

Зоологический журнал, 2020, T. 99, № 8, стр. 924-937
Размерные, возрастные, фенетические, морфофизиологические и цитогенетические характеристики популяций озерной лягушки (Pelophylax ridibundus) (Amphibia, Ranidae) загрязненных термальных водоемов Камчатки
Е. Б. Романова a, *, Е. С. Рябинина a, **, С. М. Ляпков b, ***
a Нижегородский государственный университет им. Н.И. Лобачевского
603950 Нижний Новгород, Россия
b Московский государственный университет им. М.В. Ломоносова
119234 Москва, Россия
* E-mail: romanova@ibbm.unn.ru
** E-mail: ryabinina.e.s@yandex.ru
*** E-mail: lyapkov@mail.ru
Поступила в редакцию 11.07.2019
После доработки 16.10.2019
Принята к публикации 17.12.2019
Аннотация
Сборы озерных лягушек (Pelophylax ridibundus) были проведены в двух термальных водоемах в юго-восточной части Камчатки: в сбросном канале ТЭЦ-2 г. Петропавловска-Камчатского (далее – ТЭЦ) и в сбросах термальных вод в пос. Паратунка. В водоеме ТЭЦ было выше содержание железа (в 7 раз) и марганца (в 1.2 раза) по сравнению с водоемом Паратунки, а содержание кобальта, меди и никеля – ниже. В популяции Паратунка преобладали морфы striata и hemimaculata; в популяции ТЭЦ – пятнистые (maculata), светлогорлые, светлобрюхие особи. Средние значения возраста, длины и массы тела, а также темпов роста после метаморфоза были выше и у самцов, и у самок популяции ТЭЦ по сравнению с популяцией Паратунка. Относительная масса семенников самцов из популяции ТЭЦ была значимо выше, чем у самцов из Паратунки, что можно считать компенсаторной реакцией на комплекс загрязнителей. Впервые выявлено преобладание эритроцитов с прикрепленными микроядрами у озерных лягушек популяций Камчатки, свидетельствующее о хромосомных аберрациях, индуцированных мутагенами водной среды. У лягушек из популяции Паратунка выявлены половые различия по содержанию эритроцитов с микроядрами, свидетельствующие о более высокой частоте мутаций в эритроцитах самцов по сравнению с самками. Показано возрастание доли микроядер в эритроцитах особей старших возрастов, что, по-видимому, связано с кумулятивным эффектом ксенобиотиков водной среды. Наибольший относительный вклад в общую дисперсию совокупности исследованных характеристик озерных лягушек двух популяций вносили морфофизиологические индексы (относительная масса) органов; в половые различия (только в популяции Паратунка) – цитогенетические аномалии в соматических клетках, выявляемые микроядерным тестом.
Антропогенное загрязнение окружающей среды снижает эффективность работы основных регулярных систем (нервной, иммунной, эндокринной), что способствует росту частоты хромосомных нарушений, приводит к изменению генетических показателей и нарушению цитогенетической стабильности организма. Распространенным методом в скрининге и мониторинге генотоксического загрязнения территорий и оценке хромосомных нарушений является микроядерный тест, при котором происходит учет микроядер в клетках пролиферирующих тканей организма (Жулева, Дубинин 1994; Hayashi et al., 2000; Ковалева, 2008 и др.). Такие методики были успешно применены к массовым видам бесхвостых амфибий водоемов европейской части России, том числе – к озерной лягушке (Кармазин, Пескова, 2010; Спирина и др., 2011; Romanova, Ryabinina, 2018; Романова и др., 2018а; Рябчикова и др., 2019 и др.), с целью оценки мутагенного потенциала среды обитания. Проведена дифференцированная оценка форменных элементов крови и видов микроядер в эритроцитах озерных лягушек, обитающих в водоемах Нижегородской обл., различающихся по происхождению, морфологии и химическим характеристикам. Специфические абиотические условия и антропогенная нагрузка приводили к наиболее выраженному изменению лейкоцитарного состава крови, соответствующему нейтрофильному типу лейкемоидной реакции и возрастанию доли микроядер в эритроцитах. Установлена положительная корреляция между количеством разрыхленных микроядер и содержанием в водоеме нитритов (Романова и др., 2018).
Возрастной состав относится к числу важнейших популяционных характеристик. Кроме того, поскольку рост амфибий умеренной зоны происходит ритмически, во время периода активности в теплое время года, изучение скорости постметаморфозного роста невозможно без знания возраста данной особи. В исследованиях амфибий в последние 45 лет общепринятым стал точный метод определения возраста особей, основанный на подсчете числа линий зимовок на окрашенных срезах трубчатых костей, т.е. скелетохронология, позволяющая также оценить величину ежегодных приростов данной особи (обзор – Клевезаль, Смирина, 2016). Имеется немало данных по внутрипопуляционной и межпопуляционной (географической) изменчивости размеров и темпов роста, основанных на определении возраста прудовых (Боркин, Тихенко, 1979) и озерных лягушек (Ляпков, 2016а, обзор – Фоминых, Ляпков, 2011). Анализ возраста животных имеет большое значение еще и потому, что во многих случаях необходима количественная оценка хромосомных нарушений в зависимости от возраста особей для выявления кумулятивного эффекта поллютантов, влияющих на процессы митоза и функцию кроветворения. Особенно актуальны такие исследования при изучении жизнеспособности популяций животных в условиях антропогенно трансформированных экосистем. Обитание в таких экосистемах ведет к нарушениям цитогенетического гомеостаза особей и их популяций, которые выражаются в наличии хромосомных мутаций в виде микроядер. Активность процессов кроветворения, стабильность генома и активация иммунитета амфибий могут иметь прямую связь с размерами костномозговой полости трубчатых костей бедра, голени, а также с темпами резорбции эндоста и внутренней части периоста, изменяющимися по мере взросления особей.
Озерная лягушка была завезена на Камчатку и сформировала здесь несколько самоподдерживающихся популяций в подогреваемых водоемах около 30 лет назад (Ляпков, 2014; Ляпков и др., 2017). Согласно нашим предварительным наблюдениям, успешность освоения видом новых местообитаний связана также с устойчивостью к загрязнению населяемых водоемов.
Цель работы – оценка морфофизиологических, фенотипических, гематологических характеристик и степени цитогенетических нарушений у озерных лягушек, которые населяют термальные водоемы Камчатки и различаются по полу, размеру и возрасту.
МАТЕРИАЛ И МЕТОДИКА
Материалом работы служили сборы озерных лягушек (Pelophylax ridibundus (Pallas 1771)) (49 особей), 16–19 ноября 2018 г. в двух точках юго-восточной части Камчатки: в водоеме, образованном сбросами термальных вод в пос. Паратунка (52.963350 N, 158.262578 E, далее – Паратунка, 31 особь) и в сбросном канале, вытекающем из пруда-охладителя ТЭЦ-2 г. Петропавловска-Камчатского (53.016605 N, 158.737855 E, далее – ТЭЦ, 18 особей). Две эти популяции разделены пространственно. Подробное описание их местообитаний было приведено ранее (Ляпков, 2014). Благодаря благоприятным условиям роста и развития головастиков и завершивших метаморфоз особей, регулярному размножению (в течение 8–10 месяцев в году), сформировавшиеся популяции стабильны и довольно многочисленны (Ляпков, 2016). Для данного исследования использовали только сравнительно крупных особей (с длиной тела не менее 40 мм), бóльшая часть которых была половозрелыми.
Массу тела лягушек определяли на технических весах с точностью до 0.02 г (ошибка измерения ± 0.01), а массу органов (печени, селезенки, сердца, почки, тимуса, яичников у самок (зрелые яйца в яичниках и полости тела не взвешивали) или семенников у самцов – на торсионных весах Waga Torsyjna-WT с точностью до 0.1 мг (ошибка измерения ± 0.03). Индексы органов вычисляли как отношение массы органа к массе тела (‰) (Ивантер и др., 1985).
При фенотипическом анализе популяций озерных лягушек использовали классификацию фенов по методике Ищенко (1978).
Число эритроцитов (тыс./мм3), лейкоцитов (тыс./мм3) определяли в камере Горяева общепринятым методом (Меньшиков и др., 1987). Подсчет микроядер (Жулева, Дубинин, 1994) осуществляли, используя микроскоп Meiji Techno с иммерсионным объективом при общем увеличении ×1500 (100 × 10 × 1.5), просматривая для каждой особи по 2000 эритроцитов. В связи с отсутствием в литературе четких количественных и качественных классификационных характеристик шести видов микроядер нами предложено выделять в эритроцитах амфибий с учетом формы микроядра и его расположения относительно основного клеточного ядра четыре вида микроядер: 1) хорошо оформленные, обособленные; 2) прикрепленные; 3) палочковидные и 4) разрыхленные (Романова и др., 2018а).
Изучение возрастного состава проводили с использованием стандартных методов скелетохронологии (Смирина, 1983; Smirina, 1994). У всех особей определяли возраст по ростовым слоям, которые исследовали на окрашенных гематоксилином Эрлиха поперечных срезах (толщиной 15 µ), изготовленных из середины диафиза голени. Для ретроспективной оценки длины тела в каждом из возрастов использовали величины внешнего диаметра поперечных срезов голени и диаметров каждой из линий склеивания. Эти величины определяли как среднее между минимальным и максимальным диаметром данной линии, измеренными на срезах. “Обратное расчисление” длины тела проводили с помощью наиболее часто используемого (Marunouchi et al., 2000) уравнения Даля-Леа: Li = L × Di/Dвнеш, где Li – рассчитанная длина тела в данном возрасте i, Di – диаметр соответствующей линии склеивания, Dвнеш – внешний диаметр среза, измеренный у пойманной особи, L – длина тела пойманной особи.
В исследованных водоемах в прибрежной зоне были отобраны пробы воды, в которых с помощью спектрофотометра Hach DR-2800 определены химические загрязнители: железо общее, марганец, медь, нитрит-ионы (NO2), нитрат-ионы (NO3), сульфаты, сульфиды, кобальт, никель. Относительную степень загрязнения оценивали по коэффициенту комплексности загрязненности воды, рассчитанному по 13 ингредиентам и показателям качества воды, отвечающим свободному перечню № 3 (РД 52.24.643-2002), учитывая показатели, превышающие норматив качества воды для водоемов рыбохозяйственного назначения (ПДКрыб-хоз) (Нормативы качества воды…, 2010). Норматив качества воды водных объектов рыбохозяйственного назначения был выбран нами, как наиболее жесткий из существующей нормативно-технической документации.
Предварительная проверка первичных данных показала их несоответствие нормальному распределению. Поэтому далее статистический анализ проводили методами непараметрической статистики с использованием критериев: Манна-Уитни (при парном сравнении независимых групп, U); Данна (множественный критерий при попарном сравнении групп, D), коэффициента гамма-корреляции (при анализе взаимосвязи, с учетом совпадающих значений, r) (Реброва, 2006), критерия z (при сравнении долей) и методом главных компонент с использованием пакета статистических программ STATISTICA 10.0. За величину уровня статистической значимости принимали 0.05. При проведении множественных сравнений производилась коррекция критического уровня значимости с помощью поправки Бонферрони (Гланц, 1999).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Результаты гидрохимического анализа воды исследованных местообитаний
Гидрохимические условия исследованных водоемов различались по содержанию отдельных химических элементов. Водоем ТЭЦ характеризовался высоким содержанием железа (4.15 ПДК), марганца (35 ПДК), меди (65 ПДК), никеля (4.1 ПДК), сульфидов (2.8 ПДК), кобальта (2.6 ПДК). В водоеме Паратунка выявлены превышения ПДК по меди (в 100 раз), марганцу (в 30 раз), кобальту (в 12.3 раза), никелю (в 5.4 раза) и сульфидам (в 2.2 раза). Коэффициент комплексности загрязненности воды в водоеме ТЭЦ составил 66.6%, в водоеме Паратунки – 55.5%. При этом гидрохимические показатели в искусственном водоеме ТЭЦ превышали таковые водоема Паратунки по содержанию железа (в 7 раз) и марганца (в 1.16 раза). Концентрации кобальта, меди и никеля были выше в водоеме Паратунки по сравнению с ТЭЦ в 4.73, 1.53 и 1.31 раза соответственно.
Фенотипический состав исследованных популяций
При изучении окраски спины и брюха озерных лягушек исследованных водоемов были идентифицированы следующие отдельные “морфы” (Ищенко, 1978), фактически – фены: полосатый, т.е. наличие дорсомедиальной светлой полосы (striata, s), альтернативой которому является отсутствие такой полосы (ns); пятнистый (maculata, m), с альтернативой hemimaculata (hm) – уменьшенное количество пятен на спине; nigricollis (nc) – темная окраска горла, с альтернативой albicollis (ac – светлая окраска горла; nigriventris (nv) – темная окраска брюха, с альтернативой albiventris (av) – светлая окраска брюха.
В популяции озерных лягушек ТЭЦ преобладали пятнистые, светлогорлые, светлобрюхие особи (табл. 1). В популяции Паратунка также преобладали светлобрюхие и светлогорлые особи. Анализ различий встречаемости фенотипов и фенов между популяциями выявил преобладание в популяции Паратунка доли особей striata с небольшим числом пятен на спине (hm) (табл. 1).
Таблица 1.
Встречаемость фенотипов в исследованных выборках озерных лягушек
| Популяция, показатель | s | ns | m | hm | ac | nc | av | nv |
|---|---|---|---|---|---|---|---|---|
| ТЭЦ (1) | 6/0.33 | 12/0.67 | 16/0.88 | 1/0.05 | 14/0.77 | 4/0.23 | 18/1.00 | 0 |
| Паратунка (2) | 22/0.71 | 9/0.29 | 20/0.64 | 11/0.35 | 27/0.87 | 4/0.13 | 29/0.93 | 2/0.07 |
| Сравнение долей | z1–2 = 2.56, р = 0.01 | z1–2 = 9.17, р < 0.01 | z1–2 = 1.86, р = 0.10 | z1–2 = 2.34, р = 0.02 | z1–2 = 0.85, р > 0.05 | z1–2 = 0.85, р > 0.05 | z1–2 = 1.11, р = 0.20 | – |
Согласно литературным данным, наличие светлой дорсомедиальной полосы у остромордой лягушки определяется действием одного доминантного диаллельного аутосомного гена (Ищенко, 1978; Щупак, Ищенко, 1981), сходным образом наследуется эта полоса у озерной лягушки (Berger, Smielowski, 1982), поэтому морфа striata считается хорошим фенетическим маркером, с помощью которого можно анализировать изменения генетической структуры популяции. У остромордой лягушки – широкоареального вида – частота striata сильно изменяется в различных регионах (Ищенко, 1978; Шляхтин, 1985; Лада, 2012; Файзулин, Кузовенко, 2012; Файзулин и др., 2013), а также в местообитаниях с различной степенью урбанизации (Вершинин, 1980, 1982, 1983, 2008). Направленное изменение частот фенотипов, в том числе рост доли особей striata, характерно и для зеленых лягушек, либо с изменением географической широты и долготы (Терентьев, 1962; Боркин, Тихенко, 1979), либо по мере усиления урбанизации (Жукова и др., 1986; Пескова, 1995). Показано также, что у остромордой и озерной лягушек особи этой морфы получают селективные преимущества в популяциях, населяющих антропогенно трансформированные и загрязненные территории (Вершинин, 2004; 2008).
Возрастной, размерный состав и темпы постметаморфозного роста
Данные по размерным характеристикам и возрасту исследованных популяций озерных лягушек приведены на рис. 1 и в табл. 2, 3, 4 и 5. В популяции ТЭЦ наиболее часто встречаются самцы в возрасте 4 лет и самки – в возрасте 5 лет (у самок отличие от других возрастов незначимо, только на уровне тенденции – табл. 2). В популяции Паратунка наиболее часто встречаются самцы в возрасте 3 лет и самки – в возрасте 1 года (рис. 1, табл. 3), что обусловлено включением в выборку всех особей, у которых определяли возраст и исследовали цитогематологические характеристики, в том числе и неполовозрелых. Поскольку особи Паратунки представлены чаще младшими возрастами (у самок популяции Паратунки, статистически значимо ниже, в сравнении с популяцией ТЭЦ только доля 5-летних особей, у самцов – только доля 4-летних особей – табл. 4), их средний возраст значимо более низкий и у самок, и у самцов, в сравнении с популяцией ТЭЦ (см. табл. 5).
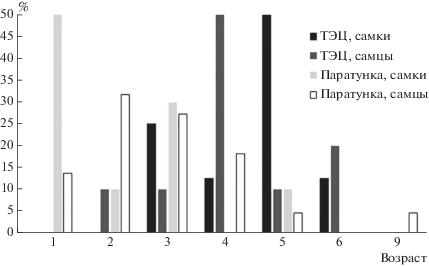
Таблица 2.
Сравнение частот (по критерию z, в ячейках таблицы приведено значение p) возрастов в пределах одного пола в популяции ТЭЦ
| Возраст | 3 | 4 | 5 | 6 |
|---|---|---|---|---|
| 2 | 0.5 | 0.033 | 0.5 | 0.269 |
| 3 | × | 0.033 | 0.5 | 0.269 |
| 4 | 0.266 | × | 0.033 | 0.088 |
| 5 | 0.159 | 0.064 | × | 0.269 |
| 6 | 0.266 | 0.5 | 0.064 | × |
Таблица 3.
Сравнение частот (по критерию z, в ячейках таблицы приведено значение p) возрастов в пределах одного пола в популяции Паратунки
| Возраст | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| 1 | × | 0.079 | 0.133 | 0.339 | 0.151 |
| 2 | 0.033 | × | 0.373 | 0.151 | 0.012 |
| 3 | 0.187 | 0.139 | × | 0.238 | 0.022 |
| 4 | – | – | × | 0.081 | |
| 5 | 0.033 | 0.5 | 0.139 | × |
Таблица 4.
Сравнение возрастного состава (по критерию z, в ячейках таблицы приведено значение p) самцов и самок в одной популяции и двух исследованных популяциях
| Возраст | ТЭЦ, самки ↔ самцы |
Паратунка, самки ↔ самцы |
Самки, ТЭЦ ↔ Паратунка |
Самцы, ТЭЦ ↔ Паратунка |
|---|---|---|---|---|
| 1 | – | 0.018 | – | – |
| 2 | – | 0.098 | – | 0.098 |
| 3 | 0.204 | 0.438 | 0.409 | 0.141 |
| 4 | 0.057 | – | – | 0.037 |
| 5 | 0.039 | 0.279 | 0.039 | 0.279 |
| 6 | 0.339 | – | – |
Таблица 5.
Размеры и возраст озерных лягушек исследованных популяций Камчатки
| Популяция | Показатель | Пол (n) | Длина тела, мм | Масса тела, г | Возраст, годы |
|---|---|---|---|---|---|
| ТЭЦ | M ± m | Самцы (10) | 78.50 ± 2.84 | 51.11 ± 5.63 | 4.20 ± 0.38 |
| M ± m | Самки (8) | 86.62 ± 4.62 | 76.01 ± 9.02 | 4.50 ± 0.37 | |
| U | 2.35 | 2.44 | 0.53 | ||
| p | 0.018 | 0.014 | 0.59 | ||
| Паратунка | M ± m | Самцы (21) | 62.76 ± 3.40 | 29.93 ± 4.82 | 3.00 ± 0.38 |
| M ± m | Самки (10) | 54.20 ± 5.45 | 23.74 ± 10.65 | 2.10 ± 0.43 | |
| U | 1.71 | 1.83 | 1.50 | ||
| p | 0.08 | 0.06 | 0.13 | ||
| Апостериорное сравнение средних для популяций значений | |||||
| D | Самцы | 2.57 | 2.45 | 2.42 | |
| p | 0.009 | 0.01 | 0.01 | ||
| D | Самки | 2.84 | 2.66 | 2.84 | |
| p | 0.004 | 0.007 | 0.004 | ||
В популяции ТЭЦ в отличие от популяции Паратунки выявлены статистически значимые половые различия по длине тела и массе тела (табл. 5).
Межпопуляционные различия в темпах роста (рис. 2) были выявлены только для самцов: в каждом из возрастов самцы ТЭЦ достигали сравнительно более крупных размеров, чем самцы Паратунки (в каждом из возрастов эти различия статистически не значимы, но все самцы популяции ТЭЦ достоверно крупнее, чем Паратунки – см. табл. 5). Кроме того, использование рассчитанных значений длины тела позволяет сделать вывод, что основной причиной более крупных размеров самок в обеих популяциях является их более быстрый, чем у самцов, рост (в обеих популяциях, в каждом из возрастов половые различия статистически незначимы, но все самки достоверно крупнее самцов – см. табл. 4).
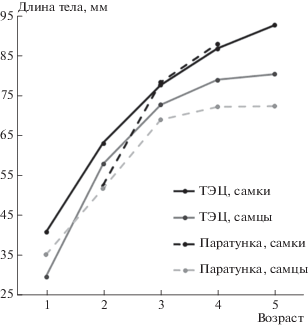
Сравнивая данные, полученные по возрастному составу, темпам роста и размерам, с данными по степени загрязненности водоемов двух исследованных популяций, можно сделать вывод, что более загрязненная среда обитания (в популяции ТЭЦ) не оказывает прямого воздействия на изученные характеристики лягушек.
В целом, размерный и возрастной состав двух исследованных популяций сходен с таковыми, установленным при исследовании летних выборок более ранних сборов (2013–2015 гг. – Ляпков, 2016а): лягушки популяции Паратунка характеризовались сравнительно мелкими размерами в отличие от популяции ТЭЦ, особи которой имели также сравнительно большой средний возраст.
Морфофизиологические характеристики
Для получения морфофизиологической характеристики, не зависящей от массы животного, и оценки состояния организма и иммунной системы в целом были рассчитаны индексы органов (масса органа/масса тела, ‰). Известно, что изменение содержания в окружающей среде микроэлементов может приводить к существенным изменениям относительной массы внутренних органов в сторону увеличения или уменьшения (Шварц, 1954). Согласно полученным нами результатам, значения относительной массы селезенки, почек, сердца и тимуса у самцов и самок исследованных популяций были сходны, что свидетельствует об отсутствии половых различий в жизнедеятельности основных органов. Относительная масса печени самок Паратунки была ниже, чем у самцов (табл. 6). Одним из показателей токсического воздействия на животных является гонадотропный эффект. Согласно литературным данным, при сравнительно невысоких уровнях токсикантов происходит адаптивное увеличение относительного размера гонад, особенно у самцов, что обеспечивает возможность поддержания численности половозрелых особей в условиях загрязнения (Пескова, 2004). Согласно нашим результатам, индекс семенников самцов популяции ТЭЦ статистически значимо превышал этот показатель у самцов Паратунки (D = 3.26, р = 0.006), что можно объяснить как аллометрической зависимостью величины индекса органа от размеров тела, так и компенсаторной реакцией организма на комплекс загрязнителей водной среды.
Таблица 6.
Морфофизиологические характеристики озерных лягушек исследованных популяций
| Показатель | Индексы органов, ‰ | |||||
|---|---|---|---|---|---|---|
| Печень | Селезенка | Гонады | Почки | Сердце | Тимус | |
| ТЭЦ, самцы (n = 10) | ||||||
| M | 43.69 | 0.89 | 31.27 | 3.78 | 4.06 | 0.04 |
| m | 2.88 | 0.24 | 2.35 | 0.28 | 0.27 | 0.001 |
| ТЭЦ, самки (n = 8) | ||||||
| M | 36.55 | 0.43 | 17.23 | 3.36 | 3.31 | 0.04 |
| m | 1.80 | 0.18 | 2.45 | 0.34 | 0.40 | 0.004 |
| U | 1.82 | 0.93 | 2.88 | 0.66 | 1.64 | 0.3 |
| р | 0.06 | 0.35 | 0.003 | 0.50 | 0.1 | 0.6 |
| Паратунка, самцы (n = 21) | ||||||
| M | 36.23 | 0.24 | 15.02 | 4.03 | 4.31 | 0.03 |
| m | 2.33 | 0.03 | 3.54 | 0.86 | 0.78 | 0.009 |
| Паратунка, самки (n = 10) | ||||||
| M | 29.43 | 0.22 | 12.86 | 3.30 | 3.18 | 0.04 |
| m | 3.56 | 0.07 | 5.05 | 0.38 | 0.46 | 0.007 |
| U | 1.96 | 1.40 | 0.23 | 0.35 | 1.28 | 0.80 |
| р | 0.04 | 0.15 | 0.81 | 0.71 | 0.19 | 0.42 |
Цитогематологические характеристики
Половых различий по содержанию в периферической крови эритроцитов и лейкоцитов у особей исследованных выборок не выявлено (табл. 7). Не было статистически значимых различий и между популяциями.
Таблица 7.
Гематологические характеристики (число клеток крови, тыс. /мм3) озерных лягушек исследованных популяций
| Показатель | ТЭЦ | Паратунка | ||
|---|---|---|---|---|
| Эритроциты, тыс./мм3 | Лейкоциты, тыс./мм3 | Эритроциты, тыс./мм3 | Лейкоциты, тыс./мм3 | |
| Самцы (n = 10) | Самцы (n = 21) | |||
| M | 152.00 | 43.00 | 145.15 | 27.42 |
| m | 50.43 | 11.57 | 33.45 | 5.26 |
| Самки (n = 8) | Самки (n = 10) | |||
| M | 132.00 | 19.25 | 73.77 | 15.55 |
| m | 30.39 | 4.75 | 27.18 | 3.96 |
| U | 0.22 | 1.42 | 1.65 | 1.51 |
| р | 0.82 | 0.15 | 0.09 | 0.12 |
Доля микроядер в эритроцитах крови у особей Паратунки статистически значимо превышала этот показатель у популяции ТЭЦ (z = 5.85, p < < 0.001) (табл. 8). В обеих популяциях преобладали микроядра прикрепленного типа, реже встречались разрыхленные и оформленные (обособленные) микроядра, палочковидные формы микроядер не были обнаружены. Различия между микроядрами, образованными разными типами нарушений генетического материала, были продемонстрированы с помощью меченных зондов (Migliore et al., 1997). Метод позволил выделять прикрепленные микроядра, включающие в себя ацентрические фрагменты (результат хромосомных аберраций) (Жулева, Дубинин, 1994) и оформленные микроядра, представленные целыми хромосомами. Образование оформленных микроядер связывают с дефектами веретена деления, что приводит к изменениям взаимоотношений с ним хромосом и возникновению анеуплоидных клеток (Ковалева, 2008; Eichenlaub-Ritter et al., 1996). Исходя из данных литературы и из собственных результатов, можно полагать, что у озерных лягушек исследованных выборок доля клеток с хромосомными аберрациями превышала долю клеток с вариантами анеуплоидии, возникающей в результате изменений взаимодействия хромосом с веретеном деления. Индукция хромосомных аберраций как один из показателей дестабилизации кариотипа свидетельствовала об активации соматического мутагенеза и о мутагенном воздействии факторов окружающей среды, более выраженном в популяции Паратунки, по сравнению с ТЭЦ.
Таблица 8.
Доля клеток с микроядрами (‰) в периферической крови самцов и самок озерных лягушек исследованных популяций Камчатки
| Популяция, пол (n) |
Доля клеток с микроядрами, ‰ | |||
|---|---|---|---|---|
| оформленными | прикрепленными | разрыхленными | Со всеми микроядрами | |
| ТЭЦ, самки (8) | 0.00037 | 0.0073 | 0 | 0.0077 |
| Паратунка, самки (10) | 0 | 0.0082 | 0.0002 | 0.0084 |
| z | 1.93 | 0.62 | 1.26 | 0.48 |
| p | 0.06 | 0.5 | 0.2 | 0.5 |
| ТЭЦ, самцы (10) | 0.0003 | 0.0034 | 0.0001 | 0.0038 |
| Паратунка, самцы (21) | 0.00004 | 0.011 | 0.00014 | 0.011 |
| z | 1.82 | 7.14 | 0.31 | 6.86 |
| p | 0.1 | <0.001 | 0.5 | <0.001 |
| ТЭЦ (18) | 0.33 | 5.16 | 0.05 | 5.55 |
| Паратунка (31) | 0.03 | 10.48 | 0.16 | 10.67 |
| z | 2.89 | 13.03 | 1.01 | 5.85 |
| р | 0.005 | <0.001 | 0.20 | <0.001 |
В популяции Паратунка выявлены значимые половые различия по содержанию прикрепленных микроядер (z = 2.71, р = 0.01) и общей доли микроядер (z = 2.69, р = 0.01) в эритроцитах крови, свидетельствующие о более высокой частоте мутаций в эритроцитах самцов по сравнению с самками. В популяции ТЭЦ также установлены значимые различия между самками и самцами по доле эритроцитов с микроядрами всех видов (z = 3.54, p = 0.001) и прикрепленных микроядкр (z = 3.69, p = 0.001). Кроме того, выявлено более высокое содержание микроядер всех видов (z = = 6.86, р < 0.001) и отдельно прикрепленных микроядер (z = 7.14, р < 0.001) в крови самцов Паратунки по сравнению с самцами ТЭЦ. Доли эритроцитов с микроядрами в крови самок двух исследованных популяций статистически значимо не различались (табл. 8).
Особый интерес представляла оценка зависимости доли эритроцитов с микроядрами от возраста особей. Сравнительный анализ внутри популяций выявил увеличение доли прикрепленных микроядер с возрастом особей. Так, в выборке озерных лягушек популяции ТЭЦ установлены статистически значимые различия между 4- и 5-летними, а также между 4- и 6-летними особями (табл. 9). В выборке из р. Паратунка случаев значимых различий доли прикрепленных микроядер в возрастных классах оказалось существенно больше (табл. 10). В табл. 9 и 10 приведены доли микроядер у особей разных возрастов и статистически значимые результаты попарного сравнения долей микроядер в эритроцитах озерных лягушек разных возрастов с применением z-критерия.
Таблица 9.
Встречаемость микроядер (‰) в эритроцитах озерных лягушек разных возрастов (самцы и самки вместе) популяции ТЭЦ
| Возраст/число особей | Встречаемость микроядер | |||
|---|---|---|---|---|
| оформленных | прикрепленных | разрыхленных | Всех микроядер | |
| 3/3 | 0.00067 | 0.004 | Не встречались | 0.0046 |
| 4/6 | 0.00016 | 0.003 | 0.00016 | 0.0035 |
| 5/5 | 0.0004 | 0.006 | Не встречались | 0.0068 |
| 6/3 | 0.0003 | 0.008 | Не встречались | 0.0086 |
| z | z4–5 = 2.48, р = 0.01 z4–6 = 3.31, р = 0.001 |
z4–5 = 5.03, р < 0.001 z4–6 = 3.20, р = 0.001 |
||
Таблица 10.
Встречаемость микроядер (‰) в эритроцитах самцов и самок озерных лягушек разных возрастов популяции Паратунки
| Возраст/число особей | Встречаемость микроядер | |||
|---|---|---|---|---|
| оформленных | прикрепленных | разрыхленных | всех микроядер | |
| 1/8 | Не встречались | 0.007 | 0.0002 | 0.0081 |
| 2/7 | Не встречались | 0.005 | 0.0002 | 0.0058 |
| 3/9 | Не встречались | 0.010 | 0.0001 | 0.0102 |
| 4/4 | Не встречались | 0.0142 | Не встречались | 0.0142 |
| 5/2 | 0.0005 | 0.029 | Не встречались | 0.0295 |
| z | z1–4 = 3.30, р < 0.001 | z1–4 = 3.15, р < 0.001 | ||
| z1–5 = 7.72, р < 0.001 | z1–5 = 7.72, р < 0.001 | |||
| z2–3 = 3.17, р = 0.001 | z2–3 = 3.01, р = 0.001 | |||
| z2–4 = 4.70, р < 0.001 | z2–4 = 4.50, р < 0.001 | |||
| z2–5 = 8.94, р < 0.001 | z2–5 = 8.89, р < 0.001 | |||
| z3–4 = 2.05, р = 0.05 | z3–5 = 6.70, р < 0.001 | |||
| z3–5 = 6.61, р < 0.001 | z4–5 = 4.04, р < 0.001 | |||
| z4–5 = 3.92, р < 0.001 | ||||
Обозначения – как в табл. 9.
Между 3-летними особями двух популяций выявлены значимые различия по содержанию прикрепленных микроядер (z = 3.085, р < 0.001) и суммарному содержанию микроядер (z = 2.83, р < < 0.001). При этом доля микроядер у озерных лягушек Паратунки в 2.5 раза превышала этот показатель лягушек ТЭЦ (рис. 3).
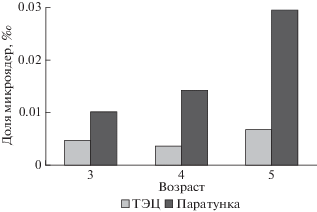
Взаимосвязь цитогематологических характеристик с размерами, возрастом и индексами органов
В качестве меры взаимосвязи исследованных характеристик использовали коэффициент парной гамма-корреляции. Полученные значения выявили положительную взаимосвязь между морфометрическими и морфофизиологическими показателями озерных лягушек исследованных популяций. Так, индекс печени обнаружил умеренную положительную статистически значимую корреляцию с длиной тела (r = 0.35), массой тела (r = 0.37), возрастом особи (r = 0.47), индексом гонад (r = 0.49), индексом сердца (r = 0.45). Умеренная положительная взаимосвязь выявлена между возрастом особи и долей оформленных микроядер. Между возрастом и суммой микроядер в эритроцитах крови обнаружена слабая положительная корреляция (табл. 11).
Таблица 11.
Значения парных коэффициентов гамма-корреляции микроядер, числа эритроцитов и числа лейкоцитов с размерами, возрастом, индексами органов и некоторыми цитогематологическими характеристиками
| Характеристика | Микроядра | Число эритроцитов | Число лейкоцитов | ||
|---|---|---|---|---|---|
| оформленные | прикрепленные | Сумма | |||
| Длина | 0.53 | 0.15 | 0.17 | 0.35 | 0.26 |
| Масса | 0.53 | 0.13 | 0.15 | 0.38 | 0.32 |
| Возраст | 0.67 | 0.21 | 0.23 | 0.30 | 0.20 |
| Индекс печени | 0.47 | 0.28 | 0.30 | 0.24 | 0.16 |
| Индекс гонад | 0.14 | 0.05 | 0.05 | 0.20 | 0.15 |
| Прикрепленные микроядра | –0.03 | – | 0.99 | 0.07 | 0.14 |
| Сумма микроядер | 0.12 | 0.99 | – | 0.06 | 0.12 |
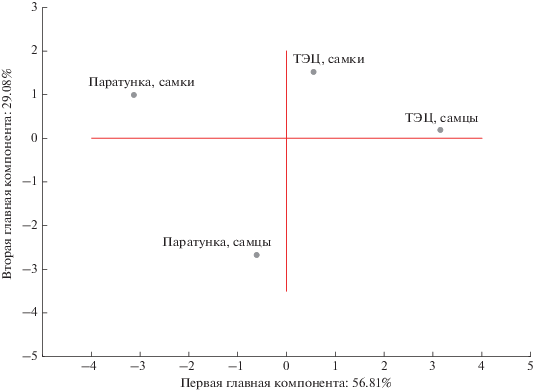
Анализ всех исследованных характеристик методом главных компонент позволил выяснить структуру зависимостей переменных и провести классификацию исследованных выборок. По критерию Кеттеля были выделены две главные компоненты (факторные оси). Первая факторная ось, соответствующая собственному значению 6.81 и описывающая приблизительно 56.81% общей дисперсии, наиболее сильно коррелировала с индексами: печени (0.98), гонад (0.91), селезенки (0.91), числом лейкоцитов (0.85), эритроцитов (0.83) и возрастом особей (0.85). Вторая факторная ось, соответствующая собственному значению 3.49 и описывающая 9.08% общей дисперсии, имела сильную положительную корреляцию с индексом тимуса (0.95), сильные отрицательные корреляции с индексами сердца (–0.85) и почек (–0.82). Сумма микроядер в эритроцитах озерных лягушек обнаружила отрицательную корреляцию с остальными характеристиками и по первой (–0.66) и по второй (–0.59) главным компонентам (рис. 4).
Рис. 4.
Распределение совокупности исследованных характеристик в пространстве 1-й и 2-й главных компонент. Звездочкой отмечены вспомогательные переменные.
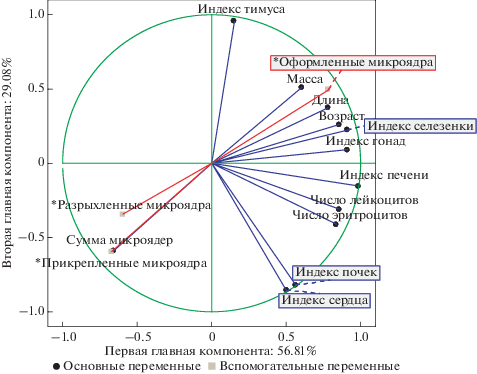
Таким образом, наибольший относительный вклад в общую дисперсию вносили морфофизиологические характеристики. В первую компоненту – индексы гонад (0.120), селезенки (0.123) и печени (0.126). Во вторую компоненту – индексы тимуса (0.23), сердца (0.20) и почек (0.20).
График рассеивания исследованных выборок в пространстве главных компонент позволил сделать заключение, что значения, соответствующие самцам и самкам популяции ТЭЦ, находятся в первом квадранте и наиболее близки друг к другу по исследованным параметрам. На удаленном расстоянии от них расположились выборки самок (второй квадрант) и самцов (третий квадрант) Паратунки (рис. 5). Наибольший вклад в дискриминацию самцов и самок этой популяции вносили цитогенетические аномалии в соматических клетках, оцениваемые микроядерным тестом.
Рис. 5.
Распределение исследованных выборок озерных лягушек в пространстве 1-й и 2-й главных компонент.
Таким образом, цитогенетическим тестом показано преобладание в мутационном спектре прикрепленных микроядер, которые, как считается, образуются в результате структурных аберраций хромосом (Жулева, Дубинин, 1994; Ковалева, 2008). Исследование зависимости частоты встречаемости микроядер от возраста не позволяет получить однозначного ответа, что обусловлено, в первую очередь, отсутствием данных о спонтанно возникающих микроядрах в возрастных группах условно-контрольных фоновых популяций. Тем не менее полученные данные свидетельствуют о выраженной тенденции к возрастанию доли микроядер в старших возрастных группах, что, по всей видимости, обусловлено более длительным воздействием на особей этих групп комплекса мутагенных факторов среды. Различные типы хромосомных аберраций формируются с вовлечением разных морфологических районов хромосом (Ковалева, 2008), что приводит к формированию микроядер разного размера. Озерные лягушки всех возрастных групп популяции ТЭЦ имеют крупные, обособленные и оформленные микроядра, в отличие от особей Паратунки. Это свидетельствует о более серьезной дестабилизации кариотипа, связанной с нарушением процессов расхождения хромосом при митозах. В мутационном спектре озерных лягушек Паратунки преобладают мелкие прикрепленные микроядра, вызванные структурными аберрациями хромосом. Появление таких микроядер может быть следствием их спонтанного формирования в группах одного и того же возраста. Кроме того, необходимо отметить, что не все типы хромосомных аберраций сопровождаются образованием микроядер, поскольку не все микроядра связаны с появлением ацентрических фрагментов хромосом.
Проведенный анализ индукции микроядер в зависимости от возраста особей и получение популяционных характеристик цитогенетического гомеостаза озерных лягушек из популяций термальных водоемов Камчатки позволяет оценить состояние иммунной системы амфибий, которая ответственна за устранение аберрантных клеток и обеспечивает выживание организмов в условиях урбанизации и повышенного загрязнения водной среды.
ВЫВОДЫ
1. Выявлены статистически значимые половые различия по длине и массе тела озерных лягушек двух исследованных популяций термальных водоемов Камчатки. Доля 5-летних самок и доля 4-летних самцов выше в популяции ТЭЦ, по сравнению с долей соответствующих половозрастных групп популяции Паратунки. Соответственно, средние значения длины тела, массы тела и возраста выше у особей популяции ТЭЦ.
2. В популяции Паратунка преобладали морфы striata и hemimaculata; в популяции ТЭЦ – пятнистые (maculata), светлогорлые, светлобрюхие особи.
3. По морфофизиологическим характеристикам (относительной массе органов) выявлено значимое превышение индекса семенников самцов популяции ТЭЦ по сравнению с самцами Паратунки, что можно считать компенсаторной реакцией на комплекс загрязнителей водной среды.
4. Впервые выявлено преобладание эритроцитов с прикреплёнными микроядрами в крови озерных лягушек термальных водоемов Камчатки, свидетельствующее о хромосомных аберрациях, индуцированных мутагенами водной среды. Выявлены половые и возрастные различия по содержанию эритроцитов с микроядрами в выборке Паратунки, свидетельствующие о более высокой частоте мутаций в эритроцитах у самцов по сравнению с самками. Показано возрастание доли микроядер в эритроцитах особей старших возрастов, что, по-видимому, связано с кумулятивным эффектом ксенобиотиков водной среды.
Список литературы
Боркин Л.Я., Тихенко Н.Д., 1979. Некоторые аспекты морфологической изменчивости, полиморфизма окраски, роста, структуры популяции и суточной активности Rana lessonae на северной границе ареала // Ананьева Н.Б. и Боркин Л.Я. (ред.). Экология и систематика амфибий и рептилий. Ленинград. С. 18–54 (Труды Зоологического института АН СССР, т. 89).
Вершинин В.Л., 2004. Гемопоэз бесхвостых амфибий – специфика адаптациогенеза видов в современных экосистемах // Зоологический журнал. Т. 83. № 11. С. 1367–1374.
Вершинин В.Л., 1980. Распределение и видовой состав амфибий городской черты Свердловска // Информационные материалы Института экологии растений и животных. Свердловск. С. 5–6.
Вершинин В.Л., 1982. Городские группировки земноводных как критерий оценки состояния мелких водоемов // Проблемы экологии Прибайкалья. Ч. 1. Иркутск. С. 19–22.
Вершинин В.Л., 1983. Видовой состав и биологические особенности амфибий ряда промышленных городов Урала. Автореф. дис. … канд. биол. наук. Свердловск. 24 с.
Вершинин В.Л., 2008. Морфа striata у представителей рода Rana (Amphibia, Anura) причины адаптивности к изменениям среды // Журнал общей биологии. Т. 69. № 1, С. 65–71.
Гланц С., 1999. Медико-биологическая статистика. М., Практика. 459 с.
Жукова Т.И., Кубанцев Б.С., Бурлаченко Т.Л., 1986. Некоторые реакции популяций озерной лягушки на пестицидное загрязнение водоемов // Антропогенное воздействие на популяции животных. Волгоград. С. 61–81.
Жулева Л.Ю., Дубинин, Н.П., 1994. Использование микроядерного теста для оценки экологической обстановки в районах Астраханской области // Генетика. Т. 30. № 7. С. 999–1004.
Ивантер Э.В., Ивантер Т.В., Туманов И.Л., 1985. Адаптивные особенности мелких млекопитающих. Л.: Наука. 320с.
Ищенко В.Г., 1978. Динамический полиморфизм бурых лягушек фауны СССР. М.: Наука, 148 с.
Кармазин А.П., Пескова Т.Ю., 2010. Использование гематологических показателей озерной лягушки Rana ridibunda (Pallas, 1771) для определения зоны токсического действия нефти // Современная герпетология. Т. 10. Вып. 1/2. С. 3–7.
Клевезаль Г.С., Смирина Э.М., 2016. Регистрирующие структуры наземных позвоночных. Краткая история и современное состояние исследований // Зоологический журнал. Т. 95. № 8. С. 872–896.
Ковалева О.А., 2008. Цитогенетические аномалии в соматических клетках // Цитология и генетика. № 1. С. 58–72.
Лада Г.А., 2012. Бесхвостые земноводные (Anura) Русской равнины: изменчивость, видообразование, ареалы, проблемы охраны. Дис. … докт. биол. наук. Казань. 424 с.
Ляпков С.М., 2014. Озерная лягушка (Pelophylax ridibundus) в термальных водоемах Камчатки // Зоологический журнал. Т. 93. № 12. С. 1427–1432.
Ляпков С.М., 2016. Места находок и состояние популяций озерной лягушки на Камчатке // Вестник Тамбовского университета. Серия: Естественные и технические науки. Т. 21. № 5. С. 1821–1824.
Ляпков С.М., 2016а. Озерная лягушка (Pelophylax ridibundus) на Камчатке: особенности местообитаний, размерного и возрастного состава популяций // “Сохранение биоразнообразия Камчатки и прилегающих морей”. Тезисы ХVII международной научной конференции. Петропавловск-Камчатский. С. 94–98.
Ляпков С.М., Ермаков О.В., Титов С.В., 2017. Распространение и происхождение двух форм озерной лягушки Pelophylax ridibundus complex (Anura, Ranidae) на Камчатке по данным анализа митохондриальной и ядерной ДНК // Зоологический журнал. Т. 96. № 11. С. 1384–1391.
Меньшиков В.В., Делекторская Л.Н., Золотницкая Р.П., 1987. Лабораторные методы исследования в клинике М.: Медицина, 368 с.
Нормативы качества воды водных объектов рыбохозяйственного значения, в том числе нормативы предельно допустимых концентраций вредных веществ в водах водных объектах рыбохозяйственного значения (утв. приказом Федерального агентства по рыболовству от 18.01.2010 г. № 20) Информационно-правой портал: Гарант.ру. [Электронный ресурс]. Режим доступа: http://www.garant.ru/products/ipo/prime/doc/2070984/#ixzz4mY1P6WRm Дата обращения: 07.07.2019.
Пескова Т.Ю., 1995. Краснобрюхая жерлянка (Bombina bombina L.) как тест объект биоиндикации пестицидного загрязнения водоемов в западном Предкавказье. Автореф. дис. … канд. биол. наук. Краснодар. 18 с.
Пескова Т.Ю., 2004. Адаптационная изменчивость земноводных в антропогенно загрязненной среде. Автореф. дис. … докт. биол. наук. Тольятти. 36 с.
Реброва О.Ю., 2006. Статистический анализ медицинских данных. М.: МедиаСфера. 312 с.
Романова Е.Б., Шаповалова К.В., Рябинина Е.С., 2018. Лейкоцитарный состав крови и микроядра в эритроцитах амфибий загрязненных водных объектов Нижегородской области // Принципы экологии. № 2. С. 125–139.
Романова Е.Б., Шаповалова К.В., Рябинина Е.С., Гелашвили Д.Б., 2018а. Лейкоцитарные индексы и микроядра в эритроцитах как популяционные маркеры иммунного статуса Pelophylax ridibundus, Pallas 1771 (Amphibia: Ranidae), обитающих в различных биотопических условиях // Поволжский экологический журнал. № 1. С. 60–75.
Рябчикова Т.Н., Дробот Г.П., Свинин А.О., Ведерников А.А., Сидушкина М.Н., 2019. Оценка цитогенетического гомеостаза озерной лягушки из водоемов заповедника “Приволжская лесостепь” с использованием микроядерного теста / Биосистемы: организация, поведение, управление: Тезисы докладов 72-й Всероссийской с международным участием школы-конференции молодых ученых (Н.Новгород, 23–26 апреля 2019 г.). Н. Новгород, Университет им. Н.И. Лобачевского. С. 194.
Смирина Э.М., 1983. Прижизненное определение возраста и ретроспективная оценка размеров тела серой жабы (Bufo bufo) // Зоологический журнал. Т. 62. № 3. С. 437–444.
Спирина Е.В., Романова Е.М., Спирина Т.М., 2011. Оценка стабильности развития и цитогенетического гомеостаза в популяциях Rana ridibunda Pall. Ульяновской области // Известия Самарского научного центра РАН. Т. 13. № 1. С. 123–128.
Терентьев П.В., 1962. Характер географической изменчивости зеленых лягушек // Труды Петергофского биологического института Ленинградского университета. № 19. С. 98–121.
Файзулин А.И., Кузовенко А.Е., 2012. Использование амфибий в мониторинге состояния окружающей среды в условиях Самарской области: фенетическая структура популяций // Известия Самарского научного центра РАН. Т. 1 (3). № 1. С. 829–833.
Файзулин А.И., Чихляев Н.В., Кузовенко А.Е., 2013. Амфибии Самарской области. Тольятти: ООО “Кассандра”. 140 с.
Фоминых А.С., Ляпков С.М., 2011. Формирование новых особенностей жизненного цикла озерной лягушки (Rana ridibunda) в условиях подогреваемого водоема // Журнал общей биологии. Т. 72. № 6. С. 403–421.
Шварц С.С., 1954. Влияние микроэлементов на животных в естественных условиях рудного поля // Труды биогеохимической лаборатории АН СССР. Т. 10. С. 76–81.
Шляхтин Г.В., 1985. Фенетический анализ окраски спины озерной лягушки // Фенетика популяций. М.: Наука. С. 173.
Щупак Е.Л., Ищенко В.Г., 1981. К генетике полиморфизма окраски у остромордой лягушки (Rana arvalis Nilss.). I. Светлая дорсо-медиальная полоса // Боркин Л.Я. (ред.). Герпетологические исследования в Сибири и на Дальнем Востоке. Ленинград: Зоологический институт Академии наук СССР. С. 128–132.
Berger L., Smielowski J., 1982. Inheritance of vertebral stripe in Rana ridibunda Pall. (Amphibia, Ranidae) // Amphibia-Reptilia. V. 3. № 2/3. P. 145–151.
Eichenlaub-Ritter U., Baart E.,Yin H., Betzendahl I., 1996. Mechanisms of spontaneous and chemically-induced aneuploidy in mammalian oogenesis: basis of sex-specific differences in response to aneugens and the r-r necessity for further tests // Mutation Research. V. 372. № 2. P. 279–294.
Hayashi M.L., MacGregor J.T, Gatehouse D.G., Adler I.D., 2000. In vivo rodent erythrocyte micronucleus assay // Environmental and Molecular Mutagenesis. V. 35 (3). P. 234–252.
Marunouchi J., Kusano T., Ueda H., 2000. Validity of back-calculation methods of body size from phalangeal bones: an assessment using data for Rana japonica // Current Herpetology. V. 19. P. 81–89.
Migliore L., Barale R., Belluomini D., 1987. Cytogenetic damage induced in human lymphocytes by adriamycin and vincristine: A comparison between micronucleus and chromosomal aberration assays // Toxicology in Vitro. V. 1. № 4. P. 247–254.
Romanova E.B., Ryabinina E.S., 2018. Cytogenetic Approach to Primary Assessment of Habitat Quality // Opera Medica et Physiologica. V. 4. № 1. P. 19–22.
Smirina E.M., 1994. Age determination and longevity in amphibians // Gerontology. V. 40. P. 133–146.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал