

Зоологический журнал, 2021, T. 100, № 11, стр. 1288-1304
Особенности строения кожи и ее дериватов у некоторых представителей подсемейства Cricetinae (Cricetidae, Rodentia): почему это важно знать
О. Ф. Чернова a, *, В. П. Куприянов a, **, Н. Ю. Феоктистова a, ***, А. В. Суров a, ****
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Ленинский пр., 33, Россия
* E-mail: chernova@sevin.ru
** E-mail: meahn@mail.ru
*** E-mail: Feoktistovanyu@gmail.com
**** E-mail: surov@sevin.ru
Поступила в редакцию 15.03.2021
После доработки 16.06.2021
Принята к публикации 20.06.2021
Аннотация
Подсемейство Cricetinae включает широкий спектр видов, различающихся по размеру, социальной организации, экологии. Морфофункциональные адаптации в значительной степени определяются особенностями кожных покровов. Исследовали кожно-волосяной покров и специфические кожные железы четырех представителей подсемейства, относящихся к двум родам, – Аllocricetulus curtatus, A. еversmanni, Cricetulus griseus и С. sokolovi. Показано, что у всех исследованных видов шерсть довольно однородная, не имеет длинного ворса, слабо разделяется на ярусы, пуховые волосы не обнаружены. Вибриссы имеют специализированную архитектонику. Выявлены три морфотипа среднебрюшной железы: железа-сумка без специализированных волос (у Allocricetulus), железа-сумка со специализированными волосами (у Phodopus) и железа-поле со специализированными волосами (у Cricetulus). Впервые у взрослых особей A. curtatus и A. eversmanni описаны специфические боковые сальные железы, располагающиеся билатерально в виде железистых полей на уровне косто-вертебрального угла. Обсуждается необходимость исследования морфологии специфических желез Cricetinae для адекватной оценки их роли в ольфакторной коммуникации.
Несмотря на многолетние планомерные исследования, накопленный большой фактический материал и разработку целого ряда теоретических обобщений (Wyatt, 2014) химическая коммуникация млекопитающих остается актуальным направлением исследований. Тем не менее многие виды млекопитающих не охвачены такими исследованиями, и зачастую получение этологических данных опережает морфологическое описание органов-источников обонятельных сигналов, что иногда приводит к парадоксальной ситуации – тестируется, описывается и обсуждается поведение реципиентов в ответ на запах секрета отсутствующих у них желез. Вот почему важно, чтобы этологические заключения опирались на морфологические и гистологические исследования источников пахучих сигналов. В этологических экспериментах по ольфакторной коммуникации и комфортному поведению хомячков подсемейства Cricetinae при интерпретации полученных данных мы столкнулись с проблемой слабой изученности их кожно-волосяного покрова и специфических кожных желез (СКЖ) – источников пахучих сигналов. Эта проблема и определила необходимость проведения данного исследования.
Рецентные хомяки подсемейства Cricetinae представляют собой отдельную филетическую линию. Это в основном обитатели открытых пространств, от уровня моря до высокогорий (4500 м) палеарктической части Евразии. Наибольшая длина тела (240–320 мм) характерна для обыкновенного хомяка (Cricetus сricetus), наименьшая (75–92 мм) – для хомячка Роборовского (Phodopus roborovskii) (Павлинов, 2006).
По последним молекулярно-генетическим и морфологическим данным подсемейство Cricetinae включает 18 видов, относящихся к девяти родам: Cricetus, Cricetulus, Allocricetulus, Mesocricetus, Tscherskia, Cansumys, Phodopus, Urocricetus и Nothocricetulus (Lebedev et al., 2018). Род хомяки обыкновенные (Cricetus Leske 1779) монотипичный, состоит из одного вида – обыкновенного хомяка (C. сricetus Linnaeus 1758). Род хомячки серые (Cricetulus Milne-Edwards 1867) включает, по современным понятиям, видовой комплекс C. barabensis sensu lato (= C. barabensis Pallas 1973) c тремя хромосомными формами – “barabensis” (2n = 20), “griseus” (2n = 22) и “pseudogriseus” (2n = 24), различающимися одной или двумя “робертсоновскими перестройками” (Kral et al., 1984; Romanenko et al., 2007; Малыгин и др., 1992), и два хорошо обособленных вида – хомячка Соколова (C. sokolovi Orlov et Malygin 1988) и длиннохвостого хомячка (C. longicaudatus Milne-Edwards 1867) (Lebedev et al., 2018). Род эверсманновы хомячки (Allocricetulus Argyropulo 1932) объединяет два вида – хомячка Эверсманна (A. eversmanni Brandt 1859) и монгольского хомячка (A. curtatus Allen 1925). Род средние хомяки (Mesocricetus Nehring1898) в классических системах включает четыре вида: золотистого (или сирийского, или переднеазиатского) хомяка (M. auratus Waterhouse 1839), хомяка Брандта (M. brandti Nehring 1898), хомяка Радде (M. raddei Nehring 1894) и хомяка Ньютона (M. newtoni Nehring 1898). Род хомячки крысовидные (Tscherskia Ognev 1914) монотипичный, представлен крысовидным хомячком (T. triton Winton 1899). Род хомяки канские (Cansumys Allen 1928) включает один вид – хомяка канского (C. canus Allen 1928) (Павлинов, 2006). Род Nothocricetulus (не имеет русского названия) – монотипичный, ранее был представлен одним видом – Cricetulus migratorius Pallas 1773 (в настоящее время Nothocricetulus migratorius Pallas 1773), возможно с несколькими полувидами или подвидами (Lebedev et al., 2018). Род Urocricetus Satunin 1902 (также не имеет русского названия) объединяет два вида: короткохвостого хомячка (U. аlticola Thomas 1917) и тибетского хомячка (U. kamensis Satunin 1903). Систематика Urocricetus пока остается дискуссионной (Lebedev et al., 2018). Относительно близки к этому роду мохноногие хомячки (Phodopus Miller 1910), с тремя видами: хомячком Роборовского (P. roborovskii Satunin 1903), хомячком Кэмпбелла (P. campbelli Thomas 1905) и джунгарским (P. sungorus Pallas 1771).
По цвету шерсти палеарктические хомяки разделяются на две группы – с “предупреждающим” и криптическим окрасом (Воронцов, 1982). К первой относят представителей Cricetus и Mesocriсetus – с ярко окрашенной шерстью спины и черным брюхом (либо черным горловым пятном), которое они демонстрируют при оборонительном поведении. Ко второй – более мелкие виды других родов. Особенно выражен криптический окрас у обитателя пустынь – P. roborovskii. Для всех представителей Cricetinae характерен мягкий, пушистый волосяной покров.
Достаточно детально кожно-волосяной покров исследован лишь у C. (N.) migratorius (Соколов, 1973) и Phodopus (Феоктистова, 2008; Феоктистова и др., 2012). Что касается других представителей подсемейства, то сведения о строении их кожно-волосяного покрова весьма ограничены.
Известно, что у многих видов млекопитающих хорошо развиты СКЖ, секрет которых зверьки оставляют на субстрате в качестве пахучих меток, играющих важную роль в ориентации как самих хозяев, так и при передаче информации другим особям, например, о занятости территории и нор (Рожнов, 2011). Очевидно, что маркирование особенно важно для территориальных и агрессивных видов. Для СКЖ характерен половой диморфизм, сезонность функционирования и возрастная изменчивость (см. Соколов, Чернова, 2001). У представителей Cricetinae имеются СКЖ разных типов, но описано лишь небольшое их число.
О функционировании СКЖ хомяков мы судим, как правило, только по косвенным данным (маркировочному поведению), так как специальных исследований, включающих, например, гландэктомию, гормонотерапию и т.д., проводилось немного. Лучше других исследованы функции СКЖ у Phodopus (Феоктистова, 2008). Показано, например, что запах секрета среднебрюшной железы (СБЖ) участвует в распознавание пола как у самцов (Васильева, 1990; Lai et al., 1996), так и у самок (Кропоткина и др., 2012), причем роль запаха этого секрета меняется в зависимости от сезона года. По нему самцы могут распознавать физиологическое состояние самок и индивидуальные особенности особей. У P. campbelli и P. sungorus в распознавании пола участвует также запах секрета желез, расположенных в углах рта. Известна уникальная роль этого секрета в регуляции пищеварения, поддержании иммунитета детенышей и скорости их развития (Феоктистова и др., 2004; Хрущова, 2007). Известно, что секрет СБЖ самок C. griseus влияет на изменение уровня тестостерона у самцов, повышая его в зимне-весенний период (Поташникова, Феоктистова, 2014).
Общее для 12 видов Cricetinae описание топографии, конфигурации (на примере P. sungorus) и микроструктуры СБЖ (на примере A. eversmanni) выполнено Воронцовым и Гуртовым (1959). Авторы указывают, что СБЖ присутствует у всех палеарктических видов хомяков, объединяемых в подтрибу Cricetini11. Эта железа расположена на осевой линии тела впереди гениталий и имеет вид небольшого щелевидного углубления, окруженного подковообразным валиком. Густой жироподобный секрет может почти полностью заполнять щель. Латеральные стороны валика покрыты шерстью, неотличимой от волосяного покрова тела. Медиальные стороны и дно щели покрывают особые короткие волоски. СБЖ состоит из большого числа полиальвеолярных голокриновых желез. Секрет выводится через протоки, открывающиеся в волосяные луковицы (на дне мешка) или непосредственно в волосяную сумку). Вся железа окружена крупным слоем соединительной ткани с мускульными волокнами. Авторы предполагают, что СБЖ настоящих хомяков произошла путем развития сальных желез на данном участке тела. Активизация функционирования СБЖ происходит в период размножения. С окончанием этого периода она уменьшается в размерах в 2–4 раза и становится практически незаметной при наружном осмотре. Из чего делается вывод о том, что СБЖ каким-то образом связана с размножением, а ее секрет может быть важен для маркирования границ индивидуального участка. Развитие СБЖ идет быстрее у детенышей из весенних выводков, приступающих к размножению в том же году, нежели у перезимовавших зверьков, у которых развитие СБЖ задерживается до периода весеннего размножения. Несмотря на довольно детальное описание “типичной” микроструктуры СБЖ, проведенное на отдельных примерах, авторы не обсуждают возможную видо- или родоспецифичность этого органа, проявление полового диморфизма. Они указывают лишь на тот факт, что наличие СБЖ, наряду с рядом других морфологических особенностей, позволяет четко очертить границы палеарктической трибы Cricetini (Воронцов, Гуртовой, 1959).
Описаны строение Гардеровых желез млекопитающих (Бодяк, 1994) и их роль в стимулировании полового поведения самцов и процептивного поведения самок Mesocricetus auratus (Соколов и др., 1994).
На наличие боковых желез (БЖ) у C. cricetus обратил внимание еще Паллас (Pallas, 1778), а Вртиш описал их гистологию (Vrtiš, 1930). БЖ формируют крупные железистые плакоды, расположенные по бокам тела. У M. auratus они представляют собой отдельные ароматические железки, происходящие от сальных желез волосяных луковиц с голокриновым типом секреции. Для этих желез характерен половой диморфизм, и они участвуют в маркировке территории (Lipkow, 1954). У самок T. triton экспериментально доказано ингибирующее влияние эстрадиола на размеры, секреторную активность и химический состав секрета БЖ (Zhang et al., 1999). Скирроу и Ризан (Skirrow, Rysan, 1976), наблюдая за поведением C. griseus, отметили, что при социальных контактах хомячки расчесывают бока задними лапами, а затем проволакивают ано-генитальную область по субстрату. Они предположили, что таким образом секрет БЖ наносится на поверхность лап, а затем распространяется при движении. Однако никаких сведений даже о наличии этого типа желез у китайского хомячка, а также о возможной роли СБЖ, секрет которой также мог переноситься на субстрат при проволакивании брюшной поверхности тела, они не приводят. По нашим собственным наблюдениям, A. curtatus и A. eversmanni демонстрируют сходные паттерны маркировочного поведения, что предполагает наличие и у них не только СБЖ, но и БЖ. Однако эти предположения основаны лишь на поведенческих наблюдениях и не могут служить доказательством участия тех или иных желез в маркировочном поведении. Необходимы комплексные исследования, включающие как макро-морфологическое, так и гистологическое описание кожно-волосяного покрова, топографии и строения СКЖ у различных видов подсемейства Cricetinae.
Цель настоящего исследования – восполнить пробел в знаниях о коже и ее производных, провести сравнительно-морфологический анализ, детально изучив кожно-волосяной покров и СКЖ четырех видов, принадлежащих к двум родам подсемейства – Cricetulus (С. griseus, C. sokolovi) и Allocricetulus (A. curtatus и A. eversmanni).
МАТЕРИАЛ И МЕТОДЫ
Все исследованные виды включены в Центр коллективного пользования “Живая коллекция диких видов млекопитающих” Института проблем экологии и эволюции им А.Н. Северцова Российской академии наук (ИПЭЭ РАН). Кроме китайского хомячка они принадлежат ко второму–четвертому поколениям лабораторного разведения от пар, отловленных в природе (Саратовская обл., Тыва и Монголия). Китайский хомячок относится к лабораторной линии и получен из зоопитомника г. Халле (Германия).
Морфологические исследования проводили в Центре коллективного пользования “Инструментальные методы в экологии” ИПЭЭ РАН с применением морфометрии, световой и сканирующей электронной микроскопии (СЭМ) и гистологии. Измерения массы тела и СКЖ проводили на отдельных выборках живых особей (табл. 1). Часть животных была подвергнута эвтаназии на аппарате Плинаркон-5 (ООО “ПрофМТ”, Санкт-Петербург), у них взяты пробы кожного покрова с холки, крестца, бока, брюха, углов рта и подошв задних лап для исследования микроструктуры кожи, волос, вибрисс и СКЖ. Материал получен от половозрелых животных: хомячка Эверсманна (3 самца и 2 самки), монгольского хомячка (2 самца и 2 самки), хомячка Соколова (2 самца) и китайского хомячка (3 самца и 6 самок). Образцы фиксировали в 10% нейтральном формалине. Подготовку гистологических препаратов осуществляли по стандартным методикам (Соколов и др., 1988) с использованием полуавтоматического специализированного гистологического оборудования фирмы “Medite” (Германия) – гистопроцессора TPC-15, заливочной станции TES-99, микротома Meditome M530. Срезы толщиной 6–8 мкм последовательно окрашивали гематоксилином Эрлиха и эозином, заливали жидким стеклом LEICA CV MOUNT. Микрофотографии препаратов изготовляли с помощью моторизованного микроскопа Keyence Biorevo BZ-9000 (“Keyence”, Япония). Образцы волос разбирали на категории и порядки под бинокулярной лупой. Остевые волосы первого порядка (ость I), вибриссы и волосы из области СБЖ исследовали с использованием СЭМ JSM 840А (“Jeol”, Япония) и TESCAN (Чехия). Для этого волосы очищали в шампуне, промывали в воде, проводили через спирты возрастающей концентрации и ацетон, высушивали в критической точке на установке Hitachi Critical Point Dryer HCP-1 (“Hitachi”, Япония), наклеивали на предметные столики. Для обеспечения электропроводимости исследуемых образцов их напыляли золотом на оборудовании S150A Sputter Coater (“Edwards”, Великобритания). Электронограммы получали с поверхности волос и с трансверсальных и сагиттальных срезов стержня. Все рисунки были масштабированы, скомпонованы и отредактированы в программе Adobe Photoshop 11 (“Adobe”, США), но изменения касались лишь масштаба, контрастности и резкости изображений.
Таблица 1.
Средние значения массы тела и размеров специфических кожных желез (по внешним измерениям) исследованных хомячков (по 5 особей каждого вида и пола)
| Вид, пол | Масса тела (X ± Sx) | Длина среднебрюшной железы (X ± Sx) | Длина боковой железы (X ± Sx) | |
|---|---|---|---|---|
| A. curtatus | Самка | 67.6 ± 2.14 | 12.2 ± 0.76 | 3.8 ± 0.59 |
| Самка | 53.0 ± 4.59 | 6.7 ± 0.17 | 2.25 ± 0.18 | |
| A. eversmanni | Самец | 74.7 ± 1.11 | 10.7 ± 0.92 | * |
| Самка | 56.0 ± 3.39 | 6.5 ± 1.02 | * | |
| C. sokolovi | Самец | 39.4 ± 2.01 | 7.8 ± 1.07 | 3.6 ± 0.22 |
| Самка | 32.9 ± 2.15 | 4.0 ± 0.25 | * | |
| C. griseus | Самец | 37.7 ± 1.38 | 6.33 ± 0.21 | * |
| Самка | 33.4 ± 1.3 | 2.75 ± 0.19 | * | |
Морфологическое описание волос и их структур соответствует общепринятой терминологии (Соколов, 1973; Соколов и др., 1988, рис. 83 ; Чернова, Целикова, 2004). Выделенные категории волос включают направляющие (самые длинные и толстые, цилиндрические, образующие ворс шерсти), остевые (как правило, имеющие расширение стержня – “гранну” в верхней половине или трети стержня) и пуховые волосы нескольких размерных порядков в зависимости от размеров и конфигурации стержня. Термин “дифференциация волосяного покрова” обозначает подразделение массы шерсти на несколько ярусов и наличие в ней волос разных категорий и порядков. Кольцевидная кутикула — одна чешуйка полностью оборачивает стержень. Лентовидная кутикула лежит поперек стержня в виде узкой ленты.
РЕЗУЛЬТАТЫ
Строение кожи. Кожа холки и крестца изученных представителей Allocricetulus и Cricetulus устроена сходно — непигментированная, тонкая (не более 226 ± 23 мкм) складчатая с тонким эпидермисом (до 23 ± 1 мкм), больше половины толщины которого составляет роговой слой (рис. 1А). Слои эпидермиса выражены слабо. Клетки росткового слоя крупные и светлые, с крупными светлыми овальными ядрами, лежащими на разных уровнях и не образующими сплошного базального ряда. Слой шиповатых клеток однорядный, клетки отличаются уплощенным ядром. Зернистый слой прерывистый, ядра его клеток темные, сморщенные, продолговатые. Дермальный слой, образующий основу кожи, состоит из сосочкового слоя, в котором залегают волосяные фолликулы и продольно вытянутые сильно извитые тонкие пучки коллагеновых волокон. Сетчатый слой не развит, сосочковый слой подстилается утолщенной (до 113 ± 45 мкм) подкожной клетчаткой. Граница между этими слоями нечеткая. Клетки подкожной клетчатки крупные — до 23 ± 1 × 34 ± ± 2 мкм. Под слоем подкожной клетчатки располагается мощный мышечный пласт толщиной до 339 ± 56 мкм. Луковицы крупных волос залегают в подкожной жировой клетчатке. Волосы располагаются пучками, а пучки собраны в группы по три–четыре в каждой. Фолликулы волос тянутся в подкожной жировой клетчатке и дермальном слое разрозненно, но в верхней трети они объединяются в одной расширенной (до 91 ± 9 мкм) волосяной сумке и выходят на поверхность кожи пучком (рис. 1А). На холке в состав пучка входят до четырех–пяти некрупных непигментированных волосков сходной толщины (11–13 мкм), с хорошо различимой пигментированной однорядной лестничной сердцевиной. Сердцевина занимает до 80% толщины стержня. На крестце в одном пучке собраны два–три крупных пигментированных волоса (диаметром до 17 ± 3 мкм) и до восьми мелких непигментированных волосков (до 11 мкм). Сальные железы развиты умеренно, они почти полностью окружают волосяной фолликул. Например, вдоль крупного волосяного фолликула длиной 113 мкм лежат две продольно вытянутые лишенные пигмента сальные железы размером 56 × 23; 56 × 34 мкм. Причем сальные железы залегают довольно глубоко от поверхности кожи (до 79 мкм).
Рис. 1.
Микроструктура кожи и желез взрослой самки Allocricetulus curtatus. A, C, D – сагиттальные срезы; B, E, F, G – трансверсальные срезы. A – холка; B – угол рта; C – бок с утолщенной зоной линьки (слева) и железистым полем (справа); D – боковая железа; E, F, G – фрагменты брюшной железы. 1 – эпидермис, 2 – дерма, 3 – подкожная жировая клетчатка, 4 – волос, 5 – альвеола сальной железы, 6 – проток железы, 7 – подкожная мускулатура. Микрофото. Окраска – гематоксилин Эрлиха-эозин. Масштаб 100 мкм.
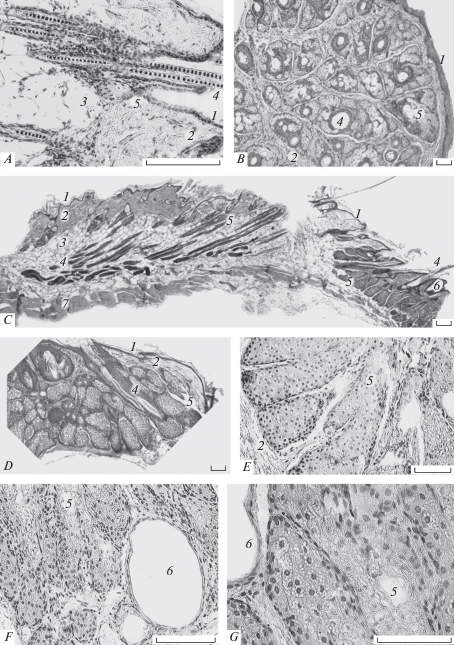
Строение кожи имеет особенности в топографически различных отделах туловища и конечностей, особенно в области залегания СКЖ. Так, у A. curtatus и C. sokolovi в углах рта присутствует утолщенная полоска кожи с растущими редкими белыми волосками. Эпидермис этой кожи (толщиной до 79 ± 6 мкм) на отдельных участках имеет упорядоченные утолщения-столбики и короткие врастания в дерму, что характерно для утолщенной кожи, постоянно подвергающейся механической нагрузке. Его основу составляет трех–четырехрядный слой шиповатых клеток, а роговой слой тонкий (не более 6 мкм). Имеется прерывистый однорядный слой зернистых клеток. Волосы растут поодиночке, а не пучками как на туловище. Каждый волос снабжен многодольчатыми увеличенными (до 350 × 245 мкм, у взрослой самки A. сurtatus) типичными сальными железами, которые отличаются длинными постепенно сужающимися или сильно расширенными протоками. Сальные железы плотно сомкнуты и образуют железистое поле (рис. 1B), размеры которого на трансверсальном срезе у взрослой самки A. curtatus достигают 1.9 ± 0.1 × 1.4 ± 0.5 мм. У C. griseus это утолщение имеет треугольную форму, на нем растут очень короткие светлые волоски, сальные железы которых незначительно увеличены (124 × 79; 214 × 23 мкм).
На мозолях подошв стоп эпидермис в несколько раз толще (до 169 ± 23 мкм у A. curtatus), чем на других участках тела, в основном за счет интенсивно отслаивающегося рогового слоя (до 115 ± ± 23 мкм). Сосочки или ячеи на внутренней поверхности эпидермиса не выражены. Имеются трех–четырехрядный слой шиповатых клеток, сплошной однорядный зернистый слой и узкий (не толще 6 мкм) блестящий слой. Мозоль образована сетчатым слоем дермы, представляющим собой плотную вязь утолщенных пучков коллагеновых волокон. В центральной части мозоли присутствуют жировые клетки, вязь пучков волокон становится рыхлой. Здесь залегают извитые и закрученные в рыхлые клубки эккриновые железы, но у зимних зверьков их секреторные отделы спавшиеся и функционируют слабо.
Волосяной покров кожи туловища всех изученных видов относительно невысокий, не имеет длинного ворса и пуховых волос, слабо разделен на ярусы.
У A. curtatus и A. eversmanni высота волосяного покрова холки одинаковая (10.0 ± 0.5 мм). У обоих видов хорошо различимы редкие черные кончики направляющих волос, на 0.5–1.0 мм выступающие над общим уровнем шерсти. Большая часть волос двуцветная: у A. curtatus нижние 2/3 стержней темно-серые, верхняя треть – от светло-серой до белой; у A. eversmanni нижние 3/4 стержней темно-серые, а верхняя четверть светло-желтая. Направляющие волосы (толщиной 34.0 ± 0.5 мкм) отличаются не только толщиной, но и двух–трехрядной пигментированной сердцевиной и черной заостренной вершиной. Имеются остевые волосы трех размерных порядков, различающиеся только толщиной стержня, соответственно 23.0 ± 0.5; 17.0 ± 0.2 и 11.0 ± 0.1 мкм. У всех волос стержень прямой без изгибов и четко выраженной гранны. Он имеет тонкую прозрачную ножку, плавно расширяется в верхней части стержня, а заканчивается тонкой и длинной вершиной, не содержащей пигмента, за исключением направляющих волос. Самое основание волос, а также их вершины лишены сердцевины и пигмента.
Волосяной покров брюшка у A. curtatus по высоте такой же, как на холке, но окрас распределен иначе: нижняя половина волос светло-серая, а верхняя – светло-желтая. У A. eversmanni высота шерсти здесь не превышает 8 ± 1 мм, а окрас волос также двуцветный: на нижней половине стержня он темно-серый, а на верхней – белый. У обоих видов на брюшке отсутствуют длинные направляющие с черными вершинами. Размерные порядки такие же, как на загривке. У A. eversmanni встречаются прямые направляющие толщиной до 45 ± 1 мкм. Стержень некоторых остей II и III в нижней части волнообразно изогнут (две-три мелкие волны).
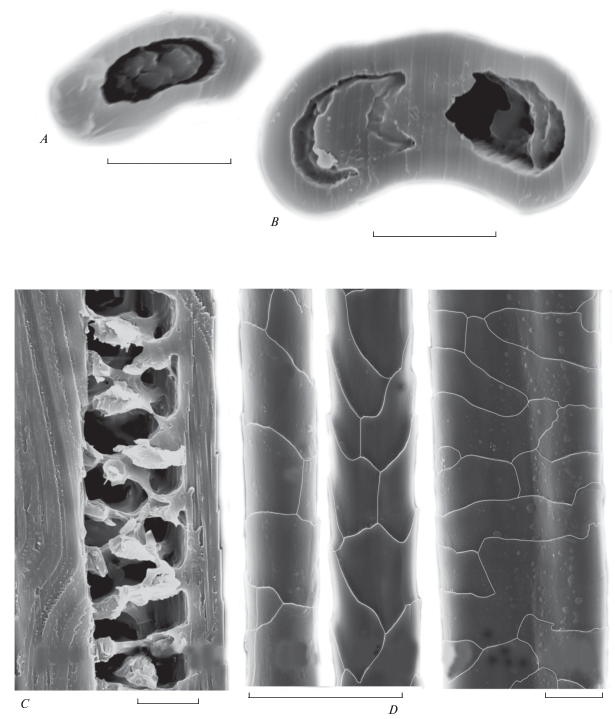
По данным СЭМ, у Allocricetulus на холке направляющие волосы имеют правильное цилиндрическое основание, но выше по стержню волос становится незначительно уплощенным, и его поперечник приобретает неправильную округлую форму (рис. 2А), а в зоне расширения стержня вентральная сторона волоса еще сильнее уплощается и поперечник становится овальным (рис. 2B). На брюшке направляющие волосы снабжены неглубокой продольной бороздкой на вентральной (направленной к поверхности тела) стороне стержня. Бороздка тянется от основания до гранны (рис. 3А). У всех направляющих волос сердцевина хорошо развита, изменяется от однорядной лестничной в основании стержня до двух–трехрядной (рис. 2C; 3B). Сердцевина в направляющих волосах загривка занимает от 57.0 ± ± 5.5% (максимум 67%) в основании стержня до 58.3 ± 16.7% (до 86%) толщины в расширенной части стержня. В направляющих волосах брюшка — соответственно 48.5 ± 6.2% и 70.5 ± 10.3%. У более тонких волос сердцевина только однорядная. В сердцевинной полости ороговевшие “клетки” − диски лежат упорядоченно, они разделены воздушными полостями такой же толщины, как и диск, лежат в один ряд или чередуясь в зависимости от толщины стержня (рис. 2С; 3B). Сам диск заполнен многочисленными овальными пигментными гранулами, что хорошо различимо на трансверсальных срезах (рис. 2A). Орнамент кутикулы неодинаков по стержню: наиболее крупные продольно вытянутые узкие чешуйки покрывают основание стержня, но к середине стержня они незначительно уплощаются и вытягиваются поперек волоса (рис. 2D; 3C).
Рис. 2.
Строение направляющего волоса холки взрослой самки Allocricetulus curtatus: A, B – трансверсальные срезы соответственно в основании и середине стержня; C – сагиттальные срезы стержня в основании и выше (слева-направо); D – кутикула от основания к середине стержня (слева-направо). СЭМ. Масштаб 10 мкм.
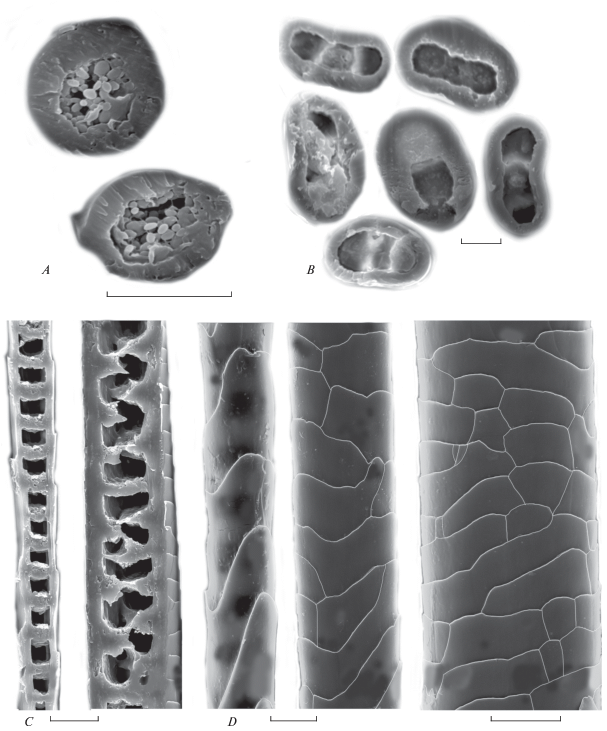
Рис. 3.
Строение направляющего волоса брюшка взрослой самки Allocricetulus curtatus: A, B – трансверсальные срезы в основании и середине стержня (слева-направо), С – сагиттальный срез середины стержня, D – кутикула от основания к середине стержня (слева-направо). СЭМ. Масштаб 10 мкм.
У C. sokolovi высота волосяного покрова достигает 13.0 ± 0.5 мм. Соотношение цветовых зон на ости первого порядка равно 4/5 (темно-серая) и 1/5 (светло-желтая). Имеются немногочисленные направляющие c черными кончиками, незначительно выступающими над остальной шерстью. Размерные порядки и конфигурация волос такие же, как у представителей Allocricetulus, однако продольная бороздка имеется у волос не только брюшка, но и загривка. Волосяной покров брюшка отличается от такового на холке как по высоте (8.0 ± 0.5 мм), так и по окрасу. Направляющие c черными кончиками отсутствуют. Окрас стержня двуцветный: нижняя половина темно-серая, верхняя половина – белая.
У C. griseus высота шерсти на холке составляет 11.0 ± 0.5 мм. Имеются редкие направляющие c черной вершиной. Размерные порядки такие же, как у других видов. Окраска волос двуцветная: нижние 2/3 волоса темно-серые, а верхняя треть – светло-желтая. На брюшке высота основной массы волос равна 7.0 ± 1.0 мм. Белые направляющие немного выступают над уровнем шерсти. Черные вершины у них отсутствуют. Окраска волос двуцветная: нижняя половина темно-серая, верхняя половина волоса – белая. Размерные порядки не отличаются от таковых на загривке. Продольная бороздка на стержне волос загривка и брюшка имеется.
У обоих видов сердцевина остей развита так же хорошо, как у волос Allocricetulus: в основании стержня она однорядная лестничная и занимает до половины толщины стержня, а в расширенной части волоса – двух-трехрядная, составляет до 90% толщины стержня.
Вибриссы у Allocricetulus имеют правильную цилиндрическую или немного уплощенную конфигурацию, снабжены слабо развитой сердцевиной, занимающей до 20% стержня. Граница между сердцевинным каналом и корой неровная. Строение сердцевины вибрисс отличается от таковой у обычных волос. У A. curtatus она имеет хаотично разбросанные неперфорированные перегородки (рис. 4А). Кутикула однородная вдоль стержня, уплощенная лентовидная (тянется поперек стержня в виде узкой ленты) и кольцевидная (одна чешуйка полностью оборачивает стержень). Ее орнамент изменяется лишь на вершине вибриссы, где чешуйки крупнее и сильно повреждены.
Рис. 4.
Строение вибрисс-усов на сагиттальных срезах (A–C) и кутикулы (D–F) волос брюшной железы: А – взрослая самка Allocricetulus curtatus; B – основание вибриссы взрослого самца Cricetulus griseus; C – то же в середине стержня вибриссы; D, F – взрослый самец C. griseus; E – разрушенный стержень волос СБЖ взрослого самца C. sokolovi. СЭМ. Масштаб 10 мкм.
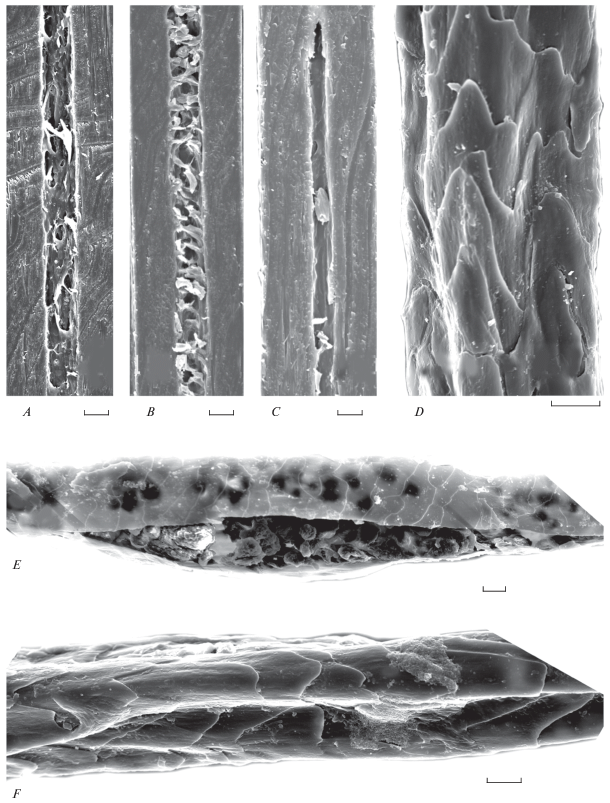
У C. griseus вибриссы со слабо развитой сердцевиной (21% толщины стержня), строение которой отличается от таковой обычных волос. В нижних отделах стержня внутренние перегородки сердцевинного канала располагаются достаточно хаотично (рис. 4B), а выше по стержню они вообще отсутствуют (рис. 4C). Кутикула такая же, как у волос Allocricetulus.
Среднебрюшная железа (СБЖ) имеется у всех исследованных в данной работе видов, ее локализацию легко определить по более темному цвету шерсти и желтовато-буроватому секрету на брюшной части тела. На живой особи можно оценить длину СБЖ (табл. 1) и ее активность по относительному количеству секрета. Однако на препарате или со стороны мездры измерения и описания можно выполнить более точно.
У обоих видов Allocricetulus СБЖ вытянута вдоль срединной медиальной части брюшка и оформлена в виде органа. Имеет форму груши с центральной щелевидной полостью; полость окружена валиками, которые толще на каудальном конце органа (рис. 6). Дно щели заполнено коричневатым секретом. На валиках растут редкие и короткие бесцветные волоски толщиной не более 17 ± 1 мкм. Они имеют однорядную непигментированную сердцевину, их стержни подвергаются лизису, а остатки входят в состав секрета СБЖ. В области СБЖ кожа утолщена до 904 мкм за счет дольчатой железистой ткани, но примыкающая к железе кожа гораздо тоньше (до 113 ± ± 45 мкм), и здесь луковицы волос с мелкими сальными железками не проникают в нижележащие слои. Эпидермис над железой утолщен (до 68.0 ± 0.6 мкм). Его сильно отслаивающийся роговой слой заполняет глубокие складки кожи, в которые выходят многочисленные крупные протоки сальных желез. В центральной складчатой щели толщина отслаивающегося рогового слоя достигает 110 ± 11 мкм. Слой шиповатых клеток состоит из двух–трех рядов. Имеется прерывистый зернистый слой. Волосы редкие, залегают глубоко в железистой ткани. Жировая клетчатка не развита, и донная часть железы непосредственно примыкает к продольным подкожным мышечным тяжам. Многочисленные кровеносные капилляры окружают и пронизывают железу. Строение отдельных желез – сложное альвеолярное с разветвленной системой депонирования и выведения секрета (рис. 1E, 1G; 5A). Форма долей, вытянутых на всю толщину железистой паренхимы, меняется от округлой до многодольчатой цилиндрической. Железистая паренхима вплоть до донной части пронизана крупными расширенными (диаметром до 125 ± 11 мкм) выводными протоками, выстланными одно–двуслойным ороговевающим эпителием (рис. 1C; 5A). У A. eversmanni расширенные протоки сливаются в центральный резервуар (шириной до 565 ± 17 мкм), который содержит отслоившийся эпидермис, фрагменты волос и слоистый эозинофильный секрет (рис. 5A). Секреторный эпителий в большинстве альвеол демонстрирует типичную голокриновую секрецию, однако в дистальных отделах некоторых альвеол процесс жирового перерождения опережает дегенерацию ядер. В цитоплазме таких альвеол присутствуют многочисленные жировые вакуоли и крупные светлые и округлые ядра без признаков деградации, что придает железе “ажурный” вид (рис. 1D).
Рис. 5.
Микроструктура среднебрюшной железы взрослых самцов Allocricetulus eversmanni (A) и Cricetulus griseus (B, D хомячков): A – трансверсальный срез донной части железы, B–D – сагиттальные срезы. Условные обозначения и окраска как на рис. 1. Микрофото. Масштаб 100 мкм.
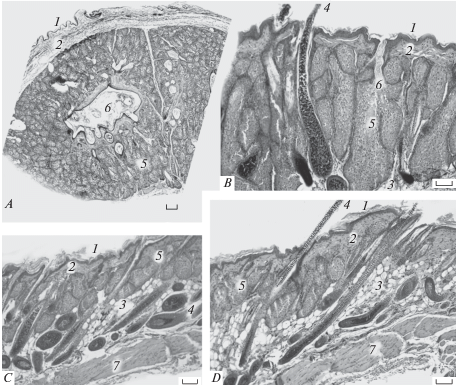
Рис. 6.
Расположение и внешний вид среднебрюшной железы (указана стрелкой) взрослого самца Allocricetulus curtatus. Фото.
У С. griseus и С. sokolovi СБЖ не оформлена в орган (рис. 7A, 7B). Она не выступает над поверхностью кожи и представляет собой железистое поле, на котором растут короткие черные волоски, резко отличающиеся по цвету, размерам и конфигурации стержня от серовато-белых волосков брюшка. Эти черные волоски многочисленные и длинные (до 8 мм). Они образуют густую, обильно покрытую прозрачным секретом “щетку”. Их цилиндрический слабоизогнутый стержень ломкий и имеет многочисленные повреждения. Вдоль всего стержня проходит бороздка (рис. 4F). Волоски выходят пучком из расширенных волосяных воронок вместе с протоками желез (рис. 5B). Конфигурация стрежня этих волосков своеобразная: стержень длиной до 3.0 ± 0.5 мм и толщиной 79–113 мкм имеет длинные непигментированные основание и шиловидную вершину, заканчивающуюся крючочком. Сам стержень сильно пигментированный, четковидный – неравномерной толщины, во многих местах он сильно разрушен, имеет признаки лизиса, и его структуры – сердцевинные перегородки и пигментные гранулы – высыпаются из стержня (рис. 4E).
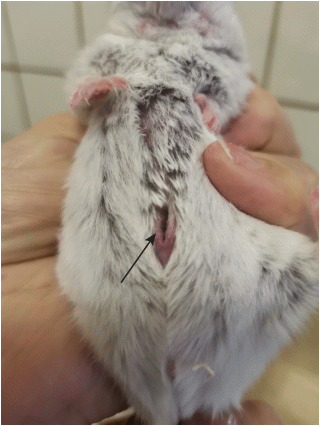
Рис. 7.
Расположение и внешний вид среднебрюшной железы (указана стрелками) взрослой самки (A) и самца (B) Cricetullus sokolovi. Фото.
У обоих видов рода Cricetulus строение волос СБЖ не отличается от таковой обычных волос, продольная бороздка имеется, но кутикула своеобразна (рис. 4D, 4F). Она образована крупными грубыми, толстыми и изломанными высокими чешуйками, на поверхности которых присутствует секрет железы. У С. griseus толщина железистого слоя на сагиттальных срезах органа достигает 1.3 ± 0.2 мм. В его состав включены многочисленные сильно пигментированные и дифференцированные луковицы волос (рис. 5B). Причем многие из них искривлены и подвергаются лизису. Диаметр фолликулов ости I достигает 134 ± 11 мкм. Сами сальные альвеолы имеют многогранную форму, вытянуты вглубь на всю толщину кожи, полиморфны, разделены толстыми соединительнотканными прослойками. Донная часть железы подстилается узким слоем жировой подкожной клетчатки, лежащей на мощных продольных мышечных тяжах. Во многих альвеолах секреторный процесс проходит типично, но не столь интенсивно, как в СБЖ A. curtatus. Здесь также отсутствуют клетки с крупными вакуолями. Имеются расширенные протоки в нижних отделах железы. В глубоко залегающих альвеолах хорошо различимы межклеточные канальцы с непрозрачной частично базофильной цитоплазмой, округлыми ядрами, что предполагает присутствие в донной части железы гепатоидных альвеол.
Боковые железы (БЖ) внешне хорошо различимы у светлоокрашенного монгольского хомячка (рис. 8А) и несколько хуже у темного хомячка Эверсманна (табл. 1). Но со стороны мездры области желез хорошо видны в виде бурых пятен у обоих видов Allocricetulus. В этом месте выявлено присутствие железистой ткани и окружающего ее толстого ободка из крупных пигментированных волос, искривленные луковицы которых погружены в сильно развитую жировую клетчатку (рис. 1C). В области ободка толщина кожи достигает 1.4 ± 0.2 мм за счет жировой клетчатки и луковиц растущих волос, в то время как в области железистого поля она вдвое меньше – 0.7 ± 0.1 мм. У самки на сагиттальном срезе кожи размеры железистого поля составляют: толщина 0.7 ± 0.5 мм; длина 1.4 ± 0.2 мм. В области БЖ кожа сильноскладчатая с неравномерно утолщенным (до 34.0 ± ± 0.6 мкм) непигментированным эпидермисом, а жировая подкожная клетчатка не развита. Железы типичные сальные, многодольчатые; отдельные альвеолы удлиненные пирамидальные с широким основанием (226 ± 34 × 113 ± 23 мкм) (рис. 1C). Система выводных протоков не развита, крупные протоки и резервуары отсутствуют. Секреторная активность железы низкая. Узкий выводной проток каждой сальной железы выходит в широкую волосяную воронку пучка тонких волос (рис. 1D).
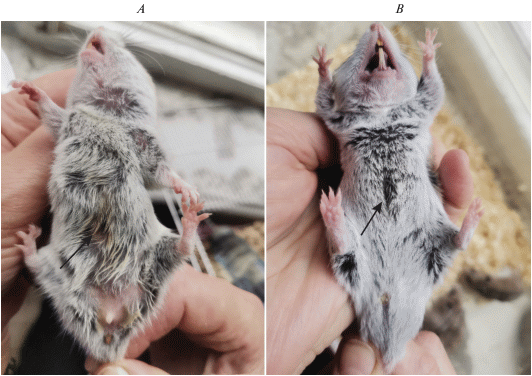
Рис. 8.
Расположение и внешний вид правосторонней боковой железы (указана стрелкой) у взрослого самца Allocricetulus curtatus. Фото.

У C. sokolovi БЖ хорошо различимы как внешне (рис. 9A), так и со стороны мездры (рис. 9B). У самцов C. griseus БЖ снаружи малозаметны из-за густой и однотипной шерсти. Но со стороны мездры здесь также с обеих сторон симметрично расположены округлые железы диаметром 5.0 ± ± 0.5 мм. У самца каждая железа окружена черным ободком из сильно пигментированных интенсивно растущих крупных луковиц волос, глубоко залегающих в хорошо развитой подкожной жировой клетчатке (рис. 5D). Сетчатый слой дермы также сильно пигментирован в центральной части органа. Толщина железистого слоя составляет 0.80–1.75 мм, он занимает всю толщу дермы до жировой клетчатки. Крупные волосяные луковицы пронизывают всю железистую паренхиму. Доли сальных желез вытянутые цилиндрические или округлые, почти не ветвящиеся (рис. 5C, 5D). Их размеры достигают 700 ± 45 × 210 ± 11 мкм. Протоки желез не расширенные, впадают в верхние отделы волосяных луковиц. Секреторная активность желез низкая. На сагиттальных срезах толщина кожи здесь достигает 770 ± 35 мкм, а железистый слой занимает лишь половину толщины кожи. Имеются утолщенные участки непигментированного складчатого эпидермиса, в основном за счет рогового слоя и слоя шиповатых клеток. Зернистый слой отсутствует. Подкожная клетчатка развита слабо, подстилается мощным тяжом подкожной мускулатуры.

ОБСУЖДЕНИЕ
Морфология кожи исследованных нами видов хомячков вполне характерна для других представителей подсемейства Cricetinae. Опушенная кожа туловища имеет все признаки тонкого интегумента: складчатая, непигментированная, со слабо выраженными слоями эпидермиса и дермы, с хорошо развитой подкожной клетчаткой, в которую проникают луковицы волос, растущих “ложными пучками.” Луковицы волос закладываются обособленно, но их стержни выходят из общей волосяной сумки. Отличительной чертой кожи хомячков служит глубокое проникновение жировой подкожной клетчатки в дермальный слой, а также залегание луковиц крупных волос в жировом слое, благодаря чему четкая граница между дермой и подкожной жировой клетчаткой не формируется, как у большинства наземных млекопитающих (Соколов, 1973). Шерсть изученных видов хомячков слабо подразделяется на ярусы и категории, а сравнение с другими представителями трибы Cricetini показывает, что она устроена все же сложнее, чем у Phodopus, у которого не обнаружены направляющие волосы (Феоктистова, 2008). Волосяной покров хомячков в целом более однороден, чем у многих других млекопитающих, например у Mus musculus, у которой имеются четыре типа волос, в том числе зигзагообразные и “awl/auchene” (см., например, Duverger, Morasso, 2009). Это может свидетельствовать об их более древнем происхождении и норном образе жизни, при котором длинный ворс мешал бы их перемещению по туннелям. В то же время сильное развитие сердцевины волос всех категорий предполагает хорошие теплозащитные свойства шерсти хомячков, что позволяет им адаптироваться к существованию в широком диапазоне климатических условий Палеарктики.
Вибриссы хомячков родов Allocricetulus и Cricetulus специализированы, в отличие от Phodopus, у которого по архитектонике они не отличаются от обычных остевых волос (одно- или двурядная лестничная сердцевина). Вероятно, в подсемействе Cricetinae процесс дивергенции сопровождался и процессом специализации структуры вибрисс, по-видимому, в связи с особенностями образа жизни и эволюционной историей. Более упрощенный внутренний дизайн вибрисс Phodopus, по сравнению с Allocricetulus и Cricetulus, согласуется с молекулярно-генетическими данными о том, что род Phodopus — наиболее древний из рецентных родов Cricetinae (Lebedev et al., 2018).
Железы в углах рта увеличены умеренно, имеют типичное для большинства грызунов расположение в виде железистого поля на полоске слабоопушенной кожи, вдающейся в слизистую углов рта (Quay, 1965; Соколов, 1973; Соколов, Чернова, 2001). Однако у исследованных видов они не достигают такого сильного развития с формированием мешотчатого органа, как у P. sungorus и P. campbelli (Соколов и др., 1993; Феоктистова, 2008). Утолщенный эпидермис этих желез характеризуется наличием упорядоченных столбчатых выростов, внедряющихся в дерму (сосочковидный эпидермис), что характерно для кожи, постоянно подвергающейся механической нагрузке при жевании и наполнении защечных мешков.
Подошвенные железы также типичные, но слабо функционирующие. Голая кожа подошв лап характерна для утолщенного интегумента, но здесь, в отличие от подошвенной кожи многих других видов грызунов, сосочковидное или ячеистое сочленение эпидермиса и дермы не развито (Соколов, 1973).
Среднебрюшная железа у всех исследованных видов родов Allocricetulus и Cricetulus СБЖ активно функционирует, и ее строение специфично для рода. У Allocricetulus СБЖ сходна с таковой Phodopus – кожный орган мешковидной конфигурации, выступающий над поверхностью кожи и имеющий центральную полость и утолщенные стенки. Однако у Phodopus боковые валики и дно железы покрыты специализированными волосами, возможно, принимающими участие в маркировочном поведении. СБЖ Allocricetulus лишена специализированных волос. У Phodopus и Allocricetulus она состоит из сложных полиальвеолярных сальных желез с разветвленной системой депонирования и выведения секрета при участии хорошо развитой подкожной брюшной мышечной системы, подстилающей дно железы. Такое строение предполагает периодичность выделения сложного экскрета: смеси секрета сальных желез, растворенных остатков волос и их пигмента, а также отслаивающихся клеток эпидермиса, которые могут составлять существенную часть его объема. Мечение СБЖ описано для Phodopus и Allocricetulus и представляет собой проволакивание брюшной области по субстрату, объектам среды (камни, ветки) или по телу других особей (Суров, 2006; Феоктистова, 2008). У Cricetulus СБЖ представляет собой лишь небольшое железистое поле, покрытое “щеткой” специализированных волос. СБЖ у видов этого рода отличается отсутствием специализированных сильно пигментированных крупных волос и слабо развитой депонирующей и транспортной системой, в том числе мышечной подстилкой. По-видимому, у представителей этого рода выделение секрета железы или отделение его от специализированных волос железистого поля происходит лишь при трении волосяной щетки о субстрат или сородичей.
Специализированные волосы СБЖ Phodopus и Cricetulus также различаются. У Phodopus боковые стенки-валики сумки покрыты обычными волосами, но на дне сумки растут особенные короткие (не более 2–5 мм), непигментированные или слабопигментированные копьевидные волоски, обильно покрытые желтоватым секретом. Короткая тонкая ножка волоса переходит в утолщенный ланцетовидный стержень (толщиной 34–58, иногда до 170 мкм), постепенно вытягивающийся в длинный, узкий, шиловидно заостренный кончик. Вдоль по стержню проходит желобок, что способствует выводу секрета СБЖ. Сердцевина и кутикула такие же, как у обычных волос (Феоктистова, 2008). У Allocricetulus и Cricetulus волосы СБЖ специализированы в большей степени, помимо бороздки для сбора секрета, они имеют специфичную кутикулу, а также характеризуются способностью к лизису и высвобождению структур сердцевины в секрет.
Таким образом, экскрет СБЖ представляет собой комплексное образование, в котором масса сального вещества смешивается с фрагментами волос (в большей степени характерно для Cricetulus), подвергшихся лизису и механическому истиранию, и c многочисленными гранулами пигмента, высвобождающегося в результате этого лизиса, а также с отшелушившимся роговым слоем эпидермиса. Вкупе эти ингредиенты значительно увеличивают объем экскрета и придают ему окраску и, вероятно, специфический запах, возможно, в результате действия симбиотических бактерий. Неудивительно, что фолликулы этих волос постоянно интенсивно растут, что приводит к их деформации.
Выявленные различия в морфологии СБЖ у представителей разных родов Cricetinae ставят под сомнение принятое в литературе клише для описания СБЖ. Обычно указывают на присутствие железы и на ее “типичное” строение, хотя теперь ясно, что как раз строение может различаться у представителей разных родов. По-видимому, назрела необходимость ревизии строения СБЖ. У многих видов остаются также невыясненными внутривидовой полиморфизм этой железы и сезонность ее функционирования.
Боковые железы представляют собой симметричные железистые поля значительной толщины, покрытые сильноскладчатым эпидермисом с активно растущими и сильно пигментированными волосами. Железистое поле состоит из плотно сомкнутых цилиндрических, практически не разветвленных желез, не имеющих сложной системы транспортировки и депонирования секрета. Исследователи, которые наблюдали маркировочное поведение C. griseus, предполагали социальную значимость секрета этих желез (Skirrow, Rysan, 1976). Однако описанное ими поведение мечения скорее можно интерпретировать как оставление на субстрате секрета не БЖ, а именно СБЖ, которая, как показано нами, развита у этого вида даже лучше БЖ.
Строение и степень развития БЖ у представителей родов Allocricetulus и Cricetulus несколько различаются. У первого каждая из желез представляет собой железистое поле небольших размеров, которое не содержит крупных пигментированных волос (хотя в примыкающей коже они имеются); доли желез имеют не цилиндрическую, а пирамидальную форму; система протоков не развита; пигментация кожи здесь отсутствует. Как и у СБЖ, мышечная подстилка железистой паренхимы хорошо выражена.
ЗАКЛЮЧЕНИЕ
При проведении исследования и сравнительного анализа строения СБЖ у разных представителей подсемейства Cricetinae выявлено три морфотипа – железа-сумка без специализированных волос (у Allocricetulus), железа-сумка со специализированными волосами (у Phodopus) и железа-поле со специализированными волосами (у Cricetulus). У Phodopus и Allocricetulus мечение железой проходит по типу оставления пахучей ”метки-печати” − оттиска с безволосой кожи на субстрате или сородичах, что обеспечивается сумчатым строением СБЖ с дозированным исторжением секрета. У Cricetulus – посредством кисти-помазка из покрытых секретом специализированных, чрезвычайно ломких волос. Все это позволяет говорить о разнообразии строения СБЖ у Cricetinae и необходимости ревизии сведений о ее морфологии.
Впервые у представителей Allocricetulus описаны БЖ, располагающиеся билатерально на боковой поверхности тела ближе к задней конечности (косто-вертебральный угол). Полученные морфологические данные дают более полное представление о кожно-волосяном покрове и степени развития СКЖ у представителей подсемейства Cricetinae. Они послужат основой для анализа экспериментальных этологических и физиологических исследований, удобными объектами которых являются некрупные и простые в содержании виды хомяков.
Не исключено, что детальное изучение морфологии кожных покровов и СКЖ у Cricetinae позволит, с одной стороны, подтвердить (или опровергнуть) данные о филогенетических отношениях родов, с другой, – выстроить картину эволюционных изменений кожно-волосяного покрова и его дериватов внутри этой таксономической группы.
Список литературы
Бодяк Н.Д., 1994. Гардеровы железы млекопитающих. Структура и функции // Известия РАН. Серия биологическая. Т. 21. № 2. С. 183–197.
Васильева Н.Ю., 1990. Функция специфических кожных желез. Функциональные и эволюционные аспекты маркировочного поведения грызунов на примере представителей п/сем. Cricetinae. Автореф. дис. … канд. биол. наук. М.: Институт эволюционной морфологии и экологии животных им. А.Н. Северцова РАН. 28 с.
Воронцов Н.Н., 1982. Низшие хомякообразные мировой фауны. Фауна СССР. Млекопитающие / О.А. Скарлато, ред. Ч. I. Морфология и экология Л.: Наука. Ленинградское отделение. 451 с.
Воронцов Н.Н., Гуртовой Н.Н., 1959. Строение среднебрюшной железы настоящих хомяков // ДАН СССР. Т. 125. № 3. С. 673–676.
Кропоткина М.В., Феоктистова Н.Ю., Найденко С.В., 2012. Сезонные особенности гормонального ответа самок двух видов хомячков рода Phodopus на химические сигналы самцов-конспецификов // Известия РАН. Серия биологическая. № 3. С. 312–319.
Малыгин В.М., Старцев Н.В., Зима Я., 1992. Кариотипы и распространение видов хомячков группы barabensis (Rodentia, Cricetinae) // Вестник Московского Университета. Серия биологическая. № 2. С. 32–39.
Павлинов И.Я., 2006. Систематика современных млекопитающих (2-е изд.). М.: Издательство Московского университета. 297 с.
Поташникова Е.В., Феоктистова Н.Ю., 2014. Сезонные особенности гормонального ответа самцов китайского хомячка (Cricetulus b. griseus) на химические сигналы самок-конспецификов // Сенсорные системы. Т. 28. № 1. С. 89–94.
Рожнов В.В., 2011. Опосредованная хемокоммуникация в социальном поведении млекопитающих. М.: Товарищество научных изданий КМК. 288 с.
Соколов В.Е., 1973. Кожный покров млекопитающих. М.: Наука. 489 с.
Соколов В.Е., Бодяк Н.Д., Суров А.В., 1994. Возможная роль Гардеровой железы в химической коммуникации хомяка (Mesocricetus auratus Waterhouse, 1839) // Известия Российской Академии наук. Серия биологическая. № 6. С. 880–888.
Соколов В.Е., Чернова О.Ф., 2001. Кожные железы млекопитающих. М.: ГЕОС. 648 с.
Соколов В.Е., Скурат Л.Н., Степанова Л.В., Шабадаш С.А., 1988. Руководство по изучению кожного покрова млекопитающих. М.: Наука. 279 с.
Соколов В.Е., Феоктистова Н.Ю., Мещерский И.Г., 1993. Влияние специфических образований в углах рта на физиолого-морфологические параметры взрослого организма хомячка Кэмпбелла (Phodopus campbelli, Thomas 1905) // Доклады РАН. Т. 330. № 5. С. 667–669.
Суров А.В., 2006. Обонятельные сигналы в половом поведении млекопитающих. Дис. … докт. биол. наук. М.: Институт проблем экологии и эволюции им. А.Н. Северцова РАН.
Феоктистова Н.Ю., 2008. Хомячки рода Phodopus. Систематика, филогеография, экология, физиология, поведение, химическая коммуникация. М.: Товарищество научных изданий КМК. 414 с.
Феоктистова Н.Ю., Ушакова Н.А., Мещерский И.Г., 2004. Участие симбиотических бактерий, обитающих в специфических комплексах в углах рта, в процессах нормальной жизнедеятельности у хомячка Кэмпбелла (Phodopus campbelli) // Доклады Академии наук. Т. 398. № 3. С. 426–429.
Феоктистова Н.Ю., Чернова О.Ф., Мещерский И.Г., 2012. Декоративные формы хомячков рода Phodopus (Mammalia, Cricetidae) – анализ распространения генетических линий и особенности изменения волосяного покрова // Журнал общей биологии. Т. 73. № 2. С. 134–150.
Чернова О.Ф., Целикова Т.Н., 2004. Атлас волос млекопитающих (Тонкая структура остевых волос и игл в сканирующем электронном микроскопе). М.: Товарищество научных изданий КМК. 429 с.
Хрущова А.М., 2007. Роль специфических экскреторных образований кожи в регуляции развития на примере хомячков р. Phodopus. Автореф. дис. … канд. биол. наук. M: Институт проблем экологии и эволюции им. А.Н. Северцова РАН. 26 с.
Duverger O., Morasso M., 2009. Epidermal patterning and induction of different hair types during mouse embryonic development // Birth Defects Research. Part C. Embryo. V. 87. (3). P. 263–272.
Kral B., Radjabli S.I., Grafodatskij A.S., Orlov V.N., 1984. Comparison of karyotypes, G bands and NORs in three Cricetulus spp. (Cricetidae, Rodentia) // Folia Zoologica. V. 33 (1). P. 85–96.
Lai S.C., Vasilieva N.Yu., Johnston R.E., 1996. Different odors providing sexual information in Djungarian hamsters: Evidens for across-odor code // Hormones and Behavior. V. 30. P. 317–323.
Lebedev V.S., Bannikova A.A., Neumann K., Ushakova M.V., Ivanova N.V., Surov A.V., 2018. Molecular phylogenetics and taxonomy of dwarf hamsters Cricetulus Milne-Edwards, 1867 (Cricetidae, Rodentia): description of a new genus and reinstatement of another. Zootaxa. V. 4387 (2). P. 331–349.
Lipkow J., 1954. Uber das Seitenorgan des Goldhamsters (Mesocricetus auratus auratus Waterh.) // Zeitschrift für Morphology und Ökologie der Tiere. Bd. 42. S. 333–372.
Pallas P.S., 1778. Novae species quadrupedum e glirium ordine cum illustrationibus variis complurium ex hoc ordine animalium. Erlangen: Wolfang Walther. 396 s.
Quay W.B., 1965. Comparative survey of the sebaceous and sudoriferous glands of the oral lips and angle in rodents // Journal of Mammalogy. V. 46. № 1. P. 23–27.
Romanenko S.A., Volobouev V.T., Perelman P.L., Lebedev V.S., Serdukova N.A. et al., 2007. Karyotype evolution and phylogenetic relationships of hamsters (Cricetidae, Muroidea, Rodentia) inferred from chromosomal painting and banding // Chromosome Research. V. 15. P. 283−297.
Skirrow M.H., Rysan M., 1976. Observation on the social behavior of the Chinese hamster, Cricetulus griseus // Canadian Journal of Zoology. V. 54. P. 361–368.
Vrtiš V., 1930. Glandular organ on the flanks of the hamster, Cricetus cricetus (L.) // Biologické spisy. Brünn: Vysoká Skola veterinari. T. 9. F. 13–2. P. 1–43.
Wyatt T.D., 2014. Pheromones and animal behavior: chemical signals and signatures. 2-nd edition. Cambridge: UK Cambridge University Press. 419 p.
Zhang J., Wang Z., Zhang Z., 1999. A preliminary study of effects of estradiol on flank glands in ratlike hamster (Cricetulus triton) // Acta theriologica sinica. V. 19. № 4. P. 262–266.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал