого подрода было связано с последующим делением зтого суперконтинента на Африку, Австралию, Индию и Южную Америку. В водоемах последнего континента виды этого подрода пока не найдены, что предоставляет автору возможность предсказать их обнаружение там в обозримом будущем.
Описание типового для подрода вида Mesocyclops (Tethymesocyclops) annae Kiefer 1930 comb. n.
Самка (рис. 4). Длина тела без щетинок 1200–1400 мкм, По внешнему краю 4-го свободного торакального сомита волосков нет. Длина генитального двойного сомита и его ширина соотносятся как 1.1–1.2. Семяприемник с широкими и длинными боковыми лопастями, изогнутыми книзу, и широким нижним отделом. Семяпроводящий (поровый) канал прямой и длинный. Каудальные ветви без волосков, отношение длины к ширине 2.2–2.5. Боковая щетинка прикреплена почти на трети длины ветвей и очень длинная. Шипиков на каудальных ветвях в местах вставки боковой и концевой наружной щетинок нет. Концевые щетинки крепкие и густо оперены волосками, соотношение их длин начиная от наружной: 1/3.5–3.8/5.0/2.4–2.6. Спинная щетинка короче наружной, наибольшая из каудальных щетинок равна или слегка длиннее уросомы. Антеннула 17-сегментная, достигает 2 торакального сомита, ее концевой членик несет широкую пилообразную гиалиновую мембрану с 2–3 глубокими вырезами. Антенны с 3-члениковым эндоподитом, базиподит на каудальной стороне несет ряд длинных шипиков у латерального края и мелких – у передней части медиального края, косой медиальный ряд отсутствует; на фронтальной стороне имеется ряд длинных шипиков у передней части латерального края и у дистальной части латерального края (близ экзоподита). Ротовые конечности типичного для циклопид строения. Плавательные ноги 1–4 пар двуветвистые и трехчлениковые. Базиподит 1 пары несет щетинку на внутреннем выросте, на четверть более длинную, чем первый сегмент эндоподита. Коксальная пластинка у ног 4 пары без волосков, по наружному краю с длинными коническими заостренными выростами. Дистальный членик эндоподита 4-й пары длинный и узкий (примерно в 3 раза длиннее ширины), на конце с двумя крепкими шипами, внутренний почти равен наружному и слегка короче членика. Рудиментная Р5 двучлениковая, базальный членик с длинной боковой щетинкой, превышающей длину медиальной щетинки дистального членика, конечный членик на конце с оперенной апикальной щетинкой, которая длиннее медиальной.
Рис. 4.
Mesocyclops (Tethymesocyclops) annae Kiefer 1930: a – генитальный сомит и Р5; b – каудальные ветви, вентрально; c – P1; d – кокса P4, каудально; e – P4 Enp3; f – базиподит A2, каудально. Масштаб (мкм): a, b – 100; c–f – 65 (по: Holynska, 2003).
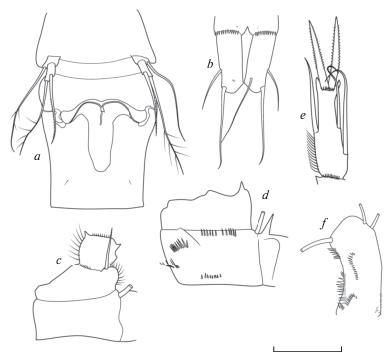
Самец неизвестен.
Распространение. Вид описан из водоема на о-ве Мадагаскар.
Ключ для определения видов подрода Mesocyclops (Tethymesocyclops) subgen. n.
1. Внутренняя щетинка CR менее чем вдвое длиннее наружной..............................................(2)
– Эта щетинка по меньшей мере в два раза длиннее наружной..............................................(3)
2. У P5 латеральная щетинка значительно длиннее медиальной щетинки.....M. (T.) yutsil Reid in Fiers et al. 1996 (Мексика)
– Соотношение длин этих щетинок обратное.................. M. (T.) chaci Fiers 1996 (Мексика)
3. На CR у основания боковой и наружной щетинок есть шипики.........M. (T.) sondoongensis Tran et Holynska 2015 (Вьетнам)
– Таких шипиков нет.................................... (4)
4. На каудальной стороне базиподита А2 латеральный ряд шипиков доходит до места прикрепления Exp........................................................... (5)
– Этот ряд шипиков не достигает Exp......... (7)
5. Поровый канал RS длинный....M. (T.) darwini Dussart et Fernando 1988 (Австралия)
– Поровый канал короткий...........................(6)
6. На гиалиновой пластинке А2 больше одной вырезки; P4 Enp3 L/W 2.5–2.9; размеры рачка 1.3–1.5 мм...............M. (T.) paludosus Lindberg 1956 (Африка)
– На гиалиновой пластинке одна мелкая вырезка; P4 Enp3 L/W около 3.2; размеры рачка 1.1–1.3 мм...............M. (T.) holynskae Karanovic 2006 (Австралия)
7. Боковые рукова RS широкие, изогнутые назад..............................................................(8)
– Рукова RS узкие, почти прямые.............. (10)
8. Внутренний выступ базиподита P4 без волосков.........M. (T.) annae Kiefer 1930 (Мадагаскар, Индия)
– Внутренний выступ базиподита P4 с дистальными волосками....................................... (9)
9. Поперечные каналы RS соединяются под почти тупым углом; на первом членике А1 нет вентральных шипиков..........M. (T.) pseudoannae Van de Velde 1987 (Новая Гвинея)
– Поперечные каналы соединяются под острым углом; на первом членике А1 есть вентральные шипики..........M. (T.) splendidus Lindberg 1943 (Индия)
10. У P5 латеральная щетинка почти равна медиальной; на CR центральная внутренняя щетинка (s2) в 4.8–5.1 раз длиннее наружной (s4), а внутренняя (s1) длиннее наружной (s4) в 2.6–2.8 раза...........M. (T.) dayakorum Holynska 2000 (Индонезия)
– у P5 латеральная щетинка короче медиальной; на CR центральная внутренняя щетинка (s2) в 4.2–4.7 раз длиннее наружной (s4), а внутренняя (s1) длиннее s4 в 2.2–2.4 раза......M. (T.) rarus Kiefer 1981 (Африка)
Подрод Мesocyclops (Neomesocyclops) Alekseev subgen.n.
Zoobank LSID: urn:lsid:zoobank.org:act:0361FD6F-1742-40E2-A213-55A19030415C
Диагноз. Члены этой группы как морфологически – щетинка на выросте базиподита Р1 и волоски хотя бы на одном из элементов просомы (Th4) или уросомы (CR), – так и территориально (Новый Свет) хорошо отделены от остальных видов рода. Единственный вид с такими признаками, описанный из Южной Африки, – М. tenuisaccus – морфологически чрезвычайно близок к массовой форме из Южной Америки – M. аnnulatus – и, на мой взгляд, является биоинвазией последнего, занесенной мореплавателями с питьевой водой в период активной работорговли (XVI–XVIII вв). К выводу об идентичности этих видов пришла и Холинска (Holynska, 2006). По сочетанию признаков опушения волосками Th4 сомита просомы и каудальных ветвей виды подрода по аналогии с подродом Pilosomesocyclops subgen. n. можно подрод Neomesocyclops subgen. n. разделить на 3 неравные по числу видов секции.
1. С волосками на обеих структурах – секция annulatus, включающая 10–12 видов, из которых 11 встречаются в Южной и Центральной Америке, 1–2 вида описаны из Мексики. Один упомянутый выше вид (M. tenuisaccus) описан из реки в Кейптауне, Южная Африка. Предположительно инвазийный характер его появления требует подтверждения молекулярно-генетическими методами.
2. Волоски имеются только на латеральной поверхности 4-го торакального сомита – секция reidae включает пока только один вид из Южной Америки.
3. Волоски имеются только на внутреннем крае каудальных ветвей – секция edax включает один вид. M. edax описан дважды – из Великих озер Северной Америки под этим названием и позднее как M. nicaraguensis из Южной Америки, откуда он вероятно и проник в Северную Америку после соединения континентов.
Центром возникновения и многообразия видов этого подрода, несомненно, следует считать Южную Америку после отделения ее от Африканской платформы около 80 миллионов лет назад. Учитывая низкую численность в двух секциях, определительный ключ составлен для подрода в целом, что не помешает вернуться к делению на секции в случае вполне возможного пополнения их видами.
Описание типового для подрода вида Mesocyclops (Neomesocyclops) edax (Forbes 1891) comb. n.
Полноценное описание голотипа M. (N.) edax, несмотря на его широкое распространение в Новом Свете, отсутствует, что определило более подробное его рассмотрение при характеристике этого вида как типового для подрода. При описании его был использован топотип самки из зоопланктонной пробы, собранной автором в районе одного из Великих озер (оз. Эри) на западе США, штат Висконсин, район г. Чикаго. Место хранения описанного топотипа № 56 055 – Федеральная коллекция Зоологического института РАН № 96-03-16: раздел “типовая коллекция видов пресноводных беспозвоночных (отделение систематики)”. При описании изменчивости самца использована статья Dahms & Fernando (1995).
Самка (рис. 5). Длина тела без каудальных щетинок 1350 (900–1450) мкм, неокрашена, соотношение длины и максимальной ширины около 3 (2.98). Просома и уросома примерно соотносятся как 1.6 : 1, отношение длины цефалосомы к ширине 1.1. По внешнему краю 4-го свободного торакального сомита нет волосков (по Холинской (2003) изредка встречаются особи с короткими малочисленными волосками, возможно атавизм). Длина генитального двойного сомита и его ширина соотносятся как 1.4 : 1. Семяприемник с широкими лопастями, нижняя часть расширена и занимает около половины площади прилегающей части сомита. Семяпровод (поровый канал) короткий продолговатой формы при просматривании сверху. Каудальные ветви узкие, отношение длины к ширине около 4, боковая щетинка прикреплена на расстоянии 2/5 длины ветвей от концевых щетинок. По внутренней поверхности ветвей расположены длинные густые волоски. Концевые щетинки крепкие и густо оперены полосками, соотношение их длин начиная от наружной: 1.0/2.8–3.1/3.6–4.2/2.4–2.8. Спинная щетинка заметно длиннее наружной (в 1.4 раза), наибольшая из каудальных щетинок короче уросомы (0.85).
Рис. 5.
Mesocyclops (Neomesocyclops) edax (Forbes 1891): a – общий вид; b – генитальный сомит и Р5; c – анальный сомит и каудальная ветвь, дорсально; d – P1; e – Р4; f – P4 Enp3; g – А1, последний сегмент; h – базиподит А2, фронтально; i – базиподит А2, каудально. Масштаб (мкм): a – 100; b – 120; c–f – 100; g–i – 65 (a, c – по: Dahms, Fernando, 1995; b, g – по: Reid, Moreno, 1999; d, e, f, h, i – ориг.).
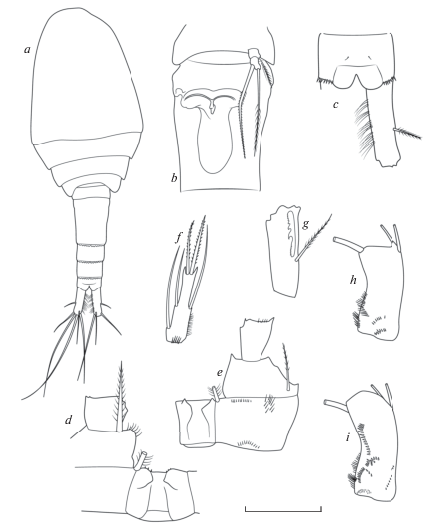
Антеннула 17-члениковая, достигает 2 торакального сомита, ее концевой членик несет широкую гиалиновую мембрану с 2–3 зазубринами, а первый членик наряду с 7 щетинками орнаментирован длинным рядом из крепких волосков. Антенны с 3-члениковым эндоподитом, базиподит по обеим сторонам несет лишь основные латеральные группы зубчиков. Второй членик эндоподита антенн несет 9 щетинок. Ротовые конечности типичного для циклопид строения, они редко используются при видовой идентификации (в этом роде никогда) и поэтому характеризуются очень кратко. Верхняя губа с 11 зубцами разной длины и редкими волосками. Мандибула изогнута и по наружному краю с 6–7 зубцами и длинной чувствительной щетинкой, покрытой редкими крепкими волосками, трехчленистый эндит с 2 длинными дистально оперенными щетинками и одной короткой голой. Максиллулы сложного строения, дистально с 4 парами зубцов и сенсилл, гнатобаза гладкая с 7 разной длины и толщины щетинками. Максиллы четырехчлениковые мощные, щетинки на сегментах располагаются попарно. Максилипеды после антеннул самые длинные, они также четырехчлениковые, несут по 2–3 щетинки, дистально покрытые крепкими волосками, первый и второй сегмент по внутренней стороне с рядом длинных крепких волосков.
Плавательные ноги 1–4 пар двуветвистые и трехчлениковые. Коксальные сегменты соединены пластинкой, которая у 4 пары по наружному краю с короткими тупыми выростами; коксальная щетинка густо оперена волосками и очень длинная. Базиподит первой пары на внутреннем выросте с длинной хорошо различимой щетинкой – важнейший признак подрода. Дистальный членик эндоподита 4-й пары длинный и узкий на конце с двумя крепкими шипами, внутренний слегка длиннее наружного, но заметно короче членика.
Рудиментная пятая пара двучлениковая, характерного для рода строения, базальный членик с длинной оперенной щетинкой, конечный членик удлиненный, на конце с тонкой оперенной щетинкой, которая несколько короче крепкой медиальной шипообразной щетики.
Самец (рис. 6). Длина тела без концевых щетинок 884 мкм с вариацией от 600 до 900 в разных популяциях. Сперматофоры крупные, окрашены в красноватый цвет. Просома и уросома примерно соотносятся как 1.8 : 1, отношение длины цефалосомы к ширине около 1.25 : 1. По внешнему краю 4-го свободного торакального сомита волосков нет. Каудальные ветви короткие, отношение длины к ширине 2.7 с вариацией 2.2–2.8, боковая щетинка прикреплена почти на середине наружного края каудальных ветвей. По внутренней поверхности ветвей, начиная от верхней трети и сгущаясь в дистальном направлении прикреплены длинные волоски. Концевые щетинки довольно длинные и густо оперены волосками. Дорсальная щетинка заметно длиннее наружной, но короче внутренней. Наибольшая из каудальных щетинок почти равна уросоме. Геникулирующие антеннулы 14-члениковые хватательного типа, приспособленные для удерживания самки. Антенны и другие ротовые конечности по строению и вооружению близки к таковым у самок. Первая пара плавательных ног на внутреннем выросте базиподита с длинной щетинкой, достигающей дистального края второго сегмента эндоподита. Конструкция других плавательных ног такая же, как у самки, но эндоподит 4 пары несколько короче, соотношение длины и ширины равно 3.1–3.4, на конце с почти равными шипами, оба короче членика. Рудиментная 5-я пара: апикальная щетинка длиннее, чем шипообразная медиальная щетинка дистального членика, латеральная щетинка базального членика заметно короче обоих придатков. Рудиментная 6-я пара представлена широкой пластинкой с тремя придатками, из которых внутренний шип почти равен по длине средней щетинке и вдвое короче наружной щетинки.
Рис. 6.
Mesocyclops (Neomesocyclops) edax (Forbes 1891), самец: a – общий вид, b – геникулирующая антеннула, c – P5 и Р6. Масштаб (мкм): a – 200; b – 120; c – 70 (по: Dahms, Fernando, 1995).
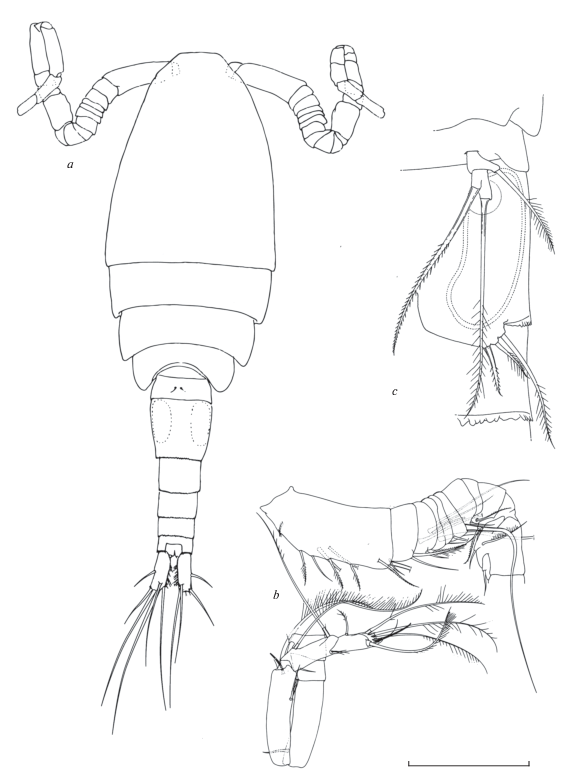
Распространение. Вид показан для озероподобных водоемов Северной Америки и как M. nicaraguens, признанный его синонимом, для Центральной и Южной Америки (Reid, Ueda, 2003).
Описание нового вида Mesocyclops (Neomesocyclops) frankfiersi Alekseev subgen. n., Alekseev sp. n.
Zoobank LSID: urn:lsid:zoobank.org:act:452772F5-ECBF-405F-90AB-C67972E67907
Типовое местообитание. Место нахождения типовой популяции – о-в Косумель, Атлантическое побережье Мексики; типовой водоем – дождевые лужи среди пальмовых зарослей на южной оконечности острова. Материал собран автором в июне 2011 г.
Типовая серия. Голотип № 56123, препарат самки, размещенный на одном стекле. Паратип № 56 124, препарат самца на одном стекле, паратипы № 56 125, 12 самок и 8 самцов в сосуде с чистым глицерином, хранящиеся в Федеральной коллекции Зоологического института РАН № 96-03-16 в разделе “Типовая коллекция водных беспозвоночных отделения систематики”.
Самка (Голотип № 56 123) (рис. 7, 8). Длина тела без каудальных щетинок 980 мкм. Тело окрашено в бурый цвет, который исчезает при спиртовом хранении и очевидно обусловлен цветом опада пальмовых листьев. Просома и уросома примерно соотносятся как 2 : 1, отношение длины цефалосомы к ширине 1.1 : 1. По внешнему краю 4-го свободного торакального сомита имеются волоски.
Рис. 7.
Mesocyclops (Neomesocyclops) frankfiersi sp. n., самка: a – общий вид; b – генитальный сомит и Р5; c – каудальные ветви, дорсально; d – А1, последние сегменты; e – базиподит А2, фронтально; f – базиподит A2, каудально; g – максилла; h – мандибула; i – максиллула. Масштаб (мкм): a – 200; b, c – 100; d–i – 40.
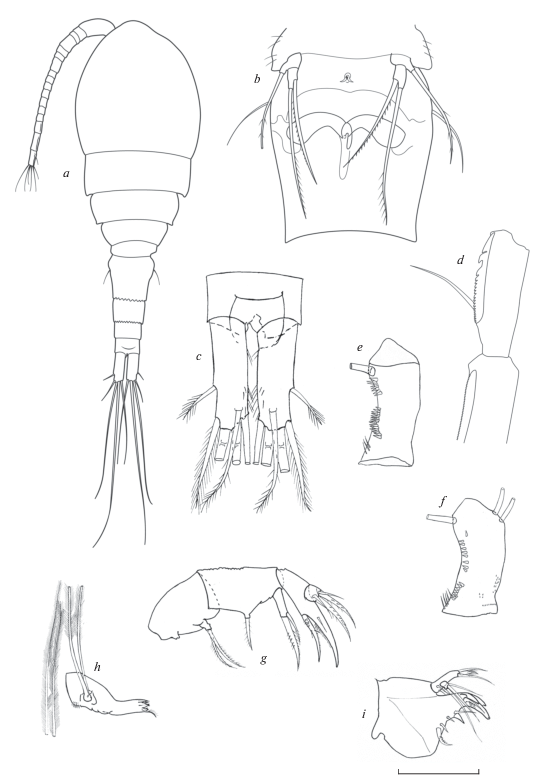
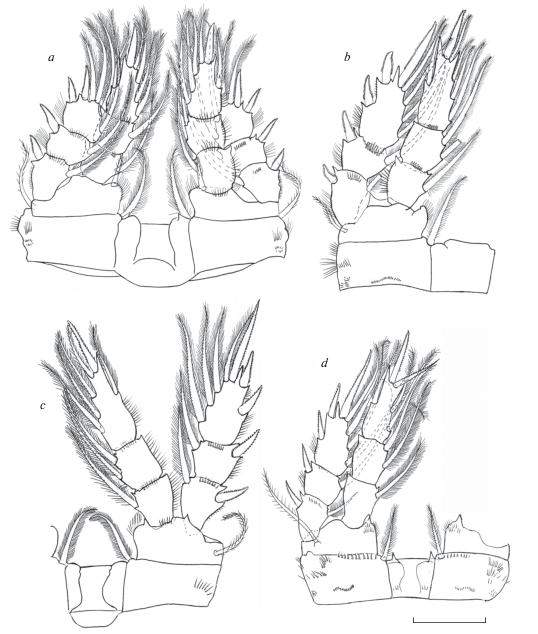
Рис. 8.
Mesocyclops (Neomesocyclops) frankfiersi sp. n., самка (продолжение): a – P1, b – P2, c – P3, d – P4. Масштаб 40 мкм.
Длина генитального двойного сомита и его ширина соотносятся как 1.1 : 1. Семяприемник с довольно широкими боковыми лопастями, повернутыми книзу, и очень узкой средней дистальной частью . Семяпровод изогнут влево и довольно длинный. Каудальные ветви с отношением длины к ширине около 3.2 (3.1–3.3), боковая щетинка прикреплена на расстоянии примерно 1/3 от терминальных щетинок. По внутренней поверхности ветвей расположены длинные, но довольно редкие волоски. Концевые щетинки крепкие и густо оперены волосками, соотношение их длин начиная от наружной: 1/4.7–5.1/6.5–7.4/3.3–3.7. Спинная щетинка в 1.15–1.25 раза длиннее наружной, наибольшая из каудальных щетинок в 1.35 раза длиннее уросомы.
Антеннула 17-члениковая, в прижатом виде достигает середины второго торакального сомита, ее концевой членик несет широкую гиалиновую мембрану, доходящую до основания средней щетинки, с одной крупной и двумя узкими зазубринами. Антенны с 3-члениковым эндоподитом. Орнаментация на каудальной поверхности базиподита: проксимальный ряд длинных шипиков на латеральном крае, дистальнее него ряд из 7–9 более коротких шипиков и ряд из 8–9 шипиков, не доходящий до основания Exp; с медиальной стороны проксимально проходит поперечный ряд крошечных шипиков и две группы мелких шипиков расположены дистальнее его. Орнаментация на фронтальной стороне базиподита состоит из проксимальной группы длинных шипиков, длинного латерального ряда из 14–16 шипиков и дистального ряда шипиков у основания Exp. Второй сегмент эндоподита антенн несет 9 щетинок. Ротовые конечности типичного для циклопид строения. Верхняя губа с 8–10 зубцами равной длины и редкими волосками (не показано). Мандибула изогнута и по наружному краю с грубыми зубцами и чувствительной щетинкой, покрытой короткими волосками, трехчленистый эндит с 2 длинными почти равной длины дистально оперенными щетинками и одной очень короткой голой. Максиллулы сложного строения, дистально с 4 парами зубцов и сенсилл, гнатобаза (щупик) гладкая с 7 разной длины и толщины щетинками. Максиллы четырехчлениковые мощные, щетинки на сегментах располагаются попарно, обычно сочетая крепкую шипообразную и тонкую чувствительную. Максилипеды (не показаны ) после антеннул самые длинные, они также четырехчлениковые, несут по 2–3 щетинки, дистально покрытые крепкими волосками, и в настоящее время не имеют заметного таксономического значения.
Плавательные ноги 1–4 пар двуветвистые и трехчлениковые. Коксальные сегменты соединены пластинкой, которая у 4 пары каудально несет волоски и по наружному краю с довольно длинными заостренными и широкими в основании выростами; коксальная щетинка густо оперена волосками и достаточно длинная. Базиподит первой пары на внутреннем выросте с короткой щетинкой, едва достигающей дистального края первого сегмента эндоподита. Дистальный членик эндоподита 4-й пары длинный и узкий (отношение длины к ширине 2.6) на конце с двумя крепкими почти равными шипами, внутренний более мощный, чем наружный (в 1.1 раза), оба шипа короче членика. Концевые щетинки этого сегмента заходят за середину ближайших к ним шипов.
Рудиментная пятая пара двучлениковая, характерного для рода строения, базальный членик короткий уплощенный с длинной оперенной щетинкой, дистальный членик удлиненный палочковидный, с апикальной щетинкой заметно длиннее боковой шипообразной щетинки, которая в свою очередь практически равна щетинке базального членика. Яйцевые мешки содержат небольшое количество крупных яиц, 20–30.
Самец. (Паратип № 56 124) (рис. 9) Длина тела без концевых щетинок 650 мкм. Цвет буроватый. Сперматофоры крупные, широкоовальной формы, красноватового или оранжевого цвета. Просома и уросома примерно соотносятся как 2 : 1, отношение длины цефалосомы к ширине около 1. По внешнему краю 4-го свободного торакального сомита волосков нет.
Рис. 9.
Mesocyclops (Neomesocyclops) frankfiersi sp. n., самец: a – общий вид; b – А1; c – P5, P6; d – Р1; e – Р4. Масштаб (мкм): a – 200; b – 70; c–e – 50.
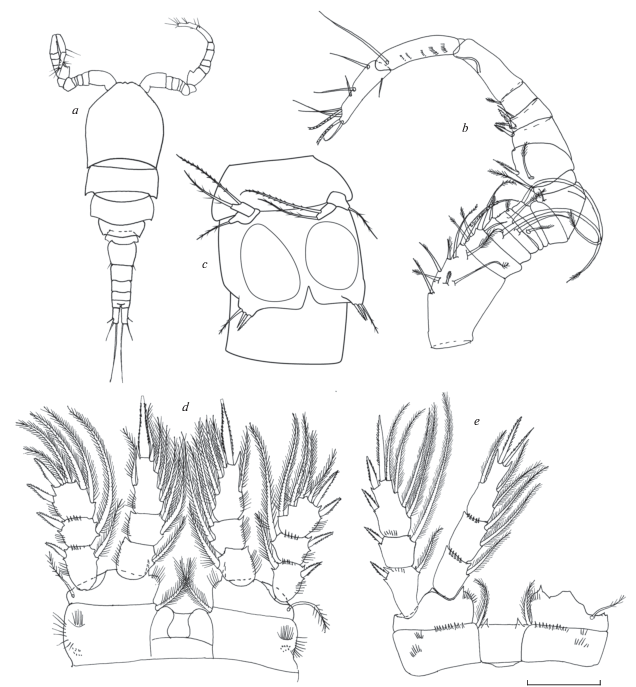
Длина генитального сомита примерно равна ширине. Каудальные ветви короткие, отношение длины к ширине 2.7 (с вариацией 2.2–2.8), боковая щетинка прикреплена почти на уровне дистальной трети наружного края ветвей. Дорсальная щетинка заметно длиннее наружной, но короче внутренней. Концевые щетинки довольно длинные и густо оперены волосками, внутренняя в 2.5 раза длиннее наружной. Каудальные ветви без волосков по внутреннему краю. Геникулирующие антеннулы 14-члениковые хватательного типа. Строение антенн и других ротовых конечностей в общем виде повторяет строение таковых у самки. Плавательные ноги в целом как у самки. Первая пара на внутреннем выросте базиподита с коротким шипом, который вдвое короче, чем у самки и доходит лишь до середины 1 сегмента эндоподита. 4-я пара сходного с самкой строения, на коксальной пластинке заостренные с широким основанием выросты. Формула зубцов и волосков на каудальной стороне коксоподита сходна с таковой у самки. Коксальная пластинка всех плавательных ног без волосков. 5 пара плавательных ног отличается от самки большей длиной шипообразной медиальной щетинки дистального членика. Рудиментарная 6-я пара в виде широкого клапана с тремя придатками, из которых внутренний шип и средняя щетинка почти равны, а наружная щетинка в 1.7 раза их длиннее.
Ареал. Новый вид пока известен только из типового местообитания. Он принадлежит к довольно богатой видами секции annulatus подрода Neomesocyclops, все представители которой обитают в водоемах Южной и Северной Америк.
Комментарий. Вид отличается от прочих членов секции сильно редуцированной щетинкой базального выроста 1-й пары плавательных ног и рядом других деталей (формой семяпровода самки, соотношением длин придатков пятой пары, строением мембранной пластинки дистального членика антеннул).
Этимология. Название вида посвящено памяти бельгийского систематика веслоногих раков Франка Фирса, безвременно ушедшего из жизни и науки.
Ключ для определения видов подрода Mesocyclops (Neomesocyclops) subgen. n.
1. На латеральной поверхности Th4 есть волоски..............................................................(2)
– Латеральная поверхность Th4 не покрыта волосками................M. (N.) edax (Forbes 1891) (Северная и Центральная Америка)
2. На медиальной поверхности каудальных ветвей есть волоски............................................ (3)
– Волосков на медиальной поверхности CR нет...............................M. (N.) reidae Petkovski 1986 (Южная и Центральная Америка)
3. Соединительная пластинка ног четвертой пары не имеет выростов.....................................(4)
– Эта пластинка снабжена парой более или менее длинных выростов................................. (10)
4. Семяприемник с длинными широкими боковыми лопастями, второй эндоподит антенн с девятью щетинками.....M. (N.) meridianalis Dussart et Frutos 1986 (Южная Америка)
– Семяприемник с узкими короткими боковыми лопастями, второй эндоподит антенн несет 7 или 8 щетинок................................................. (5)
5. Каудальные ветви на брюшной стороне несут дистально группу крепких шипиков.....M. (N.) pseudomeridianus Defaye et Dussart 1988 (Французская Гвиана)
– Такой группы шипиков на каудальных ветвях нет................................................................ (6)
6. Вентральная поверхность генитального сомита покрыта волосками................................... (7)
– Вентральная поверхность генитального сомита имеет лишь ямки.....M. (N.) evadomingoi Gutierrez-Aguirre et Suárez-Morales 2001 (Мексика)
7. Боковые лопасти семяприемника в верхней части имеют холмообразные выросты..M. (N.) venezolanus Dussart 1987 (Южная Америка)
– Боковые лопасти семяприемника без таких выростов............................................................ (8)
8. На CR имеются шипики в месте прикрепления наружной щетинки............M. (N.) brasilianus Kiefer 1933 (Центральная и Южная Америка)
– В месте прикрепления наружной щетинки шипиков нет...................................................... (9)
9. Каудальные ветви в 3.3–3.7 раза длиннее ширины..................M. (N.) meridianus (Kiefer 1926) (Центральная и Южная Америка)
– Каудальные ветви примерно в 2.9 раза длиннее ширины................M. (N.) varius Dussart 1987 (Гватемала)
10. Каудальные ветви весьма длинные, соотношение длины и ширины более пяти....M. (N.) annulatus (Wierzejski 1892) (Южная Америка)
– Каудальные ветви заметно короче, соотношение длины и ширины обычно около трех и редко достигает четырех........................................ (11)
11. Семяприемник с широкими боковыми лопастями............................................................. (12)
– Семяприемник с узкими боковыми лопастями............................................................ (13)
12. Соединительная пластинка P4 имеет волоски на каудальной поверхности; апикальная щетинка P5 не заходит за нижнюю границу генитального сомита; размер рачка около 1 мм......M. (N.) frankfiersi sp. n.
– На соединительной пластинке P4 волосков нет; апикальная щетинка очень длинная и значительно заходит за нижний край генитального сомита; размер рачка от 1.2 до 2 мм.....M. (N.) longisetus (Thiébaud 1912) (С. и Ю. Америки)
13. Генитальный сомит несет поперечные рубчики.........M. (N.) paranaensis Dussart et Frutos 1986 (Южная Америка)
– Поперечных рубчиков на генитальном сомите нет............................................................ (14)
14. Размеры рачка 0.5–0.6 мм; медиальный вырост базиподита P4 только с дистально расположенными волосками.............M. (N.) intermedius Pesce 1985 (Вест-Индия)
– Размер рачка около 1мм; медиальный вырост базиподита P4 с волосками дистально и проксимально.............M. (N.) ellipticus Kiefer 1936 (Центр. и Ю. Америка)
ЗАКЛЮЧЕНИЕ
Таким образом, насыщенный видами род Mesocyclops (78 видов) был подвергнут ревизии и разделен на четыре подрода исходя из особенностей морфологии и ареалов, занимаемых их представителями. Из водоемов о-ва Косумель (Мексика) описан новый для науки вид Mesocyclops (Neomesocyclops) frankfiersi sp. n. По сравнению с последней фаунистической сводкой (Ueda, Reid, 2003) приведенные ключи расширены на 12 таксонов (до 78 видов). Анализ рода позволяет предположить, что его наполненность видами далека от завершения. Наибольшего пополнения новыми видами следует ожидать из водоемов Юго-Восточной Азии и Южной Америки.
Список литературы
Alekseev V.R., 1993. The new subspecies of Mesocyclops leuckrti from East Siberia (Crustacea, Cyclopida: Cyclopidae) // Zoosystematica Rossica. V. 2. P. 55–58.
Alekseev V.R., 1998. Key to freshwater Cyclopidae of Russia and adjacent lands (Crustacea) // Zoosystematica Rossica. V. 7. P. 25–43.
Alekseev V.R., 2002. Copepoda. Chapter 4. A guide to tropical freshwater zooplankton. Fernando C.H. (ed.). Leiden: Backhuys Publishers. P. 123–187.
Alekseev V.R., Abramson N.I., Sukhikh N.M., 2009. Introd uction of sibling species to the ecosystem of the Baltic Sea // Doklady Biological Sciences. V. 429. № 1. P. 544–547.
Alekseev V. R., Miracle M. R., Sahuquillo M., Vicente E., 2020. Redescription of Acanthocyclops vernalis (Fischer, 1853) and Acanthocyclops robustus (Sars, 1863) from neotypes, with special reference to their distinction from Acanthocyclops americanus (Marsh, 1892) and its invasion of Eurasia // Limnetica. V. 40. P. 000–000. DOI: (in press)https://doi.org/10.23818/limn.40.xx
Besse J., 1995. Continental Drift. Tectonics Observatory. California Institute of Technology. [Электронный ресурс]. Режим доступа: http://www.tectonics.caltech.edu/ outreach/animations/drift.html. Дата обновления: 25.09.2009.
Connolly J.K., Watkins J.M., Hinchey E.K., Rudstam L.G., Reid J.W., 2019. The Asian cyclopoid copepod Mesocyclops pehpeiensis Hu 1943 reported from the western basin of Lake Erie // Journal of Great Lakes Research. V. 45. P. 196–201.
Dahms H.U., Fernando C.H., 1993. Redescription of Mesocyclops leuckarti (Copepoda, Cyclopoida), including a study of its naupliar development // Internationale Revue der Gesamten Hydrobiologie und Hydrographie. V. 78. P. 589–609.
Dahms H.U., Fernando C.H., 1995. Redescription of female and male Mesocyclops edax Forbes, 1891 (Copepoda: Cyclopoida) // Journal of Crustacean Biology. V. 15. P. 317–328.
Dussart B.H., Defaye D., 2006. World directory of Crustacea Copepoda of inland waters. II – Cyclopiformes. Leiden: Backhuys Publishers. 354 p.
Fefilova E.B., Baturina M.A., Kononova O.N., Loskutova O.A., Khokhlova L.G., Dubovskaya O.P., 2014. Long-term changes of aquatic communities in the Kharbeyskie lakes // Journal of Siberian Federal University. Biology. V. 7. № 3. P. 240–266.
Gophen M., 1984. The impact of zooplankton status on the management of Lake Kinneret (Israel) // Hydrobiologia. V. 113. P. 249–258.
Hołyńska M., 2000. Revision of the Australasian species of the genus Mesocyclops Sars, 1914 (Copepoda, Cyclopidae) // Annales Zoologici, Warszawa. V. 50. P. 363–447.
Hołyńska M., 2003. Genus Mesocyclops Sars, 1914 // Copepoda: Cyclopoida. Genera Mesocyclops and Thermocyclops. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World. Reid J.W., Ueda H. (Eds). V. 20. P. 12–213.
Hołyńska M., 2006. Phylogeny of Mesocyclops (Copepoda: Cyclopidae) inferred from morphological characters // Zoological Journal of the Linnean Society. V. 147. P. 1–70.
Kiefer E., 1981. Beitrag zur Kenntnis von Morphologie, Taxonomie und geographischer Verbreitung von Mesocyclops leuckarti auctorum // Archive fuer Hydrobioogie. Supplement. V. 62. P. 148–190.
Meleg L.N., Fiers F., Robu M., Moldovan O.T., 2012. Distribution patterns of subsurface copepods and the impact of environmental parameters // Limnologica. V. 42. P. 156–164.
Montoliu L., Miracle M.R., Elías-Gutiérrez M., 2015. Using DNA barcodes to detect non-indigenous species: the case of the Asian copepod Mesocyclops pehpeiensis Hu, 1943 (Cyclopidae) in two regions of the world // Crustaceana. V. 88. P. 1323–1338.
Peixoto R.S., Brandão L.P.M., Valadares C.F., Barbosa P.M.M., 2010. Occurrence of Kellicottia bostoniensis (Rousselet, 1908) and Mesocyclops ogunnus Onabamiro, 1957 in lakes of the Middle River Doce, MG, Brazil // Acta Limnologica Brasiliensia. V. 22. P. 356–360.
Reid J.W., Moreno D., 1999. The western and southern distribution of Mesocyclops edax (S. A. Forbes) (Crustacea: Copepoda: Cyclopoida) // Proceedings of the Biological Society of Washington V. 112. P. 581–591.
Reid J.W., Saunders, J.F., 1986. The distribution of Mesocyclops aspericornis (von Daday) in South America // Journal of Crustacean Biology V. 6. № 4. P. 820–824.
Reid J.W., Ueda H. (Eds), 2003. Copepoda: Cyclopoida: Genera Mesocyclops and Thermocyclops // Guides to the identification of the microinvertebrates of the continental waters of the world. V. 20. P. 1–318.
Suárez-Morales E., Gutiérrez-Aguirre M.A., Mendoza F. 2011. The Afro-Asian cyclopoid Mesocyclops aspericornis (Crustacea: Copepoda) in eastern Mexico with comments on the distribution of exotic copepods // Revista Mexicana de Biodiversidad. V. 82. № 1. P. 109–115.
Van de Velde I., 1984. Revision of the African species of the genus Mesocyclops Sars, 1914 (Copepoda: Cyclopidae) // Hydrobiologia. V. 109. P. 3–66.
Wyngaard G., Hołynska M., Schulte J.A., 2010. Phylogeny of the freshwater copepod Mesocyclops (Crustacea: Cyclopidae) based on combined molecular and morphological data, with notes on biogeography // Molecular Phylogenetics and Evolution. V. 55. P. 753–764.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал