

Зоологический журнал, 2020, T. 99, № 5, стр. 516-527
Сравнительный анализ рецентных субаквальных тафоценозов на примере 24 малых водоемов Шатурского и Рузского районов Московской области (Центральная Россия)
a Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: antzhar.ipee@yandex.ru
Поступила в редакцию 02.09.2019
После доработки 20.10.2019
Принята к публикации 20.10.2019
Аннотация
По результатам комплексного анализа субфоссильных остатков водорослей и водных беспозвоночных охарактеризованы тафоценозы рецентных отложений 24 малых водоемов, расположенных двумя группами (по 12 водоемов в каждой) в Рузском и Шатурском районах Московской области. Пробы верхних слоев донных отложений исследовали по методике Н.Н. Смирнова, совмещающей групповой альго-зоологический и кладоцерный анализы. Сравнение полученных результатов с собственными и литературными данными показало, что все исследованные в рамках данной работы водоемы по составу тафоценозов существенно отличаются от более крупных и глубоких озер. Относительное обилие раковинных амеб в тафоценозе может служить критерием для оценки его размеров и глубины, что может быть ценно при работе с погребенными отложениями. По составу субфоссильных комплексов ветвистоусых ракообразных возможно четко дифференцировать водоемы в соответствии с их локалитетом, что указывает на существование региональных ассоциаций Cladocera. При этом различия в субфоссильных комплексах Cladocera обнаруживаются на уровне малочисленных и редких в живых сообществах видов.
Субфоссильные остатки водорослей и беспозвоночных различных групп широко используются в качестве маркеров в палеолимнологии и палеоэкологии (Stone et al., 2011; Frolova et al., 2016; Tolotti et al., 2016). Как правило, их анализ проводят с целью реконструкции динамики состава биоценозов и дальнейшей ее экстраполяции на изменения факторов (в основном, абиотических) среды в прошлом (Millet et al., 2009; Frolova et al., 2013). Остатки организмов разных групп находятся в тафоценозе отнюдь не в тех соотношениях и составе, в которых они находились в живом сообществе (Kattel et al., 2007, López-Blanco et al., 2013). В процессе седиментации и дальнейших диагенетических преобразований отложений огромное число биологических остатков уничтожается, причем избирательно. При этом из формируемого тафоценоза могут полностью выпадать крупные группы организмов (например, пресноводные Copepoda и некоторые группы Cladocera (Жаров, Котов, 2017)), а соотношения иных может принципиально изменяться.
Вместе с тем, очевидно, что донные отложения и заключенные в них тафоценозы являются фактически единственным источником информации об истории исчезнувших и существующих ныне водных экосистем, поэтому отвергать их изучение по причине больших погрешностей – недопустимо. Тем не менее, необходимо не только критически подходить к оценке и интерпретации полученных результатов (в особенности, количественных данных), но и предпринимать специальные исследования, направленные на выявление закономерностей формирования субаквальных тафоценозов и решение вопросов их сопоставимости с живыми сообществами. В связи с этим нам представляется важным исследовать рецентные отложения водоемов, характеризующие водные ценозы последних нескольких лет или (значительно реже) десятилетий. Такой подход позволяет сопоставлять полученную тафоценотическую картину с текущей ситуацией в конкретном водоеме, а также сравнивать между собой тафоценозы водоемов, различающихся по определенным очевидным параметрам. Накопление подобных наблюдений может в дальнейшем существенно облегчить и уточнить интерпретацию данных, получаемых при работе с разрезами и кернами субаквальных отложений. Наиболее подходящими для подобных исследований объектами нам представляются именно небольшие и относительно неглубокие водоемы, поскольку они, как правило, более однородны с точки зрения морфологии котловины и экологических условий, нежели большие озера, имеющие множество участков с принципиально разными условиями. Их небольшие размеры позволяют при необходимости быстро производить оценку грунтов, растительности, рельефа дна, некоторых гидрохимических показателей и иных характеристик. Кроме того, для некоторых озер нами (Zharov et al., 2018) и другими авторами (Nevalainen, 2011) ранее была показана возможность неравномерного распределения биологических остатков на дне водоема, что существенно осложняет адекватную оценку тафоценозов. Малые водоемы, в силу перечисленных выше причин, дают основание предполагать гораздо более благоприятные условия для достаточно равномерного распределения остатков альгофлоры и беспозвоночных по площади дна в процессе седиментации.
Применяемый нами метод комплексного анализа субфоссильных остатков в донных отложениях зародился в работах Кордэ (1960), а затем существенно дополнен и сформулирован Смирновым (1978, 2010). Особенностью данного метода, как следует из его названия, является одновременный качественный и количественный учет всех доступных для определения субфоссильных остатков водорослей и водных беспозвоночных хотя бы до таксонов среднего и высокого ранга. Анализ включает три уровня, отличающихся степенью таксономической детализации. Такой подход позволяет в итоге получить список групп и видов, а также численно оценить обилие каждого из них в тафоценозе. Это выгодно отличает данный метод от широко используемых методов палеолимнологического исследования, основанных на подробном анализе только одной, реже – нескольких групп организмов. В последнем случае теряется информация о многих других компонентах субфоссильного сообщества (которая также может быть крайне полезной в рамках проводимых палеоэкологических реконструкций), а также остается невыясненным место, занимаемое исследуемой “индикаторной” группой в палеосообществе конкретного водоема.
В целом необходимо отметить, что методы палеолимнологии, основанные на анализе биологических остатков, представляются весьма перспективными для исследования современного состояния пресноводных экосистем и отдельных таксоценозов. В некоторых случаях они даже могут оказаться предпочтительнее традиционных гидробиологических методов исследования, поскольку существенно нивелируют ошибки, связанные с крайне неравномерным распределением представителей фауны в пространстве и времени. По этой причине исследование даже одной пробы донных отложений зачастую дает список групп и видов более полный, нежели стандартные исследования живых сообществ, требующие отбора “живых” проб во многих точках каждого водоема в течение длительного времени (Frey, 1960; Goulden, 1964; Смирнов, 2010; López-Blanco et al., 2013). Однако следует принимать во внимание, что не все группы гидробионтов в полной мере представлены остатками в отложениях.
В рамках данной работы мы ставили целью установление сходств и различий тафоценозов разнотипных малых водоемов (небольших прудов, в том числе водопойных, карьеров, затопленных оврагов, бочажин, ирригационных и дренажных каналов) Московской обл. Предполагалось, что сходные по определенным параметрам (таким как происхождение, форма котловины, наличие либо отсутствие течения, ихтиофауны, хозяйственное назначение и использование) водоемы будут иметь более сходные тафоценозы, нежели водоемы, значительно различающиеся по этим параметрам.
МАТЕРИАЛ И МЕТОДЫ
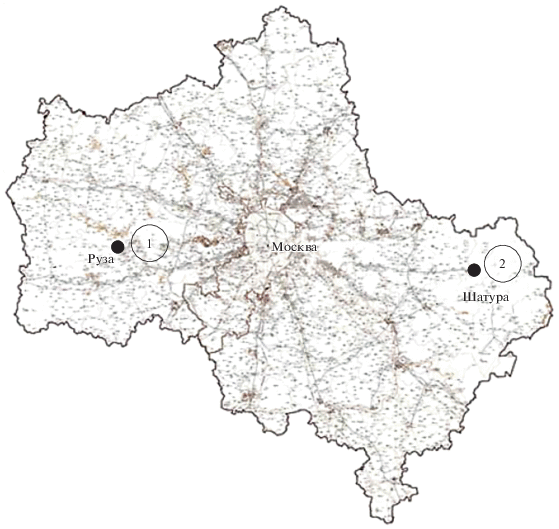
Пробы донных отложений были собраны в 2012 и 2013 гг. в 24 небольших водоемах Шатурского и Рузского районов Московской обл. В каждом районе было исследовано по 12 водоемов. Критериями в их выборе служили небольшой размер (максимальный диаметр до 50 м) и глубина (не более 2 м) и непрерывное существование (без периодов полного пересыхания) на протяжении, по крайней мере, последних 10 лет. Иные факторы, такие как происхождение, антропогенная нагрузка и загрязненность, форма котловины и т.п. не являлись селективными, в результате чего в исследование вошли довольно разнообразные водоемы – небольшие песчаные котлованы, озерца в естественных углублениях рельефа, мелиоративные и дренажные каналы, сельские пруды и т.д. Расстояние между наиболее удаленными друг от друга водоемами внутри каждой территориальной (шатурской и рузской) группы составляло около 10 км. Расстояние между двумя группами – около 200 км (рис. 1).
Рис. 1.
Расположение исследованных групп водоемов на карте Московской области: 1 – водоемы рузской группы, 2 – водоемы шатурской группы.
В наиболее глубокой точке каждого водоема (достигая ее с берега либо зайдя вброд) отбирали один образец из верхних 2–3 см донных отложений с помощью шприца объемом 50 см3, закрепленного на штанге. Образцы ничем не фиксировали и сохраняли в обводненном состоянии при температуре не выше 10°С до момента анализа.
Для исследования изготовляли временные препараты, нанося на стекло 1 каплю донного материала и 2–3 капли безводного глицерина. Пробу отложений перед этим тщательно перемешивали. Химической или физической обработке материал не подвергали, во избежание уничтожения либо утери наиболее мелких и хрупких фрагментов. Субфоссилии учитывали тотально, т.е. определению и подсчету подлежали все идентифицируемые остатки водорослей и водных беспозвоночных. В соответствии с методикой, предложенной Н.Н. Смирновым, анализ проводили в три этапа (уровня). На первом уровне учитывали все зоогенные остатки суммарно (как группу Animalia), а также остатки водорослей и цианобактерий по крупным таксономическим группам (Bacillariophyta, Desmidiales, Cyanophyta и др.). Второй этап анализа включал только зоогенные остатки, с дифференциацией их на уровне отрядов и семейств. В качестве третьего уровня выступал широко известный кладоцерный анализ, где остатки представителей Cladocera учитывались по видам. Каждый встреченный фрагмент, доступный для идентификации до соответствующего таксона, учитывали в качестве одной условной единицы. На каждом из трех уровней анализа насчитывали не менее 200 субфоссильных остатков. Таким образом, было определено относительное обилие в тафоценозе всех групп и видов гидробионтов, включенных в исследование.
Статистический анализ полученных результатов осуществляли в программах Статистика 6.0 и PAST. Были проведены кластерный анализ (UPGMA, Метод попарного внутригруппового невзвешенного среднего, с использованием индекса сходства Раупа-Крика), и анализ главных компонент (PCA). Кластеризация и анализ главных компонент были выполнены как на основе данных о присутствии видов кладоцер, так и с использованием информации об их относительном обилии в тафоценозе каждого водоема.
Достоверность различий между группами водоемов оценивали с помощью критерия Манна-Уитни.
Сравнение результатов анализа зоогенных остатков проводили, используя в качестве референтных собственные неопубликованные данные по озерам Белое-бордуковское, Черное-бордуковское, Власовское, Лемешинское, а также данные из литературных источников по озерам: Кендур (Zharov et al., 2018), Глубокое (Рузский р-н), Дубровское, Змеиное, Мотыкино, Хотавец, Пустынное (Вологодская обл.), Светлояр (Нижегородская обл.), Севан (Армения), Кинерет (Израиль), Манагуа (Никарагуа) (Смирнов, 2010).
РЕЗУЛЬТАТЫ
Трехуровневый анализ, методика проведения которого описана выше, позволил выяснить относительное обилие в тафоценозе каждой группы организмов, включенной в исследование, определить доминантные, субдоминантные и третьестепенные группы и виды.
Уровень I. Общий альго-зоологический анализ. В отложениях подавляющего большинства исследованных водоемов в изобилии были встречены как зоогенные остатки, так и остатки альгофлоры. В водоемах шатурской группы обнаружено значительное присутствие двух групп водорослей – диатомовых Bacillariophyta (среднее обилие 26%) и десмидиевых (Charophyta: Desmidiales; среднее обилие 28%), которые часто занимают доминантное положение в тафоценозе. Остатки десмидиевых присутствуют и в отложениях многих водоемов рузской группы, однако их доли здесь, как правило, невелики (4% в среднем). На диаграмме (рис. 2) показаны соотношения остатков наиболее массовых групп в отложениях исследованных водоемов. Из диаграммы видно, что отложения шатурских водоемов характеризуются более высоким содержанием остатков десмидиевых водорослей. Достоверность различий по относительному обилию Desmidiales между шатурской и рузской группами была очень высокой (U = 6, P < 0.01). Зеленые водоросли (преимущественно, представители рода Pediastrum) встречены во всех шатурских водоемах, однако везде занимают третьестепенное место, уступая вышеуказанным таксонам водорослей и остаткам беспозвоночных. В отложениях изученных водоемов Рузского р-на зеленые водоросли не обнаружены. Сине-зеленые Cyanophyta и желто-зеленые Xanthophyceae (в основном, Tribonema sp.) водоросли присутствуют в рецентных отложениях водоемов обеих территориальных групп, но их относительное обилие в большинстве случаев невелико. Тем не менее, в тафоценозах трех водоемов рузской группы остатки Tribonema доминируют. Подробные результаты общего альго-зоологического анализа приведены в табл. 1.
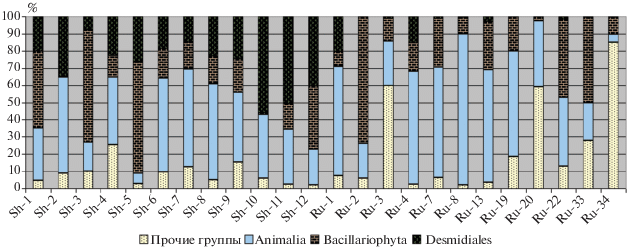
Таблица 1.
Относительное обилие остатков водорослей и зоогенных остатков в тафоценозах малых водоемов по результатам I уровня анализа
| Водоем | Группа | |||||
|---|---|---|---|---|---|---|
| Bacillariophyta | Desmidiales | Chlorophyta | Cyanophyta | Xanthophyceae | Animalia | |
| Sh-1 | 45 | 20 | 0 | 0 | 4.5 | 30.5 |
| Sh-2 | 0 | 35 | 6 | 2 | 1 | 56 |
| Sh-3 | 65 | 8 | 10 | 0 | 0 | 17 |
| Sh-4 | 12.5 | 22.5 | 12 | 13.5 | 0 | 39.5 |
| Sh-5 | 64.5 | 26.5 | 2.5 | 0.5 | 0 | 6 |
| Sh-6 | 17.5 | 18.5 | 4.5 | 2.5 | 2.5 | 54.5 |
| Sh-7 | 15.5 | 15 | 5 | 7.5 | 0 | 57 |
| Sh-8 | 16 | 23 | 4 | 1 | 0 | 56 |
| Sh-9 | 19.5 | 24.5 | 12.5 | 0 | 3 | 40.5 |
| Sh-10 | 1 | 56 | 0.5 | 4 | 1.5 | 37 |
| Sh-11 | 15 | 50.5 | 0 | 0 | 2.5 | 32 |
| Sh-12 | 37 | 40 | 1 | 1 | 0 | 21 |
| Ru-1 | 8.5 | 20.5 | 0 | 4.5 | 3 | 63.5 |
| Ru-2 | 74 | 0 | 0 | 5 | 1 | 20 |
| Ru-3 | 13.5 | 1 | 0 | 1 | 59 | 25.5 |
| Ru-4 | 16.5 | 15.5 | 0 | 2 | 0.5 | 65.5 |
| Ru-7 | 28.5 | 1 | 0 | 1 | 5.5 | 64 |
| Ru-8 | 9.5 | 0.5 | 0 | 1.5 | 0.5 | 88 |
| Ru-13 | 28 | 3 | 0 | 3.5 | 0 | 65.5 |
| Ru-19 | 20 | 0 | 0 | 15.5 | 3 | 61.5 |
| Ru-20 | 2.5 | 0 | 0 | 20.5 | 38.5 | 38.5 |
| Ru-22 | 45.5 | 1.5 | 0 | 5 | 8 | 40 |
| Ru-33 | 49.5 | 0.5 | 0 | 3 | 25 | 22 |
| Ru-34 | 10 | 0.5 | 0 | 2.5 | 82.5 | 4.5 |
Уровень II. Групповой зоологический анализ. Результаты данного этапа исследования представлены в табл. 2. Наиболее массовыми зоогенными компонентами тафоценозов во всех случаях оказались остатки Cladocera и раковинных амеб (главным образом, представителей отряда Arcellinida). Остатки представителей других групп беспозвоночных были немногочисленны, однако регулярно встречались в отложениях водоемов обеих территориальных групп. Исключение составили раковинные инфузории Tintinnidae, присутствовавшие в отложениях только одного водоема (Ru-13).
Таблица 2.
Относительное обилие остатков беспозвоночных различных групп в тафоценозах малых водоемов по результатам II уровня анализа
| Водоем | Группа | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cladocera | Chaoboridae | Chironomidae | Ceratopogonidae | Insecta aliae | Arcellinida | Turbellaria | Bryozoa | Ostracoda | Acari | Tintinnidae | |
| Sh-1 | 60 | 0.5 | 3.5 | 0 | 2 | 30 | 2.5 | 0.5 | 1 | 0 | 0 |
| Sh-2 | 34.5 | 0 | 1 | 0 | 3.5 | 59.5 | 1.5 | 0 | 0 | <0.5 | 0 |
| Sh-3 | 59.5 | 0 | 1 | <0.5 | 1.5 | 37.5 | 0.5 | <0.5 | 0 | 0 | 0 |
| Sh-4 | 38.5 | 0 | 1.5 | <0.5 | 4.5 | 53 | 1 | 0.5 | 1 | <0.5 | 0 |
| Sh-5 | 40.5 | 0 | 0.5 | 0 | 1 | 57.5 | 0.5 | 0 | 0 | 0 | 0 |
| Sh-6 | 45.5 | 0.5 | 2.5 | 1 | 2.5 | 45 | 2.5 | 0 | 0.5 | 0 | 0 |
| Sh-7 | 29 | 0 | 0.5 | <0.5 | 2.5 | 66 | 2 | 0 | 0 | 0 | 0 |
| Sh-8 | 56 | <0.5 | 5.5 | <0.5 | 4.5 | 32 | 2 | <0.5 | 0 | 0 | 0 |
| Sh-9 | 42 | 1 | 2 | 0 | 6.5 | 46.5 | 1.5 | 0.5 | <0.5 | 0 | 0 |
| Sh-10 | 71.5 | 0.5 | 1 | <0.5 | 1.5 | 24 | 1.5 | <0.5 | <0.5 | <0.5 | 0 |
| Sh-11 | 53.5 | 0 | 1.5 | <0.5 | 2.5 | 41 | 1.5 | 0 | 0 | 0 | 0 |
| Sh-12 | 45.5 | 0 | 0.5 | 1 | 2 | 45.5 | 4 | 0.5 | 1 | 0 | 0 |
| Ru-1 | 28 | 0 | 2.5 | 1 | 12 | 56.5 | 0 | 0 | 0 | 0 | 0 |
| Ru-2 | 5.5 | 0 | 1.5 | 0 | 10 | 81 | 2 | 0 | 0 | 0 | 0 |
| Ru-3 | 15 | 0 | 2.5 | <0.5 | 5 | 76.5 | 1 | 0 | <0.5 | <0.5 | 0 |
| Ru-4 | 15 | <0.5 | 0.5 | <0.5 | 5 | 78 | 0.5 | 0.5 | 0.5 | 0 | 0 |
| Ru-7 | 16 | 0 | 1.5 | 0 | 3 | 79 | 0.5 | 0 | 0 | 0 | 0 |
| Ru-8 | 7 | 0 | 1 | <0.5 | 1.5 | 89.5 | 0.5 | 0 | 0.5 | <0.5 | 0 |
| Ru-13 | 33 | 0 | 1 | 0 | 4.5 | 50.5 | 0 | 0 | 0 | 0 | 11 |
| Ru-19 | 22 | 0 | 1 | <0.5 | 5 | 71 | 1 | 0 | <0.5 | <0.5 | 0 |
| Ru-20 | 15 | 0 | 0.5 | 0.5 | 12 | 71 | 0.5 | 0 | 0.5 | 0 | 0 |
| Ru-22 | 27 | 0.5 | 0.5 | 0.5 | 9.5 | 60.5 | 1.5 | 0 | <0.5 | <0.5 | 0 |
| Ru-33 | 17 | 0 | 0 | <0.5 | 10 | 69.5 | 1.5 | 0 | 2 | 0 | 0 |
| Ru-34 | 26 | 0 | 2.5 | 0 | 15 | 52 | 1 | 0 | 3.5 | 0 | 0 |
Относительное обилие раковин тестацид оказалось достаточно высоким во всех исследованных водоемах. При этом в отложениях водоемов Рузского р-на их доли были практически во всех случаях выше (в среднем 70%), чем в шатурских (45%). Различия между выборками в высокой степени достоверны (U = 2, P < 0.01).
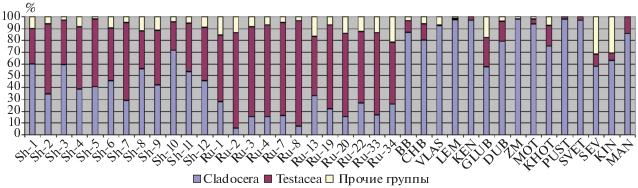
Сравнение результатов по 24 исследованным малым водоемам с собственными неопубликованными данными о тафоценозах относительно больших подмосковных озер, а также с литературными сведениями об отложениях озер иной локализации (Смирнов, 2010), показало, что в осадках крупных и глубоких водоемов относительное обилие субфоссильных раковинных амеб обычно существенно ниже (см. рис. 3). Различия между выборками высоко достоверны (U = 1, P < 0.01).
Рис. 3.
Относительное обилие субфоссильных остатков Cladocera, раковинных амеб и прочих групп беспозвоночных в тафоценозах малых водоемов и 15 озер.
Уровень III. Кладоцерный анализ. Исследование субфоссильных остатков выявило, по меньшей мере, 29 видов ветвистоусых ракообразных в отложениях исследованных водоемов. Некоторые остатки были идентифицированы только до группы видов в силу невозможности установления видовой принадлежности из-за отсутствия надежных определительных признаков. Например, остатки крупных видов рода Alona (A. affinis и A. quadrangularis), мелких видыов рода Alona (A. rectangula, A. guttata, A. costata и A. rustica) учитывали суммарно как группы “Alona large sized” и “Alona small sized” соответственно, а также фрагменты экзоскелетов дафний (Daphnia sp.).
Присутствие видов ветвистоусых ракообразных в отложениях исследованных водоемов представлено в табл. 3.
Таблица 3.
Присутствие видов Cladocera в тафоценозах малых водоемов
| Водоем | Вид | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bosmina cf. longirostris | Daphnia sp. | Ceriodaphnia sp. | Diaphanosoma sp. | Sida crystallina | Simocephalus sp. | Ophryoxus gracilis | Chydorus sphaericus s.l. | Alona sp. (large sized) | Alona sp. (small sized) | Alona intermedia | Alona guttata | Alona rectangula | Rhynhotalona falcata | Disparalona rostrata | Monospilus dispar | Graptoleberis testudinaria | Alonella excisa | Alonella exigua | Alonella nana | Camptocercus rectirostris | Acroperus harpae s. l. | Oxyurella tenuicaudis | Kurzia latissima | Eurycercus lamellatus | Leydigia sp. | Pleuroxus sp. | Pleuroxus trigonellus | Pleuroxus truncatus | Pleuroxus uncinatus | Pleuroxus laevis | |
| Sh-1 | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||||
| Sh-2 | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||||||
| Sh-3 | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||||||
| Sh-4 | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||||
| Sh-5 | + | + | + | + | + | + | + | ||||||||||||||||||||||||
| Sh-6 | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||
| Sh-7 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||
| Sh-8 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||||||||
| Sh-9 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||
| Sh-10 | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||||
| Sh-11 | + | + | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||||
| Sh-12 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||
| Ru-1 | + | + | + | + | + | + | + | + | + | + | + | ||||||||||||||||||||
| Ru-2 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-3 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-4 | + | + | + | + | + | + | + | + | + | ||||||||||||||||||||||
| Ru-7 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-8 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-13 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-19 | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||||
| Ru-20 | + | + | + | + | + | + | |||||||||||||||||||||||||
| Ru-22 | + | + | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||
| Ru-33 | + | + | + | + | + | + | + | + | + | + | |||||||||||||||||||||
| Ru-34 | + | + | + | + | + | + | + | ||||||||||||||||||||||||
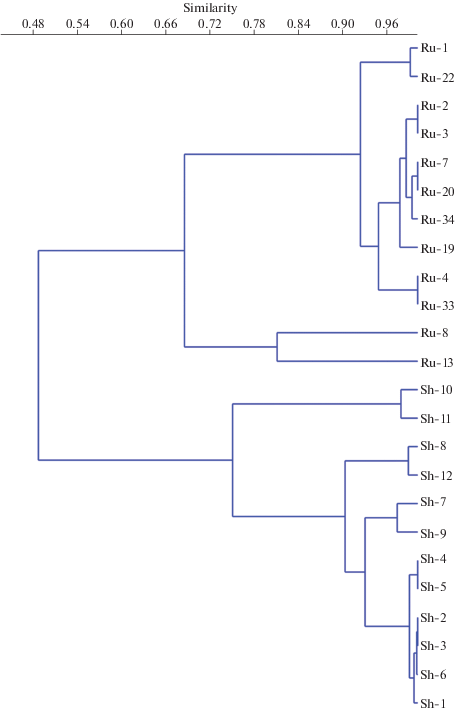
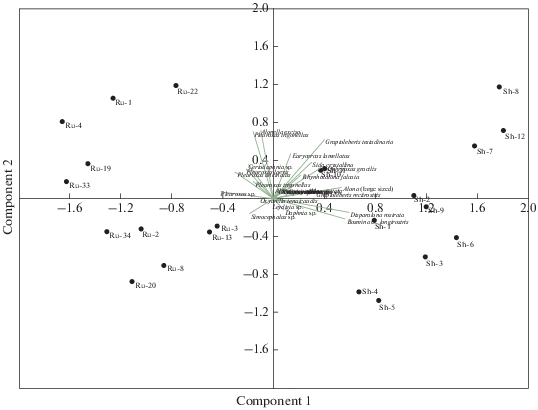
Кластерный анализ и анализ главных компонент были применены к данным по присутствию видов Cladocera в исследованных тафоценозах. В обоих случаях наблюдалось разделение водоемов на две группы, в полном соответствии с их территориальной принадлежностью (рис. 4, 5). Четкая классификация водоемов по локалитету имела место только при использовании данных о присутствии/отсутствии видов. Введение данных об относительном обилии видов приводило к перегруппировке водоемов в единый массив, без формирования каких-либо отчетливых групп и кластеров.
ОБСУЖДЕНИЕ
На всех трех примененных уровнях анализа тафоценозов между водоемами двух территориальных групп обнаружены существенные различия. Состав субфоссильной альгофлоры на уровне крупных таксонов был довольно сходным, однако показатели среднего относительного обилия некоторых из них значительно разнились. Так, в отложениях водоемов шатурской группы гораздо обильнее были представлены десмидиевые водоросли (преимущественно, представители родов Staurastrum, Cosmarium, Xanthidium, Euastrum, Closterium, Penium), среднее относительное обилие которых внутри этой группы составило 28%, против 4% в водоемах рузской группы (P < 0.01). По всей видимости, это связано с тяготением многих десмидиевых к болотам и водоемам с кислой водой (Brook, 1981; Coesel, Meesters, 2013). В водоемах Шатурского р-на pH воды, как правило, ниже 6, что связано с обилием торфяников и болот. Примечательно, однако, что даже в водоемах, не имеющих прямого контакта с торфянистыми грунтами и заболоченными территориями, присутствуют десмидиевые водоросли. Вероятно, это обусловлено геохимическими свойствами подстилающих пород и грунтовых вод.
Большая распространенность и более высокие показатели обилия желто-зеленых водорослей Xanthophyta (представленных преимущественно Tribonema sp.) в водоемах рузской группы связаны, по-видимому, с тем, что эти водоемы в большинстве своем расположены на открытых пространствах, а наибольшего развития нитчатки Tribonema достигают в хорошо освещенных и прогреваемых мелководных участках. Водоемы шатурской группы находятся преимущественно среди лесных массивов, и поэтому более затенены.
Зоогенные остатки в отложениях водоемов обеих групп были представлены комплексом таксонов, в целом характерным для пресных водоемов средней полосы России. Лидирующее положение по относительному обилию в тафоценозах занимали ветвистоусые ракообразные и раковинные амебы, причем последние были достоверно (P < 0.01) более многочисленны в водоемах рузской группы. Возможно, причиной этому является в среднем несколько меньшая глубина рузских водоемов (преимущественно в пределах 0.5–1 м) по сравнению с шатурскими (в среднем около 1.5 м). При исследовании многих других водоемов ранее мы отмечали, что относительное обилие раковин тестацид в донных отложениях в некоторой степени связано с глубиной водоема – в тафоценозах более глубоких водоемов тестацид, как правило, несколько меньше. На это же указывают данные, полученные при изучении пространственного распределения различных зоогенных остатков в отложениях озера Кендур (Zharov et al., 2018), где доля тестацид уменьшалась с возрастанием глубины и удаленности от берега.
Для проверки предположения о взаимосвязи обилия тестацид в отложениях с глубиной и размерами водоема было проведено сравнение результатов, полученных в ходе данного исследования, с литературными сведениями о составе тафоценозов крупных озер, исследованных по аналогичной методике, а также с собственными (ранее не опубликованными) данными по некоторым озерам Шатурского р-на (см. рис. 3). Было установлено, что в тафоценозах малых водоемов относительное обилие раковин тестацид достоверно выше (P < 0.01), чем в озерах. Вероятно, такая ситуация обусловлена тем, что крупные и глубокие водоемы имеют большой объем пелагиали, обычно густо населенной планктонными видами кладоцер. Раковинные амебы в большинстве своем являются донными организмами и обитателями зарослей, т.е. так или иначе связаны с подводными субстратами (хотя существуют и некоторые планктонные виды) (Мазей, Цыганов, 2006). Возрастание объема пелагиали, таким образом, приводит к увеличению количества остатков ветвистоусых ракообразных (в основном за счет планктонных видов) или других пелагических организмов на единицу площади дна и, как следствие, уменьшению доли тестацид в формирующемся тафоценозе. Некоторую роль может также играть попадание в донные отложения аллохтонных раковин тестацид с прилегающих к водоему влажных биотопов, которое может вызывать заметное повышение их обилия в тафоценозе небольшого водоема (при наличии обширных заболоченных участков на побережье), однако вряд ли окажется существенным в масштабах крупного озера. Таким образом, доля раковин тестацид в субаквальных тафоценозах может служить своеобразным маркером при идентификации отложений палеоводоемов. Однако следует иметь в виду, что их высокое относительное обилие в тафоценозе может являться результатом деградации остатков других массовых групп (например, кладоцер). Такая ситуация наблюдалась нами, в частности, в степных эфемерных водоемах, в отложениях которых раковины тестацид были супердоминантами, а иногда являлись единственными зоогенными компонентами тафоценозов (Жаров, Котов, 2017). В данном случае их чрезвычайное преобладание было вызвано, по всей очевидности, сразу несколькими факторами: (1) преобладанием в планктоне эфемерных водоемов представителей Daphniidae, остатки которых известны плохой сохраняемостью (Sarmaja-Korjonen, 2007; Nykänen et al., 2009), (2) деятельностью детритофагов Triops cancriformis, способных уничтожать хитиновые остатки других ракообразных, и (3) сезонным пересыханием водоемов, также приводящем к разрушению некоторых хитиновых остатков. Необходимо отметить, что известны редкие случаи доминирования раковинных амеб и в озерных тафоценозах (Кордэ, 1960; Smirnov, 1986), однако все они относились лишь к отдельным периодам в истории существования данных озер. Их преобладание в этих случаях могло быть обусловлено тафономическими причинами (например, деградацией остатков кладоцер) или значительным падением уровня воды в водоемах.
Одной из причин уменьшения доли раковинных амеб в тафоценозах с возрастанием глубины может служить и снижение их видового разнообразия в рецентных сообществах на больших глубинах, показанное ранее на примере нескольких Шатурских озер (Tsyganov et al., 2019). В качестве объяснения этого явления рассматриваются менее благоприятные условия для бентосных сообществ на больших глубинах, а также снижение аккумуляции раковинок, переотложенных со средних и малых глубин из-за их низких флотационных свойств (там же).
Анализ результатов исследования субфоссильных кладоцер методами кластерного анализа (рис. 4) и анализа главных компонент (рис. 5) выявил, что по качественному составу субфоссильных комплексов Cladocera водоемы рузской и шатурской групп четко обособляются. При этом значительную роль играют преимущественно редкие и малочисленные виды (за исключением Bosmina longirostris), тогда как доминантные могут быть общими – Chydorus sphaericus s. l. и B. longirostris. Ch. sphaericus является широко распространенным эвритопным видом, обитающим как в зарослях макрофитов, так и в пелагиали, B. longirostris – планктонный вид, обитающий, главным образом, в пелагиали небольших водоемов (Смирнов, 2010). Преобладание этих видов в тафоценозах является типичной для многих водоемов ситуацией (Birks et al., 1976; Смирнов, 1978; 2010). B. longirostris оказалась более характерна для водоемов шатурской группы, что, возможно, связано с их несколько большей глубиной и, следовательно, бо́льшим объемом пелагиали.
В шатурских водоемах были отмечены, по меньшей мере, 10 видов, не найденных в отложениях водоемов рузской группы. Это Diaphanosoma brahyurum, Sida crystallina, Ophryoxus gracilis, Camprocercus rectirostris, Acroperus harpae (s. l.), Monospilus dispar, Rhynhotalona falcata, Disparalona rostrata, Alona intermedia и Alonella nana.
Представители рода Alona из группы мелких (“small sized”) видов по большинству субфоссильных остатков трудноопределимы, поэтому A. guttata и A. rectangula, присутствие которых отмечено в табл. 3 только для шатурских водоемов, могут в действительности присутствовать и в водоемах рузской группы, однако не быть при этом учтенными из-за отсутствия диагностически значимых субфоссильных остатков. То же касается и видов рода Pleuroxus (P. truncatus, P. uncinatus, P. trigonellus и P. laevis), которые по некоторым субфоссильным фрагментам трудноразличимы. Их состав можно считать адекватно выясненным только для отложений рузских водоемов, где они встречаются существенно чаще.
В отложениях водоемов рузской группы в качестве единичных находок были отмечены Leydigia sp., Oxyurella tenuicaudis, Diaphanosoma brachyurum и Simocephalus cf. vetulus, не обнаруженные в шатурских водоемах. Последние два вида являются широко распространенными, и их отсутствие в отложениях прочих водоемов обеих территориальных групп вызвано, скорее всего, тафономическими причинами. Как и другие представители семейств Sididae и Daphniidae, они известны плохой сохраняемостью и слабой представленностью в тафоценозах.
Alonella nana является регулярным и зачастую массовым компонентом в отложениях шатурских водоемов, тогда как в рузских, по-видимому, отсутствует. Как и в случае с бо́льшим распространением в шатурских водоемах десмидиевых водорослей, причина этого кроется, вероятно, в слабокислых мягких водах, характерных для этого района. Многие авторы отмечали приуроченность этого литорального вида к водам с низкими значениями pH (Sandøy, Nilssen, 1986; Walseng, Schartau, 2001). Менее многочисленным, но также более характерным для водоемов шатурской группы, является зарослевый вид Graptoleberis testudinaria, который, по некоторым данным, также предпочитает водоемы со слабокислой водой (Fryer, 1993).
M. dispar, R. falcate и D. rostrata являются бентосными видами, населяющими преимущественно открытые участки литорали (Van De Bund, Spaas, 1996). Присутствие их в шатурских водоемах может быть обусловлено наличием во многих из них мелководий с песчаным слабо заиленным дном, свободных от зарослей макрофитов.
Таким образом, субфоссильные комплексы Cladocera в водоемах двух территориальных групп отличаются главным образом на уровне субдоминантных и редких видов, представляющих бентосную и зарослевую фауну. Ранее многими авторами предпринимались попытки классифицировать субфоссильные комплексы ветвистоусых ракообразных по преобладающим родам и видам (см. Смирнов, 2010) с присвоением им соответствующих названий, по аналоги с тем, как это делал Литынски (по: Sramek-Husek, 1962) для живых ассоциаций кладоцер. Чаще всего отмечается преобладание в тафоценозе эвритопных и пелагических видов – Chydorus sphaericus (тип тафоценоза “Chydoretum”), Bosmina longirostris, B. coregoni (“Bosminetum”). Реже преобладают литоральные виды, причем доминирование в субфоссильных комплексах известно для очень немногих из них – Acroperus sp. (Ravera, Parise, 1978), Alona affinis (Смирнов, 2010), Alonella excisa (Koff et al., 2005), Alonella nana (Sarmaja-Korjonen et al., 2006), Pleuroxus uncinatus (Смирнов, 1978) и Monospilus sp. Результаты нашего исследования показывают, что в классификации сообществ кладоцер важную роль могут играть малочисленные виды, что подтверждает целесообразность применения анализа остатков в донных отложениях. Исследования живых сообществ традиционными гидробиологическими методами могут не выявить присутствие этих видов.
ЗАКЛЮЧЕНИЕ
Комплексный анализ биологических остатков в донных отложениях показал высокую эффективность в выявлении сходств и различий разнотипных водоемов по составу и структуре их тафоценозов. Возможность достижения подобных результатов с помощью стандартных гидробиологических методов исследования сомнительна, т.к. в таком случае было бы проблематично выяснить взаимные соотношения организмов различных групп в сообществах, а также избежать больших погрешностей, связанных с неоднородностью их пространственного и временно́го распределения. В данном исследовании показана возможность выявления региональных особенностей состава субаквальных тафоценозов на разных таксономических уровнях. В связи с этим данный палеолимнологический подход может стать методом выбора при проведении био- и палеогеографических исследований водных организмов. Данные о различиях и их вероятных причинах, полученные при сравнении рецентных тафоценозов, могут быть полезны при интерпретации результатов исследования более древних субаквальных отложений.
Список литературы
Жаров А.А., Котов А.А., 2017. Тафоценозы эфемерных водоемов степной зоны Европейской части России по данным альго-зоологического анализа рецентных отложений // Известия Российской академии наук. Серия биологическая. № 3. С. 312–321.
Кордэ Н.В., 1960. Биостратификация и типология русских сапропелей. М.: Изд-во АН СССР. 219 с.
Мазей Ю.А., Цыганов А.Н., 2006. Пресноводные раковинные амебы. М.: Товарищество научных изданий КМК. 300 с.
Смирнов Н.Н., 1978. Методы и некоторые результаты исторической биоценологии ветвистоусых ракообразных // Экология сообществ озера Глубокого. Отв. ред. Поляков Г.Д. М.: Наука. С. 105–173.
Смирнов Н.Н., 2010. Историческая экология пресноводных зооценозов. М.: Товарищество научных изданий КМК. 225 с.
Birks H.H., Whiteside M.C., Stark D.M., Bright R.C., 1976. Recent paleolimnology of three lakes in northwestern Minnesota // Quaternary Research. V. 6. № 2. P. 249–272.
Brook A.J., 1981. The biology of desmids. Berkeley: Univ. of California Press. V. 16.
Coesel P.F.M., Meesters K.J., 2013. European Flora of the Desmid Genera Staurastrum and Staurodesmus: Identification Key for Desmidiaceae-Morphology-Ecology and Distribution-Taxonomy. Zeist: KNNV Publishing. 358 p.
Frey D.G., 1960. On the occurrence of cladoceran remains in lake sediments // Proceedings of the National Academy of Sciences of the USA. V. 46. № 6. P. 917–920.
Frolova L., Nazarova L., Nigamatzyanova G., Mukhametgaliev N.R., Krylov P., Frolova A., 2013. Environmental and climatic factors influencing the distribution of sub-fossil cladocerans in thermokarst lakes in northeastern Siberia, Russia // Environmental Radioecology and Applied Ecology. V. 19. № 3–4. P. 3-9.
Frolova L., Ibragimova A., Fedorova I., 2016. Stratigraphy of Cladocera in a core from a Yamal peninsula lake (Arctic Russia) // International Multidisciplinary Scientific GeoConference: SGEM: Surveying Geology & Mining Ecology Management. V. 2. P. 579–585.
Fryer G., 1993. Variation in acid tolerance of certain freshwater crustaceans in different natural waters // Hydrobiologia. V. 250. № 2. P. 119–125.
Goulden C.B., 1964. The history of the Cladoceran fauna of Esthwaite Water (England) and its limnological significance // Archiv für Hydrobiologie. Bd. 60. № 1. S. 1–52.
Kattel G.R., Battarbee R.W., Mackay A., Birks H.B., 2007. Are cladoceran fossils in lake sediment samples a biased reflection of the communities from which they are derived? // Journal of Paleolimnology. V. 38. P. 157–181.
Koff T., Punning J.M., Sarmaja-Korjonen K., Martma T., 2005. Ecosystem response to early and late Holocene lake-level changes in lake Jjuusa, Southern Estonia // Polish Journal of Ecology. V. 53. № 4. P. 553–570.
López-Blanco C., Miracle M.R., Vicente E., 2013. Is there a bias between contemporary and subfossil cladoceran assemblages? // Limnetica. V. 32. P. 201–214.
Millet L., Arnaud F., Heiri O., Magny M., Verneaux V., Desmet M., 2009. Late-Holocene summer temperature reconstruction from chironomid assemblages of Lake Anterne, northern French Alps // The Holocene. V. 19. № 2. P. 317–328.
Nevalainen L., 2011. Intra-lake heterogeneity of sedimentary cladoceran (Crustacea) assemblages forced by local hydrology // Hydrobiologia. V. 676. P. 9–22.
Nykänen M., Vakkilainen K., Liukkonen M., Kairesalo T., 2009. Cladoceran remains in lake sediments: a comparison between plankton counts and sediment records // Journal of Paleolimnology. V. 42. P. 551–570.
Ravera O., Parise G., 1978. Eutrophication of Lake Lugano ‘read’ by means of planktonic remains in the sediment // Schweizerische Zeitschrift für Hydrologie. V. 40. № 1. P. 40–50.
Sandøy S., Nilssen J.P., 1986. A geographical survey of littoral crustacea in Norway and their use in paleolimnology // Hydrobiologia. V. 143. № 1. P. 277–286.
Sarmaja-Korjonen K., 2007. Subfossil shell margins and tail spines of Daphnia in Finnish lake sediments–is Daphnia underrepresented in Cladocera analysis // Studia Quaternaria. V. 24. P. 61–64.
Sarmaja-Korjonen, K., Nyman M., Kultti S., Väliranta M., 2006. Palaeolimnological development of Lake Njargajavri, northern Finnish Lapland, in a changing Holocene climate and environment // Journal of Paleolimnology. V. 35. № 1. P. 65–81.
Smirnov N.N., 1986. History of the zoocenosis of Lake Glubokoe according to animal remains in the bottom sediments // Hydrobiologia. V. 141. № 1–2. P. 143–144.
Sramek-Husek R., 1962. Die mitteleuropäischen Cladoceren-und Copepodengemeinschaften und deren Verbreitung in den Gewässern der CSSR // Sbornik VSCHT Technology Vody. V. 6. № 1. P. 99–133.
Stone J.R., Westover K.S., Cohen A.S., 2011. Late Pleistocene paleohydrography and diatom paleoecology of the central basin of Lake Malawi, Africa // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 303. № 1–4. P. 51–70.
Tolotti M., Milan M., Szeroczyńska K., 2016. Subfossil Cladocera as a powerful tool for paleoecological reconstruction // Advances in Oceanography and Limnology. V. 7. № 2. P. 125–130.
Tsyganov A.N., Malysheva E.A., Zharov A.A., Sapelko T.V., & Mazei Y.A., 2019. Distribution of benthic testate amoeba assemblages along a water depth gradient in freshwater lakes of the Meshchera Lowlands, Russia, and utility of the microfossils for inferring past lake water level // Journal of Paleolimnology. V. 62. № 2. P. 137–150.
Van De Bund W.J., Spaas S.J.H., 1996. Benthic communities of exposed littoral sand-flats in eighteen Dutch lakes // Netherland Journal of Aquatic Ecology. V. 30. № 1. P. 15–20.
Walseng B., Schartau A.K.L., 2001. Crustacean communities in Canada and Norway: comparison of species along a pH gradient //Acid rain 2000. Springer, Dordrecht. P. 1319–1324.
Zharov A.A., Khasanov B.F., Kotov A.A., 2018. Spatial heterogenity of the taphocenosis in lake Kendur, Moscow region, Russia, based on the results of complex zoological and cladoceran analyses // Zoologichesky zhurnal. V. 97. № 11. P. 1330–1339.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал