

Зоологический журнал, 2020, T. 99, № 6, стр. 603-611
Поселения сердцевидки Cerastoderma edule (Linnaeus 1758) (Mollusca, Bivalvia) литорали залива Леврие, Мавритания
А. В. Гущин a, *, П. К. Гудвард b, **
a Институт океанологии им. П.П. Ширшова РАН
117997 Москва, Россия
b Institute for Marine Resources and Ecosystem Studies of Wageningen University Research
1970AB IJmuiden, Netherlands
* E-mail: Poseidon-47@rambler.ru
** E-mail: pcgoudswaard@hotmail.com
Поступила в редакцию 04.07.2019
После доработки 15.10.2019
Принята к публикации 15.01.2020
Аннотация
Изучены поселения сердцевидки съедобной (Cerastoderma edule) в заливе Леврие (Мавритания) на южной границе ареала этого вида. Поселения в заливе Леврие располагаются на литорали в закрытых бухтах, где прибойное воздействие на поселение моллюсков минимально, но хорошо выражено воздействие прилива, осушающего часть поселения. Стацией сердцевидки являются преимущественно песчаные и в меньшей степени песчано-илистые грунты. Изучены морфологические особенности, размерно-массовый состав, численность и биомасса моллюсков в поселении. Анализируются различия размерно-массового состава, численности и биомассы на участке, где сердцевидка добывается, и на участке, где ее запас не эксплуатируется. Приводятся численность и биомасса организмов бентоса, сопутствующих сердцевидке.
Сердцевидка съедобная (Cerastoderma edule (Linnaeus 1758)) обитает в водах литорали побережья Европы от побережья Баренцева моря до Средиземного и Черного морей (Генельт-Яновский, 2010; Tebble, 1966; Hayward, Ryland, 1995; Beukema, Dekker, 2006). В водах Северной Африки сердцевидка встречается на литорали Марокко (Lloris, Rucabado, 1998) и Мавритании (Wolff et al., 1993). Наиболее высокая численность сердцевидки – несколько тысяч экз./м2 – на литоралях Голландии, Англии и Ирландии (Tebble, 1966; Coosen et al., 1994; Ramon, 2003). Южной границей ареала сердцевидки считается Сенегал (Tebble, 1966; Hayward, Ryland, 1995), но найти упоминание о конкретном месте поселения сердцевидки на литорали Сенегала авторам не удалось. Обследование литорали от мыса Тимирис до р. Сенегал, включая дельту р. Сенегал со стороны Мавритании, проведенное авторами в 2003–2004 гг., показало, что поселений сердцевидки на литорали океанического побережья нет. Таким образом, вероятно, южная граница ареала сердцевидки проходит через залив Арген и входящий в него на севере залив Леврие.
Практически на всем ареале обитания сердцевидка служит объектом промысла. В ряде работ (Генельт-Яновский, 2010; Coosen et al., 1994; Ramon, 2003) подтверждается распространенное мнение, что численность и биомасса этого моллюска уменьшаются к границам ареала. Однако на литорали бухт Этуаль и Архимед, где сложились определенные местные условия, это не так, а численность сердцевидки вполне соизмерима с численностью, отмеченной для побережья Европы за исключением литорали Голландии (Dabouineau, Ponsero, 2009).
Целью данной работы является представление данных по морфологии, численности и биомассе сердцевидки на южной границе ареала, описание связи с условиями обитания и определение воздействия промысла на поселение.
МАТЕРИАЛ И МЕТОДИКА
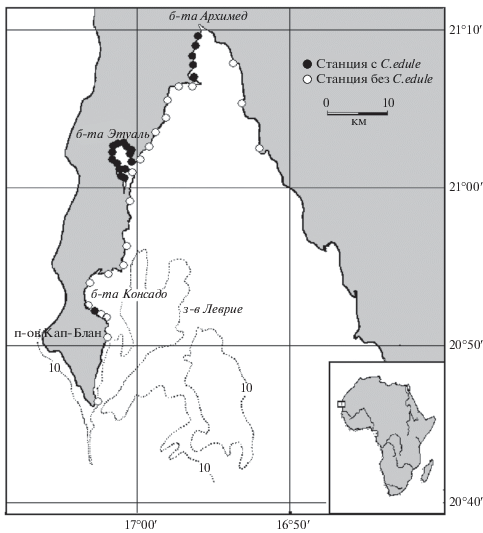
Изучение поселений сердцевидки литорали11 залива Леврие проводилось авторами в июне–августе 2004 г. Всего была выполнена 41 станция, из которых сердцевидка была найдена на двадцати двух (рис. 1). На станции в бухте Консадо и двух станциях в бухте Этуаль сердцевидка была обнаружена единично, эти станции были исключены из анализа. Было изучено 1245 экз. (474 экз. в бухте Архимед, 771 экз. в бухте Этуаль) с определением длины раковины (L) и массы моллюска (P). Для 30 экз. были определены длина раковины (L), высота раковины (H), ширина раковины (E) и масса моллюска (P).
Взятие проб вели с помощью металлической рамки площадью в 1 м2 и приурочивали к периоду отлива, когда обнажалась большая часть литорали. Положение (координаты) станций определялись с помощью GPS. Критерием выбора места для станции был состав грунта (песок), расстояние от предыдущей станции (150–250 м). В остальном места взятия проб носили случайный характер. На каждой станции отбирали 9–15 проб в радиусе 5–25 м от центрального положения станции. Грунт выбирали до глубины 15 см и промывали через сетной фильтр с ячеей 5 мм, что позволяло достоверно собрать все бентосные организмы размером более 5 мм. Все пробы доставляли в лабораторию, где замораживали для последующей камеральной обработки. Перед анализом моллюсков размораживали, для чего их выкладывали на фильтровальную бумагу, где полностью осушали. Для определения индивидуальной массы моллюска использовали электронные весы “Shimadzu ELB-300” точностью 0.01 г. Размеры моллюсков (длина, ширина и высота раковины) измеряли штангенциркулем с точностью до 0.1 мм.
На каждой станции определяли среднюю биомассу всех встреченных видов моллюсков.
(1)
${{B}_{S}} = \sum {{({{b}_{{{{z}_{1}}}}} + {{b}_{{{{z}_{2}}}}} + \ldots + {{b}_{{{{z}_{i}}}}})} \mathord{\left/ {\vphantom {{({{b}_{{{{z}_{1}}}}} + {{b}_{{{{z}_{2}}}}} + \ldots + {{b}_{{{{z}_{i}}}}})} {\sum ({{S}_{{{{z}_{1}}}}} + {{S}_{{{{z}_{2}}}}} + \ldots + {{S}_{{{{z}_{i}}}}}),}}} \right. \kern-0em} {\sum ({{S}_{{{{z}_{1}}}}} + {{S}_{{{{z}_{2}}}}} + \ldots + {{S}_{{{{z}_{i}}}}}),}}$где: BS – средняя биомасса каждого вида моллюска на станции (г/м2);
${{b}_{{{{z}_{1}}}}},{{b}_{{{{z}_{2}}}}},...,{{b}_{{{{z}_{i}}}}}$ – биомасса пробы каждого вида моллюска, взятой на станции (г);
${{S}_{{{{z}_{1}}}}},{{S}_{{{{z}_{2}}}}},...,{{S}_{{{{z}_{i}}}}}$ – площадь, с которой была взяты проб (м2).
Плотность поселения (экз./м2) моллюсков на станции определяли по тому же принципу, что и биомассу.
РЕЗУЛЬТАТЫ
Условия обитания сердцевидки на литорали залива Леврие
Залив Леврие располагается к северо-востоку от мыса Кап-Блан и имеет две закрытые бухты Этуаль и Архимед (см. рис. 1). Западная сторона залива имеет достаточно узкую литораль шириной от 10 до 60 м. Северная и восточная стороны залива имеют более пологий уклон дна и здесь ширина литорали составляет 50–250 м. Ширина литорали изменяется в течение года, что связано с годовым циклом сизигийных и квадратурных приливов.
Приливы в заливе имеют неправильный полусуточный характер. Максимально зарегистрированная амплитуда прилива составила 2.5 м, высшая точка прилива составляла + 2.3 м, низшая – 0.2 м от уровня моря (Hopper, 2007).
Условия среды обитания в зоне литорали характеризуются ежедневными перепадами температуры и солености воды. Вода, поступающая во время прилива из более глубоководной части залива, имеет температуру 18–24°С (сказывается воздействие Канарского апвеллинга), и соленость 35–36‰. В дневное время вода на литорали прогревается и испаряется, что вызывает повышение температура воды в закрытых бухтах до 24–30°С и солености до 37–38‰.
Поселения сердцевидки приурочены к пескам фракций 0.2–0.65 мм с небольшой примесью алевритов и отсутствуют в зоне чистых алевритов и пелитов, поэтому их нет в северной части бухты Архимед и восточной части залива Леврие, где грунты представлены алевритами и пелитами. В слое песка глубже 5–10 см наблюдается дефицит кислорода, песок приобретает серо-зеленый оттенок и выраженный запах сероводорода. В период осушения литорали в отлив сердцевидка погружается в грунт так, что над раковиной находится 2–3 см грунта. В небольших количествах сердцевидка встречается в зарослях морских трав Zostera noltii и Cymodocea nodosa на локальных свободных от трав участках.
Сердцевидка в заливе Леврие была обнаружена на трех участках литорали. Самый южный участок – южная часть бухты Консадо. Здесь на станции было найдено всего три живых моллюска (ввиду низкой численности моллюсков эта станция в исследованиях не учитывалась), но пустые раковины с разрушившимся конхиолоиновым слоем обычны для верхней зоны литорали южного берега бухты. Поселение сердцевидки в этом месте существует с периода неолита, о чем говорят массовые находки раковин в кухонных кучах на стоянках человека неолита, которые датируются периодом 7000–5000 лет до н. э. (Гущин и др., 2007) и которые располагались рядом со станцией. По-видимому, поселение сердцевидки бухты Консадо находится на грани исчезновения из-за загрязнений от города и порта Нуадибу.
В бухте Этуаль поселение сердцевидки занимает практически всю песчаную литораль берега бухты и протоки, соединяющей бухту и обширную зону марша на юге. В протоке сердцевидка перестает встречаться до места перехода в марши.
В бухте Архимед сердцевидка встречается в западной части, но отсутствует в северной и восточной частях бухты, где грунты литорали состоят из алевритов и пелитов.
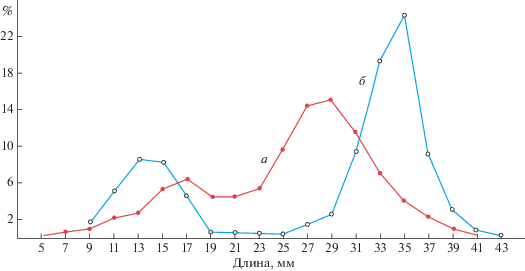
Поселения сердцевидки, обитающей в бухтах Этуаль и Архимед, сходны по многим признакам, но отличаются по средней численности и биомассе, по средним размерам и средней массе моллюсков, по максимальной численности и биомассе. Сравнение размерных рядов моллюсков бухт Этуаль и Архимед показывает, что они имеют определенную закономерность. В том и другом случае размерный ряд представлен кривой, имеющей две модальные группы, но в бухте Этуаль кривая более сглажена и модальные группы менее выражены. Наблюдается низкая численность моллюсков группы 19–25 мм, высокая у группы 9–17 мм и наиболее высокая у группы 25–31 мм в бухте Этуаль и 31–37 мм в бухте Архимед. Наиболее крупные (старые) моллюски встречаются в бухте Архимед (рис. 2).
Состав фауны литорали мест обитания сердцевидки
Фауна литорали песков бухт Этуаль и Архимед (на станциях) представлена несколькими видами макробентоса (табл. 1). Моллюск Dosinia lupinus занимает второе место по встречаемости после сердцевидки – 75% случаев. Mitra cornicula и Turritella ligar встречались в 20% случаев, Persicula cingulata в 10% случаев, Marginella deliciosa efasciata и Marginella glabella в 5% случаев. Представители Amphipoda и Polychaeta встречались в 5% случаев. Крупные гастроподы Cymbium pero, Cymbium cymbium, Purgilina morio обычны на литорали, но в случайных пробах на станциях встречены не были.
Таблица 1.
Состав сообщества сердцевидки бухт Этуаль и Архимед
| Станция | Координаты, с.ш./з.д. | Дата | Виды | Плотность, экз./м2 | Биомасса, г/м2 |
|---|---|---|---|---|---|
| Бухта Этуаль | |||||
| 1 | 21°01′027/17°00′943 | 25.06.2004 | Cerastoderma edule | 73 | 513.0 |
| Всего | 73 | 513.0 | |||
| 2 | 21°01′829/17°01′535 | 02.07.2004 | Cerastoderma edule | 35 | 46.2 |
| Dosinia lupinus | 13 | 17.8 | |||
| Mitra cornicula | 4 | 5.3 | |||
| Turritella ligar | 3 | 3.6 | |||
| Всего | 55 | 72.9 | |||
| 3 | 21°01′848/17°01′539 | 03.07.2004 | Cerastoderma edule | 3 | 9.0 |
| Dosinia lupinus | 7 | 9.4 | |||
| Persicula cingulata | 5 | 7.0 | |||
| Mitra cornicula | 1 | 0.4 | |||
| Всего | 16 | 25.8 | |||
| 4 | 21°02′025/17°01′568 | 04.07.2004 | Cerastoderma edule | 16 | 126.6 |
| Dosinia lupinus | 92 | 176.1 | |||
| Всего | 108 | 302.8 | |||
| 5 | 21°02′310/17°01′650 | 09.07.2004 | Cerastoderma edule | 64 | 202.3 |
| Dosinia lupinus | 26 | 12.8 | |||
| Persicula cingulata | 2 | 0.8 | |||
| Mitra cornicula | 600 | 2.3 | |||
| Всего | 692 | 218.2 | |||
| 6 | 21°02′520/17°01′559 | 17.07.2004 | Cerastoderma edule | 35 | 232.4 |
| Dosinia lupinus | 58 | 50.4 | |||
| Polychaeta | 23 | 0.1 | |||
| Всего | 282.9 | ||||
| 7 | 21°02′543/17°01′303 | 18.07.2004 | Cerastoderma edule | 75 | 539.3 |
| Dosinia lupinus | 1 | 2.6 | |||
| Amphipoda | 4 | 0.1 | |||
| Всего | 80 | 542 | |||
| 8 | 21°02′620/17°01′493 | 17.07.2004 | Cerastoderma edule | 94 | 851.9 |
| Всего | 94 | 851.9 | |||
| 9 | 20°02′571/17°01′270 | 20.08.2004 | Cerastoderma edule | 17 | 185.4 |
| Dosinia lupinus | 17 | 25.8 | |||
| Всего | 34 | 211.3 | |||
| 10 | 21°02′591/17°00′738 | 20.08.2004 | Cerastoderma edule | 41 | 214.8 |
| Dosinioa lupinus | 11 | 12.0 | |||
| Mitra cornicula | 1 | 0.2 | |||
| Turrittella ligar | 4 | 1.0 | |||
| Marginella glabella | 2 | 0.8 | |||
| Всего | 59 | 228.8 | |||
| 11 | 21°02′457/17°00′446 | 20.08.2004 | Cerastoderma edule | 14 | 40.6 |
| Dosinia lupinus | 1 | 0.3 | |||
| Всего | 15 | 40.9 | |||
| 12 | 21°02′242/17°00′194 | 20.08.2004 | Cerastoderma edule | 25 | 196.5 |
| Dosinia lupinus | 1 | 1.1 | |||
| Всего | 27 | 197.6 | |||
| 13 | 21°02′000/17°00′143 | 20.08.2004 | Cerastoderma edule | 50 | 257.7 |
| Dosinia lupinus | 1 | 0.2 | |||
| Всего | 51 | 257.9 | |||
| 14 | 21°01′630/17°00′205 | 20.08.2004 | Cerastoderma edule | 6 | 48.0 |
| Dosinia lupinus | 1 | 1.0 | |||
| Всего | 7 | 49.0 | |||
| Бухта Архимед | |||||
| 1 | 21°09′510/16°55′708 | 24.07.2004 | Cerastoderma edule | 68 | 462.9 |
| Dosinia lupinus | 8 | 16.1 | |||
| Всего | 76 | 479 | |||
| 2 | 21°07′813/16°56′299 | 24.07.2004 | Cerastoderma edule | 56 | 610.7 |
| Dosinia lupinus | 1 | 0.7 | |||
| Всего | 57 | 611.4 | |||
| 3 | 21°06′563/16°56′496 | 24.07.2004 | Cerastoderma edule | 5 | 41.9 |
| Turritella ligar | 21 | 23.8 | |||
| Всего | 26 | 65.8 | |||
| 4 | 21°06′711/16°56′575 | 20.08.2004 | Cerastoderma edule | 19 | 228.9 |
| Dosinia lupinus | 2 | 4.8 | |||
| Marginella deliciosa | 1 | 0.2 | |||
| Turritella ligar | 13 | 10.5 | |||
| Всего | 35 | 244.4 | |||
| 8 | 21°08′728/16°55′930 | 20.08.2004 | Cerastoderma edule | 30 | 453.0 |
| Всего | 30 | 453.0 | |||
В бухте Этуаль на песчаных грунтах сердцевидка была найдена на 14 станциях. На 12 станциях сердцевидка была доминирующим видом. Только на станциях номер 3 и 4, расположенных на границе зарослей морских трав Z. noltii и C. nodosa, где грунты в большей степени состояли из алевритов, руководящим видом была Dosinia lupinus.
В бухте Архимед на всей литорали, представленной песчаными грунтами, сердцевидка была доминирующим видом.
Сердцевидка в поселениях распределялась мозаично. Численность и биомасса моллюсков сильно различалась на близко расположенных местах.
В поселениях моллюски были представлены особями длиной раковины от 4.1 до 42.3 мм и массой от 0.11 до 25.04 г (рис. 2, 3).
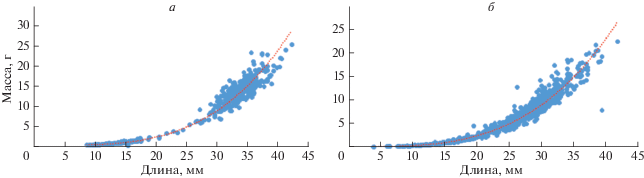
Соотношение масса – длина раковины у сердцевидки для бухты Этуаль описывается степенным уравнением P = 0.0001 L3.2833 при величине аппроксимации r 2 = 0.9795; для бухты Архимед P = 0.00006 L3.4889 при величине аппроксимации r 2 = 0.9821, где P – масса моллюска, L – длина раковины.
Соотношение высоты раковины (H) к длине раковины (L) описывается уравнением H = = 0.9566 L0.9733 при величине аппроксимации r 2 = = 0.9202. Соотношение ширины раковины (E) к длине раковины (L) описывается уравнением E = 0.6367 L1.0385 при величине аппроксимации r 2 = 0.8112.
Средняя численность по станциям в бухте Этуаль выше, чем в бухте Архимед, но средняя биомасса выше в бухте Архимед (табл. 2). Сердцевидка бухты Архимед массивнее сердцевидки бухты Этуаль (табл. 3).
Таблица 2.
Средняя численность и биомасса сердцевидки в поселениях бухт Этуаль и Архимед
| Показатель | Количество станций | Среднее | Минимум | Максимум |
|---|---|---|---|---|
| Бухта Этуаль | ||||
| Численность, экз./м2 | 14 | $\frac{{39.17 \pm 8.36}}{{28.96}}$ | 3.00 | 94.00 |
| Биомасса, г/м2 | 14 | $\frac{{268.40 \pm 71.53}}{{247.78}}$ | 9.00 | 851.90 |
| Бухта Архимед | ||||
| Численность, экз./м2 | 5 | $\frac{{35.60 \pm 11.639}}{{26.02}}$ | 5.00 | 68.00 |
| Биомасса, г/м2 | 5 | $\frac{{359.48 \pm 100.11}}{{223.86}}$ | 41.90 | 610.70 |
Таблица 3.
Средняя длина и масса сердцевидки в поселениях бухт Этуаль и Архимед
| Показатель | Количество станций | Среднее | Минимум | Максимум |
|---|---|---|---|---|
| Бухта Этуаль | ||||
| Длина, мм | 771 | $\frac{{25.28 \pm 0.249}}{{6.91}}$ | 3.90 | 41.80 |
| Масса, г | 771 | $\frac{{6.50 \pm 0.16}}{{4.51}}$ | 0.01 | 22.34 |
| Бухта Архимед | ||||
| Длина, мм | 474 | $\frac{{27.97 \pm 0.43}}{{9.47}}$ | 8.60 | 42.30 |
| Масса, г | 474 | $\frac{{9.76 \pm 0.30}}{{6.53}}$ | 0.04 | 25.04 |
ОБСУЖДЕНИЕ
Считается, что показатели численности и стабильности поселений сердцевидки по всему ареалу уменьшаются к границам ареала. Генельт-Яновский (Генельт-Яновский, Полоскин, 2004; Генельт-Яновский, 2010) сравнил численность сердцевидки Мурманского побережья Баренцева моря (4–20 экз./м2) с ее обилием в центральной части ареала, где численность составляет тысячи экз./м2 (Coosen et al., 1994; Ramon, 2003). Численность в заливе Леврие не достигает показателей численности в центральной части ареала, но вполне соотносится с численностью в других частях побережья Европы (Dabouineau, Ponsero, 2009).
Наблюдаемые различия в биомассе и численности сердцевидки залива Леврие в Мавритании, куда входят бухты Этуаль и Архимед, можно объяснить тем, что популяция сердцевидки бухты Этуаль эксплуатируется, а популяция бухты Архимед не используется. Сборщики моллюсков отбирают крупные экземпляры моллюска, а мелкие отбрасывают, таким образом моллюски перераспределяются по литорали. Часть мелких моллюсков погибает, не имея возможности зарыться в осушенный песок. Промысел привел к тому, что численность размерных групп длиной более 31 мм (наиболее привлекательный для промысла размер) в бухте Этуаль снижается, и сердцевидка длиной 23–33 мм составляет основу поселения. Моллюски длиной более 35 мм встречаются только в нижней зоне литорали, более труднодоступной для собирателей моллюсков, так как большую часть времени они находятся под водой. В бухте Архимед (популяция не эксплуатируется) моллюски длиной 30–40 мм составляют большую часть популяции.
Наиболее вероятной причиной малой численности сердцевидки небольших размеров в период наблюдений является то, что зона литорали служит местом концентрации тысяч птиц, которые питаются мелкими моллюсками. Здесь наблюдаются кулики Calidris alba, C. alpina, C. canutus, C. ferruginea, C. minuta, Haematopus ostralegus, кроншнеп Numenius phaeopus и фламинго Phoenicopterus ruber, которые питаются организмами бентоса на литорали (Swennen, 1990; Zwarts et al., 1990). Птицы ограничивают численность сердцевидки, но существует возможность пополнения популяции за счёт того, что сердцевидка размножается практически круглогодично, а массовое появление птиц сезонно. Наибольшее количество перелетных птиц встречается в заливе Арген, куда входит залив Леврие, в марте–мае и сентябре–январе (Wolff, Smit, 1990; Gowthorpe, 1993). Таким образом, остается время, когда пресс птиц на популяцию сердцевидки менее выражен. Часть молоди моллюсков становится пищей для рыб. Однако выедание молоди сердцевидки рыбами относительно невелико. Из 8 видов бентосоядных рыб, постоянно встречающихся на литорали залива, только 3 вида (Diplodus bellottii, Solea senegalensis, Eucinostomus melanopterus) используют в пищу двустворчатых моллюсков, в том числе сердцевидку (Гущин, 2013). Вероятно, птицы являются главными хищниками, потребляющими сердцевидку в зоне литорали.
ЗАКЛЮЧЕНИЕ
Поселения сердцевидки, обитающей в бухтах Этуаль и Архимед, сходны по многим признакам, но различаются по средней численности и биомассе, по средним размерам и средней массе моллюсков, по максимальной численности и биомассе. Сравнение размерных рядов сердцевидки бухт Этуаль и Архимед показывает, что между ними существует определенное сходство. В том и другом случае размерный ряд представлен кривой, имеющей две модальные группы, но в бухте Этуаль кривая более сглажена и модальные группы менее выражены. Наблюдается низкая численность моллюсков группы 19–25 мм, высокая у группы 9–17 мм и наиболее высокая у группы 25–31 мм в бухте Этуаль и 31–37 мм в бухте Архимед. Наиболее крупные (старые) моллюски встречаются в бухте Архимед (см. рис. 2).
Поселения сердцевидки, обитающей в южной части ареала этого вида, в заливе Леврие располагаются на литорали в закрытых бухтах с минимальным прибойным и выраженным приливным воздействием. Поселения встречаются на песчаных грунтах. На большинстве станций, где грунты представлены песчаными фракциями, сердцевидка является доминирующим видом. В бухте Этуаль средняя биомасса и численность сердцевидки 268.4 г/м2 и 39.2 экз./м2, средняя длина и масса моллюска 25.3 мм и 6.5 г. В бухте Архимед средняя биомасса и численность сердцевидки 359.5 г/м2 и 35.6 экз./м2, средняя длина и масса моллюска 27.9 мм и 9.8 г. Различия в средней численности и биомассе, в средней длине и массе моллюска, вероятно, связаны с тем, что популяция сердцевидки бухты Этуаль эксплуатируется, а популяция бухты Архимед нет. Небольшая численность молоди сердцевидки в момент исследования, возможно, связана с выеданием моллюсков небольших размеров птицами и в меньшей степени рыбами. В этом случае пополнение популяции молодью осуществляется в июне–августе и феврале, в месяцы минимального присутствия перелетных птиц в заливе Леврие.
Список литературы
Генельт-Яновский Е.А., 2010. Популяционная биология и распространение Cerastoderma edule (Linnaeus, 1758) на Северо-Восточной границе ареала (Мурманское побережье Баренцева моря). Автореф. дис. … канд. биол. наук. С.-П.: Государственный университет. 18 с.
Генельт-Яновский Е.А., Полоскин А.В., 2004. Современное состояние популяции Cerastoderma edule (L.) на литорали Дальнего пляжа (Баренцево море, Восточный Мурман) // Вестник С.-Петербургского университета. Т. 3. № 4. С. 17–23.
Гущин А.В., 2013. Питание молоди рыб литорали залива Арген, Мавритания // Вопросы ихтиологии. Т. 53. № 6. С. 718–725. https://doi.org/10.1134/S0032945213050068
Гущин А.В., Гущина Т.А., Клепицкая А.М., Шаврина И.А., 2007. Человек у океана: история длиной в 8000 лет // Вестник Российского государственного университета им. И. Канта. Естественные науки. Калининград. Т. 7. С. 43–49.
Beukema J.J., Dekker R., 2006. Annual cockle Cerastoderma edule production in the Wadden Sea usually fails to sustain both wintering birds and a commercial fishery // Marine Ecology Progress Series 309. P. 189–204.
Coosen J., Twisk F., van der Tol M.W.M., Lambeck R.H.D., van Stralen M.K., Meire P.M., 1994. Variability in stock assessment of cockles (Cerastoderma edule L.) in the Oosterschelde (in 1980-1990) in relation to environmental factors // Hydrobiologia. V. 282/283. Kluwer Academic Publishers, Dordrecht. P. 381–395.
Dabouineau L., Ponsero A., 2009. Synthesis on biology of Common European Cockle Cerastoderma edule. hal-00581394. [Электронный ресурс]. Режим доступа: https://hal.archives-ouvertes.fr/file/index/docid/581394/ filename/Synthesis_on_biology_of_Europeen_cockle. pdf Дата обновления: 20.06.2019
Gowthorpe P., 1993. Une visite au Parc National du Banc d’’Arguin: itineraries présentation des principales composantes naturalles // Mauritania, Nouakchott: Parc National du Banc d’Ar\guin. 193 p.
Hopper M., 2007. WXTide32 – a free Windows tide and current prediction program. [Электронный ресурс]. Режим доступа: http://wxtide32.com/ Дата обновления: 20.06.2019
Hayward P.J., Ryland J.S., 1995. Handbook of the marine fauna of Nord West Europe. Oxford: Oxford University Press. 816 p.
Lloris D., Rucabado J., 1998. Guide d’Identification des resources marines vivantes du Maroc. Rome: FAO. 263 p.
Ramon M., 2003. Population dynamics and secondary production of the cockle Cerastoderma edule (L.) in a back barrier tidal flat of Wadden Sea // Scientia Marina. V. 67 (4). P. 429–443.
Swennen C., 1990. Oystercatchers feeding on giant blood cockles on the Banc d’Arguin, Mauritania // Ardea. V. 78 (1/2). P. 53−62.
Tebble N., 1966. British bivalve seashells. A Handbook for Identification. British Museum. Oxford. 213 p.
Wolff W.J., Duiven A.G., Duiven P., Esselink P., Abou Gueye, Meijboom A., Moerland G., Zegers J., 1993. Biomass of macrobenthic tidal flat fauna of the Banc d’Arguin, Mauritania // Hydrobiologia. Kluwer Academic Publishers, Dordrecht. V. 258. P. 151–164.
Wolff W.J., Smit C.J., 1990. The Banc d’Arguin, Mauritania, as an environment for coastal birds // Ardea. V. 78 (1/2). P. 17−38.
Zwarts L., Blomert A.M., Ens B.J., Hupkes R., van Spanje T.M., 1990. Why do waders reach high feeding densities on the intertidal flats of the Banc d’Arguin, Mauritania // Ardea. V. 78 (1/2). P. 39–52.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал