

Зоологический журнал, 2021, T. 100, № 1, стр. 104-114
Внутривидовая изменчивость формы строения третьего нижнего предкоренного зуба северной пищухи (Ochotona hyperborea (Pallas 1811))
А. Е. Гусев a, *, М. П. Тиунов a
a Федеральный научный центр биоразнообразия наземной биоты
Восточной Азии ДВО РАН
690022 Владивосток, Россия
* E-mail: gusevmamoru@gmail.com
Поступила в редакцию 31.05.2019
После доработки 03.07.2019
Принята к публикации 19.12.2019
Аннотация
Проанализирована изменчивость формы жевательной поверхности третьего нижнего предкоренного зуба (p3) у современных представителей северной пищухи (Ochotona hyperborea). Выявлена изменчивость трех основных признаков, характеризующих этот зуб: форма перешейка, соединяющего антероконид и постероконид, форма антероконида и форма постероконида. Построена комбинативная решетка, показывающая распределение морфотипов жевательной поверхности p3 на основании различающих их признаков. На севере и юге ареала доминируют особи с различными морфотипами жевательной поверхности p3.
Северная пищуха Ochotona hyperborea (Pallas 1811) относится к одному из наиболее широко распространенных видов Северной Азии. Ее обитание связано преимущественно с каменистыми россыпями на открытых и покрытых лесом и кустарником горных склонах, побережьях рек и морском побережье. Широкое распространение этого вида естественно привлекало исследователей к изучению различных аспектов его изменчивости. Проводились изучение географической изменчивости черепа (Lissovsky, 2003), молекулярно-генетические (Lissovsky, 2014), кариологические (Воронцов, Иваницкая, 1973; Картавцева и др., 2014) и биоакустические исследования (Kawamichi, 1981; Lissovsky, Lissovskaya, 2002; Лисовский, 2005). Отдельный интерес представляет изучение внутривидовой изменчивости третьего нижнего предкоренного зуба p3, наиболее часто используемого при идентификации ископаемых пищух, остатки которых зачастую представлены только отдельными зубами. Проведенное недавно изучение изменчивости методом геометрической морфометрии формы антероконида и передних складок (пара- и протофлексид) у 4 современных видов пищух показало отсутствие значимых различий между исследуемыми видами по форме этих структур, что связывается с их значительной внутривидовой изменчивостью (Volkova, Lissovsky, 2018). В то же время, в этой работе высокая внутривидовая изменчивость только фиксируется, а не рассматривается все пространство морфологических состояний формы третьего нижнего предкоренного зуба у исследуемых видов. Работы по изучению внутривидовой изменчивости формы этого зуба отсутствуют, что значительно осложняет как проведение диагностики ископаемых пищух, так и сравнительное изучение его морфотипической изменчивости во времени. Количество находок северной пищухи в ископаемом состоянии увеличивается (Лисовский, Сердюк, 2004; Панасенко, Тиунов, 2010; Калмыков, 2015; и др.), поэтому возникает настоятельная необходимость проведения анализа всего морфопространства формы жевательной поверхности третьего нижнего предкоренного зуба, прежде всего у современных представителей этого вида.
МАТЕРИАЛЫ И МЕТОДЫ
Изучение третьего нижнего предкоренного зуба (p3) северной пищухи проводилось на коллекционных материалах лаборатории териологии ФНЦ Биоразнообразия ДВО РАН и Зоологического музея МГУ.
В целом, было просмотрено 183 третьих нижних премоляров северной пищухи из 10 локалитетов.
Исследование проводилось в несколько этапов. На первом этапе рассчитывалось распределение размерных характеристик зуба. Затем выделялись признаки, с помощью которых можно охарактеризовать морфотипическую изменчивость жевательной поверхности исследуемого зуба. Для различных состояний этих признаков рассчитывалась их частота встречаемости. На последнем этапе проводились сравнения и анализ наиболее часто встречающихся морфотипов.
В работе использовался один из методов многомерной статистики, а именно последовательный дискриминантный анализ. Последовательность анализа заключалась в объединении исследуемых выборок при помощи функции матричной классификации и расстояния Махаланобиса. Для вычисления достоверности расстояний Махаланобиса было принято использовать значение уровня значимости p-level, равное 0.01, так как объем выборки n > 100 (Наследов, 2004). Для вычисления достоверности различий между выборками по размерным характеристикам предкоренных зубов был рассчитан критерий Стьюдента. Так как статистические различия между самцами и самками отсутствуют (Лисовский, Сердюк, 2004; Lissovsky, 2014; Volkova, Lissovsky, 2018), выборки анализировали без разделения по полу. Для выполнения расчетов использовалось программное обеспечение Statistica 13.
В данной работе использовались девять промеров (рис. 1): 1 – ширина зуба, 2 – длина зуба, 3 – расстояние между наиболее назальной точкой переднего сегмента и наиболее глубоко расположенной точкой буккальной петли, 4 – ширина перешейка между передним и задним сегментами, 5 – ширина буккальной части переднего сегмента – расстояние между наиболее назальной и буккальной точками переднего сегмента, 6 – расстояние между наиболее буккальной точкой второго сегмента и самой выпуклой частью с лингвальной стороны, 7 – длина буккальной петли третьего сегмента – расстояние между наиболее буккальной точкой третьего сегмента и наиболее глубоко расположенной точкой задней буккальной петли, 8 – расстояние между наиболее буккальными точками второго и третьего сегментов, 9 – расстояние между наиболее назальной точкой переднего сегмента и самой выпуклой частью с лингвальной стороны.
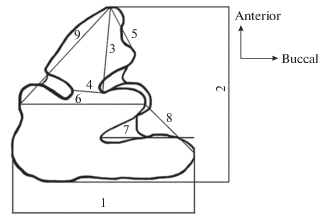
Все промеры и рисунки жевательной поверхности зубов производились только у взрослых особей с помощью бинокулярного микроскопа Zeiss Stemi SV 6 с окулярным измерения представлены в миллиметрах. Для составления морфологической классификационной таблицы проводилось сканирование рисунков, полученных на микроскопе, с последующей обработкой в программе Photoshop CS6.
Морфологическое исследование
В связи с отсутствием работ по выделению фенотипов жевательной поверхности зубов у пищуховых, поиск наиболее важных варьирующих признаков для данного исследования составлялся по аналогии с тем, как это осуществлялось в других работах по млекопитающим (Круковер, 1989; Поздняков, 2005; 2011; Гимранов, 2015; Гимранов, Косинцев, 2017). Ниже приводится подробное описание различных состояний используемых в работе признаков жевательной поверхности третьего нижнего предкоренного зуба северной пищухи.
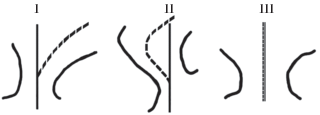
Форма перешейка, соединяющего антероконид и постероконид.Изменение этого признака зависит от расположения по отношению друг к другу пара- и протофлексида; длины и формы этих флексидов и вершины антероконида. В данной работе используется три положения перешейка: I – изогнутый, II – S-образный, III – прямой.
Для изогнутого перешейка характерно низкое положение протофлексида к парафлексиду, за счет чего от начала середины перешейка можно провести изогнутую линию до вершины антероконида. При S-образном перешейке в нижнем положении находится парафлексид и, следовательно, линия от начала перешейка до вершины антероконида имеет изогнутую форму (рис. 2 ). При прямом перешейке парафлексид и протофлексид находятся один напротив другого и образуют прямую линию, от начала перешейка до вершины антероконида можно провести прямую линию.
Рис. 2.
Схема положения перешейка относительно вершины антероконида: I – изогнутый перешеек, II – S-образный перешеек, III – прямой перешеек. Пунктирная линия показывает направление перешейка относительно вершины антероконида.
Форма антероконида. Так как в процессе жевания у пищух большая нагрузка приходится именно на антероконид (Гуреев, 1964), то эта структура является достаточно изменчивой. В данной работе выделяются восемь вариантов строения антероконида (рис. 3).

Тип a. Антероконид в виде ромба, без выраженной либо с едва заметной входящей складкой на лабиальной стороне.
Тип a2. Антероконид в виде ромба без выраженных отростков и складок.
Тип b. Антероконид закруглен, с выраженным отростком с лингвальной или лабиальной сторон.
Тип c1. Антероконид в виде овала с коротким отростком на лабиальной стороне.
Тип c2. Антероконид в виде овала без выраженных отростков.
Тип d. Антероконид в виде ромба или трапеции. Отличием от остальных типов является наличие входящей складки с лабиальной стороны, плоской верхней частью складки парафлексида и длинной лингвальной стороной.
Тип e. Антероконид ромбовидной формы, с вытянутой вершиной, имеет входящую складку и небольшой отросток на лабиальной стороне.
Тип f. Антероконид трапециевидной формы, с выраженными отростками с лингвальной и лабиальной сторон. Вершина антероконида плоская либо конической формы. В редких случаях отростки с лингвальной и лабиальной сторон могут быть едва заметны.
Форма постероконида. Наиболее характерными изменяющимися структурами являются форма протоконида, наличие/отсутствие мезофлексида (рис. 4).
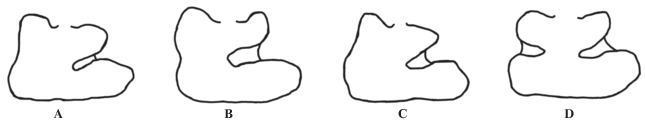
Тип A. Широкий протоконид, с вытянутой в прямом направлении вершиной. Является наиболее часто встречающимся в исследуемой выборке.
Тип B. Широкий протоконид, закруглен, и своей вершиной вытянут вверх.
Тип C. Протоконид очень короткий и имеет заостренную, треугольную форму.
Тип D. Единственный тип с наличием предполагаемого аберрантного признака – наличие мезофлексида. Является наиболее редким по всей исследуемой выборке.
Разложив признаки по состояниям, мы получаем комбинативную решетку, показывающую распределение морфотипов жевательной поверхности р3 по различающим их признакам. Такая решетка позволяет увидеть пределы разнообразия формы жевательной поверхности третьего нижнего премоляра северной пищухи и наглядно показывает, какие ячейки остаются пустыми и какие морфотипы еще можно ожидать.
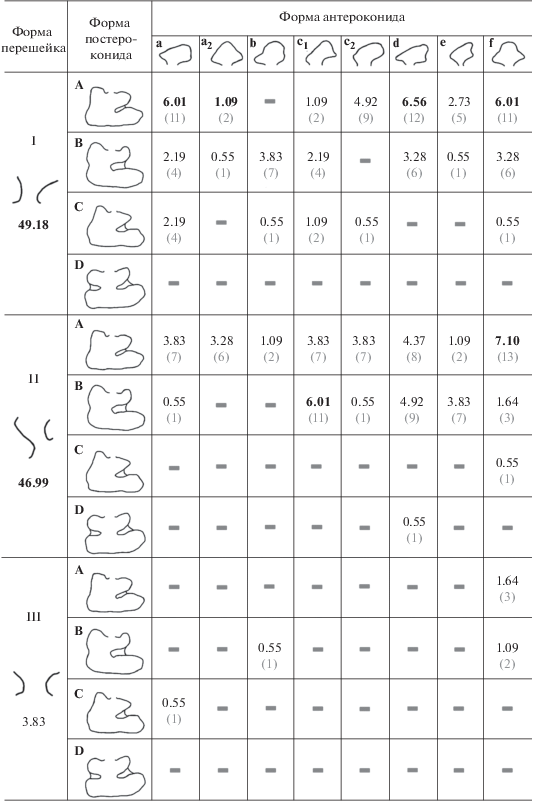
Определяющими критериями для морфологической классификации служат форма перешейка, соединяющего антероконид и постероконид; тип структуры антероконида и постероконида (табл. 4). Классификационная таблица представляет собой прямоугольную сетку, из 8 вертикальных рядов, равным числу типов антероконида, и 12 горизонтальных строк. Для большей компактности таблицы распределение признака по форме перешейка решено было расположить в левой части таблицы. За счет этого ряды типов постероконида разделены на три группы, пронумерованные римскими цифрами. В каждую вертикальную и горизонтальную ячейки помещены графические отображения формы перешейка; типов антероконида и постероконида, с присвоением им буквенного обозначения.
В полученные от пересечения рядов и строк ячейки помещены графические схемы рисунка жевательной поверхности зубов. Каждой модели соответствует конкретный рисунок жевательной поверхности зубов.
При нахождении любого нового состояния признака у современных или ископаемых зубов классификационная таблица может изменяться и дополняться. Также возможно, что некоторые модели жевательной поверхности третьего нижнего премоляра не существуют в принципе, и ячейки останутся пустыми.
Любой морфотип можно записать в виде следующего выражения:
где X – цифровое обозначение формы перешейка, y – буквенное обозначение формы антероконида, Z – буквенное обозначение формы постероконида.РЕЗУЛЬТАТЫ
Статистический анализ
При проведении последовательного дискриминантного анализа исследуемые выборки классифицировались между собой в несколько этапов. После первого этапа выборки с Чукотки, Камчатки и Полярного Урала оказались статистически отличны от остальных групп, так как их значения уровня значимости p-level < 0.01 (табл. 1). Эти выборки были объединены в общую группу.
Таблица 1.
Квадраты расстояний Махаланобиса
| Группы | Группы | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Чукотка | Камчатка | Полярный Урал | Амурская область | Тува | Якутия | Забайкалье | Хабаровский край | Приморский край | Сахалин | |
| Чукотка | 1.42 (0.00) |
5.75 (0.02) |
12.88 (0.00) |
5.27 (0.00) |
7.94 (0.00) |
6.05 (0.00) |
7.78 (0.00) |
10.74 0.00 |
11.37 (0.00) |
|
| Камчатка | 1.42 (0.00) |
5.97 (0.02) |
8.87 (0.00) |
3.22 (0.00) |
4.85 (0.00) |
3.16 (0.00) |
4.28 (0.00) |
7.54 (0.00) |
6.60 (0.00) |
|
| Полярный Урал | 5.75 (0.02) |
5.97 (0.02) |
22.75 (0.00) |
10.21 (0.00) |
14.64 (0.00) |
12.26 (0.00) |
11.63 (0.00) |
19.12 (0.00) |
16.77 (0.00) |
|
| Амурская область | 12.88 (0.00) | 8.87 (0.00) |
22.75 (0.00) |
4.67 (0.00) |
2.10 (0.27) |
2.85 (0.05) |
9.55 (0.00) |
2.38 (0.61) |
8.96 (0.00) |
|
| Тува | 5.27 (0.00) |
3.22 (0.00) |
10.21 (0.00) |
4.67 (0.00) |
1.08 (0.56) |
2.33 (0.02) |
7.74 (0.00) |
7.03 (0.00) |
8.27 (0.00) |
|
| Якутия | 7.94 (0.00) |
4.85 (0.00) |
14.64 (0.00) |
2.10 (0.27) |
1.08 (0.56) |
1.26 (0.30) |
6.09 (0.00) |
3.73 (0.16) |
6.44 (0.00) |
|
| Забайкалье | 6.05 (0.00) |
3.16 (0.00) |
12.26 (0.00) |
2.85 (0.05) |
2.33 (0.02) |
1.26 (0.30) |
3.94 (0.00) |
2.36 (0.43) |
3.44 (0.09) |
|
| Хабаровск | 7.78 (0.00) |
4.28 (0.00) |
11.63 (0.00) |
9.55 (0.00) |
7.74 (0.00) |
6.09 (0.00) |
3.94 (0.00) |
4.72 (0.09) |
3.72 (0.14) |
|
| Приморский край | 10.74 (0.00) |
7.54 (0.00) |
19.12 (0.00) |
2.38 (0.61) |
7.03 (0.00) |
3.73 (0.16) |
2.36 (0.43) |
4.72 (0.09) |
6.86 (0.04) |
|
| Сахалин | 11.37 (0.00) |
6.60 (0.00) |
16.77 (0.00) |
8.96 (0.00) |
8.27 (0.00) |
6.44 (0.00) |
3.44 (0.09) |
3.72 (0.14) |
6.86 (0.04) |
|
Следующий повторный этап дискриминантного анализа включал в себя оставшиеся не классифицированные группы. Конечным результатом таких последовательных этапов стало объединение всех исследуемых выборок в три большие группы: северную, южную и восточную (рис. 5). Северная группа состоит из образцов зубов пищух Камчатки (28), Чукотки (68) и Полярного Урала (4). Южную группу составляют образцы зубов пищух южной части Якутии (15), Забайкалья (22), Амурской обл. (9) и Тувы (15). К восточной группе были отнесены образцы зубов пищух из Хабаровского края (11), Приморского края (5) и Сахалина (5).
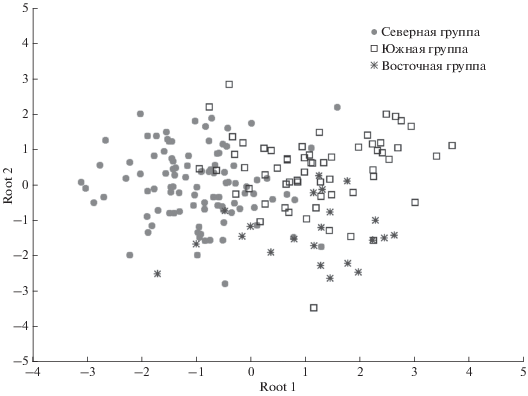
Образованные группы почти полностью повторяют распределение северной пищухи по акустическим расам (Лисовский, 2005). Отличием является то, что образцы из Амурской обл. находятся в южной группе, а в разделении по акустическим расам эта область относится к восточной группе.
Исследованные зубы северной группы мельче подобных из южной и восточной групп (табл. 2). Размеры зубов из восточной и южной групп схожи между собой. При этом расстояние Махаланобиса между южной и восточной группами значительно меньше. Таким образом образцы из этих групп наиболее близки друг к другу по размерным характеристикам нижнего предкоренного зуба, относительно северной группы (табл. 3).
Таблица 2.
Размеры зубов p3 (мм) северной пищухи Ochotona hyperborea
| Признак | Северная группа | Южная группа | Восточная группа | |||
|---|---|---|---|---|---|---|
| Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | |
| 1 | 1.16 ± 0.09 | 0.94–1.44 | 1.31 ± 0.12 | 1.00–1.52 | 1.33 ± 0.10 | 1.00–1.52 |
| 2 | 1.05 ± 0.09 | 0.84–1.30 | 1.23 ± 0.10 | 1.00–1.36 | 1.24 ± 0.11 | 1.00–1.36 |
| 3 | 0.44 ± 0.07 | 0.12–0.60 | 0.54 ± 0.07 | 0.32–0.62 | 0.53 ± 0.08 | 0.32–0.62 |
| 4 | 0.14 ± 0.03 | 0.06–0.20 | 0.13 ± 0.03 | 0.10–0.24 | 0.16 ± 0.04 | 0.10–0.24 |
| 5 | 0.28 ± 0.06 | 0.16–0.46 | 0.40 ± 0.08 | 0.22–0.56 | 0.40 ± 0.09 | 0.22–0.56 |
| 6 | 0.77 ± 0.09 | 0.56–0.96 | 0.92 ± 0.09 | 0.70–1.08 | 0.91 ± 0.10 | 0.70–1.08 |
| 7 | 0.55 ± 0.07 | 0.42–0.74 | 0.65 ± 0.09 | 0.54–0.74 | 0.66 ± 0.06 | 0.54–0.74 |
| 8 | 0.47 ± 0.06 | 0.32–0.78 | 0.54 ± 0.06 | 0.40–0.60 | 0.52 ± 0.06 | 0.40–0.60 |
| 9 | 0.74 ± 0.06 | 0.58–0.98 | 0.85 ± 0.08 | 0.60–0.90 | 0.80 ± 0.08 | 0.60–0.90 |

Частота встречаемости морфотипов в классификационной таблице
После выделения наиболее значимых морфологических признаков была построена классификационная морфологическая таблица жевательной поверхности третьих нижних премоляров северной пищухи (табл. 4). Всего было обнаружено 39 комбинаций по трем признакам.
Поскольку выявленные морфотипы объединяют в себя различное число экземпляров, была рассчитана частота встречаемости, отображаемая в табл. 5. Большее число моделей относится к первой группе с изогнутым перешейком (49.18%). В этой группе встречаются комбинации a-A, d-A и f-A, с частотами встречаемости выше 6%. В этой группе также встречается и наибольшее количество морфотипов с С-формой постероконида. Во второй группе преобладают комбинации морфотипов c1-B и f-A. Наименьшее число морфотипов находится в третьей группе. В этой группе нет доминирующих морфотипов.
Встречаемость различных форм перешейка
Географическая изменчивость структуры жевательной поверхности третьего нижнего премоляра северной пищухи может быть охарактеризована по частоте встречаемости морфотипов в исследуемой выборке (табл. 6).
Таблица 6.
Число и частота встречаемости форм перешейка, соединяющего антероконид и постероконид
| Группа | Положение перешейка | Nобщ | |||
|---|---|---|---|---|---|
| Изогнутый | S-образный | Прямой | |||
| Северная | N | 56 | 39 | 5 | 100 |
| % | 56.00 | 39.00 | 5.00 | ||
| Южная | N | 25 | 36 | 0 | 61 |
| % | 40.98 | 59.02 | 0 | ||
| Восточная | N | 9 | 11 | 2 | 22 |
| % | 40.91 | 50.00 | 9.09 | ||
Преобладание изогнутого перешейка характерно только для северной группы и составляет 56%. У южной и восточной групп преобладает S-образный перешеек, 59.02 и 50% соответственно.
Встречаемость различных форм антероконида
Полный набор форм антероконида встречается только в северной и южной группах (табл. 7). Преобладающими формами для северной группы являются формы a (25%) и c1 (23%). У южной группы форма d встречается в 31% случаях, а форма f почти в 33% случаев. Формы a и c имеют низкие значения частоты встречаемости. У пищух из восточной группы отсутствуют формы a, a2, b, но при этом преобладающими являются такие же формы, как и в южной группе d (32%) и f (40%).
Таблица 7.
Число (N) и частота встречаемости (%) различных форм антероконида
| Группа | a | a2 | b | c1 | c2 | d | e | f | Nобщ | |
|---|---|---|---|---|---|---|---|---|---|---|
| Северная | N | 25 | 2 | 10 | 23 | 15 | 10 | 4 | 11 | 100 |
| % | 25.00 | 2.00 | 10.00 | 23.00 | 15.00 | 10.00 | 4.00 | 11.00 | ||
| Южная | N | 3 | 7 | 1 | 2 | 1 | 19 | 8 | 20 | 61 |
| % | 4.92 | 11.48 | 1.64 | 3.28 | 1.64 | 31.15 | 13.11 | 32.79 | ||
| Восточная | N | 0 | 0 | 0 | 1 | 2 | 7 | 3 | 9 | 22 |
| % | 0 | 0 | 0 | 4.55 | 9.09 | 31.82 | 13.64 | 40.91 | ||
Встречаемость различных форм постероконида
У всех трех групп форма постероконида A является преобладающей и включает в себя более половины экземпляров в каждой группе (табл. 8). Морфотип C в основном характерен для северной группы, а единственный экземпляр с наличием мезофлексида был встречен в восточной группе (4.55%).
Таблица 8.
Число (N) и частота встречаемости (%) морфотипов постероконида
| Группа | A | B | C | D | Nобщ | |
|---|---|---|---|---|---|---|
| Северная | N | 56 | 34 | 10 | 0 | 100 |
| % | 56.00 | 34.00 | 10.00 | 0.00 | ||
| Южная | N | 38 | 22 | 1 | 0 | 61 |
| % | 62.30 | 36.07 | 1.64 | 0 | ||
| Восточная | N | 12 | 9 | 0 | 1 | 22 |
| % | 54.55 | 40.91 | 0 | 4.55 | ||
Исходя из вышесказанного, распределение преобладающих морфотипов для выделенных групп можно записать в следующем виде: для северной группы – [I-a-A] и [I-c1-A]; для южной и восточной групп – [II-d-A] и [II-f-A].
ЗАКЛЮЧЕНИЕ
Для северной части ареала северной пищухи характерны особи, имеющие меньший размер третьего нижнего предкоренного зуба, что отмечалось и другими авторами (Ербаева, 1988; Lissovsky, 2003). Особи, обитающие на юге и востоке ареала, близки между собой по размерам и строению жевательной поверхности третьего нижнего предкоренного зуба. Как было показано ранее, имеет смысл рассматривать вопросы доминирования только отдельных признаков, а не комбинации из состояний признаков (Коваленко, 2003).
Таким образом, на севере ареала доминируют особи северной пищухи с изогнутым перешейком, f-формой антероконида и А-формой постероконида третьего нижнего предкоренного зуба. На юге и востоке преобладают особи с S-образным перешейком, f-формой антероконида и А-формой постероконида.
Список литературы
Воронцов Н.Н., Иваницкая Е.Ю., 1973. Сравнительная кариология пищух (Lagomorpha, Ochotonidae) Северной Палеарктики // Зоологический журнал. Т. 52. Вып. 4. С. 584–588.
Гимранов Д.О., 2015. Морфотипическая изменчивость зубов Mates martes и Martes zibellina // Зоологический журнал. Т. 94. Вып. 5. С. 579–587.
Гимранов Д.О., Косинцев П.А., 2017. Морфотипическая изменчивость резцов бурого (Ursus arctros) и белого (Ursus maritimus) медведей (Carnivora, Ursidae) // Зоологический журнал. Т. 96. Вып. 5. С. 547–562.
Гуреев А.А., 1964. Фауна СССР (Зайцеобразные). Москва, Ленинград: Наука. 276 с.
Ербаева М.А., 1988. Пищухи Кайнозоя. Москва: Наука. 222 с.
Калмыков Н.П., 2015. Млекопитающие обрамления озера Байкал в палеонтологической летописи. Зайцеобразные и грызуны (Lagomorpha and Rodentia, Mammalia) // Байкальский Зоологический журнал. Иркутск. Т. 2. Вып. 17. С. 7–16.
Картавцева И.В., Шереметьева И.Н., Гуськов В.Ю., Вакурин А.А., Кумакшева Е.В., Фрисман Л.В., 2014. К уточнению таксономического положения северной пищухи Ochotona hyperborea Сихоте-Алиня // Вестник ДВО РАН. Вып. 2. С. 79–85.
Коваленко Е.Е., 2003. Эффект нормы признака и его теоретическое значение // Эволюционная биология: история и теория. СПб. Вып. 2. С. 66–87.
Круковер А.А., 1989. Строение и морфотипическая изменчивость жевательной поверхности зубов некорнезубых полевок // Институт геологии и геофизики СО АН СССР. Т. 12. С. 38.
Лисовский А.А., 2005. Сравнительный анализ Акустического репертуара и изменчивости звуковых сигналов пищух (Ochotona, Mammalia) группы Alpina-Hyperborea // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 110. Вып. 6. С. 12–26.
Лисовский А.А., Сердюк Н.В., 2004. Определение позднеплейстоценовых пищух (Ochotona, Lagomorpha, Mammalia) группы alpina-hyperborea из Денисовской пещеры (Алтай) по переднему нижнему предкоренному зубу (P3) // Палеонтологический журнал. Вып. 38. № 6. С. 89–95.
Наследов А.Д., 2004. Математические методы психологического исследования. Санкт-Петербург: Речь. 392 с.
Панасенко В.Е., Тиунов М.П., 2010. Население мелких млекопитающих (Mammalia: Eulipotyphla, Rodentia, Lagomorpha) на южном Сихотэ-Алине в позднем плейстоцене и голоцене // Вестник ДВО РАН. Вып. 6. С. 60–67.
Поздняков А.А., 2005. Структура морфотипической изменчивости M3 серых полевок (Microtus Schrank, 1798) // Систематика, палеонтология и филогения грызунов. СПб. Т. 306. С. 102–114.
Поздняков А.А., 2011. Структура морфологической изменчивости (на примере морфотипов жевательной поверхности первого нижнего коренного зуба серых полевок) // Журнал общей биологии. Т. 72. Вып. 2. С. 127–139.
Поздняков А.А., Литвинов Ю.Н., Лопатина Н.В., 2004. Эколого-морфологическая изменчивость алтайских популяций плоскочерепной полевки Articola strelzowi Kastschenko (Rodentia, Cricetidae, Arvicolinae) // Сибирский экологический журнал. Вып. 4. С. 579–587.
Čermák S., Obuch J., Benda P., 2006. Notes on the genus Ochotona in the Middle East (Lagomorpha: Ochotonidae) // Lynx (Praha). V. 37. P. 51–66.
Kawamichi T. 1981. Vocalisation of Ochotona as a taxonomic character // Proseedings of the World Lagomorph Conference. Ontario. P. 324–339.
Lissovsky A.A., Lissovskaya E.V., 2002. Diagnostic of picas (Lagomorpha, Ochotonidae, Ochotona) from the Putorana Plateau, Eastern Siberia // Russian Journal of Theriology. V. 1. № 1. P. 37–42.
Lissovsky A.A., 2003. Geographical variation of skull characters in pikas (Ochotona, Lagomorpha) of the alpina-hyperborea group // Acta Theriologica. V. 48. № 1. P. 11–24.
Lissovsky A.A., 2014. Taxonomic revision of pikas Ochotona (Lagomorpha, Mammalia) at the species level // Mammalia. V. 78. № 2. P. 199–216.
Volkova V.R., Lissovsky A.A., 2018. The structure of morphological variation in the shape of occlusal surface of permanent teeth in pikas Ochotona (Lagomorpha: Ochotonidae) // Russian Journal of Theriology. V. 17. № 2. P. 91–99.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал