

Зоологический журнал, 2020, T. 99, № 5, стр. 528-543
Голоценовые насекомые Урала, центра и севера Западно-Сибирской равнины
Е. В. Зиновьев *
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: zin62@mail.ru
Поступила в редакцию 02.09.2019
После доработки 15.09.2019
Принята к публикации 20.10.2019
Аннотация
Обобщен энтомологический материал из голоценовых местонахождений Урала, Предуралья, центра и севера Западно-Сибирской равнины. Приведены данные по 38 местонахождениям, большая часть из которых имеет аллювиальный генезис. Также изучены остатки насекомых из торфяников и зоогенных отложений. Показано, что в большинстве аллювиальных местонахождений комплексы жуков соответствуют современным комплексам жуков, проживающим на этих же территориях. Динамика энтомофаун, связанная со сменой ландшафтно-климатических условий, наблюдается только на границах современных природных зон, таких как тундровая и таежная. В климатический оптимум голоцена именно на таких пограничных территориях отмечено продвижение к северу целого ряда видов, связанных с лесами, в том числе и с древесной растительностью. В пределах таежной зоны комплексы жуков из исследованных нами голоценовых отложений аллювиального и болотного генезиса идентичны современным фаунам соответствующих природных зон. Основные изменения в пределах конкретных разрезов могут объясняться сукцессионными процессами, связанными с динамикой мезоландшафтов в пределах смены русел рек, формированием стариц, последующим облесением. При этом изменения в составе растительных сообществ в голоцене, фиксируемые на палинологическом материале, не выражены в составе комплексов насекомых. Это продемонстрировано на примере двух торфяников на Полярном Урале (точка “озеро Перевальное”) и Южном Ямале (точка Нюлсавэйто). Приведенные в настоящей работе данные в целом соответствуют сведениям, полученным для территории Западной и Восточной Европы, а также Северо-Восточной Сибири. В зоогенных же отложениях (погадки птиц, норовища) могут быть найдены виды, изменения ареалов которых можно связать с климатическими изменениями в голоцене.
Голоцен характеризуется формированием природной зональности в ее современном состоянии. Данный период можно определить в качестве межледниковья, в котором находится современная цивилизация, его продолжительность составляет 10–12 тыс. лет (Хотинский, 1977, 1981; Динесман и др., 1979; Velichko et al., 1992 и др.). В системе хронологии квартера он является относительно коротким интервалом теплого климата, который наступил после длительного холодного периода и, возможно, предваряет новое похолодание. При этом динамика изменений климата, реконструированная по соотношению изотопов кислорода О18/О16, не показывает принципиальных изменений в голоцене по сравнению с предыдущим периодом (окончание МИС2) (Хотинский, 1977).
Исследование истории биогеоценозов в голоцене делится на три направления: история отдельных компонентов, развитие структуры экосистем и изучение истории взаимоотношения экосистем (Динесман и др., 1979).
Процессы развития экосистем наиболее детально исследованы на спорово-пыльцевом материале на Урале и в Западной Сибири (Волкова и др., 1989; Панова, 1996; Бляхарчук, 2000; Blyakharchuk, 2003 и др.), в Западной и Восточной Европе (Huntley, 1988, 1990; Bradshaw et al., 2003; Usinger, 2004; Голубева, 2008 и др.). Более того, полученные данные легли в основу палеоэкологических реконструкций для территории всей Северной Евразии (Tarasov et al., 1999 и др.). Детально исследованы процессы становления фаун крупных и мелких млекопитающих (Динесман, 1976; Смирнов, 1993, 1996; Бачура, Струкова, 2002; Фадеева, 2002; Косинцев, 2009; Kuzmina, 2009; Sommer et al., 2011 и др.). Кроме того, изучена динамика фаун водных беспозвоночных, в частности остракод (Хазин и др., 2016), и других групп.
Одной из наиболее информативных групп животных, позволяющих выявлять специфику развития ландшафтно-климатических условий квартера в целом, являются насекомые. Обилие их остатков в отложениях аллювиального, озерно-аллювиального генезиса может являться одним из положительных моментов, позволяющих привлекать их для палеоэкологических и палеогеографических реконструкций. Анализ ископаемых энтомофаун является важной частью комплексных исследований динамики развития природных сообществ в плейстоцене и голоцене. Они позволяют проводить реконструкции как на уровне фундаментальных экологических ниш, так и мезо- и макроклимата. Поскольку одним из важных лимитирующих факторов распространения насекомых является тип растительных сообществ, то в ряде случаев они позволяют детализировать характер растительного покрова (Назаров, 1984). Ввиду того, что хитин относительно хрупок, вероятность его переотложения и смешивания разновозрастного материала значительно ниже, чем у растений и млекопитающих. Это очень важно для реконструкций плейстоценовых наземных экосистем.
Цель настоящей работы – оценить информационную ценность жесткокрылых для воссоздания процессов развития ландшафтов в контексте развития климатической обстановки голоцена, определить типы захоронений, откуда можно было бы получать наиболее ценный палеонтологический материал для этих целей. Основой настоящей статьи являются данные, полученные автором при изучении четвертичных насекомых Урала, центра и севера Западно-Сибирской равнины, использованы также литературные сведения по этой же территории.
ХАРАКТЕРИСТИКА РАЙОНА ИССЛЕДОВАНИЙ
Работа основана на изучении остатков насекомых (главным образом, жесткокрылых), найденных в голоценовых отложениях. Энтомологический материал собран из соответствующих отложений Урала, Предуралья (Большеземельская тундра), северной и центральной частей Западной-Сибирской равнины.
Данная территория включает в себя несколько природных зон – от лесотундр, южных и типичных тундр на севере (местонахождения Гыданского п-ова, Ямала, Большеземельской тундры и Полярного Урала) до южнотаежных лесов (Усть-лог, местонахождения в долине р. Чусовая в окр. г. Екатеринбурга). Это обусловило широкое разнообразие природных условий и необходимость учета региональных особенностей современных энтомофаун для тех мест, откуда собран материал. При характеристике современных фаун использованы как данные собственных сборов (Зиновьев, Рябицев, 2000; Зиновьев, Нестерков, 2003; Зиновьев, 2007 и др.), так и сведения, взятые из литературных источников (Бухкало и др., 2011; Ухова, Ольшванг, 2014 и др.).
МАТЕРИАЛЫ И МЕТОДЫ
Отбор проб на энтомологический анализ и их дальнейшая камеральная обработка осуществлялись по стандартным методикам, принятым для данного типа анализа (Киселев, 1987). Определение найденных остатков производилось по эталонным коллекциям Зоологического музея Института экологии растений и животных УрО РАН.
Принадлежность отложений к голоцену определялась несколькими способами. Все пробы брались из разрезов, представляющих геоморфологический уровень пойменной террасы. Для части местонахождений были получены радиоуглеродные даты, представленные в табл. 1. В тексте приводятся калиброванные датировки, для чего была использована программа CalPal Online (www.calpal-online/de). Кроме того, для точек в долине р. Яда-Яходыяха получены даты гибели деревьев непосредственно из тех слоев, откуда брались пробы (табл. 1).
Таблица 1.
Абсолютные датировки для местонахождений региона
| Номер | Местонахождение | Координаты | Тип даты |
Абсолютный возраст, лет назад (л. н.) | Лаб. номер даты | ||
|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | некалиброванная | калиброванная | ||||
| 51 | Перевальное | 66°51′ | 65°41′ | 14С | 24 820 ± 750 | 29 587 ± 863 | СОАН 4535 |
| 9030 ± 80 | 10 127 ± 125 | LU-6425 | |||||
| 9270 ± 110 | 10 464 ± 142 | LU-6424 | |||||
| 52 | Нюлсавэйто | 67°32′ | 70°10′ | 14С | 5620 ± 188 | 6395 ± 405 | ИЭРЖ_67к |
| 6081 ± 207 | 6934 ± 491 | ИЭРЖ-69с | |||||
| 7041 ± 281 | 7913 ± 490 | ИЭРЖ-69см | |||||
| 7291 ± 219 | 8108 ± 431 | ИЭРЖ-71с | |||||
| 8179 ± 231 | 9033 ± 517 | ИЭРЖ-72 | |||||
| 8182 ± 227 | 9085 ± 296 | ИЭРЖ-79 | |||||
| 53 | Шапкина-1102 | 67°28′ | 54°36′ | 14С | 7030 ± 110 | 7850 ± 103 | ГИН-9443 |
| 5920 ± 50 | 6754 ± 62 | ГИН-9443 | |||||
| 54 | Ванзеват | 64°10′ | 66°03′ | 14С | 8350 ± 300 | 9297 ± 387 | B-7064 |
| 55 | Аган-4068/2 | 62°06′ | 77°55′ | 14С | 2200 ± 150 | 2206 ± 183 | IPAE-96 |
| 9770 ± 300 | 11 264 ± 493 | IPAE-97 | |||||
| 11 400 ± 350 | 13 337 ± 363 | IPAE-98 | |||||
| 56 | Горбуновский торфяник | 57°50′ | 59°58′ | 14С | 6990 ± 40 | 6990 ± 40 | ГИН-14 125 |
| 8200 ± 40 | 9161 ± 78 | ГИН-14 128 | |||||
| 9140 ± 40 | 10 312 ± 58 | ГИН-14 083 | |||||
| 58 | Яхаядато | 67°46′ | 70°55′ | Дендр. | 3050 л. до н. э. | ||
| 59 | Яхаеларпэйто | 67°41′ | 70°58′ | Дендр. | ~3000 л. до н. э | ||
| 60 | Порсыяха-1 | 67°36′ | 70°50′ | Дендр. | 3000 л. до н. э. | ||
| 61 | Ядаяходыяха-1 | 67°35′ | 70°36′ | Дендр. | 1794 л. до н. э. | ||
| 27 | Зеленый остров | 62°29′ | 81°51′ | 14С | 10 780 ± 70 | 12768 ± 72 | Le-8972 |
| 22 100 ± 325 | 26669 ± 681 | СОАН-7550 | |||||
| 62 | Ядаяходыяха-4 | 67°33′ | 70°40′ | Дендр. | 3533 л. до н. э. | ||
| 63 | Ядаяходыяха-5 | 67°32′ | 70°40′ | Дендр. | 4639 л. до н. э. | ||
| 683 л. до н. э. | |||||||
| 64 | Ядаяходыяха-8 | 67°31′ | 70°40′ | Дендр. | 3769 л. до н. э. | ||
| 78 | Шипичное | 61°05′ | 60°33′ | 14С | 5770 ± 60 | 6573 ± 69 | СОАН 4539 |
| 79 | Черемухово-1 | 60°24′ | 60°03′ | 14С | 3210 ± 25 | 3428 ± 24 | СОАН 4798 |
| 80 | Богословский карьер | 59°46′ | 59°58′ | 14С | 4400 ± 60 | 5050 ± 142 | ГИН-84 |
| 86 | Усть-Лог | 56°57′ | 57°25′ | 14С | 2664 ± 100 | 2752 ± 135 | SPb_920 |
| Примечания. Тип даты: 14С – радиоуглеродная, дендр. – дендрохронологическая (по дате гибели дерева, которому соответствует слой). Калибровка дат осуществлялась при помощи программы CalPal Online (www.calpal-online/de). | |||||||
Интерпретация видовых списков насекомых основана на возможности отнесения обнаруженных в отложениях фрагментов к современным видам. Для представителей этого отряда характерны высокая степень таксономического разнообразия, широкий спектр экологических требований. Это обусловливает возможность детальной реконструкции как на локальном, так и на региональном уровнях и соотнесения полученных данных с этапами геологической истории, позволяет рассматривать насекомых в качестве одного из наиболее перспективных источников информации, используемого для целей реконструкции климата.
В четвертичных отложениях встречаются группы, населяющие как наземные биотопы, так и пресноводные водоемы (озера, реки, небольшие лужицы и т.д.). К первым относятся большинство жесткокрылых, обитающих в основном в наземных биогеоценозах, а также перепончатокрылые (Hymenoptera, Formicoidea). Водные насекомые относятся к нескольким отрядам, причем наиболее распространены исследования комаров-звонцов или хирономид (Diptera, Chironomidae), используемых для характеристики озерных отложений разных периодов квартера, в том числе и голоцена (Brooks, 2003; Ilyashuk, Ilyashuk, 2004; Ilyashuk et al., 2005; Gathorne-Hardy et al., 2009; Brooks et al., 2012 и др.). Кроме них рассматриваются остатки ручейников (Trichoptera), а также водные группы жесткокрылых (семейства Dytiscidae, Haliplidae, Gyrinidae, Dryopidae, Elmidae, частично Hydrophilidae и др.).
Для повышения степени достоверности реконструкций на основе энтомологического анализа необходимо привлечение данных по изучению современных энтомофаун как изучаемого региона, так и других территорий, где встречаются найденные в отложениях виды. Особое значение могут иметь результаты собственных сборов, позволяющие оценивать не только их распространение, но и характер предпочитаемых ими биотопов.
Использование данных по современному распространению насекомых дает возможность выделить виды жуков, позволяющие реконструировать широкий спектр биотопов в местах формирования отложений. В частности, существует целый комплекс видов жесткокрылых, связанных с конкретными породами деревьев: сосны (Callirus abietis (L.), Pissodes pini (L.), P. piniphilus (Hbst.), Rhyncolus ater (L.) и др.), ели (Phoelotribus spinulosus (Rey.) и др.), лиственницы (Hylobius excavatus (Laich.) и др.), а также лиственных – березы (Platycerus caraboides (L.), Sinodendron cylindricum (L.), Scolytus ratzeburgi Jans., Deporaus betulae (L.) и др.). Т. е., среди жуков существует достаточное число видов, позволяющих реконструировать определенные типы лесов. Но это возможно только в тех случаях, если бы в захоронение попадали именно эти виды без так называемого “засорения” интразональными формами, связанными с приводными и заболоченными биотопами.
Совокупность локальных тафономических факторов в конкретных местах захоронения, в том числе степень развитости околоводной растительности, может существенным образом искажать видовой состав реально существовавших фаун за счет привнесения большого числа околоводных и водных форм с участков, примыкающих к местам захоронения (Назаров, 1984). Это характерно для условий именно гумидного климата, где околоводная растительность развита достаточно сильно. В то же время в тундрах приводная растительность, как правило, развита слабо, что обусловливает бóльшую вероятность попадания в захоронения насекомых, обитавших на плакорах (Назаров, 1984). В результате этого состав энтомокомплексов будет отражать более сильное развитие ксероморфных формаций, а состав соответствующих тафоценозов будет реально приближен к реально существовавшим зональным группировкам насекомых.
Таким образом, при характеристике энтомологического материала нужно принимать во внимание специфику формирования отложений разных типов, положение местонахождений в системе современной природной зональности. Кроме того, для уточнения индикаторной ценности тех или иных таксонов необходимо привлечение как можно большего числа данных по современным жукам как с тех территорий, где проводятся палеонтологические исследования, так и с других участков современных ареалов представленных в слоях видов. Причем это касается не только специфики современного распространения отдельных групп насекомых, но и состава сообществ насекомых в целом, населяющих как зональные, так и интра- и азональные ландшафты.
РЕЗУЛЬТАТЫ
В настоящей работе рассматривается энтомологический материал из 39 голоценовых местонахождений с территории Урала, Предуралья, центральной и северной частей Западной Сибири (рис. 1). Использованы главным образом оригинальные данные, полученные в ходе полевых исследований с 1988 по 2013 годы. Кроме того, привлечены литературные сведения по ряду точек с северной части Западно-Сибирской равнины (Киселев и др., 1982; Ерохин, 1988; Киселев, 1988; Назаров, 1989). Часть проб была отобрана непосредственно автором, кроме того, использован материал, представленный В.В. Стефановским (ООО “Уралгеология”), Н.К. Пановой и Т.Г. Антипиной (Институт леса УрО РАН, г. Екатеринбург), А.В. Бородиным, Е.П. Извариным, С.В. Зыковым, С.С. Трофимовой и Н.Г. Ерохиным (ИЭРиЖ УрО РАН). С территории Большеземельской тундры пробы на энтомологический и карпологический анализы были собраны Н.Г. Смирновым (ИЭРиЖ УрО РАН).
Рис. 1.
Местонахождения голоценовых насекомых на Урале и в центральной и северной частях Западно-Сибирской равнины: 1 – Юрибей (Киселев, Друк, Криволуцкий, 1982), 2 – Нюлсавэйто, 3 – Шапкина-1102, 4 – Море-Ю (обн14), 5 – Яхаядато, 6 – Яхаеларпэйто, 7 – Порсыяха-1, 8 – Ядаяходыяха-1, 9 – Ядаяходыяха-4, 10 – Ядаяходыяха-5, 11 – Ядаяходыяха-8, 12 – Лесмиеган-9132, 13 – Лесмиеган-9142/2, 14 – Лесмиеган-9146/2, 15 – Лесмиеган-9168/1, 16 – Ванзеват, 17 – Етыпур, 18 – Аган-3081, 19 – Аган-4068/2, 20 – Аган-1065, 21 – Ватьеган, 22 – Эллееганский перекат, 23 – Черничный ручей, 24 – Аган-1082/2, 25 – Кульеган-2247 (верх), 26 – Шипичное, 27 – Богословский карьер, 28 – Черемухово, 29 – Чусовая-1, 30 – Чусовая-2, 31 – Чусовая-3, 32 – Чусовая-4, 33 – Озеро Перевальное, 34 – Горбуновский торфяник, 35 – Ноябрьск, 36 – Аганский увал-2, 37 – Шигирский торфяник, 38 – Тэва (Ерохин, 1988), 39 – Усть-Лог.
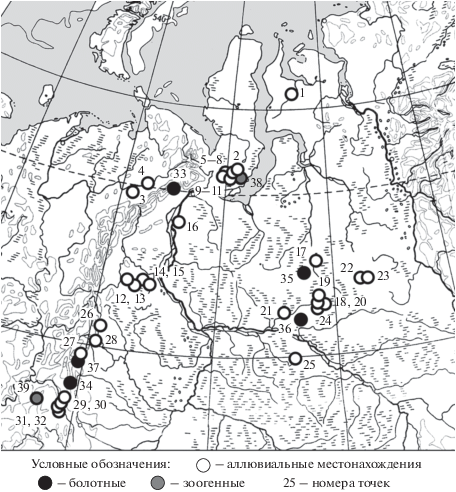
Большинство местонахождений (31) имеет аллювиальный и озерно-аллювиальный генезис, 6 точек – болотный (торфяники) и 2 точки – зоогенный (точки Усть-Лог и Тэва). Для части точек получены абсолютные даты на основе радиоуглеродного анализа, а также по датам гибели деревьев, ассоциированных с соответствующими слоями (табл. 1).
Из самых северных районов Западно-Сибирской равнины, соотносимых с подзоной современных типичных тундр, описаны фауны жуков, аналогичные современным. В частности, с севера Гыданского п-ова С.В. Киселевым была представлена фауна арктического типа, полностью соответствующая современным (рис. 1, т. 1). На это, в частности, указывают находки таких видов как Pterostichus sublaevis (J. Sahlb.), P. costatus (Men.), P. vermiculosus (Men.), Stereocerus rubripes Dej, Amara alpina (Payk.), Chrysolina cavigera Sahlb. и других, обитающих здесь и в настоящее время (Киселев и др., 1982).
В голоценовых слоях местонахождения Нюлсавэйто (рис. 1, т. 2), имеющих серию радиоуглеродных дат от 6395 ± 405 (ИЭРЖ-67к), 7291 ± 219 (ИЭРЖ-71с) и 9085 ± 296 (ИЭРЖ-79) (табл. 1), представлены комплексы насекомых северобореального типа (Панова и др., 2010). В них доминируют полизональные виды, тогда как арктобореальные насекомые единичны (Hypnoidus arcticus Gandeze). Найденные в торфяных отложениях виды муравьев подсемейств Formicidae и Myrmicidae могут быть обитателями как лесотундры и кустарниковой (южной) тундры, так и таежной зоны. Принципиально иным является видовой состав насекомых из нижнего слоя, подстилающего торфяную толщу (не имеющего радиоуглеродной даты), в нем доминируют арктобореальные виды жуков (Diacheila polita (Fa1d.), Pterostichus cf. tundrae Tsch., Pt. ventricosus Esch., Pt. cf. pinguedineus Esch.). Доминирование криофильных насекомых и отсутствие бореальных групп позволяет предполагать, что отложения формировались в условиях более холодного климата по сравнению с лежащими выше торфяными. На основе полученных данных можно предположить, что смена климатической обстановки произошла примерно на рубеже 10–9 тыс. л. н., поскольку образец 6 взят из отложений, расположенных ниже слоя, датированного по радиоуглероду 9033 ± 517 (ИЭРЖ-72) и 9085 ± 296 л. н. (ИЭРЖ-79) (Панова и др., 2010; Зиновьев, 2016) (табл. 1).
В составе энтомокомплексов местонахождения Шапкина-1102 (Большеземельская тундра, левый берег в 7 км к юго-западу от устья р. Вэснию; рис. 1, т. 3), отнесенных к раннему и среднему голоцену на основании калиброванных радиоуглеродных дат в 7850 ± 103 (ГИН-9443) и 6754 ± 62 л. н. (ГИН-9443) (Смирнов и др., 1999), отмечено доминирование бореальных и полизональных видов насекомых с транспалеарктическими типами ареалов (Clivina fossor (L.), Platynus mannerheimi (Dej.), Acidota crenata (F.), и др.). Здесь же присутствует ряд арктобореальных форм (Bembidion hastii C.R.Sahlb., Pterostichus brevicornis (Kby.), Р. (Cryobius) sp. и др.). По соотношению таксонов надвидового уровня и ландшафтных групп энтомокомплексы данного местонахождения целесообразнее всего отнести к северобореальному лесному типу фаун, а среднегодовые температуры выше современных на 2°С. Таким образом, рассматриваемые энтомокомплексы отражают существование на территории современной Большеземельской тундры в раннем голоцене лесных сообществ северотаежного типа, распространенных в настоящее время южнее Северного Полярного круга (Смирнов и др., 1999). Относительно высокое содержание в пробах жуков семейства Elmidae указывает на то, что отложения формировались в зоне влияния проточного водоема, поскольку они обитают исключительно в проточной воде.
В составе среднеголоценового энтомокомплекса местонахождения Море-Ю, (Большеземельская тундра, правый берег р. Вэснию, у ее впадения в р. Море-Ю; рис. 1, т. 4) отмечено полное отсутствие как арктических, так и ксилобионтных бореальных видов. Энтомологические данные отражают существование участка заболоченной поймы реки в пределах северной части современной бореальной зоны (Зиновьев, 2016).
Условия более теплого по сравнению с современным климата реконструируются и по энтомологическим данным для бассейна р. Яда-Яходыяха (Южный Ямал, подзона южных или кустарничковых тундр; в пойме развиты лиственничные леса). Отсюда было описано 7 точек (рис. 1, точки 5–11), эти точки приурочены в основном к отложениям, ассоциированным с остатками полуископаемой древесины (Зиновьев и др., 2001). Выявленные комплексы жуков относятся в основном к субарктическому лесотундровому и бореальному лесному типам по классификации фаун, предложенной Зиновьевым (Zinovyev, 2006). В них отмечен целый ряд ксилобионтных форм жуков, в том числе связанных с елью (короеды Hylugrops palliatus Gyll., Phoelotribus spinulosus Rey.) и лиственницей (Orthotomicus suturalis Gyll.). Максимальное содержание ксилобионтных видов отмечается в местонахождении Ядаяходыяха-8 (рис. 1, т. 11), возраст которой определен по дате погибшего дерева как 3769 г. до н. э. (табл. 1). Таким образом, комплексы жуков из этих местонахождений показывают продвижение к северу лесных видов жуков, что обусловлено распространением здесь древесной растительности, которая позже сместилась к югу (Зиновьев и др., 2001).
Для местонахождений, расположенных в пределах таежной зоны, описаны фауны жуков, аналогичные современным на этих же территориях.
Так, в среднем течении р. Лесмиеган (приток р. Сыни) насекомые найдены в четырех точках, отнесенных к голоцену – Лесмиеган-9132, Лесмиеган-9142/2, Лесмиеган-9146/2 и Лесмиеган-9168/1 (рис. 1, т. 12–15). Их возраст определен на основе принадлежности изученных местонахождений к геоморфологическому уровню пойменной террасы; радиоуглеродные даты для этих точек не были получены. Основу представленных в них энтомокомплексов составляют бореальные лесные (Pterostichus mannerheimi (Dej.), Р. diligens (Sturm), Notiophilus fasciatus Maekl., Amara brunnea (Gyll.) и др.), арктобореальные (Agonum bicolor (Dej.), Pterostichus (Cryoblus) sp.), а также полизональные гигрофильные виды (Bembldion obliquum (Ol.), Patrobus septentrionis, Р. assimilis, Agonum fuliginosum (Pz.), Cercyon spp., Olophrum spp., Acidota crenata (Mnnh.) и др.). Это указывает на определенное сходство сообществ, существовавших на территории долины р. Лесмиеган в среднем и позднем голоцене, с современными экосистемами этого района, поскольку ареалы всех найденных видов охватывают территорию современной северотаежной зоны, куда и входит эта территория. Выявленные же различия между группировками насекомых можно объяснить особенностями локальных стаций, соответствующих участкам формирования исследуемых слоев (Зиновьев, 2016). Аналогичные данные получены и для раннеголоценового местонахождения Ванзеват, расположенного в Нижнем Приобье (рис. 1, т. 16) и для т. Етыпур в среднем течении одноименной реки (рис. 1, т. 17).
На территории Среднего Приобья голоценовые местонахождения приурочены к долинам рек Аган и ее притоков, а также Глубокий Сабун (рис. 1, т. 18–26) (Бородин и др., 1994; Зиновьев, 2016 и др.). В составе тафоценозов голоценовых отложений средней и юга северной тайги (местонахождения долин рек Глубокий Сабун, Аган, Ватьеган и Кульеган) присутствуют преимущественно бореальные и полизональные виды жуков, характерные и для современных лесных, пойменных и болотных биотопов этих же территорий. Маркерами лесных сообществ являются жужелицы Notiophilus fasciatus Makl., Calathus micropterus Duft., C. erratus C.R.Sahlb., долгоносики Rhyncolus ater (L.), Anthonomus varians (Payk.), Pissodes spp. и др. В целом, представленные комплексы также соответствуют современным колеоптерофаунам с этих же территорий (Зиновьев, Нестерков, 2003; Зиновьев, 2007 и др.). Найденные в них фрагменты насекомых отнесены к видам, населяющим изучаемую территорию и в настоящее время. Карпологические и палинологические данные, полученные для этих точек, также указывают на сходство растительных ассоциаций с современными флорами этой территории (Бородин и др., 1994). При этом согласно ботаническим данным, на территории бассейна р. Аган в среднем голоцене были распространены березовые леса с сосной и елью, а в верхнем – сосново-березовые с участием ели (Архипов, Волкова, 1994). К сожалению, энтомологические данные не позволяют делать подобного рода заключения, поскольку в составе энтомокомплексов преобладают именно интразональные виды жуков, связанные с приводными и заболоченными биотопами. Т. е., в отличие от растительности (реконструированной по результатам спорово-пыльцевого анализа), группировки насекомых в этот период не претерпевали существенных изменений и были во многом идентичны рецентным энтомокомплексам, населяющим данный район. Их характерной особенностью является полное отсутствие арктических видов насекомых, доминирование таежных и интразональных форм, а также находки отдельных аркто-бореальных форм, таких как жужелица Diacheila polita Fald., распространенных ныне в тундpовой зоне и в северотаежных лесах (Бородин и др., 1994). Динамика энтомокомплексов, которую можно сопоставить с плейстоцен-голоценовым переходом, описана лишь на примере местонахождения Аган-4068/2 (рис. 1, т. 19) в верховьях одноименной реки (Зиновьев, 2005), где арктические и субарктические виды жуков были обнаружены в нижних слоях, датированных по радиоуглероду в 11 264 ± 493 (IPAE-97) и 13 337 ± 363 л. н. (IPAE-98) (табл. 1).
На Северном Урале известны три точки, отнесенные к среднему (Шипичное, Богословский угольный карьер; рис. 1, т. 26, 27) и позднему (Черемухово 1; рис. 1, т. 28) голоцену. Так, в районе Богословского угольного карьера в окрестностях г. Карпинска Свердловской обл. были исследованы аллювиальные отложения р. Турьи (Трофимова, Зиновьев, 1999). Для верхней (пойменной) фации исследуемого разреза по растительному детриту получена калиброванная радиоуглеродная дата в 5050±142 л. н. (ГИН-84). Отсюда описан комплекс жуков, представленный полизональными и бореальными (Scolytus sp., Trechus secalis (Pk.), Trichapion simile Kby) видами жуков. Эта фауна соответстввует современным энтомокомплексам с этих же территорий (Трофимова, Зиновьев, 1999).
Несколько иные выводы получены для точки Шипичное (бассейн р. Лозьва вблизи одноименного поселка в Ивдельском р-не Свердловской обл.), имеющей калиброванную радиоуглеродную дату в 6573 ± 69 л. н. (СОАН 4539; Зиновьев, Фадеев, 2002; табл. 1). Во всех трех образцах обнаружены однотипные комплексы насекомых, характеризующиеся доминированием полизональных и бореальных видов. В целом тафоценозы данного местонахождения соответствуют современным энтомокомплексам таежной зоны Северного Урала, описанным в энтомологической литературе (Козырев, 1997; Козырев и др., 2000 и др.). Видовой состав найденных насекомых отражает биотопы типа северотаежных лесов, соседствующих с поймой реки, на что, в частности, указывает находка жужелицы Oodes helopioides F. Значительный интерес представляет находка в пробе 3 данной точки (откуда и была получена радиоуглеродная дата) переднеспинок жужелицы Poecilus (Derus) sp. Представители этого подрода обычны в позднеплейстоценовых отложенях региона (Зиновьев, 2003; Зиновьев и др., 2007; Zinovyev, 2011 и др.), но отсутствуют в составе современных энтомофаун Урала и Западной Сибири. Можно предположить, что данные остатки могли быть переотложены в эти слои из более древних отложений.
В точке Черемухово 1 (старица р. Сосьвы в 6 км к югу от пос. Черемухово Свердловской обл.) обнаружены фауны жуков бореального лесного типа (Бородин и др., 2001; Струкова и др., 2006). Фрагменты насекомых взяты из слоев, один из которых имеет калиброванную дату в 3428 ± 24 л. н. (СОАН 4798; Зиновьев, 2003) (табл. 1). В них представлены виды, встречающиеся в настоящее время на этой же территории и населяющие как пойменные, так и лесные биотопы. Отмечено относительно большое число жуков-короедов (семейство Scolytidae), найденных в трех пробах этого местонахождения. Среди них выделены Carphoborus rossicus Sem. и Phloeotribus spinulosus (Rey.), связанные с елью Другой особенностью тафоценозов данного местонахождения является относительно высокая численность видов семейства Elmidae, связанных с проточными водоемами (Зиновьев, 2003). Это характерно для энтомокомплексов из отложений, формировавшихся в зоне действия предгорных и горных рек, например Лозьвы (точка Шипичное) или Шапкиной в Приуральской Субарктике (Смирнов и др., 1999).
На правом берегу р. Чусовая (1 км выше пос. Верхнемакарово, окр. г. Екатеринбурга) из четырех местонахождений насекомых (рис. 1, т. 29–32), отнесенных к голоцену (Зиновьев, 2003), также описаны идентичные современным фауны насекомых. Все четыре точки приурочены к береговым обрывам, представляющим геоморфологический уровень пойменной террасы (высота до 3 м) (Зиновьев, 2003). Насекомые, найденные в этих слоях, в настоящее время встречаются на территории Среднего Урала и Зауралья. Тем не менее, в пределах всех этих разрезов выявлена определенная смена энтомокомплексов, которую, однако, можно объяснить сукцессионными изменениями. Так, в точке Чусовая-1 из нижнего образца (глубина 1.95–2.1 м) описан энтомокомплекс, характерный для торфяного болота, в пробе с глубины 1.85–1.95 м отмечены виды, населяющие заболоченные леса, и, наконец, в образце с глубины 1.65–1.75 м найденные насекомые позволяют воссоздать биотоп типа заболоченного луга. Схожие изменения в составе энтомокомплексов указаны и для остальных разрезов Чусовая-2, Чусовая-3 и Чусовая-4, расположенных соответственно в 1.3, 1.5 и 2 км выше по течению от пос. Верхнемакарово. Из оторфованного суглинка в верхней части точки Чусовая 3 описана фауна жуков бореального лесного типа. В местонахождении Чусовая-2 отмечены виды жуков, связанные с водоемами (вертячки рода Gyrinus) и болотами (стафилиниды подсемейства Аlеосhаrinае). Лугово-болотные комплексы насекомых описаны также из нижних слоев точек Чусовая-4 и Чусовая-3.
Таким образом, в большинстве местонахождений таежной зоны описанные фауны насекомых соответствуют современным энтомокомплексам с этих же территорий. Выявленные изменения в пределах конкретных разрезов можно связать с сукцессионными процессами при формировании разных элементов пойменного ландшафта – переход от участков старицы реки к болоту или сосновому лесу. Климатические изменения, выявляемые на основе палинологических данных, не отражены в динамике комплексов насекомых.
При этом в большинстве случаев анализ видового состава насекомых из аллювиальных местонахождений голоценового возраста в пределах таежной зоны позволяет реконструировать в первую очередь набор локальных стаций непосредственно вблизи места захоронения. Это, как правило, интразональные биотопы – прибрежные участки, старицы, в ряде случаев – леса. Однако говорить о типах леса в том виде, как это можно выявить в ходе отловов современных жуков (см. выше), не приходится. Можно лишь реконструировать наличие тех или иных пород деревьев (сосна, ель, лиственница, береза), однако их соотношение, выявляемое на основе анализа палиноспектров, установить нельзя. Даже при увеличении выборок все равно будут доминировать интразональные формы, сглаживающие общий облик фаун. В ряде случаев можно выявить сукцессионные изменения – переход от участков заболоченной поймы реки к хвойному лесу, однако они далеко не всегда обусловлены климатической динамикой и отражают, скорее всего, процессы трансформации биотопов при изменениях русла. Таким образом, климатические изменения в том виде, в каком они имели место в голоцене, в большинстве случаев не могут быть выявлены на энтомологическом материале, поскольку в захоронения попадают виды, прежде всего, из околоводных биотопов, где колебания макроклимата могут быть достаточно сильно сглажены. Доминирование же интразональных насекомых в пробах не позволяет выявить даже смены типов леса, если таковые были в местах формирования энтомологического материала.
Аналогичная ситуация показана и для торфяников. Остатки насекомых в отложениях данного генезиса встречаются в них достаточно часто, однако в ходе наших исследований также не была выявлена динамика энтомокомплексов, которую можно было бы надежно сопоставить с климатическими изменениями на данных территориях в голоцене.
Это, в частности, показано для точки “озеро Перевальное” (рис. 1, т. 33), расположенной в восточной части массива Рай-Из (Панова и др., 2003). Нижние слои данного местонахождения (с глубин 140–160 и 180–190 см) имеют радиоуглеродные даты, соответственно, 9030 ± 80 (LU-6425) и 9270 ± 110 л. н. (LU-6424) (табл. 1). Из голоценового торфа описаны комплексы насекомых, аналогичные современным на этой территории. Арктические и арктобореальные виды здесь не представлены. Ксилобионтные насекомые обнаружены только в средних и нижних частях данного местонахождения, что согласуется с данными палинологического и карпологического анализов (Панова и др., 2003).
При анализе энтомологического материала из двух торфяников (Ноябрьск-1 и Аганский увал-2; рис. 1, т. 34 и 35) описаны комплексы жуков, соответствующие современным группировкам жесткокрылых с этих же территорий (Зиновьев, Рябицев, 2000). В первом местонахождении доминируют виды, связанные с переувлажненными, в том числе и заболоченными биотопами (Plateumaris sericea (L.), Agabus spp., Notaris aethiops F. и др.).
Крайне интересные (хоть и немногочисленные) выборки насекомых были получены из Горбуновского торфяника на Среднем Урале (рис. 1, т. 36), ранее описанного в литературе (Старков и др., 1979; Лаптева, 2010; Панова, 2011; Антипина и др., 2013). Энтомологический материал взят из культурных слоев II, III и IV в нижней части данного местонахождения. Максимальное число остатков извлечено из культурного слоя IV, имеющего 14С дату 9140 ± 40 л. н. (ГИН-14 083). Представленные виды имеют современное бореальное и полизональное распространение и связаны с болотами (Pterostichus aterrimus (Hbst.)), березовыми лесами (Cimbex spp., Platycerus caraboides), а также с интразональными биотопами в пределах таежной (лесной) зоны (Phratora sp., Plagiodera versicolora (L.), Agelastica alni (L.), Altica sp., Protaetia metallica (Hbst.). Представленный список отражает условия климата, аналогичного современному на данной территории, и может указывать на распространение березовых лесов с зарослями ивы. В культурном слое II (14С дата 6990 ± 40 л. н. (ГИН-14 125)) отмечены виды жужелиц, связанных с лесо-болотными (Carabus menetriesi Humm.) и заболоченными биотопами (Chlaenius sulcicollis (Payk.), Agabus sp.). Помимо них в слое обнаружены хорошо сохранившиеся фрагменты бронзовки Protaetia metallica, населяющей разнотравные луга, в том числе и в поймах рек (причем личинки этого вида населяют трухлявую древесину). Видовой состав данного местонахождения показывает наличие участков заболоченного березового леса, что соотносится с палинологическими данными, полученными ранее для этой же части разреза (Панова, 2011; Антипина и др., 2013).
Немногочисленные остатки жуков из верхней части Шигирского (Калатинского) торфяника (окр. г. Кировграда Свердловской обл.; рис. 1, т. 37) отнесены к видам, имеющим современное полизональное и бореальное распространение. Согласно результатам анализа экологических требований найденных видов, условия в период формирования отложений тождественны современным на этой территории, указывая, в том числе и на наличие древесной растительности (ели) на этом участке.
При анализе энтомологического материала часто возникает проблема низкой насыщенности торфа остатками насекомых. Это во многих случаях не позволяет давать развернутые характеристики ландшафтно-климатических условий, особенно если на это накладываются локальные микро- и мезоклиматические факторы (например, сглаживание крайних значений температур и увеличение влажности в поймах, что обусловливает широкое развитие околоводной растительности). И даже при получении богатого материала (точки Ноябрьск-1, Аганский увал-2) он не позволяет выявить какие-либо изменения в составе энтомокомплексов, которые можно было бы связать с процессами смены типов лесной растительности (Бородин и др., 1994).
Основной причиной отсутствия четко выраженных изменений в составе энтомокомплексов можно считать слабую степень динамики ландшафтно-климатических условий, отраженную в соответствующей изотопно-кислородной шкале голоцена (Хотинский, 1977).
Энтомологический материал из зоогенных захоронений, к которым можно отнести птичьи погадки, экскременты песцов и барсуков, в настоящее время исследован недостаточно. Тем не менее, именно найденные в этих типах захоронений остатки насекомых могут быть весьма ценным источником информации для палеоэкологических и палеотемпературных реконструкций. Так, анализ остатков насекомых из голоценовых барсучьих нор Русской равнины позволил сделать заключения о более теплом, по сравнению с современным, климате на основе находок ряда термофильных видов жуков, таких как Dendroxena quadrimaculata (Scop.) (Динесман, 1968).
В среднеголоценовом местонахождении Тэва на Южном Ямале (возраст определен в пределах 6000 лет; рис. 1, т. 38), описанном Ерохиным (1988), преобладают полизональные и арктобореальные виды насекомых, здесь же отмечены как арктические (Pterostichus sublaevis), так и бореальные лесные формы (Sericoda bogemanni (Gyll.), Hylobius albosparsus Boh., Camponotus herculeanus (L.)), достоверно отсутсвующие в составе современных энтомофаун Южного Ямала. Представленный энтомокомплекс отражал, судя по всему, существование хвойных (скорее, лиственничных) лесов северобореального типа (Ерохин, 1988), что соответствует данным, полученным для местонахождений с долин рек Ядаяходыяха и Порсыяха (Зиновьев и др., 2001). Присутствие арктической жужелицы P. sublaevis в составе энтомокомплекса северобореального типа было отмечено нами для отложений более раннего возраста, в частности местонахождения Белая гора, датированного стадиалом брёруп (МИС5с), т. е., около 100 тысяч лет назад (Зиновьев и др., 2016).
И, наконец, энтомокомплекс зоогенного местонахождения Усть-Лог (рис. 1, т. 39), имеющего калиброванная 14С дата 2450–3100 л. н. (табл. 1) можно характеризовать как лесостепной. В его составе отмечены степные (Harpalus cf. froelichi Sturm, Oodescelis polita (Sturm) и др.) и лесные жесткокрылые (Callirus pinastri (L.), Pterostichus oblongopunctatus (F), а также муравьи родов Formica и Camponotus. Доминирующими видами являются долгоносик Otiorhynchus velutinus Germ и божья коровка Adalia bipunctata (L.). Находка фрагментов навозника в этом местонахождении надкрылья навозника весеннего (Trypocopris vernalis (L.)) может свидетельствовать о несколько более теплом и гумидном климате в данный период (Зиновьев и др., 2014).
По всей вероятности, именно в зоогенных отложениях могут быть представлены виды, обитавшие на плакорах и в большей степени зависимые в своем распространении от динамики климата в голоцене.
Таким образом, в составе энтомокомплексов из местонахождений юга лесной зоны изучаемого региона присутствуют те же виды, что населяют эту территорию и в настоящее время. Преобладающей широтной группой в этих фаунах являются полизональные виды, а доминирующей экологической группой – интразональные насекомые, в составе которых наиболее массовыми являются околоводные насекомые. Что касается таксономического состава, то одной из самых массовых групп являются стафилины, среди которых больше всего фрагментов относится к трибе Omaliinae (в т.ч. жукам рода Olophrum). Одним из маркеров теплого климата является жужелица Trechus secalis, обычная для голоценовых отложений центральной части изучаемого региона и обычная в современных энтомокомплексах интразональных и отчасти лесных биотопов южной и юга средней тайги (Зиновьев, 2007; Бухкало и др., 2011; Ухова, Ольшванг, 2014 и др.).
При этом даже в точках, отнесенных к пребореальному (7–8 тыс. лет назад) периоду, на границе с поздним неоплейстоценом уже фиксируются находки бореальных видов жуков, связанных с лесами и болотами в пределах тех самых зон, где в настоящее время эти точки находятся. Например, в точке Горбуновский торфяник на Среднем Урале отмечен целый ряд жуков, характерных именно для средне- и южнотаежных лесов (Carabus menetriesi, Placycerus caraboides, Agelastica alni).
ОБСУЖДЕНИЕ
Сопоставление энтомологических и палеоботанических данных из голоценовых отложений региона
В ряде местонахождений образцы на энтомологический анализ были отобраны совместно с палинологическими и карпологическими пробами. Представляется целесообразным оценить то, насколько в таких случаях динамика энтомокомплексов могла соответствовать изменениям в составе спорово-пыльцевых спектров в пределах одного и того же разреза.
Было проведено сопоставление палинологических и энтомологических данных по местонахождениям Нюлсавэйто и озеро Перевальное (Панова и др., 2003, 2010), которые имеют, соответственно, аллювиальный и болотный генезис отложений.
Так, в местонахождении Нюлсавэйто было выделено восемь палинозон (ПЗ), показывающих изменения в составе лесной растительности на протяжении голоцена. Так, палинозоне 8 (ПЗ8) соответствуют спорово-пыльцевые спектры лиственнично-елово-березовой лесотундры с кустарничково-ерниковым покровом и соотносящиеся с окончанием бореального – началом атлантического периодов голоцена, в соответствии с радиоуглеродными датами (8179 ± 2231 и 7290 ± 219 л. н.). ПЗ7 – редкостойные лиственнично-березово-еловые леса, соответствующие первой половине атлантического периода. ПЗ6 – березовые леса с елью и лиственницей; ПЗ5 – таежные леса с преобладанием ели, примесью лиственницы и березы, относящиеся к климатическому оптимуму голоцена; ПЗ4 – лиственнично-елово-березовые леса, ПЗ3 – лиственнично-березово-еловые редколесья в условиях похолодания и увлажнения климата; ПЗ2-ПЗ1 – растительность лиственнично-елово-березовой лесотундры (зона 2), которая сменяется кустарниковой и травяно-кустарничково-ерниковой тундрой (зона 1), что соответствует нарастающему похолоданию суббореального и субатлантического периодов голоцена (Панова и др., 2010). Пробы на жуков соответствуют следующим палинозонам: пр. 6 взята ниже ПЗ8, пр. 5 соответствует ПЗ6, пр. 3 и 4 – ПЗ6, пр. 2 – ПЗ4-5 и пр. 1 – ПЗ2-1.
Взятые из этих же слоев жуки в целом отражают условия относительно теплого климата и распространения интразональных биотопов. Видовой состав насекомых из проб 1–5 данного местонахождения отличается от современного состояния энтомокомплексов на этих территориях с одной стороны (Зиновьев, Ольшванг, 2003; Ольшванг, 1992 и др.) и тафоценоза пробы 6 – с другой. Само по себе отсутствие индикаторных для холодного климата видов еще не может говорить о том, что их там не было, однако выборка из пробы 3, напротив, показывает обилие муравьев, которые более характерны именно для лесов.
Данные энтомологического и палинологического анализов сопоставлены также на примере торфяника “озеро Перевальное” (Панова и др., 2003). Здесь не была показана динамика энтомокомплексов в пределах разреза, хотя, согласно палинологическим данным, здесь было несколько периодов смены типов растительных ассоциаций, обусловленных соответствующими климатическими изменениями (табл. 2).
Таблица 2.
Сопоставление палинологических и энтомологических данных для местонахождения Нюлсавэйто (Панова и др., 2010)
| Глубина, см | Дата (некалиброванная) | Номер палинозоны (ПЗ) | Спорово-пыльцевые спектры | Энтомологические данные |
|---|---|---|---|---|
| 23–0 | Нет даты | ПЗ2-1 | Лиственнично-елово- березовая лесотундра, сменяющаяся кустарниковой и травяно-кустарничково- ерниковой тундрой | Нет данных |
| 40–23 | 5620 ± 188 | ПЗ3 | Лиственнично- березово-еловые редколесья | Малое число остатков, отнесенных к интразональным таксонам жуков. Находки фрагментов муравья Lasius niger, распространение которого можно сопоставить с бореальной зоной |
| 60–40 | 6081 ± 207 | ПЗ4 | Лиственнично- елово-березовые леса | Малое число остатков, в том числе жужелиц Agonum и плавунцов Hydroporus |
| 80–60 | ПЗ5 | Таежные леса с преобладанием ели, примесью лиственницы и березы | ||
| 140–80 | ПЗ6 | Березовые леса с елью и лиственницей | Фауна насекомых бореального интразонального типа с доминированием фрагментов муравья Formica gagathoides | |
| 225–140 | Нет даты | ПЗ7 | Редкостойные лиственнично-березово-еловые леса | Нет данных |
| 250–225 | 7290 ± 219 8179 ± 2231 | ПЗ8 | Лиственнично-елово- березовые лесотундры с кустарничково- ерниковым покровом | Энтомокомплекс бореального типа, в том числе с участием муравьев Formica gagathoides |
| 350 | Нет даты | Нет данных | Фауна насекомых субарктического типа | |
В частности, на энтомологическом материале не выявлены те небольшие похолодания, которые отмечены на примере палинокомплексов нижней (190–250 см) и верхней (35–0 см) частей разреза. Палинокомплексы нижней части торфяника (190–250 см) указывают на тундровую травяно-кустарниковую растительность в сочетании с лиственнично-березовым редколесьем, тогда как палинокомплекс верхней части указывает на условия современной лесотундры, что никак не отражено на энтомологическом материале. В целом выявленная структура энтомокомплексов характерна для верховых болот таежной зоны Западно-Сибирской равнины. Этот факт, а также отсутствие в разрезе арктических и целого ряда аркто-альпийских видов, обитающих на Полярном Урале в настоящее время, свидетельствуют о более теплых климатических условиях времени формирования основной толщи торфа по сравнению с современными.
Аналогичные выводы получены на карпологическом материале. Выделенный комплекс макрофоссилий основной толщи торфа характеризует растительность заболоченного березового леса с елью и лиственницей и более теплый и мягкий климат по сравнению с современным (Панова и др., 2003).
Таким образом, климатические изменения, происходившие в голоцене и выявленные на палеоботаническом материале, далеко не всегда могут быть показаны на основе анализа энтомологических данных.
Сравнение с энтомологическими данными из голоценовых местонахождений Европы и Северо-Восточной Сибири
Для Западной и Восточной Европы в этот период также реконструировано окончательное формирование энтомокомплексов современного облика. Изучению голоценовых отложений Европы посвящено большое число работ (Buckland, 1979; Назаров, 1984, 1989; Atkinson et al., 1987; Lemdahl, Gustavsson, 1997; Lemdahl, 2000; Buckland et al., 2004; Whitehouse, 2004, 2006; Ponel et al., 2007 и др.). Комплексы жуков из голоценовых отложений Европы сопоставляются с современными на тех же территориях, указывая на принципиальное сходство природных условий с их современным состоянием.
В работе Назарова (1984) по голоценовым местонахождениям Беларуси также описаны фауны жуков, соответствующие современным энтомокомплексам на данной территории и свидетельствующие о сходном с современным климате. В частности, к среднему голоцену отнесена точка Слобода Двинская, откуда описана фауна жуков, которая представлена исключительно лесными (Callirus abietis (L.)) и полизональными (Clivina fossor (L.), Hydrobius fuscipes (L.), Lathrobium fovulum Steph., Gonioctena affinis Gyll. и др.) видами. Это позволяет реконструировать биотопы типа речной протоки, поросшей мезофитной растительностью и ивняком. В этой же работе отмечается, что видовой состав голоценовых насекомых определяется в первую очередь спецификой локальных стаций, непосредственно граничащих с местом захоронения.
Это утверждение соответствует нашим данным по динамике комплексов жуков с территории таежной зоны (центр Западно-Сибирской равнины, Северный и Средний Урал). Судя по видовому составу видов, представленных в местонахождениях аллювиального и болотного генезиса, можно с уверенностью говорить, что захоронение в большинстве случаев происходило в интразональных ландшафтах (мезоландшафтах или фациях), таких как поймы рек или болота. В них макроклиматические факторы могли быть существенным образом сглажены по сравнению с плакорными участками. В то же время в зоогенных отложениях изменения в составе комплексов жуков, определяемые климатическими флуктуациями, могут быть выявлены. На это, в частности, могут указывать находки отдельных термофильных видов жуков, не встречающихся в настоящее время на этой территории. Например, в местонахождении Усть-Лог, расположенном в пределах Красноуфимской степи, отмечена находка неморального навозника Trypocopris vernalis, достоверно отсутствующего на этой территории в настоящее время (Зиновьев и др., 2014). В голоценовых экскрементах барсука с территории Московской обл. Медведевым (1976) был описан целый комплекс видов, характерных именно для современных энтомофаун региона, а также несколько таких термофильных жуков как жужелицы Carabus coriaceus L., C. hortensis L., мертвоед Dendroxena quadrimaculata и др.
Для территории Северо-Восточной Сибири (Sher, Kuzmina, 2007) описаны голоценовые фауны, принципиально отличающиеся от энтомокомплексов из более ранних местонахождений данного региона. Анализ фаун из ряда голоценовых точек показал, в том числе, и возможности перезахоронения материала в отложениях этого возраста из более ранних слоев. Это, в частности, отмечено для долгоносиков родов Coniocleonus и Stephanocleonus, обладающих прочными хитиновыми покровами. В самих же голоценовых отложениях региона отмечаются те же виды, что представлены в современных фаунах в настоящее время, в том числе и связанные с древесной растительностью. Резкие отличия голоценовых фаун от позднеплейстоценовых отмечены в местонахождении Яна-RHS (Янская стоянка), где в пробе, датированной 9790 ± 110 л. н. (ЛЕ-8915), исчезает большинство криофильных видов жуков, обычных в нижних пробах, зато появляются обитатели заболоченных биотопов (стафилиниды рода Olophrum), а также короед Phoelotribus spinulosus (Rey), обычный на ели, но, вероятно, обитавший и на лиственнице. При этом массовый в нижних образцах пилюльщик Morychus viridis составляет не более 0.8% от общего числа особей (Зиновьев и др., 2015).
В работах Киселева (1981; Kiselev, Nazarov, 2009) описано несколько точек, отнесенных к голоцену (Дуванный яр, Айон, Крестовка). Так, для местонахождения Айон показана динамика комплексов жуков в пределах голоцена. В пробе, соотнесенной с бореальным периодом голоцена (некалиброванная дата 8470 ± 140 л. н. (МГУ-601)), представлены арктические, арктобореальные, суббореальные степные (Coniocleonus ferrugineus Fahr., Stephanocleonus eruditus Fast., S. fossulatus F.-W., S. foveifrons Chevr.) и бореальные (Bromius obscurus (L.), Altica spp.) виды жуков. Здесь же обнаружены остатки муравьев рода Formica. Это позволило Киселеву предположить, что данный слой формировался в условиях, сложившихся на границе тундровой и лесной зон. Однако в пробах, которые ассоциированы с атлантическим периодом (некалиброванные даты 6980 ± 110 л. н. (МГУ-956) и 5680 ± 90 л. н. (МГУ-595)) доминируют арктические и арктобореальные виды жуков, что позволяет определять облик данного комплекса как “тундровый” (т. е., принадлежащий к арктическому типу фаун). И, наконец, в слое с датой в 1290 ± 200 л. н. (МГУ-583) найдена фауна жуков, которая сопоставлена с современным населением “кустарничково-травянистых” (южных) тундр.
ВЫВОДЫ
1. Голоценовые изменения в составе растительных сообществ таежной зоны Западно-Сибирской равнины и Урала, показанные на палинологическом материале, практически не выявлены на энтомологическом материале из аллювиальных местонахождений и торфяников.
2. Динамика энтомофаун, связанная со сменой ландшафтно-климатических условий, наблюдается только на границах современных природных зон, таких как тундровая и таежная. В климатический оптимум голоцена именно на таких пограничных территориях отмечено продвижение к северу целого ряда видов, связанных с лесами, в том числе и с древесной растительностью.
3. В пределах таежной зоны комплексы жуков из исследованных нами голоценовых отложений аллювиального и болотного генезиса идентичны современным фаунам соответствующих природных зон. Основные изменения в пределах конкретных разрезов могут объясняться сукцессионными процессами, связанными с динамикой мезоландшафтов в пределах смены русел рек, формированию стариц, последующему облесению.
4. Динамика ареалов отдельных видов жуков, которую можно связать с климатическими изменениями, пока выявлена лишь на примере зоогенных захоронений, поскольку в них с большей долей вероятности попадают виды, обитавшие на плакорах, по сравнению с отложениями аллювиального и болотного генезисов.
5. Голоценовые комплексы жуков позволяют реконструировать в первую очередь локальные условия осадконакопления, в том числе типы водоемов (стоячие или проточные), набор биотопов на участках вблизи мест захоронения, дают возможность дополнить характеристики растительного покрова, в том числе показать наличие отдельных элементов древесной растительности.
Список литературы
Антипина Т.Г., Панова Н.К., Чаиркина Н.М., 2013. Динамика природной среды в голоцене по данным комплексного анализа VI разреза Горбуновского торфяника // Известия Коми научного центра УрО РАН. Вып. 4 (16). Сыктывкар. С. 89–97.
Архипов С.А., Волкова B.C., 1994. Геологическая история, ландшафты и климаты плейстоцена Западной Сибири. Новосибирск: ИГиГ СО РАН. 106 с
Бачура О.П., Струкова Т.В., 2002. Остатки млекопитающих из местонахождения Черемухово-1: (раскоп 4) // Фауна Урала в плейстоцене и голоцене: сборник научных трудов. Екатеринбург. С. 37–55.
Бляхарчук Т.А., 2000. История растительности юго-востока Западной Сибири в голоцене по данным ботанического и спорово-пыльцевого анализа торфяных отложений // Сибирский экологический журнал. Вып. 5. С. 659–668.
Бородин А.В., Зиновьев Е.В., Быкова Г.В., Корона О.М., 1994. Материалы к характеристике наземных экосистем бассейна реки Аган, Аганского и Сибирских увалов в позднечетвертичное время. – М. Деп. в ВИНИТИ 11.01.1994, № 83-В94. 198 с.
Бородин А.В., Струкова Т.В., Трофимова С.С., 2001. Историко-экологические исследования на Северном Урале // Уральский север в панораме тысячелетий. Материалы историко-краеведческой конференции. Североуральск. С. 44–52.
Бухкало С.П., Галич Д.Е., Сергеева Е.В., Алемасова Н.В, 2011. Конспект фауны жуков южной тайги Западной Сибири (в бассейне Нижнего Иртыша). М.: Товарищество научных изданий КМК. 267 с.
Волкова В.С., Бахарева В.А., Левина Т.П., 1989. Растительность и климат голоцена Западной Сибири // Палеоклиматы позднеледниковья и голоцена. М.: Наука. С. 90–95.
Голубева Ю.В., 2008. Климат и растительность голоцена на территории республики Коми Институт геологии Коми научного центр // Литосфера. Вып. 2. С. 124–132.
Динесман Л.Г., 1968. Изучение истории биогеоценозов по норам животных. М.: Наука. 100 с.
Динесман Л.Г., 1976. Голоценовая история биогеоценозов Русской равнины в позднем антропогене // История биогеоценозов СССР в голоцене. М.: Наука. С. 122–132.
Динесман Л.Г., Нейштадт М.И., Флеров К.К., 1979. Изучение голоценовой истории биогеоценозов в связи с проблемами биосферы // Общие методы изучения истории современных экосистем. М.: Наука. С. 7–13.
Ерохин Н.Г., 1988. Находки остатков жесткокрылых в позднечетвертичных отложениях Южного Ямала // Современное состояние и история животного мира Западно-Сибирской низменности. Свердловск: изд-во УрО АН СССР. С. 123–126.
Зиновьев Е.В., Рябицев А.В., 2000. К фауне жесткокрылых Сибирских увалов (Западная Сибирь) // Материалы к познанию фауны и флоры Ямало-Ненецкого автономного округа. Научный вестник. Вып. 4 (II). Салехард. С. 51–55.
Зиновьев Е.В., Гилев А.В., Хантемиров Р.М., 2001. Изменения энтомофаун Южного Ямала в связи с динамикой северной границы леса в голоцене // Энтомологическое обозрение. Т. 80. Вып. 4. С. 843–851.
Зиновьев Е.В., Фадеев Ф.А., 2002. Реконструкция условий формирования голоценовых отложений местонахождения Лозьва-1 (Северный Урал) на основе энтомологических данных // Плейстоценовые и голоценовые фауны Урала. Вып. 2. Челябинск: Рифей. С. 24–35.
Зиновьев Е.В., 2003. Обзор местонахождений четвертичных насекомых Урала и Зауралья // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского Университета. С. 86–97.
Зиновьев Е.В., Нестерков А.В., 2003. Видовой состав жесткокрылых (Insecta: Coleoptera) Заповедно-Природного Парка “Сибирские Увалы” // Экологические исследования восточной части Сибирских Увалов: Сборник научных трудов ЗПП “Сибирские Увалы”. Вып. 2. Нижневартовск: Изд-во “Приобье”. С. 83–118.
Зиновьев Е.В., Ольшванг В.Н., 2003. Жуки севера Западно-Сибирской равнины, Приполярного и Полярного Урала // Биологические ресурсы Полярного Урала. Научный вестник. Вып. 3 (II). Салехард. С. 37–60.
Зиновьев Е.В. 2005. Материалы к характеристике раннеголоценовых энтомокомплексов Среднего Приобья // Евразиатский энтомологический журнал. Т. 4. Вып. 4. С. 283–292.
Зиновьев Е.В., 2007. Локальные фауны жужелиц (Coleoptera: Trachypachidae, Carabidae) Среднего Приобья и прилегающих территорий // Биологические ресурсы и природопользование. Вып. 10. Сургут: Изд-во Дефис. С. 134–148.
Зиновьев Е.В., Корона О.М., Стефановский В.В., 2007. Реконструкция условий формирования позднеплейстоценовых отложений местонахождения Андрюшино (низовья реки Тавды) по энтомологическим и карпологическим данным // Уральский геологический журнал. Т. 56. Вып. 2. С. 27–43.
Зиновьев Е.В., Изварин Е.П., Иванов А.В., Улитко А.И., 2014. Находка остатков навозника Trypocopris vernalis (Linnaeus, 1758) (Coleoptera, Geotrupidae) в позднеголоценовых орнитогенных захороненниях на Среднем Урале (Кунгурско-Красноуфимская лесостепь) // Евразиатский энтомологический журнал. Т. 13. Вып. 1. С. 11–14.
Зиновьев Е.В., Павлова Е.Ю., Питулько В.В., 2015. Граница плейстоцена и голоцена западной части Яно-Индигирской низменности по энтомологическим данным (на примере Янской стоянки) // Фундаментальные проблемы квартера, итоги изучения и основные направления дальнейших исследований: Материалы IX Всероссийского совещания по изучению четвертичного периода (г. Иркутск, 15–20 сентября 2015 г.). Иркутск. С. 170–173.
Зиновьев Е.В., 2016. Обзор местонахождений позднеплейстоценовых и голоценовых насекомых Нижнего Приобья и прилегающих к нему территорий // Вестник НВГУ. Вып. 2. С. 23–35.
Зиновьев Е.В., Бородин А.В., Трофимова С.С., Шейнкман В.С., Русаков А.В. и др., 2016. Позднеплейстоценовые энтомокомплексы разреза Белая Гора (Западная Сибирь, р. Вах) и их экологическая характеристика // Евразиатский энтомологический журнал. Т. 15. Вып. 5. С. 483–498.
Киселев С.В., 1987. Отбор образцов на палеоэнтомологический анализ // Комплексные биостратиграфические исследования: Учебное пособие. М.: Изд-во МГУ. С. 21–26.
Киселев С.В., 1988. Плейстоценовые и голоценовые жесткокрылые Западной Сибири) // Современное состояние и история животного мира Западно-Сибирской низменности. Свердловск: изд-во УрО АН СССР. С 97–118.
Киселев С.В., Друк А.Я., Криволуцкий Д.А., 1982. О фауне жуков и панцирных клещей из захоронений мамонтов // Юрибейский мамонт. М.: Наука. С. 44–53.
Козырев, А.В. 1997. Итоги изучения жужелиц (Coleoptera, Carabidae) Урала и сопредельных территорий // Успехи энтомологии на Урале. Сборник научных трудов. – Екатеринбург: Аэрокосмоэкология. С. 44–50.
Козырев А.В. Козьминых В.О., Есюнин С.Л., 2000. Состав локальных фаун жужелиц (Coleoptera, Carabidae) Урала и Приуралья // Вестник Пермского университета. Биология. Вып. 2. С. 165–215.
Косинцев П.А., 2009. Костные остатки млекопитающих из археологического памятника Усть-Полуй (раскопки 1993-1995 гг.) // Археозоологические исследования на Урале и в Сибири. Екатеринбург–Челябинск: Рифей. С. 127–137.
Лаптева Е.Г., 2010. Палинологические исследования на VI разрезе Горбуновского торфяника в 2007 году // Древности Горбуновского торфяника. Охранные археологические исследования на Среднем Урале: сборник научных статей. Вып. 6. Екатеринбург: Банк культурной информации. С. 157–163.
Медведев Л.Н., 1976. О составе энтомокомплексов из голоценовых копролитов барсука в Подмосковье // История биогеоценозов СССР в голоцене. М.: Наука, С. 183–193.
Назаров В.И., 1984. Реконструкция ландшафтов Белоруссии по палеоэнтомологическим данным (антропоген) // Труды Палеонтологического института АН СССР. Т. 205. 96 с.
Назаров В.И., 1989. Климат голоцена некоторых районов СССР (по данным энтомологии) // Палеоклиматы позднеледниковья и голоцена. М.: Наука. С. 76–79.
Ольшванг В.Н., 1992. Структура и динамика населения насекомых Южного Ямала. Екатеринбург: Изд-во УрО РАН. 104 с.
Панова Н.К., 1996. История развития лесной растительности на Урале в голоцене // Лесообразовательный процесс на Урале. Труды Института леса УрО РАН. Екатеринбург. С. 26–49.
Панова Н.К., 2011. Динамика природной среды в голоцене по данным палинологического анализа стоянки Береговая 2 на Горбуновском торфянике (Средний Урал) // Экология древних и традиционных обществ. Тюмень: Изд-во ИПОС СО РАН. Вып. 4. С. 62–64.
Панова Н.К., Трофимова С.С., Антипина Т.Г., Зиновьев Е.В., Гилев А.В., Ерохин Н.Г., 2010. Динамика растительности и экологических условий в голоцене на Южном Ямале (по данным комплексного анализа отложений реликтового торфяника) // Экология. № 1. С. 22–30.
Панова Н.К., Янковска В., Корона О.М., Зиновьев Е.В., 2003. О динамике растительности и экологических условий на Полярном Урале в голоцене // Экология. № 4. С. 248-260.
Смирнов Н.Г., 1993. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: Наука. 62 с.
Смирнов Н.Г., 1996. Разнообразие мелких млекопитающих Северного Урала в позднем плейстоцене и голоцене // Материалы и исследования по истории современной фауны Урала. Екатеринбург: изд-во “Екатеринбург”. С. 39–83.
Смирнов Н.Г., Андреичева Л.Н., Корона О.М., Зиновьев Е.В., Головачев И.Б., и др., 1999. Материалы к характеристике биоты Приуральской Субарктики в голоценовом оптимуме // Биота Приуральской Субарктики в позднем плейстоцене и голоцене. Екатеринбург: Изд-во “Екатеринбург”. С. 23–60.
Старков В.Ф., Хотинский Н.А., Алексашенко Н.А., Калинина И.В., 1979. Раскопки на Горбуновском торфянике // Археологические открытия 1978 года. М: Наука. С. 200.
Струкова Т.В., Бачура О.П., Бородин А.В., Стефановский В.В., 2006. Первые находки фауны млекопитающих в аллювиально-спелеогенных образованиях позднего неоплейстоцена и голцена Северного Урала (Черемухово-1) // Стратиграфия. Геологическая корреляция. Т. 14. Вып.1. С. 98–108.
Трофимова С.С., Зиновьев Е.В., 1999. Находки остатков насекомых и макроостатков растений из голоценовых аллювиальных местонахождений Северного Урала // Развитие идей академика С.С. Шварца в современной экологии. Материалы конференции. Екатеринбург: Изд-во “Екатеринбург”. С. 195–199.
Ухова Н.Л., Ольшванг В.Н., 2014. Беспозвоночные животные Висимского заповедника. Аннотированный список видов. Екатеринбург: Изд-во Раритет. 284 с.
Фадеева Т.В., 2002. Ископаемая микротериофауна многослойного местонахождения “Пещера Дыроватый камень на р. Вишере” // Фауна Урала в плейстоцене и голоцене. Екатеринбург: Изд-во Университет. С. 85–110.
Хазин Л.Б., Хазина И.В., Кривоногов С.К., Кузьмин Я.В., Прокопенко А.А., и др., 2016. Климатические изменения на юге Западной Сибири в голоцене по результатам анализа ассоциаций остракод // Геология и геофизика. Т. 57. Вып. 4. С. 729–742.
Хотинский Н.А., 1977. Голоцен Северной Евразии: опыт трансконтинентальной корреляции этапов развития растительности и климата: К X Конгрессу INQUA (Великобритания, 1977). М.: Наука. 199 с.
Хотинский Н.А., 1981. Следы прошлого ведут в будущее: очерки палеогеографа. М.: Мысль. 160 с.
Atkinson T.C., Briffa K.R., Coope G.R., 1987. Seasonal temperatures in Britain during the past 22,000 years, reconstructed using beetle remains // Nature. V. 325. P. 587–592.
Blyakharchuk T.A., 2003. Four new pollen sections tracing the Holocene vegetational development of the southern part of the West Siberian Lowland // The Holocene. V. 13. P. 715–731.
Bradshaw R.H.W., Hannon G.E., Lister A.M., 2003. A long-term perspective on ungulate–vegetation interactions // Forest Ecology and Management. V. 181. P. 267–280.
Brooks S.J., 2003. Chironomid analysis to interpret and quantify Holocene climate change // Global change in the Holocene. Birks J., Battarbee R., Mackay A., Oldfield F. (Eds). London: Hodder Arnold publ. P. 328–341.
Brooks S.J., Matthews I.P., Birks H.H., Birks H.J.B., 2012. High resolution Lateglacial and early-Holocene summer air temperature records from Scotland inferred from chironomid assemblages // Quaternary Science Reviews. V. 41. P. 67–82.
Buckland P.C., 1979. Thorne Moors: a palaeoecological study of a Bronze Age site (a contribution to the history of the British insect fauna) // University of Birmingham, Department of Geography Occasional Publication. V. 8. 173 p.
Buckland P.C., Panagiotakopulu E., Buckland P.I., 2004. Fossil insects and the Neolithic: methods and potential // Antaeus. V. 27. P. 235–252.
Gathorne-Hardy F.J., Erlendsson E., Langdon P.G., Edwards K.J., 2009. Lake sediment evidence for late Holocene climate change and landscape erosion in western Iceland // Journal of Paleolimnology. V. 42. P. 413–426.
Huntley B., 1988. Glacial and Holocene vegetation history – 20 ky to present. Europe // Vegetation History. Huntley B., Webb T. III (Eds) Kluiver: Dordrecht. P. 341–383.
Huntley B., 1990. European vegetation history: palaeovegetation maps from pollen data – 13000 ky BP to present // Journal of Quaternary Science. V. 5. P. 103–122.
Ilyashuk E.A., Ilyashuk B.P., 2004. Analysis of chironomid remains from lake sediments in paleoecological reconstruction // Water Resources. V. 31. P. 203–214.
Ilyashuk E.A., Ilyashuk B.P., Hammarlund D., Larocque I., 2005. Holocene climatic and environmental changes inferred from midge (Diptera: Chironomidae, Chaoboridae, Ceratopogonidae) records at Lake Berkut, southern Kola Peninsula, Russia // The Holocene. V. 15. P. 897–914.
Kiselev S.V., Nazarov V.I., 2009. Late Cenozoic insects of northern Eurasia // Paleontological Journal. V. 43. P. 723–850.
Kuzmina E.A., 2009. Late Pleistocene and Holocene small mammal faunas from the South Trans-Urals // Quaternary International. V. 201. P. 25–30.
Lemdahl G., 2000. Late-glacial and early Holocene insect assemblages from sites at different altitudes in the Swiss Alps – implications on climate and environment // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 159. P. 293–312.
Lemdahl G., Gustavsson G., 1997. Lateglacial and middle Holocene Coleoptera assemblages from coastal environments in south-western Sweden // Entomologiske Tidskrift. V. 118. P. 117–187.
Ponel Ph., Gandouin E., Coope G.R., Andrieu-Ponel V., Guiter F., et al., 2007. Insect evidence for environmental and climate changes from Younger Dryas to Sub-Boreal in a river floodplain at St-Momelin (St-Omer basin, northern France), Coleoptera and Trichoptera // Palaeogeography, Palaeoclimatology, Palaeoecology. V. 245. P. 483–504.
Sher A., Kuzmina S., 2007. Beetle records: Late Pleistocene of northern Asia // Encyclopedia of Quaternary Science. Amsterdam: Elsevier, P. 246–267.
Sommer R.S., Benecke N., Löugas L., Nelle O., Schmölcke U., 2011. Holocene survival of the wild horse in Europe: a matter of open landscape? // Journal of Quaternary Science. V. 26. № 8. P. 805–812.
Tarasov P.E., Guiot J., Cheddadi R., Andreev A.A., Bezysko L.G., et al., 1999. Climate in northern Eurasia 6000 years ago reconstructed from pollen data // Earth and Planetary Science Letters. V. 171. P. 635–645.
Usinger H., 2004. Vegetation and climate of the lowlands of northern Central Europe and adjacent areas around the Younger Dryas – Preboreal transition – with special emphasis on the Preboreal oscillation // Hunters in a Changing World. Environment and Archaeology of the Pleistocene–Holocene Transition (ca. 11 000–9000 B. C.) in Northern Europe. Terberger T., Eriksen B.V. (Eds). Marie Leidorf Verlag: Rahden. P. 1–26.
Velichko A.A., Klimanov V.A., Borzenkova I.I., 1992. Climates between 6,000 and 5,500 yr B.P. // Atlas of Palaeoclimates and Palaeoenvironments of the Northern Hemisphere. Frenzel B., Pecsi M., Velichko A.A. (Eds). Budapest: INQUA/Hungarian Academy of Sciences. P. 137–139.
Whitehouse N.J., 2004. Mire ontogeny, environmental and climatic change inferred from fossil beetle successions from Hatfield Moors, eastern England // The Holocene. V. 14. P. 79–93.
Whitehouse N., 2006. The Holocene British and Irish ancient forest fossil beetle fauna: implications for forest history, biodiversity and faunal colonisation // Quaternary Science Reviews. V. 25. № 15. P. 1755–1789.
Zinovyev E.V. 2006. Problems of ecological interpretation of Quaternary insect faunas from the central part of Northern Eurasia // Quaternary Science Reviews. V. 25. P. 1821–1840.
Zinovyev E.V., 2011. Sub-fossil beetle assemblages associated with the “mammoth fauna” in the Late Pleistocene localities of the Ural Mountains and West Siberia // ZooKeys. V. 100. P. 149–169.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал