

Зоологический журнал, 2021, T. 100, № 10, стр. 1159-1164
Разнообразие пневматизации ушной капсулы у большой песчанки (Rhombomys opimus, Rodentia, Gerbillinae)
Е. Г. Потапова *
Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: lena-potapova@yandex.ru
Поступила в редакцию 13.09.2020
После доработки 19.10.2020
Принята к публикации 29.10.2020
Аннотация
Описаны специфика и разнообразие пневматизации ушной капсулы у Rhombomys. Специфика проявляется в а) слабом вздутии трубки наружного слухового прохода, б) отсутствии предмеатальной камеры, в) значительном развитии нижней мастоидной камеры и г) образовании особого мастоидно-тимпанального кармана в результате обратного проникновения полости мастоида в барабан. Условиями формирования этого кармана являются значительное смещение назад купола барабанного кольца и наличие широкой тимпано-мастоидной септы. Разнообразие пневматизации ушной капсулы проявляется в разных соотношениях размеров полостей, расположенных в эпитимпанальном, мастоидном и барабанном отделах капсулы. Это разнообразие обусловлено разной последовательностью и разной интенсивностью вздутия этих отделов. Выделены признаки, разные сочетания которых составляют спектр данного разнообразия. Предложен способ их описания. Предварительная оценка изменчивости показала, что песчанки из разных географических точек различаются и по размерным характеристикам ушной капсулы, и по качественным показателям ее пневматизации. Возможно, эти различия отражают экологическую специализацию вида.
Пневматизация ушной капсулы – один из механизмов настройки среднего уха на восприятие определенного акустического спектра (Dallos, 1973; Moore, 1981). Гипертрофированное вздутие капсулы повышает чувствительность уха к низким частотам и считается адаптацией к освоению аридных территорий, где условия звукопередачи наиболее благоприятны для низкочастотного спектра, и где высокая восприимчивость к звукам этого диапазона – один из важнейших факторов выживания (Lay,1972; Павлинов, 1988; Павлинов и др., 1990; Webster, Plassmann, 1992; Momenzadeh et al., 2008; Tabatabaei Yazdi et al., 2014, 2015; и др.). Пневматизация может осуществляться за счет раздувания барабана и (или) формирования дополнительных камер в эпитимпаникуме и мастоиде. Функционально значимо именно изменение объема полости среднего уха, тогда как разнообразие путей пневматизации и характер дифференциации этой полости в значительной степени обусловлены спецификой базовой конструкции уха и историей развития группы. Это дает основание использовать данные по строению ушной капсулы в исследованиях по филогении и систематике грызунов.
Ушная капсула у Gerbillnae изучена достаточно полно и в функциональном, и в филогенетическом аспектах (Lay, 1972; Павлинов, 1980; Павлинов и др., 1990; Pavlinov, 2001, 2008; Потапова, 2016). Выяснены пути и способы ее пневматизации. На уровне рода описана структура разнообразия мастоида и предложены наиболее вероятные сценарии его эволюционных преобразований. Филогенетическая оценка этих материалов учтена при построении системы Gerbillinae (Павлинов и др., 1990; Pavlinov, 2001, 2008).
На основании многочисленных исследований ушной капсулы грызунов (хомяков, песчанок, сонь, белок, роющих, и мн. др.) показано, что филогенетическая составляющая в структуре ее разнообразия, как правило, наиболее четко проявляется на родовом уровне и выше. Однако анализ межвидового и внутривидового разнообразия мастоида у песчанок рода Gerbilliscus, с начальной стадией его пневматизации, показал пригодность таких исследований для оценки межвидовых взаимоотношений и изучения географической изменчивости (Потапова, 2016). Полученные данные оказались важными для понимания морфогенеза слуховой капсулы песчанок и внесли коррективы в существующие представления о семофилезе мастоида в этой группе. Это позволило по-новому оценить некоторые родственные связи внутри Gerbillinae и отчасти смягчить противоречия между морфологическими (Pavlinov, 2001, 2008) и молекулярно-генетическими (Chevret, Dobigny, 2005; Colangelo et al., 2007; Alhajeri et al., 2015) трактовками филогении группы.
У песчанок с увеличенной ушной капсулой внутривидовая изменчивость качественных характеристик ее пневматизации исследована слабо. Косвенно о ней можно судить по изменчивости формы капсулы (Tabatabaei Yazdi et al., 2014). Но, как правило, анализ географической изменчивости черепа у этих песчанок учитывает, в основном, размерные показатели пневматизации (Momenzadeh et al., 2008; Nanova, 2014; и др.).
Цель данной публикации – на качественном уровне описать разнообразие мастоидной пневматизации у Rhombomys opimus (Lichtenstein 1823) – одного из самых высокоспециализированных видов песчанок с сильно вздутой ушной капсулой.
МАТЕРИАЛ И МЕТОДИКА
Использованы материалы из коллекций Зоологического музея МГУ (ЗММУ). Отпрепарированы 48 черепов большой песчанки: удалена часть покровной стенки капсулы между септами (рис. 1). Места сбора материала: Гурьевская обл. (3 экз.); северо-западное Приаралье (6 экз.); Туркмения: центральные Каракумы (3), Марыйская обл. (5), Карабиль (9 экз.); Семиречье (7 экз.); Монголия: Южно-Гобийский аймак (12 экз.), Увэр-Хангайский аймак (3). Линии крепления основных септ на покровной стенке капсулы видны при ее внешнем осмотре. Поэтому для рекогносцировочной оценки географической изменчивости дополнительно исследованы не препарированные черепа из выбранных точек. Для сравнения рассмотрены виды песчанок из родов Psammomys и Meriones.
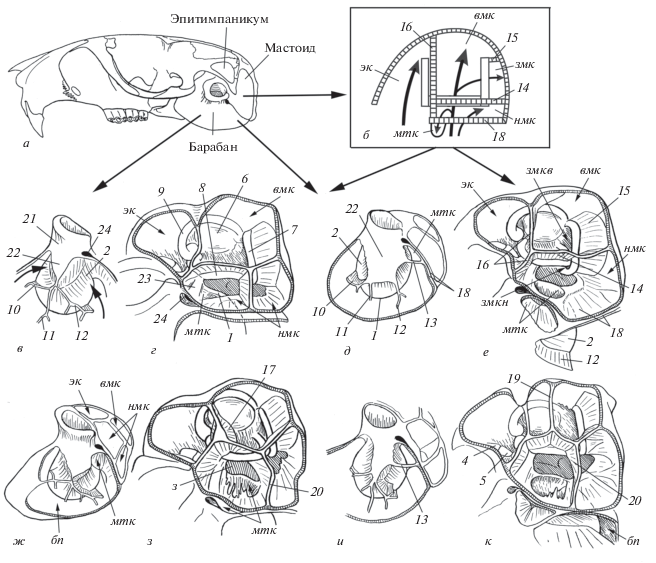
Рис. 1.
Пневматизация ушной капсулы у Rhombomys opimus: а – череп; б – схема пневматизации мастоида; в, д, ж, и – оформление слухового прохода, вид сбоку и немного спереди и снизу; г, е, з, к – мастоид, вид сбоку и немного сверху и сзади; в, г – по экз. S-113151 (ЗММУ) Южная Туркмения, Карабиль; д, е – S-60 770, Гурьевская обл.; ж, з – S-119 273, (ЗММУ) Монголия, Южно-Гобийский аймак; и, к – S-130 285, (ЗММУ) Монголия, Южно-Гобийский аймак. Обозначения: бп – барабанная полость; вмк – верхняя мастоидная камера; змк –задний мастоидный карман и вход в него из верхней (змкв) и нижней (змкн) мастоидных камер; мтк – мастоидо-тимпанальный карман; нмк – нижняя мастоидная камера, эк – эпитимпанальная камера; 1, 2 – барабанное кольцо (1) и его купол (2); 3 – вход в нижнюю мастоидную камеру из барабанной полости; 4 – молоточек; 5 – наковальня; 6 –парафлоккулярная ямка; 7–9 – полукружные каналы: боковой (7), задний (8), передний (9); 10–13 – септы барабанного кольца: передняя (10), средняя (11), задняя (12), стенка мастоидно-тимпанального кармана (13); 14–20 – септы мастоида: септа бокового канала (14), септа заднего канала (15), мастоидная септа (16) и ее конусовидное расщепление (17), тимпано-мастоидная (18), дополнительная септа в верхней (19) и в нижней (20) мастоидной камере; 21, 22 – трубка слухового прохода (21) и однослойная часть его стенки (22); 23 – трубка лицевого канала; 24 – шилососцевидное отверстие, for. stylomastoideum.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Ушная капсула у Rhombomys крупная и конструктивно высоко организована. Черты специализации проявляются и в строении костного чехла внутреннего уха, и в особенностях пневматизации капсулы. По размерам капсулы относительно длины черепа большая песчанка уступает лишь некоторым видам Meriones и Psammomys с гипертрофированно раздутой капсулой. Увеличение размеров капсулы у Rhombomys обеспечено вздутием всех ее отделов (барабана, мастоида, эпитимпаникума – рис. 1а, 1б).
Своеобразие барабанного отдела большой песчанки проявляется в оформлении слухового прохода. По сравнению с другими песчанками у Rhombomys трубка наружного слухового прохода максимально оттянута (рис. 1в, 21) и слабо вздута. Ее пневматизация никогда не достигает той же степени, что у некоторых видов Meriones или Psammomys. Часть нижней стенки трубки всегда остается однослойной (рис. 1в–1д, 22). Вздутие трубки идет несколькими путями: спереди, сзади (стрелки на рис. 1в) и, в меньшей степени, снизу. Оно либо равномерное, либо более интенсивное сзади. Воздушный карман в передней стенке трубки, так называемая предмеатальная камера, не формируется. В результате вздутия барабанное кольцо, annulus tympanicus (рис. 1д, 1), обособляется от покровной стенки и крепится по краю образовавшегося костного купола, cupula annuli tympanicus (рис. 1д, 2), который свешивается внутрь барабанной полости. Купол барабанного кольца связан с наружной стенкой барабана тремя септами: передней, средней и задней (рис. 1в–1д, 10–12). Их размеры, расположение и взаимосвязь варьируют в зависимости от того, какие из названных выше путей пневматизации преобладают.
Барабанное кольцо у Rhombomys сдвинуто в мастоид и соединяется с краем всегда широкой тимпано-мастоидной септы (рис. 1г–1е, 1, 18). Такое смещение создает условия для формирования характерного для этого вида мастоидно-тимпанального кармана (рис. 1б, 1е–1з, мтк). Сходные взаимоотношения этой септы и барабанного кольца, но без образования полноценного кармана, отмечены у Psammomys. Среди Meriones барабанное кольцо и тимпано-мастоидная септа не перекрываются. Эта септа, как правило, неширокая, причем у видов с увеличенными барабанами (M. meridianus, M. libycus и др.) она наиболее узкая.
Пневматизация эпитимпано-мастоидного отдела у Rhombomys осуществляется по тому же типу (рис. 1б), что и у других “высших” песчанок (Meriones, Cheliones, Brachiones, Psammomys и др). Расширение барабанной полости происходит одновременно в двух направлениях: спереди и сзади. Спереди (над слуховым проходом) она заходит в этитимпаникум, где образуется эпитимпанальная камера (рис. 1б, эк). Сзади она двумя потоками (изнутри и снаружи бокового канала) проникает в мастоид, где формируются две основных камеры: верхняя и нижняя мастоидные (рис. 1б, 1г, 1е, вмк, нмк).
Эпитимпанальная камера крупная, значительно раздута и в поперечном, и в продольном направлениях. Мастоидная септа (рис. 1б, 1е, 16), которая полностью разделяет полости эпитимпаникума и мастоида, сдвинута немного назад и проходит по дну парафлоккулярной ямки. Вся ампула и часть трубки переднего полукружного канала лежат в эпитимпанальной полости. Этот канал соединен только с дном парафлоккулярной ямки с помощью септы, вправленной внутрь его арки. Как правило, он не связан ни с внутренней, ни с наружной стенкой камеры. Лишь иногда короткий гребень соединяет его с крышей этой полости.
Обе мастоидные камеры крупные. Они бывают равноценными по размерам или одна может быть больше другой. Каждая из них может быть одинарной или двойной (рис. 1г, 1з, 1к). Дополнительная перегородка в верхней мастоидной камере возникает либо при расщеплении мастоидной септы (рис. 1з, 17), либо изначально формируется как дополнительное ребро жесткости на наружной стенке мастоида (рис. 1к, 19). В обоих случаях она свободно свешивается внутрь камеры и не перегораживает ее полностью. Обе части верхней мастоидной камеры сообщаются между собой и связаны с барабанной полостью через отверстие в арке бокового полукружного канала.
Задний мастоидный карман (рис. 1б, змк) довольно крупный, имеет форму конуса или цилиндра. Его стенка (“септа заднего канала” – рис. 1б, 1е, 15) широкая и независимо от формы кармана крепится к задней стенке капсулы, что типично для всех Rhombomiini и отличает их от Gerbillini. Основная часть заднего кармана у Rhombomys образуется как расширение верхней мастоидной камеры назад через арку заднего полукружного канала (рис. 1е, змкв). Кроме того, может формироваться еще и небольшой дополнительный задний карман (рис. 1е, змкн), который через отверстие в перекрестье бокового и заднего полукружных каналов сообщается не с верхней, а с нижней мастоидной камерой. Оба кармана имеют общую наружную стенку, но изнутри разделены перегородкой.
Нижняя мастоидная камера (рис. 1б, нмк) крупная даже при ее минимальном, по меркам данного вида, развитии. Она бывает меньше верхней мастоидной камеры, но чаще сопоставима с ней по размерам или превосходит ее. При раздувании мастоида часто именно эта камера обеспечивает максимальный прирост объема полости. У песчанок других родов, например Meriones, в большей степени увеличивается верхняя мастоидная камера. Значительное развитие именно нижней мастоидной камеры – характерная особенность Rhombomys. Часто эта камера двойная. Внешний вид, размеры и расположение дополнительной перегородки варьируют. Это может быть небольшой гребень, переходящий с септы бокового канала на покровную стенку, или широкая пластинка, свешивающаяся внутрь камеры (рис. 1з, 1к, 20).
Расширение этой камеры может приводить к обратному проникновению мастоидной полости в тимпанальную позади лицевого канала (рис. 1г, 23). В результате образуется так называемый мастоидно-тимпанальный карман, который сообщается только с нижней мастоидной камерой и не связан напрямую с полостью барабана. Он может выглядеть как глубокая ямка (рис. 1ж, 1з, мтк) или как небольшая камера (рис. 1д, 1е, мтк), дно которой образовано барабанной костью. Наличие или отсутствие данного кармана зависит от интенсивности вздутия барабана и мастоида. Если пневматизация трубки слухового прохода начинается раньше и (или) идет интенсивнее, чем вздутие мастоида, то полное обособление барабанного кольца под шилососцевидным отверстием происходит из барабанной полости (рис. 1в, 2). Это препятствует обратному впячиванию нижней мастоидной камеры в эту область, и мастоидно-тимпанальный карман не образуется (рис. 1г, 1). Если, наоборот, пневматизация мастоида происходит быстрее, чем раздувание барабанного пузыря, обособление барабанного кольца под шилососцевидным отверстием осуществляется из мастоида (рис. 1д, 1е, 1и, 1к, мтк). Образуется тимпано-мастоидный карман, который полностью отделен от полости барабана. Его нижняя стенка соединяет купол барабанного кольца с покровной стенкой барабанного пузыря и играет роль дополнительной септы (рис. 1д, 1и, 13). Размеры этого кармана варьируют.
ЗАКЛЮЧЕНИЕ
Как и следовало ожидать, характер и параметры внутривидовой изменчивости мастоида у большой песчанки иные, нежели у песчанок Gerbilliscus с начальной стадией пневматизации мастоида (Потапова, 2016). Межвидовая и внутривидовая изменчивость у Gerbilliscus проявляется в признаках, которые определяют типы пневматизации мастоида у песчанок в целом. В пределах этого рода намечены все пути трансформации мастоида и представлен практически весь спектр вариантов его дифференциации, характерный для этой группы. Частично этот спектр реализован и на внутривидовом уровне, но внутри вида он менее полный и может различаться у разных видов.
В отличие от Gerbilliscus, у Rhombomys изменчивость мастоида не затрагивает его типовые характеристики. В ее спектре нет вариантов, схожих с другими характерными для Gerbillinae типами его строения. Пневматизация ушной капсулы у Rhombomys соответствует строго одному типу, свойственному и другим “высшим” песчанкам, но в деталях обладает выраженной видовой спецификой. Эта специфика проявляется: а) в слабом вздутии трубки наружного слухового прохода, б) в отсутствии предмеатальной камеры и в) в обратном проникновении мастоидной полости в барабанную, которое приводит к образованию мастоидно-тимпанального кармана. Развитие этого кармана предопределяют три характерные для Rhombomys особенности: г) смещение назад купола барабанного кольца, д) широкая тимпано-мастоидная септа и е) значительное развитие нижней мастоидной камеры.
Разнообразие в строении ушной капсулы у Rhombomys отражает неравномерность ее пневматизации в разных направлениях и проявляется, прежде всего, в разной степени вздутия ее отделов: наружного слухового прохода барабана, эпитимпаникума и обеих частей мастоида. У этого вида наибольшее раздувание происходит в эпитимпаникуме и мастоиде. Пневматизация слухового прохода незначительная, диапазон ее изменчивости довольно узкий. Но в пределах этого диапазона степень пневматизации трубки слухового прохода (“слабая”, “умеренная”, “сильная”) можно оценивать по площади однослойного участка ее стенки (рис. 1в, 22 и 1д, 22) или по размерам свободной части купола барабанного кольца (рис. 1в, 2 и 1д, 2). Этот показатель важен для сравнения последовательности и интенсивности раздувания барабана и мастоида. У Rhombomys увеличение полости среднего уха может начинаться 1) с пневматизации стенки слухового прохода с последующим вздутием мастоида (рис. 1в, 1г), 2) с интенсивного раздувания мастоида с последующей пневматизацией трубки (рис. 1д, 1е, 1и, 1к) и 3) с одновременного раздувания в обоих направлениях (рис. 1ж, 1з).
Об интенсивности вздутия эпитимпанального и мастоидного отделов можно судить по площади наружной стенки соответствующих камер. Однако для описания пневматизации капсулы по качественным показателям важна оценка относительных размеров этих камер (в терминах “меньше”, “равна”, “больше”) при их попарном сравнении друг с другом: эпитимпанальной с верхней мастоидной, эпитимпанальной с нижней мастоидной, верхней мастоидной с нижней мастоидной. Кроме того, необходимо учитывать, представлены или нет дополнительные перегородки в камерах, а также отмечать наличие или отсутствие воздушных карманов: “дополнительного заднего мастоидного” и “мастоидно-тимпанального”. Спектр разнообразия пневматизации ушной капсулы у данного вида определяется количеством вариантов с разным сочетанием всех этих параметров.
Рекогносцировочная оценка географической изменчивости ушной капсулы у Rhombomys показывает различия данной конструкции в разных точках ареала вида, несмотря на высокий уровень индивидуальной изменчивости в каждой точке.
В наибольшей степени различаются песчанки из Семиречья и Монголии. У песчанок из Семиречья капсула наименее увеличена. Пневматизация слухового прохода широко варьирует: представлен весь спектр переходов от едва наметившегося до почти полного обособления от стенки барабанного кольца. Часто это обособление происходит со стороны барабана. Мастоидно-тимпанальный карман, как правило, есть, но может и отсутствовать. Нижняя мастоидная камера меньше верхней мастоидной или они равноценны. Дополнительных перегородок в обеих камерах нет. Септа заднего канала конусовидно расходится. Дополнительный задний мастоидный карман не развит.
У песчанок из Монголии барабаны наиболее вздуты (рис. 1ж–1к). Пневматизация стенок слухового прохода максимальная в рамках видового диапазона. Мастоидно-тимпанальный карман, как правило, крупный, но может и отсутствовать. Нижняя мастоидная камера намного крупнее верхней и обычно разделена на две части. Верхняя мастоидная камера, как правило, тоже двойная. Септа заднего мастоидного кармана прямая. Дополнительный задний карман бывает развит. Песчанки из других точек занимают промежуточное положение между песчанками Семиречья и Монголии. Среди них выделяются песчанки из Карабиля более вздутым барабаном и часто более значительным развитием верхней мастоидной камеры (рис. 1в, 1г). Своеобразие песчанок из других точек оценить трудно из-за высокого уровня индивидуальной изменчивости.
Таким образом, различия в пневматизации ушной капсулы у песчанок из разных географических точек связаны как с изменением объема барабанной полости (т.е. с функционально значимым параметрами), так и с различиями качественных характеристик раздувания. Наибольшее расхождение наблюдается у песчанок из тех географических точек, которые заметно различаются по экологически значимым для вида параметрам среды. Вполне возможно, что описанная выше морфологическая дифференциация ушной капсулы в значительной степени отражают экологическую специализацию вида. Чтобы оценить роль адаптивной и исторической составляющих в проявлении географической изменчивости важно провести статистический анализ с учетом не только размерных, но и качественных характеристик пневматизации.
Список литературы
Павлинов И.Я., 1980. Эволюция и таксономическое значение строения костного среднего уха в подсемействе песчанок Gerbillinae (Rodentia: Cricetidae) // Бюллетень Московского общества испытателей природы, отдел биологический. Т. 85. № 4. С. 20–33.
Павлинов И.Я., 1988. Эволюция мастоидного отдела слухового барабана у пустынных грызунов // Зоологический журнал. Т. 67. № 5. С. 739–750.
Павлинов И.Я., Дубровский Ю.А., Россолимо О.Л., Потапова Е.Г., 1990. Песчанки мировой фауны. Москва: Наука. 368 с.
Потапова Е.Г., 2016. Разнообразие пневматизации мастоида у песчанок родов Taterillus и Gerbilliscus (Rodentia, Gerbillidae): морфогенетический и филогенетический аспекты // Сборник трудов Зоологического музея МГУ. Т. 38. С. 356–379
Alhajeri B.H., Hunt O.J., Steppan S.J., 2015. Molecular systematics of gerbils and deomyines (Rodentia: Gerbillinae, Deomyinae) and a test of desert adaptation in the tympanic bulla // Journal Zoological Systematics and Evolutionary Research. V. 53. № 4. P. 312–330.
Chevret P., Dobigny G., 2005. Systematics and evolution of the subfamily Gerbillinae (Mammalia, Rodentia, Muridae) // Molecular Phylogenetics and Evolution. V. 35. № 3. P. 674–688.
Colangelo P., Granjon L., Taylor P.J., Corti M., 2007. Evolutionary systematics in African gerbilline rodents of the genus Gerbilliscus: Inference from mitochondrial genes // Molecular Phylogenetics and Evolution. V. 42. № 3. P. 797–806.
Dallos P., 1973. The auditory periphery biophysics and physiology. New York–London: Academic Press. 548 p.
Lay D.M., 1972. The anatomy, physiology, functional significance and evolution of specialized hearing organs of gerbilline rodents // Journal of Morphology. V. 38. № 1. P. 41–120.
Momenzadeh M., Darvish J., Toutoniyan F., Sarmad M., Sadeghi Shakib F., 2008. Investigation of climatic effects on the shape and volume of tympanic bulla of Meriones libycus and Meriones persicus (Muridae: Rodentia) from northeastern Iran: an evolutionary approach // Iranian Journal of Animal Biosystematics. V. 4. № 1. P. 63–73.
Moore W.J., 1981. The mammalian skull. London–New York: Cambridgen Univ. Press. 369 p.
Nanova O.G., 2014 Geographical variation in the cranial measurements of the midday jird Meriones meridianus (Rodentia: Muridae) and its taxonomic implications // Journal of Zoological Systematics and Evolutionary Research. V. 52. № 1. P 75–85.
Pavlinov I.Ya., 2001. Current concepts of gerbillid phylogeny and classification // African small mammals. Proc. 8th Internat. Symp. African small mammals. Paris: IRD Ed. P. 141–149.
Pavlinov I.Ya., 2008. A review of phylogeny and classification of Gerbillinae (Mammalia: Rodentia). Moscow: Moscow Univ. Publ. 68 p.
Tabatabaei Yazdi F., Adriaens D., Darvish J., 2014. Cranial phenotypic variation in Meriones crassus and M. libycus (Rodentia, Gerbillinae), and a morphological divergence in M. crassus from the Iranian Plateau and Mesopotamia (Western Zagros Mountains) // European Journal of Taxonomy. V. 88. P. 1–28.
Tabatabaei Yazdi F., Colangelo P., Adriaens D., 2015. Testing a long-standing hypothesis on the relation between the auditory bulla size and environmental conditions: a case study in two jird species (Muridae: Meriones libycus and M. crassus) // Mammalia. V. 79. № 2. P. 185–200.
Webster D.B., Plassmann W., 1992. Parallel evolution of low frequency sensitivity in old world and new world desert rodents // D.B. Webster, A.N. Popper and R.R. Fay, eds. The evolutionary biology of hearing. Berlin–Heidelberg–New York: Springer. P. 625–631.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал