

Зоологический журнал, 2020, T. 99, № 9, стр. 1047-1052
Использование деревьев лесными полевками (Rodentia, Cricetidae) в европейской северной тайге
А. А. Калинин *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: benguan@yandex.ru
Поступила в редакцию 27.07.2019
После доработки 22.10.2019
Принята к публикации 16.01.2020
Аннотация
Древесную активность мелких млекопитающих изучали в северной тайге на западном макросклоне Северного Урала. Работы проводили в августе. За три года было помечено 279 млекопитающих 7 видов в живоловках, расположенных на уровне земли и на стволах деревьев на высоте 1.5–2 м. На деревьях зарегистрировано 2 особи рыжей полевки (Myodes glareolus) и 51 особь красной полевки (M. rutilus). Деревья используют преимущественно оседлые зверьки в пределах своего домашнего участка. Уловистость полевок на деревьях достоверно связана с плотностью вида в наземном ярусе. Использование деревьев красной полевкой не зависело ни от вида деревьев, ни от толщины ствола. Половозрастной состав полевок, зарегистрированных на деревьях, не отличался от состава полевок, отмеченных в наземном ярусе. После ночного дождя уловистость полевок в верхних ловушках значительно возрастала. Посещение деревьев полевками может быть связано с тем, что эти зверьки питаются древесными лишайниками. В лесах, где наблюдается значительное обрастание деревьев лишайниками, можно ожидать массового использования этих деревьев лесными полевками.
Полевки рода Myodes могут активно использовать деревья. Это хорошо известно для европейской рыжей полевки (Myodes glareolus Schreber 1780), которую неоднократно отлавливали в ловушках, установленных на деревьях (Holisova, 1969; Montgomery, 1980; Tattersall, Whitbread, 1994; Buesching et al., 2008; Бобрецов и др., 2019), обнаруживали в искусственных гнездовьях и гнездах птиц (Формозов, 1948; Juškaitis, 1999), в ряде работ исследовали способности этого вида к лазанью экспериментально (Trebatická et al., 2008; Karantanis et al., 2017). Известна способность к лазанию и для других представителей рода, например полевки Гаппера (M. gapperi Vigors 1830) отлавливались на высоте до 5 м (Getz, Ginsberg, 1968), а калифорнийские рыжие полевки (M. californicus Merriam 1890) отмечались на высоте 10.8 м (Swingle, 2005). Красные полевки (M. rutilus Pallas 1779) так же отлавливались на деревьях на высоте 1.5–2 м в штате Аляска (Nations, Olson, 2015) и в верхнепечорской тайге (Бобрецов и др., 2007; 2019), в Восточной Сибири этих полевок отмечали на вершинах кедров (Реймерс, Воронов, 1963). Хотя лесные полевки могут посещать деревья, лазают они хуже, а на деревьях отмечаются реже, чем некоторые другие виды, относящиеся к наземным мелким млекопитающим. В Европе при сравнительных исследованиях показано, что лесные (Apodemus uralensis Pallas 1811) и, особенно, желтогорлые мыши (A. flavicollis Melchior 1834) чаще используют деревья (Holisova, 1969; Montgomery, 1980; Buesching et al., 2008) и лучше приспособлены к лазанию (Karantanis et al., 2017). В Северной Америке чаще использует деревья белоногий хомячок (Peromyscus leucopus Rafinesque 1818) (Getz, Ginsberg, 1968).
В некоторых работах предполагается, что полевки залезают на деревья, спасаясь от хищников. Действительно, в экспериментах показано, что в присутствии ласки (Mustela nivalis Linnaeus 1766) рыжие полевки больше времени проводили в верхнем ярусе (Trebatická, 2008). Но в природе наблюдали только желтогорлых мышей, которые при нападении ласки взбирались на деревья, в то время как у рыжих полевок такого поведения отмечено не было (Jędrzejewski et al., 1992). Считается, что использование древесного яруса позволяет уменьшить конкуренцию между видами и в результате увеличить перекрытие в горизонтальной плоскости (Лукьянова, Бобрецов, 2008; Albanese et al., 2011). Способность к лазанью позволяет получить доступ к дополнительным ресурсам, в частности к древесным лишайникам (Nations, Olson, 2015). Лишайники занимают значительное место в рационе лесных полевок. Например, у полевки Гаппера рацион на 90% состоит из грибов и лишайников, причем лишайники употребляют чаще зимой (Dubay et al., 2008). Лишайники поедаются рыжими полевками (Формозов, 1948; Ивантер, 1975) и красными полевками (Вольперт, Шадрина, 2012; Докучаев, 2012; Ивантер, Моисеева, 2015; Nations, Olson 2015), причем у красных полевок древесные лишайники рода Usnea могут составлять основу питания: у 57% красных полевок лишайники составляли свыше 70% содержимого желудка (Докучаев, 2009).
Таким образом, хотя сам факт использования деревьев лесными полевками рода Myodes известен, крайне мало работ, позволяющих количественно оценить роль древесного яруса в экологии этих видов.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили в августе в течение трех лет на западном макросклоне Северного Урала в долине р. Илыч в предгорном районе Печоро-Илычского государственного природного биосферного заповедника (62.6° с.ш., 58.9° в.д.), который относится к северной тайге камско-печорско-западноуральских темнохвойных лесов (Карпенко, 1980). Участок расположен в пихтово-еловом зеленомошно-разнотравном лесу со значительной примесью кедра.
Отловы проводили трапиковыми беспружинными живоловками (Щипанов, 1986), которые позволяют отлавливать весь комплекс видов мелких млекопитающих, включая насекомоядных (Щипанов и др., 2008). В качестве приманки использовали по несколько зерен крупы геркулес, смоченных нерафинированным растительным маслом. На земле живоловки расставляли в линию, с расстоянием между ловушками 7.5 м. Проверки проводили в светлое время суток через 1.5 ч, два раза подряд, затем ловушки оставляли до следующего дня ненастороженными, доступными для посещения животными (Щипанов и др., 2000). Учетная линия на земле состояла из 100 ловушек, общая протяженность 750 м. Около каждой нечетной ловушки на стволе дерева на высоте 1.5–2 м закреплялась аналогичная живоловка (рис. 1). Таким образом линия учетов на деревьях состояла из 50 живоловок, расставленных через 15 м. Проверки на деревьях проводили 2 раза в сутки, ловушки не закрывали.

Всего за время учетов отмечено 279 млекопитающих 7 видов, давших 1127 регистраций. Общий объем материала приведен в табл. 1.
Таблица 1.
Количество особей разных видов, отмеченных на уровне земли и на деревьях
| Вид | 2005 | 2006 | 2007 | |||
|---|---|---|---|---|---|---|
| Земля | Деревья | Земля | Деревья | Земля | Деревья | |
| Myodes glareolus | 7 | 2 | 0 | 0 | 0 | 0 |
| M. rutilus | 83 | 26 | 99 | 25 | 2 | 0 |
| Microtus agrestis | 0 | 0 | 1 | 0 | 0 | 0 |
| Sorex araneus | 1 | 0 | 21 | 0 | 20 | 0 |
| S. caecutiens | 3 | 0 | 20 | 0 | 17 | 0 |
| S. minutus | 1 | 0 | 3 | 0 | 0 | 0 |
| S. tundrensis | 0 | 0 | 1 | 0 | 0 | 0 |
Расчет показателей плотности оседлого населения (особей/га) при мечении на линиях живоловок проводили по описанной ранее методике (Калинин, 2012). Получаемые на линиях данные хорошо соответствуют результатам учетов на больших площадках (Калинин и др., 2018).
Сравнение двух независимых групп проводили Манн–Уитни тестом (U – Mann–Whitney U-test), несколько независимых выборок сравнивали критерием Крускал-Уоллиса (H – Kruskal-Wallis test), парные линейные связи оценивали по коэффициенту корреляции Спирмана (rs – Spearman rank order correlation). Соотношение долей оценивали по критерию Пирсона (χ2 – Pearson Chi-square test). Значение среднего приводится с показателями стандартного отклонения (Mean ± SD).
РЕЗУЛЬТАТЫ
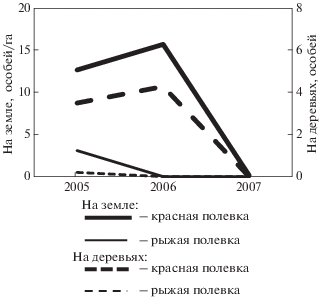
За все время проведения учетов в ловушках, установленных на деревьях, отмечены 51 особь красной полевки и 2 особи рыжей полевки. Уловистость полевок на деревьях с высокой достоверностью связана с плотностью вида, рассчитанной по данным учетов на уровне земли (рис. 2), коэффициент корреляции Спирмана rs = 0.95, p = = 0.003. В годы, когда численность была минимальной (для красной полевки это 2007, а для рыжей полевки – 2006 и 2007), лесных полевок вообще не наблюдали в учетах на деревьях. Такая синхронность в изменении численности животных на деревьях и на земле отмечалась и другими исследователями (Бобрецов и др., 2007; Buesching et al., 2008), при этом в местах совместного обитания красных и рыжих полевок наблюдалась явная избирательность в посещении деревьев разными видами. Доля поимок на них красной полевки по сравнению с долей поимок рыжей была выше, чем на земле (Лукьянова, Бобрецов, 2008; Бобрецов и др., 2019), что может говорить о большей склонности красных полевок к использованию верхних ярусов.
Рис. 2.
Плотность оседлых красных и рыжих полевок в учетах на земле и количество поимок на деревьях.
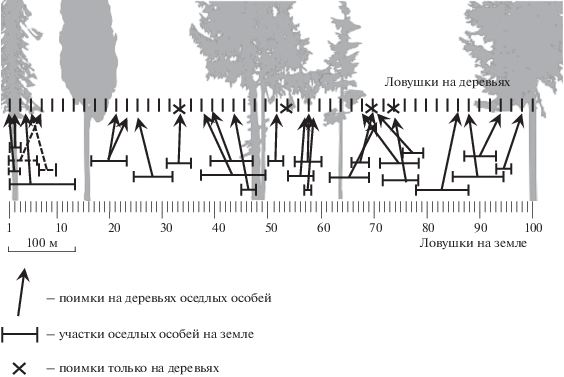
При проведении учетов в живоловках оседлые зверьки дают неоднократные регистрации, что позволяет выделить их индивидуальные участки. Простейший способ выделения границ индивидуальных участков зверьков на линиях мечения – выделение участков по крайним точкам. Необходимо учитывать, что такой способ расчета значительно снижает реальную величину использования пространства оседлыми животными, особенно при небольшом числе регистраций (Щипанов и др., 2000). Большинство полевок, отмеченных на деревьях, ловились либо на территории своих наземных участков, либо в непосредственной близости от них (рис. 3). Можно предположить, что деревья входят в структуру участка и регулярно посещаются оседлыми особями.
Рис. 3.
Индивидуальные участки красных (сплошная линия) и рыжих (пунктир) полевок на земле и посещение ими деревьев в 2005 г.
При общей низкой численности рыжих полевок в районе учетной линии и на деревьях их было отловлено крайне мало. Достаточный материал для статистической обработки получен только для красных полевок (табл. 2). Среди красных полевок, отлавливаемых на деревьях, 68.6% особей отлавливали так же и на земле, причем большинство из них (91.4%) дали на земле 2 и более регистрации, что позволяет отнести их к оседлым особям.
Таблица 2.
Количество особей красной полевки, отмеченных на уровне земли и на деревьях
| Год | На земле | На деревьях | ||||
|---|---|---|---|---|---|---|
| всего особей | с 2 и более поимками | всего особей | из них на земле | из них на земле с 2 и более поимками | только на деревьях | |
| 2005 | 83 | 52 | 26 | 22 | 21 | 4 |
| 2006 | 99 | 60 | 25 | 13 | 11 | 12 |
| 2007 | 2 | 1 | 0 | 0 | 0 | 0 |
| Всего | 184 | 113 | 51 | 35 | 32 | 16 |
При сравнении выборок, регистрируемых на земле и на деревьях, показано, что соотношение полов красных полевок, оцененное по критерию Пирсона, в этих выборках не различался: χ2 = = 2.59, df = 1, p = 0.11. Самцы и самки этого вида одинаково используют деревья, всего в верхних ловушках отмечены 21 самец и 30 самок.
Деревья посещают все возрастные группы полевок. При мечении в живоловки можно уверено выделить 3 группы: дети (juv), молодые (sad) и взрослые (ad). Всего на деревьях зарегистрированы 4 детеныша, 10 взрослых и 37 молодых особей красной полевки, что не отличается по составу от отловленных на уровне земли: χ2 = 2.85, df = 2, p = 0.24.
В верхнем ярусе ловушки устанавливали на различных породах деревьев (ель, пихта, береза) с диаметром стволов 8–35 см. Использование деревьев красной полевкой, оцененное по общему количеству регистраций, не зависело ни от вида деревьев (Критерий Крускал-Уоллиса H = 1.59, N = 50, p = 0.45), ни от толщины ствола (коэффициент корреляции Спирмана rs = 0.05, p = 0.75).
На результативность учетов верхними ловушками значительное влияние оказывала погода. При наличии ночного дождя уловистость полевок в верхних ловушках значительно возрастала. Использованы данные только за 2005 и 2006 годы, когда были поимки в верхних ловушках. В дождливые ночи (n=6) уловистость составила 8.33 ± ± 3.6 поимок за сутки на 50 ловушек, а в сухие (n = 11) 1.54 ± 1.7. Различия в Манн-Уитни тесте достоверны U = 3.12, p = 0.002. Нужно отметить, что возрастание активности полевок в древесном ярусе не связана с возрастанием общей активности. В эти же дни количество регистраций полевок на земле не различалось (соответственно 34.4 ± 5.9 и 37.9 ± 10.7 регистраций на 100 ловушек).
ОБСУЖДЕНИЕ
Таким образом, лесные полевки, в частности красная полевка, могут активно использовать верхние ярусы леса, поднимаясь по стволам деревьев. В ряде случаев полевки могут устраивать убежища на деревьях, в том числе используя гнезда птиц или искусственные гнездовья (Формозов, 1948; Juškaitis, 1999). Полевки посещают различные виды деревьев, при этом в верхнем ярусе отмечены все половозрастные группы. По всей видимости использование деревьев связано с обилием древесных лишайников, в том числе рода Usnea, которые широко распространены в данном регионе. Лишайники являются одним из основных компонентов зимнего питания красных полевок, хотя поедаются ими и в другие сезоны (Докучаев, 2009; Nations, Olson, 2015). При этом как рыжие, так и красные полевки могут делать запасы лишайников в виде аккуратно скрученных шариков (Формозов, 1948; Докучаев, 2012). Красных полевок ранней весной и зимой неоднократно наблюдали кормящимися лишайниками на стволах деревьев на высоте 0.3–2 м или встречали следы такой деятельности (Бобрецов и др., 2004; 2019). Выраженная лихенофагия лесных полевок приводит к активному использованию ими деревьев при наличии на них значительного количества лишайников. Увеличение активности полевок на деревьях после дождей может быть связано с повышением привлекательности переувлажненных лишайников в качестве пищевого объекта.
При проведении учетов на деревьях нами не было зарегистрировано никаких других видов мелких млекопитающих, кроме лесных полевок. В некоторых исследованиях отмечается присутствие в древесном ярусе землероек-бурозубок (Atkins et al., 2018). При этом на деревьях обычно отмечают малую бурозубку (Sorex minutus L. 1766) (Holisova, 1969; Juškaitis, 1999; Stepánková, Vohralík, 2009). Мы также наблюдали в Тверской обл. сеголетка малой бурозубки, который после выпуска из живоловки ушел на ствол сосны, свободно перемещался по стволу, обследовал трещины коры, отыскал и съел имаго комара (сем. Tipulidae), в течение 10 мин поднялся на высоту 5 м, где был потерян из виду. Средняя бурозубка (Sorex caecutiens Laxmann 1788) в небольшом количестве была отмечена при отловах на деревьях в предгорной тайге Северного Урала (Бобрецов и др., 2019). В данном эксперименте в учетах на земле было зарегистрировано 87 особей землероек бурозубок 4 видов, при этом на деревьях они не отмечены.
Использование деревьев наземными мелкими млекопитающими, в частности лесными полевками, может иметь региональные особенности. У красных полевок питание древесными лишайниками в основном значимо для северных регионов, особенно в Заполярье (Докучаев, 2009). Для этих регионов характерно обильное обрастание деревьев эпифитными лишайниками, и именно здесь можно ожидать наиболее высокую древесную активность красных полевок.
Список литературы
Бобрецов А.В., Лукьянова Л.Е., Сокольский С.М., 2007. Использование лесными полевками верхних ярусов леса в верхнепечорской тайге // Териофауна России и сопредельных территорий (VIII съезд Териологического общества). М.: Товарищество научных изданий КМК. С. 48.
Бобрецов А.В., Нейфельд Н.Д., Сокольский С.М., Теплов В.В., Теплова В.П., 2004. Млекопитающие Печоро-Илычского заповедника. Сыктывкар: Коми книжное издательство. 464 с.
Бобрецов А.В., Петров А.Н., Быховец Н.М., 2019. Мелкие млекопитающие на деревьях: результаты учетов животных в предгорной тайге Северного Урала // Вестник ИБ Коми НЦ УрО РАН. № 2. С. 10–15.
Вольперт Я.Л., Шадрина Е.Г., 2012. Мелкие млекопитающие северо-востока Сибири. Новосибирск: Наука. 246 с.
Докучаев Н.Е., 2009. О лихенофагии красной полевки (Clethrionomys rutilus) // Зоологический журнал. Т. 88. № 8. С. 1013–1016.
Докучаев Н.Е., 2012. Запасливые полевки // Природа. № 8. С. 63–66.
Ивантер И.В., 1975. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука. 246 с.
Ивантер Э.В., Моисеева Е.А., 2015. К экологии красной полевки (Clethrionomys rutilus Pall.) на юго-западной периферии ареала // Труды Карельского научного центра РАН. № 1. С. 37–47.
Калинин А.А., 2012. Оседлая и нерезидентная составляющая численности массовых видов мелких млекопитающих по данным учета на линиях живоловок // Зоологический журнал. Т. 91. № 6. С. 759–768.
Калинин А.А., Олейниченко В.Ю., Купцов А.В., Александров Д.Ю., Демидова Т.Б., 2018. Пространственная изменчивость оседлого и нерезидентного населения мелких млекопитающих в лесном массиве // Зоологический журнал. Т. 97. № 4. С. 474–485 (Kalinin A.A., Oleinichenko V.Y., Kouptsov A.V., Aleksandrov D.Y., Demidova T.B., 2018. Spatial variability of resident and nonresident populations of small mammals on a forest plot // Biology Bulletin. V. 45. № 9. P. 1012–1022).
Карпенко А.С., 1980. Камско-печорско-западноуральские темнохвойные леса // Растительность Европейской части СССР. Л.: Наука. С. 93–101.
Лукьянова Л.Е., Бобрецов А.В., 2008. Локальное распределение численности симпатрических видов лесных полевок в микросредовых условиях дестабилизированных и стабильных местообитаний // Успехи современной биологии. Т. 128. № 5. С. 517–528.
Реймерс Н.Ф., Воронов Г.А., 1963. Насекомоядные и грызуны Верхней Лены. Иркутск: Иркутское кн. изд-во. 192 с.
Формозов А.Н., 1948. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930-1940 гг. // Фауна и экология грызунов. М.: Московское общество испытателей природы. Вып. 3. С. 3–110.
Щипанов Н.А., 1986. К экологии малой белозубки (Crocidura suaveolens) // Зоологический журнал. Т. 66. № 7. С. 1051–1060.
Щипанов Н.А., Калинин А.А., Олейниченко В.Ю., Демидова Т.Б., Гончарова О.Б., Нагорнев Ф.В., 2000. К методике изучения использования пространства землеройками-бурозубками // Зоологический журнал. Т. 79. № 3. С. 362–371.
Щипанов Н.А., Литвинов Ю.Н., Шефтель Б.И., 2008. Экспресс-метод оценки локального биологического разнообразия сообщества мелких млекопитающих // Сибирский экологический журнал. Т. 15. № 5. С. 783–791 (Shchipanov N.A., Litvinov Y.N., Sheftel B.I., 2008. Rapid method for estimating local biodiversity of a community of small mammals // Contemporary Problems of Ecology. V. 1. № 5. P. 596–602).
Albanese S., Rodríguez D., Ojeda R.A., 2011. Differential use of vertical space by small mammals in the Monte Desert, Argentina // Journal of Mammalogy. V. 92. № 6. P. 1270–1277.
Atkins E., Swetnam R., Mitchell P., Dover J., 2018. Surveying small mammals in urban hedges // Mammal Communications. № 4. P. 7–15.
Buesching, C.D., Newman C., Twell R., Macdonald D.W., 2008. Reasons for arboreality in wood mice Apodemus sylvaticus and bank voles Myodes glareolus // Mammalian Biology. V. 73. № 4. P. 318–324.
Dubay S.A., Hayward G.D., Martinez del Rio C., 2008. Nutritional value and diet preference of arboreal lichens and hypogeous fungi for small mammals in the Rocky Mountains // Canadian Journal of Zoology. V. 86. № 8. P. 851–862.
Getz L.L., Ginsberg V., 1968. Arboreal behaviour of the red-back vole, Clethrionomys gapperi // Animal Behaviour. V. 16. № 4. P. 418–424.
Holisova V., 1969. Vertical movements of some small mammals in a forest // Zoologické listy. V. 18. P. 121–141.
Jędrzejewski W., Jędrzejewska B., McNeish E., 1992. Hunting success of the weasel Mustela nivalis and escape tactics of forest rodents in Białowieża National Park // Acta Theriologica. V. 37. № 3. P. 319–328.
Juškaitis R., 1999. Mammals occupying nestboxes for birds in Lithuania // Acta Zoologica Lituanica. V. 9. № 3. P. 19–23.
Karantanis N.E., Rychlik L., Herrel A., Youlatos D., 2017. Arboreal gaits in three sympatric rodents Apodemus agrarius, Apodemus flavicollis (Rodentia, Muridae) and Myodes glareolus (Rodentia, Cricetidae) // Mammalian Biology. V. 83. P. 51–63.
Montgomery W.I., 1980. The use of arboreal runways by the woodland rodents, Apodemus sylvaticus (L.), A. flavicollis (Melchior) and Clethrionomys glareolus (Schreber) // Mammal Review. V. 10. № 4. P. 189–195.
Nations J.A., Olson L.E., 2015. Climbing behavior of northern red-backed voles (Myodes rutilus) and scansoriality in Myodes (Rodentia, Cricetidae) // Journal of Mammalogy. V. 96. № 5. P. 957–963.
Stepánková J., Vohralík V., 2009. Vertical activity of the yellow-necked mouse (Rodentia: Muridae) at edge of a mixed forest // Folia Zoologica. V. 58. № 1. P. 26–35.
Swingle J.K., 2005.Daily activity patterns, survival, and movements of red tree voles (Arborimus longicaudus) in western Oregon. M.S. thesis, Oregon State University, Corvallis. 118 p.
Tattersall F., Whitbread S., 1994. A trap-based comparison of the use of arboreal vegetation by populations of bank vole (Clethrionomys glareolus), woodmouse (Apodemus sylvaticus) and common dormouse (Muscardinus avellanarius) // Journal of Zoology. V. 233. № 2. P. 309–314.
Trebatická L., Sundell J., Tkadlec E., Ylönen H., 2008. Behaviour and resource use of two competing vole species under shared predation risk // Oecologia. V. 157. № 4. P. 707–715.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал