

Зоологический журнал, 2021, T. 100, № 12, стр. 1401-1421
Изменения состава и относительного обилия копытных Евразийской лесостепной зоны в голоцене
В. В. Гасилин a, *, М. М. Девяшин a, **, Н. А. Пластеева a, ***, А. Н. Созонтов a, ****
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: gasilinv@yandex.ru
** E-mail: devjashinm@yandex.ru
*** E-mail: natalya-plasteeva@yandex.ru
**** E-mail: a.n.sozontov@gmail.com
Поступила в редакцию 13.08.2020
После доработки 23.07.2021
Принята к публикации 29.07.2021
Аннотация
Обобщен и изучен свод литературных данных о видовом составе и количестве голоценовых костных находок копытных в пищевых остатках промысловой добычи, собранных при раскопках древних поселений лесостепной зоны Евразии. Использованы данные по остаткам копытных из более чем 570 археологических памятников. В результате анализа количественных показателей обнаружено, что высокое сходство видовых составов копытных лесостепных провинций в среднем–позднем голоцене сочеталось с различиями в относительном обилии фоновых видов – кабана (Sus scrofa L. 1758), благородного оленя (Cervus elaphus L. 1758), косули европейской (Capreolus capreolus L. 1758) и косули сибирской (C. pygargus Pallas 1771), лося (Alces alces L. 1758). Значимым оказался географический фактор широтного направления действия. Установлено, что в лесостепи от ее Паннонской островной провинции до Среднерусской провинции относительное обилие благородного оленя и кабана в голоцене выше, чем в лесостепи от Заволжской провинции до Обской. Состав фоновых видов лесостепи сохранился до текущего столетия.
Необходимость количественного анализа изменений последних тысячелетий в обилии животных, часть видов которых вымерла, связана с ускоренным ростом антропогенного влияния на природную среду и тем, что исчезновению вида предшествует продолжительная фаза его затухания. Источником данных для анализа прошлого современной фауны признаются результаты в области археозоологии.
В последнее столетие на территории бывшего СССР и соседних государств исследовано множество археологических памятников, небольшая часть материалов которых изучалась на предмет роли охоты и животноводства у древнего населения. Свод опубликованных данных о количестве остеологических остатков в культурных слоях древних поселений важных в хозяйственном отношении животных в настоящее время позволяет в общих чертах наметить основные направления изменений в фауне копытных млекопитающих умеренного пояса евразийского континента на протяжении голоцена.
Исследование определено природной зоной, поскольку известна зависимость копытных от типов растительности и преобладающих ландшафтов. Выбор лесостепной зоны обусловлен ее переходным характером между закрытым ландшафтом лесов и открытым ландшафтом степи, большим флористическим и фаунистическим богатством, позволяющим косвенно оценивать в смежных зонах распространенность видов, у которых здесь проходит граница ареала. Кроме того, вследствие видового разнообразия зона выделяется устойчивостью экосистем, которая иллюстрируется множеством растительных реликтов и рефугиумов, как открытых, так и закрытых ландшафтов прошлого.
В лесостепи обнаружено гораздо большее количество древних поселений от неолита до средних веков и нового времени, чем в любой из соседних зон. Вместе со степью она представляла коридор для переселений древних человеческих коллективов. Это объясняется ее условиями, более благоприятными как для раннего скотоводства, так и для развитого земледелия. Несмотря на это, исследователи-зоологи редко ставят задачу описания и преобразования фауны лесостепи вне контекста истории охоты (Цалкин, 1963; Бибикова, 1963, 1975, 1975а; Верещагин, Барышников, 1980; Явшева, 2006; Гасилин, Косинцев, 2009). Справедливо это в отношении и других зон (Верещагин, 1956; Динесман, 1982; Бибикова, 1984; Косинцев, 2002; Косинцев, Явшева, 2009; Косинцев, Бачура, 1999; Bachura et al., 2019). Независимо от зонального членения широко изучается на примерах различных регионов постледниковая история фауны копытных Северной Евразии (Верещагин, 1959; Паавер, 1965; Верещагин, Русаков, 1979; Девяшин, Косинцев, 2013; Пластеева, Клементьев, 2018; Бачура, Косинцев, 2020; Lorenzen et al., 2011) и изменения их ареалов (Громова, 1931, 1935; Бурчак-Абрамович, 1957; Бибикова, 1960, 1962; Верещагин, 1967; Бибикова, Белан, 1981; Benecke, 2005; Schmölcke, Zachos, 2005; Sommer еt al., 2008, 2009, 2011, 2014; Косинцев и др., 2013; Crees et al., 2016). Однако во всех случаях очень редко анализируются количественные изменения структуры населения копытных по обилию их костных остатков (Паавер, 1965; Верещагин, Русаков, 1979; Кузьмина, 1988; Явшева, 2006; Гасилин, Косинцев, 2009; Гасилин, 2013; Девяшин, Косинцев, 2013; Rosvold et al., 2012), по минимальному числу особей11 (Динесман, 1982) и по динамике площади ареалов (Crees et al., 2016). Редкость таких работ в сравнении с изысканиями, основанными на описании состава фаун и границ ареалов, объясняется трудоемкостью сбора необходимого объема материала и недостаточной методической базой исследования. Вместе с тем количественная составляющая голоценового разнообразия копытных природной зоны содержит сведения о хронологических и географических масштабах значимых для него изменений.
Цель нашей работы – анализ изменений состава и структуры населения копытных лесостепной зоны во второй половине голоцена.
ПРИРОДНЫЕ УСЛОВИЯ РАЙОНА ИССЛЕДОВАНИЯ
Лесостепь, охватывающая колки и крупные лесные массивы, луговые степи и остепненные луга, является особой ландшафтной зоной и среди других зон выделяется рядом характеристик (Мильков, 1950), в частности на небольшие колебания климата она способна реагировать перераспределением пространства между лесными и степными участками. Общие контуры, положение и широтная протяженность зоны в умеренном поясе определены климатическим градиентом (средние летние температуры, осадки и влажность воздуха). Он обусловливает подъем южной границы зоны на несколько градусов в верхние широты при движении от Приднестровья к Тоболо-Иртышскому междуречью и ее спуск к Приобью. Геоморфологические условия и геологическое строение коры определяют очертания границ между провинциями и долготную протяженность зоны (Мильков, 1950): расширение лесостепи в долготном направлении до 450 км в пределах Среднерусской возвышенности, сужение до 250 км в западносибирских равнинах и новое расширение вдоль возвышенностей Приобья. Примечательно, что изменения границ лесостепи – подъем и спуски, расширения и сужение – описывают дугу.
Лесостепь включает наиболее сложные экосистемы умеренного пояса Северного полушария и может быть определена как самостоятельный биом (Мордкович, 2006; Erdős et al., 2019). По причине сильнейшей агрикультурной трансформации южной полосы пояса выявляемые исследователями природные границы лесостепи спорны и иногда реконструируются с очень широким охватом территорий (Erdős et al., 2019). С произвольным объединением островов лесостепи правобережного Поднепровья в единое целое с левобережным массивом, небольшими изменениями в положении границы между Среднерусской и Приволжской провинциями, а также с пренебрежением наличия островов широколиственных лесов в Днепровской и Среднерусской провинциях и сосновых лесов в Обской провинции геоботаническая схема членения зоны нами принята по работам отечественных специалистов (Юго-Восток европейской …, 1971; Гвоздецкий, Михайлов, 1978; Мильков, Гвоздецкий, 1986) в ее восстановленных границах (Атлас. Карта растительности, 1981; Fekete et al., 2002; Иллеш, 2006).
Полоса лесостепи протянулась по равнинам Евразии с небольшими перерывами от 18° в.д. до 105° в.д. В ней выделяют ряд провинций, сменяющих одна другую в широтном направлении (вдоль параллели). Ниже указано (табл. 1), на какие орографические элементы приходятся провинции в принятых границах.
Таблица 1.
Орографическая приуроченность отрезков лесостепи, их площадь и объемы выборок
| Лесостепная провинция |
Орография | Площадь, тыс. км2 |
k | Абсолютное количество | |||||
|---|---|---|---|---|---|---|---|---|---|
| местонахождений | костных остатков | ||||||||
| а | б | в | а | б | в | ||||
| 1. Паннонская | Средне-Дунайская низменность, в границах Венгрии | 22.1 | 4.90 | 48 | 54 | 6 | 6625 | 563 | 184 |
| 2. Днепровская | Южно-Молдавская равнина, Подольская, Приднепровская возвышенности, равнины Приднепровской низменности | 42.1 | 2.54 | 49 | 56 | 2 | >6180 | 3715 | 24 |
| 3. Среднерусская | Среднерусская возвышенность, западная часть Окско-Донской равнины | 53.3 | 1.01 | 18 | 36 | – | >700 | >4120 | – |
| 4. Приволжская | Восточная часть Окско-Донской равнины, Приволжская возвышенность | 32.8 | 0.18 | 2 | 4 | – | 5 | 17 | – |
| 5. Заволжская | Высокое (Предуралье) и Низменное Заволжье | 48.3 | 2.86 | 81 | 57 | – | 2834 | 810 | – |
| 6. Красноуфимская и Месягутовская | Крайний юг Среднего Урала и Южный Урал | 5.6 | 1.25 | 7 | – | – | 703 | – | – |
| 7. Тобольская | Урало-Тобольское плато | 28.2 | 1.03 | 16 | 7 | 6 | 1048 | 268 | 1108 |
| 8. Ишимская | Ишимская равнина | 41.2 | 0.73 | 22 | 6 | 2 | 1079 | 1123 | 20 |
| 9. Барабинская | Барабинская низменность | 31.3 | 0.42 | 8 | 4 | 1 | 3128 | 121 | 1765 |
| 10. Обская и Кузнецкая | Приобское плато, Кузнецкая котловина, Бийско-Чумышская возвышенность и Салаирский кряж | 45.6 | 1.18 | 24 | 29 | 1 | >1460 | >1390 | 17 |
| 11. Ачинско-Красноярско-Минусинская | Приаргинская равнина и котловины Минусинской впадины | 31.3 | 0.35 | 9 | 2 | – | >580 | ? | – |
| 12. Канская | Канско-Рыбинская котловина | 10.9 | 0.28 | 2 | – | 1 | 4645 | – | 47 |
| 13. Балаганско-Иркутская | Иркутско-Черемховская равнина и Лено-Ангарское плато | 5.5 | 2.73 | 7 | 5 | 3 | >670 | >3100 | 513 |
Примечания. k – отношение числа местонахождений к площади участков, умноженное на 1000; а – средний голоцен; б – поздний голоцен; в – средний–поздний голоцен; > – действительная сумма костей больше указанной ввиду отсутствия в некоторых публикациях количества костей в местонахождениях;? – количество костных остатков неизвестно.
МАТЕРИАЛ
Работа основана на данных о находках костей копытных в древних поселениях, расположенных в восстановленных геоботаническими методами границах лесостепной зоны. Границы зоны восстановлены для ближайших столетий, предшествовавших ее повсеместному агрикультурному освоению.
Мы исходим из допущения, что выборки костей копытных из культурных слоев археологических памятников бытового типа (стоянки, селища, городища): 1) принадлежат животным, обитавшим в окрестностях поселения; 2) представляют собой пищевые отходы, в которые сходным образом попадали остатки туш всех добытых видов; 3) происходят от особей, добыча которых есть результат разумной экономии усилий по преследованию, результат обычного случайного обнаружения и выслеживания зверя в охотничьих угодьях, а не необходимого (например, предписанного особым обрядом) поиска и преследования особей конкретного вида, невзирая на доступность других копытных; 4) отражают совокупную добычу в течение годового цикла; 5) являются остатками жертв, способы и орудия лова которых не обеспечивали преимущества добычи одних видов копытных перед другими. Из сделанных допущений следует, что остатки охотничьей добычи отражают видовой состав, а соотношение их количеств, называемое в работе долей, отражает относительную численность животных в естественной среде с искажением тафономической природы, которое не препятствует обнаружению основных направлений изменения численности.
Материал, вероятно, включает остатки массовых жертв копытных, добытых на путях сезонных миграций: в некоторых местонахождениях, характеризующихся сравнительно большой выборкой находок, один вид копытного обнаруживает значительное превосходство в количестве костей. Местонахождения, где более сотни экземпляров костей одного вида превосходят половину выборки всех остатков копытных, составили порядка 10%, что при выбранных методах расчета не сказывается на результатах.
В результате отсутствия палеоботанических работ, обобщающих данные по смещениям на протяжении голоцена границ зон, смежных с исследуемой зоной, мы, ограничивая район исследования, исходили из предположения, что границы лесостепи менялись во времени с частотой и в масштабах, которыми здесь можно пренебречь. По некоторым данным (Динамика ландшафтных компонентов …, 2002), основная часть лесостепи к середине голоцена располагалась в принятых в работе границах, хотя, вероятно, была сужена в меридиональном направлении и занимала меньшую площадь.
Район исследования в соответствии с геоботаническими провинциями лесостепной зоны разбит для анализа на участки, площадь которых рассчитана средствами геоинформационной системы (QGIS, 2020). Отношение числа местонахождений к площади участков (показатель k табл. 1) отражает изученность каждого из участков района исследования: наиболее изучена Паннонская лесостепь (Венгрия), наименее – Приволжская (районы Тамбовской, Воронежской, Пензенской, Саратовской, Ульяновской, Самарской областей и Татарстана). Фаунистические материалы для Красноуфимской лесостепи неизвестны. Распределение местонахождений в пределах участков неравномерно (рис. 1) и, очевидно, связано с археологической изученностью территорий, обратно пропорциональной расстоянию от крупных городов с научными центрами.
Рис. 1.
Провинции и островные участки лесостепной зоны Евразии (с запада на восток нумерованы соответствующие им отрезки в переходных от светлого к темному тонах) и средне-позднеголоценовые местонахождения копытных (белые и черные значки). Номера лесостепных провинций соответствуют номерам в табл. 1. В тексте и подрисуночных подписях сложные названия лесостепных провинций заменены на простые – по наименованию одной из составляющих провинцию лесостепей. Контуры Паннонской лесостепи взяты по: Иллеш, 2006. Прочее с небольшими изменениями по: Атлас. Карта растительности, 1981.
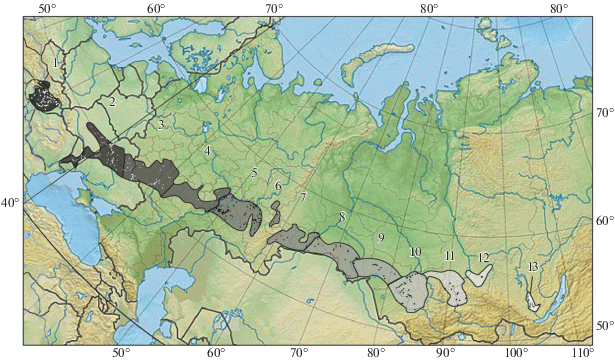
Использованы результаты палеофаунистических определений для местонахождений, расположенных в одной из провинций изучаемой природной зоны: Паннонская лесостепь (Bokonyi, 1974; Bartosiewicz, 1995, 1999, 2003; Choyke et al., 2004; The Early Neolithic …, 2007; Gál, 2009; Bartosiewicz, Gál, 2010; Horváth, 2012; Lyublyanovics, 2015), Днепровская лесостепь (Бибикова, 1953; Пiдоплiчко, 1956; Третьяков, 1959; Цалкин, 1966, 1967, 1970; Даниленко, 1969; Корнеев, 1969; Тимченко, 1970, 1972; Максимов, 1972; Белан, 1975, 1978, 1982; Збенович, 1980; Давид, 1982; Журавлев, 1989, 1990, 1998, 2000, 2000а, 2001, 2009; Журавльов, Котова, 1996, 1998; Журавльов и др., 1996; Цвек, 1997; Журавлев, Лысенко, 2000; Журавлев, Маркова, 2000; Котова, 2002; Колода, Фоменко, 2004; Шрамко и др., 2006; Бондаренко, 2008; Горбаненко и др., 2008; Пряхин, 2008; Сиволап, 2009; Колода, Горбаненко, 2012; Колода, Кройтор, 2012; Пуголовок и др., 2015; Тимків, 2015; Старобогатов, 2018; Бейлекчи, Чирков, 2019; Гак и др., 2019), Среднерусская лесостепь (Цалкин, 1963, 1966, 1969; Пряхин, 1976; Кузьмина, Каспаров, 1987; Чубур, Шинаков, 2006; Чубур, Новик, 2010; Антипина, 2011; Чубур, Радюш, 2013; Кабанов, 2014; Кройтор, Колода, 2014; Чубур, 2014, 2018, 2018а; Горчыца, Чубур, 2016; Чубур, Тропин, 2017; Яниш и др., 2018), Приволжская лесостепь (Петренко, 1984, 2003), Заволжская лесостепь (Цалкин, 1958; Петренко, 1984, 2003; Косинцев, 1988; Мосин, 1996; Гасилин, 2002, 2005, 2005a; Моргунова и др., 2017), Месягутовская лесостепь (Косинцев, Варов, 1996), Тобольская лесостепь (Потемкина, 1985; Косинцев, 1988; Косинцев, Кисагулов, 2018), Ишимская лесостепь (Цалкин, 1972; Косинцев, 1988), Барабинская лесостепь (Молодин, 1981, 1985; Молодин, Соболев, 1986; Косинцев, 1988; Молодин и др., 1990; Васильев и др., 1999; Молодин, Парцингер, 2009), Обская лесостепь (Троицкая, 1976; Шамшин, 1986; Сидоров, 1989, 1989а; Абдуганеев, 1996; Шамшин, Гальченко, 1997; Косинцев, 2002; Васильев, Новикова, 2005; Васильев и др., 2007, 2011; Ситников и др., 2007), Иркутская лесостепь (Ермолова, 1978; Пластеева, Клементьев, 2018).
Местонахождения сгруппированы в соответствии с климатической периодизацией голоцена (Хотинский и др., 1991), согласно которой рассмотрены составляющие средний голоцен атлантический (8000–4600 л. н.) и суббореальный (4600–2600 л. н.) периоды среднего голоцена и представляющий поздний голоцен субатлантический (2600–200 л. н.) период. Верхний предел для продолжающегося субатлантического периода принят условно, в качестве обозначения приблизительного времени окончания формирования изученных антропогенных отложений. Данные об остеологических материалах из местонахождений рубежа среднего и позднего голоцена, а также о недифференцированных материалах нескольких его периодов использованы только в анализе встречаемости видов в местонахождениях.
Морфологическая идентификация костных остатков нескольких видов имеет методические проблемы. Различение костных остатков зубра (Bison bonasus L. 1758), бизона (B. priscus Bojanus 1825), тура (Bos primigenius Bojanus 1825) и домашнего быка (Bos taurus L. 1758) проводится с большими затруднениями и не по всем костям скелета, а потому полученные для них результаты определений наименее точны. Дикая и домашняя формы лошади морфологически не различаются, и поэтому в настоящей работе дикая лошадь (Equus ferus Boddaert 1785) исключена из анализа.
В отношении проблемы перекрывания ареалов европейской (Capreolus capreolus L. 1758) и сибирской косули (C. pygargus Pallas 1771) в древности предполагается, что в Днепро-Донском междуречье проходила зона симпатрии (Цалкин, 1963): от Днепра на запад – европейская косуля, от Дона на восток – сибирская. Поскольку дифференциация остатков двух видов из проблемной области в обозреваемых работах не проведена, в нашем исследовании эти близкородственные виды-викариаты рассматриваются совместно.
В литературе есть указания на остатки дзерена (Procapra gutturosa Pallas 1777) в Алтайском крае в двух местонахождениях конца среднего голоцена (Кирюшин и др., 1988; Шамшин, Гальченко, 1997), но поскольку в публикациях не приведены надежные основания для идентификации нового для лесостепной зоны вида, в нашей работе сведения о находках дзерена не учитываются.
Данные по численности копытных территории РФ в 20–21 вв. взяты для административных областей, располагающихся преимущественно в лесостепной зоне (Данилкин, 1999, 2002; Состояние охотничьих ресурсов …, 2011): Белгородская, Воронежская, Липецкая, Тамбовская и Пензенская области пересечены Среднерусским отрезком зоны, Ульяновская, Самарская области, Башкортостан и Татарстан – Заволжским отрезком, Челябинская и Курганская – Притобольским, Курганская и Омская – Ишимским, Омская и Новосибирская – Барабинским, а Новосибирская пересекается Обским отрезком зоны. В таком же порядке сгруппированы и суммированы величины поголовья копытных, за исключением двух последних сочетаний, которые сгруппированы по парам и объединены – данные о поголовье Омской и Новосибирской областей противопоставлены данным по костным остаткам в местонахождениях Барабинского и Обского отрезков лесостепи. Аналогичные данные с территории Украины (Колиснык, 1990; Волох, 2016) имеются только для лесной (Волынская обл.) и степной (использованы данные по Днепровской, Донецкой, Луганской, Николаевской, Одесской областям севера зоны) частей страны. Совокупные данные для территории Венгрии (Csanyi, Lehoczki, 2009) нельзя подчинить какой-либо зоне, однако ввиду того что государство в основном располагается в изучаемой зоне, они использованы в анализе. Относительная численность рассчитана как доля вида в поголовье последних десятков лет, когда в результате предпринятых мер численность их возросла и стала стабилизироваться: для Венгрии с 1969 по 2005 гг., для лесной территории Украины с 1978 по 1986 гг., для степной – с 1975 по 2015 гг., для РФ с 1986 по 2010 гг.
МЕТОДЫ
Для измерения относительного обилия рассчитаны два показателя, выраженных в долях. Первый показатель, собственно доля вида, – отношение числа костей вида к сумме костей всех копытных в местонахождении. Второй показатель, встречаемость вида, – отношение числа местонахождений, где были найдены костные остатки вида, к общему числу местонахождений в отрезке зоны.
Первый показатель положен в основу анализа структуры населения копытных лесостепи в голоцене. В расчет показателя не включены местонахождения, выборки костей из которых принадлежали одному виду копытных или содержали менее 5 экземпляров. Оценки влияния факторов пространства, времени и их взаимодействия на соотношение костных остатков видов в местонахождениях рассчитаны на основе матрицы расстояний Брея-Кёртиса по алгоритму PERMANOVA (Anderson, 2001; Oksanen et al., 2019) – многомерного непараметрического аналога дисперсионного анализа с перестановочным тестом (999 пермутаций). Структура населения копытных проанализирована при помощи ординации методом главных координат PCoA (Legendre, Gallagher, 2001; Legendre, Legendre, 2012) на той же матрице.
Второй показатель использован для оценки встречаемости копытных в провинциях лесостепи в голоцене. В его расчет включены местонахождения, для которых имелось указание на наличие конкретного вида среди костных остатков копытных, т.е. все без исключения местонахождения из свода данных (табл. 1).
Доли видов среди фоновых копытных лесостепи в голоцене – относительное обилие видов, выраженное величиной первого показателя, – сравнены с величинами относительной численности поголовья копытных в пределах каждого участка и района исследования в целом.
Участки района исследования, содержащие менее 25 местонахождений, использованы не во всех анализах.
Расчеты и визуализация выполнены в среде программирования R 4.0.2 (R Core Team, 2020) при помощи пакетов ape (Paradis, Schliep, 2019) и vegan (Oksanen et al., 2019) для обработки многомерных данных (коэффициенты сходства, PCoA, PERMANOVA), а также пакета ggplot2 (Wickham, 2016) и коллекции пакетов tidyverse (Wickham et al., 2019) для визуализации и предварительной обработки данных. Все скрипты и исходные данные доступны на репозитории GitHub по адресу: https://github.com/ANSozontov/Gasilin_2020
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На высоком уровне значимости показано (табл. 2), что наибольшее значение коэффициента детерминации r 2 получено по фактору пространства “провинция лесостепи”, около половины дисперсии объяснено действием неучтенных факторов, наконец, менее 10% дисперсии определены временны́м фактором “период голоцена” и его взаимодействием с фактором “провинция лесостепи”.
Таблица 2.
Результаты анализа PERMANOVA
| Фактор | Df | F. Model | r 2 | p-value |
|---|---|---|---|---|
| Провинция лесостепи | 11 | 22.8 | 0.43 | 0.001 |
| Период голоцена | 3 | 4.4 | 0.02 | 0.001 |
| Взаимодействие факторов | 20 | 1.9 | 0.07 | 0.001 |
| Остатки | 284 | – | 0.48 | – |
| Всего | 318 | – | 1.00 | – |
Структура населения копытных лесостепи в голоцене рассмотрена в отношении к его периодам (рис. 2) – значительное перекрывание относящихся к ним трех групп местонахождений указывает на небольшую величину их различий.
Рис. 2.
Ординация местонахождений с группировкой по периодам голоцена. Алгоритм PCoA, расстояние Брея-Кёртиса. Распределение местонахождений: А – атлантического периода, Б – суббореального периода, В – субатлантического периода. Провинции лесостепи: 1 – Паннонская, 2 – Днепровская, 3 – Среднерусская, 4 – Заволжская, 5 – Тобольская, 6 – Ишимская, 7 – Барабинская, 8 – Обская, 9 – Иркутская.
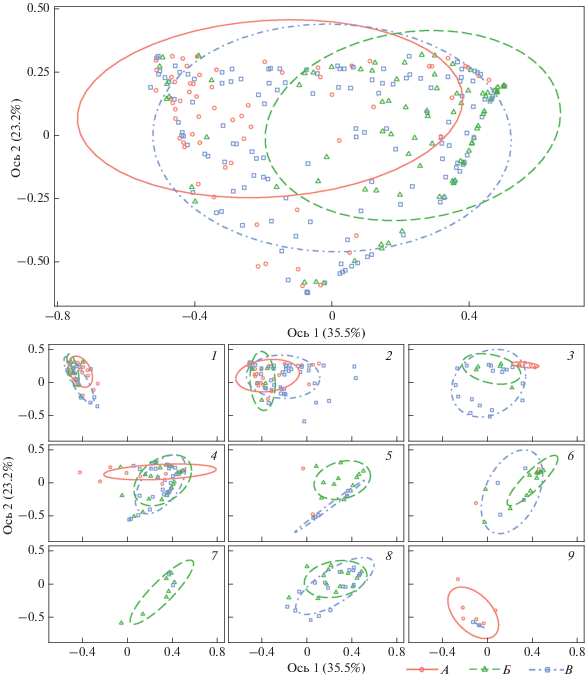
Справедливо это и для паттернов, выделенных для провинций лесостепи (рис. 2).
Структура населения, выраженная через отношение местонахождений к провинциям лесостепи, оказалась наиболее специфичной в Паннонской островной провинции: наблюдается обособленность группы ее местонахождений, перекрывающихся с группой Днепровской и, незначительно, с группой Среднерусской провинции (рис. 3).
Рис. 3.
Ординация местонахождений с группировкой по лесостепным провинциям. Алгоритм PCoA, расстояние Брея-Кёртиса. Периоды: 1 – атлантический, 2 – суббореальный, 3 – субатлантический. Провинции лесостепи: А – Паннонская, Б – Днепровская, В – Среднерусская, Г – Заволжская, Д – Тобольская, Е – Ишимская, Ж – Обская.
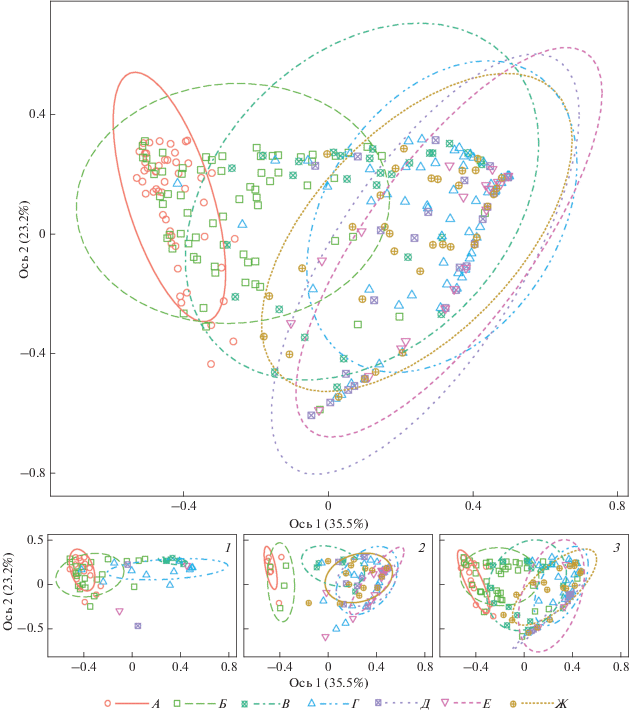
В свою очередь Днепровская группа в значительно большей мере перекрывается со Среднерусской и частично – с остальными группами. Заволжская группа перекрывает сибирские группы, выказывая с ними сходство. Из трех приведенных сибирских групп Обская перекрывается с западными группами более всего. Судя по частным паттернам, выделенным для отдельных периодов голоцена (рис. 3), характер различий между провинциями не был стабильным – в атлантическом и суббореальном периодах различия между Паннонской и Днепровской провинциями с одной стороны и прочими отрезками зоны с другой стороны наибольшие, а в субатлантическом периоде все различия наименьшие. В атлантическом периоде обе западные провинции более походили на Заволжскую, а с суббореального периода – на соседнюю Среднерусскую провинцию. Только местонахождения Паннонской лесостепи максимально обособлены во все изученные периоды.
Поскольку местонахождения из разных провинций в наибольшей мере дифференцированы по первой оси, мы проанализировали зависимость этой первой главной координаты от долготы. Географический фактор статистически значим (p < 0.001) и в линейной, и в полиномиальной модели второй степени, однако коэффициент детерминации r 2 (0.33 и 0.68 соответственно) и информационный критерий Акаике (66.4 и –174.4) дают основания считать полиномиальную модель предпочтительной (рис. 4).
Рис. 4.
Полиномиальная и линейная модели зависимости первой главной координаты (ось 1) местонахождений от долготы. Параметры моделей приведены в тексте. Местонахождения малых островных лесостепей включены в анализ, но в диаграмме не отражены. Провинции лесостепи: А – Паннонская, Б – Днепровская, В – Среднерусская, Г – Заволжская, Д – Тобольская, Е – Ишимская, Ж – Обская, З – Иркутская, И – полиномиальная модель, К – линейная модель.
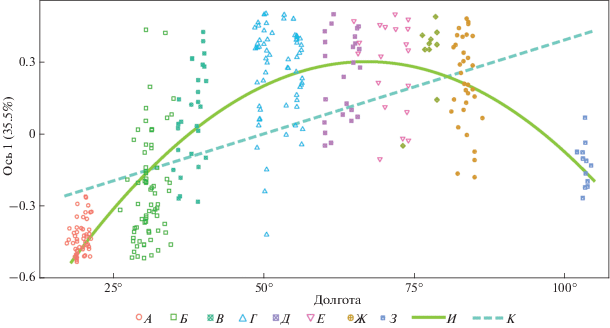
Диаграмма помогает зрительно оценить связь между местонахождениями, характеризующимися конкретными соотношениями количеств остатков разных видов и географическим положением. От Паннонской провинции до Среднерусской линия тренда поднимается, от Заволжской провинции до Барабинской – выполаживается, поскольку соответствующие местонахождения имеют близкие значения по первой оси главных координат, а с Обской провинции до островной Иркутской линия тренда понижается. Паннонская и Иркутская островные провинции далеко отстоят от сплошной полосы лесостепи, но их исключение из анализа не отменило параболическую модель: характер распределения относительного обилия копытных в провинциях лесостепной зоны обнаруживает симметрию.
Результаты демонстрируют, что различные доли костных остатков видов копытных распределены по местонахождениям голоцена не хаотично, но в значительной мере определены нахождением последних в провинции лесостепи. Упорядоченность данных оправдывает сделанные методические допущения. Главным же фактором изменений следует признать географический, в основном действующий в широтном направлении, вдоль зоны. С ним тесно связаны климат и рельеф. Они формируют зону и вызывают ее пространственные изменения, тоже имеющие дугообразный характер.
Динамика относительного обилия видов, выраженная первым показателем, отразила сложность организации структуры населения копытных в зоне. Близость по первой главной координате местонахождений Заволжской и западносибирских провинций (рис. 3, 4) связана с низкой долей кабана и благородного оленя, а также тура, и с отсутствием кулана (в Заволжской присутствовал).
Видовой состав и встречаемость копытных в провинциях лесостепи в голоцене. Фоновыми видами среди копытных животных евразийской лесостепной зоны в среднем и позднем голоцене следует признать европейскую и сибирскую косуль, лося, благородного оленя и кабана: от других копытных их отличают постоянное присутствие и обычно высокая встречаемость (второй показатель) среди местонахождений любой из провинций лесостепи, за исключением Паннонской, где и поныне отсутствует лось.
В наиболее насыщенной местонахождениями Паннонской провинции (табл. 1; рис. 1) оказался беднейший для большинства лесостепных отрезков состав видов (что объясняет ее положение на рис. 3), но только здесь для среднего голоцена отмечен европейский плейстоценовый осел (E. hydruntinus Regalia 1907) (табл. 3).
Таблица 3.
Встречаемость костных находок копытных в лесостепных провинциях, %
| Вид | 1. Паннонская | 2. Днепровская | 3. Среднерусская | 4. Приволжская | 5. Заволжская | 6. Месягутовская | 7. Тобольская | 8. Ишимская | 9. Барабинская | 10. Обская | 11. Минусинская | 12. Канская | 13. Иркутская |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Кабан | 47 | 69 | 74 | 0 | 25 | 50 | 34 | 20 | 38 | 33 | 17 | 0 | 13 |
| Кабарга | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 67 | 33 | 13 |
| Благородный олень | 87 | 83 | 45 | 0 | 6 | 0 | 14 | 10 | 15 | 31 | 33 | 100 | 80 |
| Косули | 51 | 67 | 55 | 33 | 48 | 50 | 76 | 57 | 77 | 61 | 25 | 100 | 100 |
| Гигантский олень | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| Лось | 0 | 50 | 77 | 83 | 80 | 100 | 90 | 90 | 100 | 83 | 25 | 100 | 67 |
| Северный олень | 0 | 0 | 2 | 0 | 9 | 50 | 17 | 20 | 8 | 9 | 0 | 0 | 13 |
| Тур | 44 | 21 | 4 | 0 | 6 | 0 | 3 | 7 | 8 | 2 | 0 | 0 | 0 |
| Зубр | 0 | 12 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Сайга | 0 | 2 | 4 | 0 | 16 | 0 | 10 | 23 | 23 | 7 | 8 | 0 | 0 |
| Европейский осел | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Кулан | 0 | 6 | 2 | 0 | 4 | 0 | 7 | 0 | 0 | 7 | 0 | 0 | 13 |
| Всего местонахождений | 108 | 107 | 53 | 6 | 138 | 2 | 29 | 30 | 13 | 54 | 12 | 3 | 15 |
Впрочем, восточнее вид устанавливали в синхронных местонахождениях в зоне широколиственных лесов Центрально-Молдавской возвышенности в долине р. Реут (Давид, 1982) и в Приазовских степях (Котова, 2002). Следует предполагать, что в Днепровской лесостепной провинции вид также обитал. В это же время именно в Паннонской провинции наиболее многочислен тур, который здесь мало уступал по встречаемости фоновым видам и фактически к таковым относился, в отличие от европейского осла. На первом месте по встречаемости на этом западном форпосте зоны и в среднем, и в позднем голоцене находился благородный олень.
Для голоцена Днепровской лесостепи, помимо фоновых видов и тура, отмечены зубр и такие представители открытых пространств, как кулан (E. hemionus Pallas 1775) и сайга (Saiga tatarica L. 1766) (табл. 3). Как и в Паннонской провинции, по встречаемости здесь преобладал благородный олень. Лось, как и в настоящее время, занимал среди фоновых видов последнее место по этому показателю. Тур встречался в два с лишним раза реже, чем на западе.
В Среднерусской провинции кроме копытных, указанных для Днепровской провинции, обитал северный олень (Rangifer tarandus L. 1758), который мог здесь появляться заходами, вероятно, в особо многоснежные зимы (табл. 3). Видовой состав этой провинции наиболее богат видами. Начиная со Среднерусской и далее на восток до Обской провинции, лось становится самым многочисленным видом среди копытных. В Среднерусской провинции он лишь немного обходит кабана, встречаемость которого нарастала от Паннонской провинции и достигла здесь максимума, впервые обнаружив значительный разрыв с величиной встречаемости косули. Встречаемость благородного оленя резко снизилась – среди фоновых видов с этого отрезка лесостепи на восток до Обского включительно он на последнем месте.
В Заволжской провинции в голоцене обитали те же виды, что и в Среднерусской, за исключением зубра. Видовая идентификация находок зубра в степной зоне (Петренко, 1984), на наш взгляд, нуждается в проверке. Доминирование лося здесь и в провинциях, лежащих восточнее, хорошо выражено. Косули и кабан в западных провинциях занимали по встречаемости второе или третье место, причем величины их встречаемости различались слабо (табл. 3). Начиная с Заволжской лесостепи и далее на восток, косуля занимает второе место после лося. Только на восточной окраине зоны, в Иркутской островной лесостепи, косуля поднимается на первое место.
Видовые составы копытных Заволжской, Тобольской и Обской лесостепей идентичны. Отсутствием кулана и наличием гигантского оленя (Megaloceros giganteus Blumenbach 1799) отличаются от них составы видов Ишимской и Барабинской провинций (в Барабинской лесостепи остатки гигантского оленя обнаружены только в составе погребального комплекса могильника). Рефугиумы гигантского оленя существовали вплоть до середины атлантического периода (Lister, Stuart, 2019).
В сибирских провинциях лесостепи в среднем-позднем голоцене сохраняется порядок следования фоновых видов по их встречаемости, установленный для Заволжской лесостепи: лось, косуля, кабан, благородный олень.
Видовые составы копытных для Месягутовской, Минусинской, Канской и Иркутской островных лесостепей характеризуются обусловленной методическими причинами неполнотой, но включают, как минимум, три фоновых вида, среди которых всегда присутствуют лось и косуля. Начиная с Минусинской и кончая Иркутской провинцией, в их состав видов входит горно-таежный азиатский вид – кабарга (Moschus moschiferus L. 1758). Учитывая небольшие площади островных лесостепей (табл. 1), где расположены местонахождения с остатками зверя, нельзя исключить, что добыт он в характерных для него природных условиях за пределами изучаемой зоны.
В результате анализа выявлено, что видовые составы отрезков лесостепей в среднем-позднем голоцене во многом идентичны, наиболее устойчивые различия между ними проявляются в присутствии европейского осла в Паннонской лесостепи в среднем голоцене и отсутствии там лося, а также в возможных заходах кабарги в лесостепь от Минусинского отрезка до Иркутского островного ее участка. Только в историческое время резко сократили ареалы и исчезли в лесостепи благородный олень, северный олень, лось, зубр, сайга и кулан, во многих местах исчезли косули (Данилкин, 1999, 2005). Географические различия в среднем-позднем голоцене проявляются, главным образом, в постепенных изменениях встречаемости на протяжении зоны тура и фоновых видов. Заметим, что состав фоновых видов сохранился до нашего времени, хотя абсолютная их численность на территории РФ еще находится в затянувшемся процессе восстановления, обнаруживая сильную зависимость от антропогенного фактора, а именно, от управления ресурсами и организации охотничьего хозяйства (Данилкин, 2018). Эти виды, будучи широко распространенными эвритопами, находят в лесостепи благоприятную среду обитания, т.к. предпочитают сплошным массивам разреженные леса с разнообразием стаций, которые используются ими в течение года: питательное разнотравье и возможность спасения от гнуса на открытой продуваемой местности лугостепей летом, укрытие от врагов и веточный корм в лесных колках зимой, а также возможность избежать многоснежных участков благодаря разнообразию ландшафтов этой зоны. Фоновые виды, а также домашний скот, являются пищевыми конкурентами (Данилкин, 1999).
Доли видов среди фоновых копытных лесостепи в голоцене. Оценено относительное обилие многочисленных видов в крупных лесостепных провинциях. Настоящий анализ позволяет, по сути, рассмотреть непосредственно вклад каждого из фоновых видов (по средним значениям первого показателя) в структуру населения, в частности в характер ее движения со сменой провинций. Но поскольку число местонахождений и количество в них костных остатков одного вида, а вслед за ними и оба изучаемых показателя обилия взаимосвязаны, можно принять, что вклады в географическое изменение относительного обилия в общем уже оценены выше по встречаемости фоновых видов, и остановиться на вкладе видов в фактор времени “период голоцена”. Каждая диаграмма динамики обилия вида по периодам голоцена завершается позицией вида в последние десятилетия, которая установлена по данным учета поголовья в охотничьих хозяйствах (рис. 5).
Рис. 5.
Доля фоновых видов копытных в крупных провинциях лесостепной зоны в среднем и позднем голоцене и их относительная численность в последние десятилетия (по: Данилкин, 1999, 2002; Csanyi, Lehoczki, 2009; Состояние охотничьих ресурсов …, 2011; величинам долей костных остатков видов в Днепропетровской лесостепи противопоставлены величины относительной численности поголовья в лесной (Колиснык, 1986) и степной (Волох, 2016) частях Украины, разница между которыми оказалась недостоверной). Периоды: А – атлантический, Б – суббореальный, В – субатлантический, Г – 20–21 вв. * – единичная находка вида. Рамка – границы межквартильного размаха (25–75%), отрезок внутри рамки – медиана, отрезки снаружи рамки – минимальное и максимальное значения, точки – выбросы.
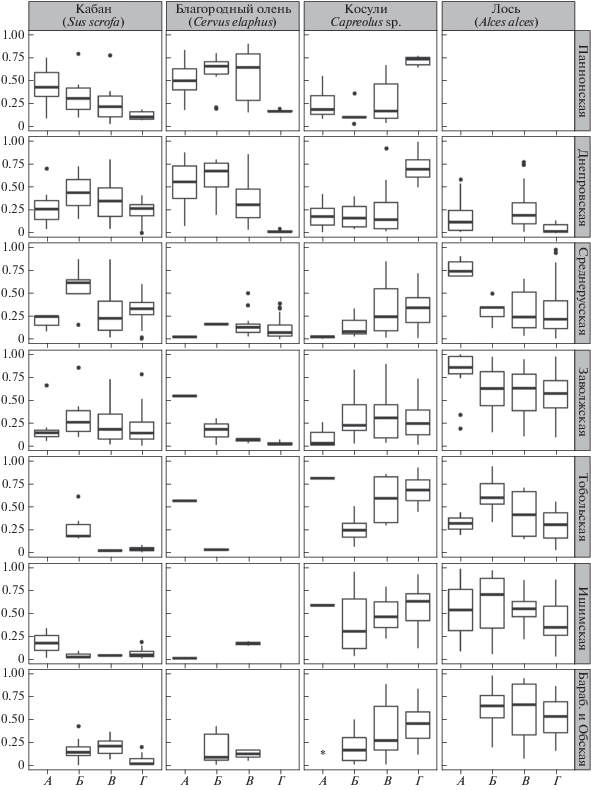
В Паннонской провинции лесостепи с атлантического периода снижалась средняя доля кабана, и в 20–21 вв. это снижение продолжилось (рис. 5). Доля благородного оленя поднялась к наступлению суббореального периода. В 20–21 вв. величина относительной численности поголовья оленя значимо ниже его доли в голоцене, что обусловлено высокой относительной (и абсолютной) численностью поголовья европейской косули, величина которой, наоборот, значимо превосходит среднюю долю косули в голоцене.
В Днепровской лесостепи доля кабана (в атлантическом периоде меньшая, чем в Паннонской лесостепи) немного поднималась в суббореальном периоде, но в целом оставалась стабильной, поскольку разница между долями невелика. Относительная численность его в рецентных лесной и степной зонах Украины близка по величине установленным для голоцена долям в лесостепи. Доля благородного оленя значимо снизилась от среднего голоцена к позднему и, судя по данным для соседних зон, продолжила значительное снижение в 20–21 вв. Стабильная в лесостепной зоне доля видов косуль, исходя из данных по степной зоне Украины, резко поднялась в 20–21 вв., подняв косуль на первое место по численности. Малочисленный в голоцене лось стал еще более редок в наши дни. Примечательно, что вид отсутствует в суббореальном периоде (изучены два местонахождения среднего суббореального и восемь – позднего суббореального периода).
В Среднерусской, как и в Днепровской провинции, наибольшая доля кабана в суббореальном периоде. Динамика доли вида в Среднерусской провинции в целом схожа с динамикой в предыдущем отрезке зоны. Доля благородного оленя минимальна среди видов. Доля косуль нарастала и продолжила тренд в 20–21 вв. Доля относительно многочисленного в атлантическом периоде лося (в отдельных местонахождениях уступает кабану) в дальнейшем снижается. Доля вида в 20–21 вв. близка его доле в субатлантическом периоде.
В Заволжской провинции доля кабана в суббореальном периоде максимальна, как и в двух предыдущих отрезках зоны, но в дальнейшем снижается, хотя различия ее значений от периода к периоду малы. Доля благородного оленя в атлантическом периоде здесь выше, чем в Среднерусской провинции. Она снижалась до 20–21 вв. Доля косули сибирской, минимальная в атлантическом периоде, значимо поднялась в суббореальном и более достоверно не изменялась. В суббореальном и субатлантическом периоде доля вида здесь выше, чем в Среднерусской провинции. Доля лося в Заволжской провинции, наибольшая среди видов, принимала максимальные значения, как и в Среднерусской провинции, в атлантическом периоде, затем она существенно снизилась. Но не ниже соответствующих значений, полученных для Среднерусской провинции. Доля вида в 20–21 вв. близка долям в двух предыдущих периодах.
В Тобольской провинции кабан не отмечен в атлантическом периоде, представленном лишь двумя местонахождениями. Доля кабана, вновь наибольшая в суббореальном периоде, позже снижается. Резкое снижение доли благородного оленя можно оценить только для среднего голоцена. Вид исчезает с позднего голоцена (отсутствует во всех пяти местонахождениях раннего и двух местонахождениях среднего-позднего субатлантического периода). В отличие от Заволжской в Тобольской провинции доля косули, наибольшая среди копытных в атлантическом периоде, резко снизилась в суббореальном и заметно поднялась в субатлантическом периоде, не выказывая разницы с нынешним состоянием относительной численности поголовья. Доля лося в противоположность доле косули резко поднялась в суббореальном периоде, а затем обнаружила некоторое снижение: в 20–21 вв. она существенно ниже доли в суббореальном периоде, но немного ниже, чем в субатлантическом периоде.
В Ишимской провинции, как на западе в Тобольской и на востоке в Барабинской и Обской провинциях, доли кабана и благородного оленя существенно уступают долям косули и лося, самым многочисленным видам в сибирских отрезках зоны всех периодов. Благородный олень здесь в голоцене крайне редок (и отсутствует в двадцати местонахождениях всего суббореального периода), а в 20–21 вв. он здесь и восточнее, в Обской лесостепи, не обитает. Доля косули с суббореального периода к 20–21 вв. нарастает, но различия ее значений между периодами невелики. Доля лося снова меняется в противофазе изменениям доли косули, и с суббореального периода снижается, но поскольку различия долей лося между периодами тоже невелики, можно считать, что относительное обилие обоих видов оставалось стабильным.
В Барабинской и Обской провинциях доля кабана в 20–21 вв. представляется, вопреки намеченному в голоцене тренду, сниженной (рис. 5). В единственном местонахождении атлантического периода вид отсутствует. Благородный олень – редкий, как и кабан, – также обитал здесь в суббореальном и субатлантическом периоде, но в 20–21 вв. его поголовье не восстановлено. Доля сибирской косули и лося оказались в согласии с трендами, обнаруженными для сибирских провинций зоны, расположенных западнее.
Лось в большинстве лесостепных провинций – наиболее многочисленный вид как в среднем, так и в позднем голоцене, но его доля проявляет слабую тенденцию к уменьшению во времени (рис. 6).
Рис. 6.
Общие тенденции изменения в лесостепной зоне долей костных остатков фоновых видов копытных в среднем и позднем голоцене и относительная численность копытных в последние десятилетия. Периоды: А – атлантический, Б – суббореальный, В – субатлантический, Г – 20–21 вв.
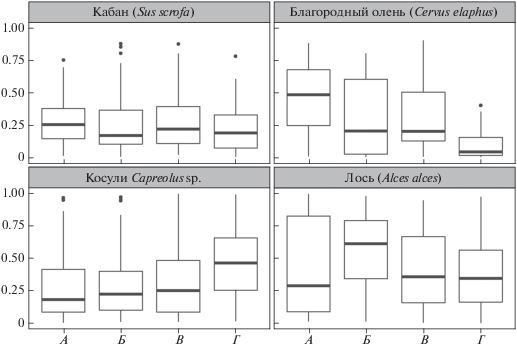
Доля косуль велика среди фоновых видов в голоцене, кроме того, она имела слабовыраженную тенденцию к нарастанию, в то время как доля благородного оленя – к снижению. Оба тренда нашли продолжение в последние десятилетия (рис. 6), причем доля благородного оленя значительно ниже, чем в позднем голоцене. Можно заключить, что благородный олень медленнее восстанавливает свою численность после истребления, которому был подвергнут в последние два–три столетия, и более других фоновых видов копытных нуждается в настоящее время в охране. Относительно сдержанное обилие кабана характеризуется стабильностью во времени (рис. 6).
Относительное обилие последних двух видов снижено в Заволжской и в сибирских провинциях лесостепи (рис. 5). Одной из причин малочисленности в лиственных массивах западносибирских лесостепей приверженного к лучшим в кормовом и защитном отношениях угодьям кабана (Данилкин, 2002), является недостаток материала для устройства зимних лежек (Баранов, 1990). Ареал благородного оленя под антропогенным прессом отступил к началу 20 в. с этих территорий, и до сих пор не восстановлен в западносибирских провинциях. Вместе с тем нами показано, что и прежнее относительное обилие вида как в западносибирских провинциях, так и в Заволжской лесостепи невелико. Можно предположить, что избегающий заболоченных территорий марал находил недостаточно пригодных биотопов в западносибирской лесостепи с ее плоскоравнинным рельефом, обусловливающим слабую дренированность почв (Гвоздецкий, Михайлов, 1978), в отличие от пищевого конкурента – лося (Данилкин, 1999), который предпочитает озерные долины и заболоченные низины с их водно-болотной растительностью. Восточнее р. Оби благородный олень несколько более обилен, что может объясняться соседством гор, где, как во многих других горных районах умеренного пояса, вид и в настоящее время находит благоприятное сочетание факторов (Данилкин, 1999). В частности, летом в горах зверь выбирает остепненные склоны и луга, иными словами, те же лесостепные биомы.
ЗАКЛЮЧЕНИЕ
Изучение массива количественных данных по костным остаткам копытных из древних поселений обнаружило его организованность, объясняемую сложной структурой населения видов, которые обитали в разных провинциях лесостепной зоны в среднем-позднем голоцене.
При высоком сходстве видовых составов лесостепных провинций выявленные в них паттерны относительной численности копытных определяются, главным образом, географическим фактором широтного направления. Так, в Европе соседние провинции проявляют меньшее сходство, тогда как Заволжская провинция и провинции Сибири к западу от р. Обь схожи в большей степени и представляются единым кластером. Среднерусская лесостепь занимает промежуточное положение между европейскими и сибирскими звеньями, обнаруживая большее сходство с последними. Географические различия провинций обусловлены распространенностью и относительной численностью фоновых видов. Ключевыми являются различия между западными провинциями (от Паннонской до Среднерусской) и восточными (от Заволжской до Барабинской): в последних кабан и благородный олень встречаются значительно реже. К востоку – в Обской в Иркутской лесостепи – благородный олень немного более многочислен.
Изменения во времени сказались, в первую очередь, на составе копытных изученной зоны. К концу среднего голоцена вымерли европейский плейстоценовый осел, гигантский олень и тур. Наибольшее обеднение видового состава копытных лесостепи произошло в историческое время. Обнаруженные для голоцена тенденции изменения относительного обилия фоновых видов во многих провинциях зоны оказались продолженными на рубеже истекшего и нынешнего столетий. Это, вероятно, свидетельствует о направлении и постоянстве ведущих природных факторов, которые определяют среду обитания и динамику относительной численности видов.
Список литературы
Абдуганеев М.Т., 1996. О хозяйстве населения лесостепного и предгорного Алтая в скифское время // Археология, антропология и этнография Сибири. Сб., посвященный памяти антрополога А.Р. Кима. Барнаул: Изд-во АГУ. С. 145–153.
Антипина Е.Е., 2011. Археозоологические коллекции из поселений поздней бронзы на территории Тульской области. Приложение 3 // Гак Е.И. Поселения эпохи бронзы на северной окраине донской лесостепи. М.: ГИМ. С. 166–201.
Атлас. Карта растительности, 1981. Географический атлас. Отв. ред. Колосова Л.Н. М.: ГУГК. С. 143.
Баранов П.В., 1990. Акклиматизация кабана в Западной Сибири // V съезд Всесоюз. Териол. об-ва АН СССР. Т. 3. М. С. 75–76.
Бачура О.П., Косинцев П.А., 2020. Фауна крупных млекопитающих Центральной части Восточно-Европейской равнины в голоцене // Зоологический журнал. Т. 99. № 5. С. 569–585.
Бейлекчи В.С., Чирков А.Ю., 2019. Раскопки гумельницкого поселения у пгт. Тараклия в Молдове в 1984–1985 гг. // Вестник Московского государственного областного университета. Сер. История и политические науки. М.: Изд-во МГОУ. № 5. С. 8–26.
Белан Н.Г., 1975. Фауна зарубинецких поселений Каневщины // Природная обстановка и фауны прошлого. Киев: Наукова Думка. Вып. 9. С. 108–128.
Белан Н.Г., 1977. До iсторii мисливства i твариництва у племен Правобережної України у I тисячолiттi н.е. // Археологiя. № 24. С. 29–37.
Белан Н.Г., 1978. Фауна городища Монастырек на Среднем Днепре // Использование методов естественных наук в археологии. Сб. науч. трудов. Киев: Наукова Думка. С. 96–109.
Белан Н.Г., Бибикова В.И., 1980. Лошадь мезолита Северо-западного Причерноморья // Археологические исследования на Украине в 1978-1979 гг.: тез. докл. XVIII конф. ИА АН УССР (Днепропетровск, апрель 1980 г.). Днепропетровск. С. 37.
Бєлан Н.Г., 1982. Фауна Трахтемирівського городища // Археологія. № 38. С. 50–63.
Бибикова В.И., 1953. Фауна раннетрипольского поселения Лука-Врублевецкая // Материалы и исследования по археологии СССР. М.-Л.: Изд-во АН СССР. Т. 38. 460 с.
Бибикова В.И., 1960. О распространении дикого кабана в четвертичном периоде // Бюллетень Комиссии по изучению четвертичного периода. № 25. С. 107–112.
Бибикова В.И., 1962. Остатки субфоссильного тарпана на Украине // Основные проблемы териологии: Эволюция, зоогеография, экология и морфология млекопитающих и происхождение домашних животных. № 6. С. 97–114.
Бибикова В.И., 1963. Из истории голоценовой фауны позвоночных в Восточной Европе // Природная обстановка и фауны прошлого. Киев: Изд-во АН УССР. № 1. С. 119–146.
Бибикова В.И., 1975. О смене некоторых компонентов фауны копытных на Украине в голоцене // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 80. № 6. С. 67–72.
Бибикова В.И., 1975а. Фауна копытных из мезолитических поселений Северо-Западного Причерноморья // 150 лет Одесскому археолог. музею АН УССР: тез. докл. юбил. конф. Киев: Наукова думка. С. 24–25.
Бибикова В.И., 1984. Копытные Северного Причерноморья в раннем голоцене (по материалам археологических раскопок) // Проблемы изучения истории современных биогеоценозов. М.: ИЭМЭЖ. С. 171–203.
Бибикова В.И., Белан Н.Г., 1983. К истории охотничьего промысла на Украине // Вестник зоологии. № 6. С. 14–20.
Бибикова В.И., Белан Н.Г., 1981. Раннеголоценовый тарпан на территории Северо-Западного Причерноморья // Вестник зоологии. № 3. С. 21–26.
Бондаренко В.Л., 2008. Результаты археозоологического изучения костного материала из раскопок поселений II–I тыс. до н.э. бассейна Северского Донца // Проблемы археологии Восточной Европы. К 85-летию Б.А. Шрамко. Харьков. С. 39–41.
Бурчак-Абрамович Н.И., 1957. Ископаемые быки Старого Света // Труды Естественно-исторического музея имени Г. Зарадаби. Баку: Изд-во АН АзССР. Т. 1. Вып. 11. 263 с.
Васильев С.К., Кирюшин К.Ю., Ситников С.М., Семибратов В.П., 2011. Фаунистические остатки из поселения Новоильинка-3 (по материалам раскопок 2010 года) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 17. Новосибирск: Изд-во ИАЭТ СО РАН. С. 148–151.
Васильев С.К., Мартынович Н.В., Чемякина М.А., 1999. Охота по данным остеологического анализа материалов памятника Омь 1 // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Материалы VII Годовой итоговой сессии Института археологии и этнографии СО РАН. Декабрь 1999 г. Новосибирск: Изд-во ИАЭТ СО РАН. С. 119–120.
Васильев С.К., Новикова А.В., 2005. Фаунистические остатки с памятников Ояшкинского археологического микрорайона // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 11. Новосибирск: Изд-во ИАЭТ СО РАН. С. 265–269.
Васильев С.К., Орлова Л.А., Кузьмин Я.В., 2007. Местонахождение фауны крупных млекопитающих сартанского времени на реке Орда (Ордынский р-н Новосибирской области) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 13. С. 29–32.
Верещагин Н.К., 1967. Геологическая история лося и его освоение первобытным человеком // Биология и промысел лося. М.: Россельхозиздат. Сборник 3. С. 3–37.
Верещагин Н.К., 1959. Млекопитающие Кавказа: история формирования фауны. М.-Л.: Изд-во АН СССР. 703 с.
Верещагин Н.К., 1956. О прежнем распространении некоторых копытных в районе смыкания европейско-казахстанских и центральноазиатских степей // Зоологический журнал. Т. 35. № 10. С. 1541–1553.
Верещагин Н.К., Барышников Г.Ф., 1980. Ареалы копытных фауны СССР в антропогене // Труды ЗИН АН СССР. Т. 93. С. 3–20.
Верещагин Н.К., Русаков О.С., 1979. Копытные Северо-Запада СССР (история, образ жизни и хозяйственное использование). Л.: Наука. 308 с.
Волох А.М., 2016. Охотничьи звери степной Украины. Монография: в 2-х кн. Херсон: Гринь Д. С. Кн. 2. 572 с.
Гак Е.И., Антипина Е.Е., Лебедева Е.Ю., Кайзер Э., 2019. Хозяйственная модель поселения среднедонской катакомбной культуры Рыкань-3 // Российская археология. № 2. С. 19–34.
Гасилин В.В., 2002. Новое археозоологическое местонахождение – именьковский памятник селище Ош-Пандо-Нерь. Бюллетень Самарская Лука. Самара. № 12. С. 125–135.
Гасилин В.В., 2005. Фауна предыменьковских и раннеименьковских памятников лесостепного Поволжья // Сташенков Д.А. Оседлое население Самарского лесостепного Поволжья в I–V вв. н.э. М. С. 56–62.
Гасилин В.В., 2005. Фауна Старомайнского городища именьковской культуры // Краеведческие записки. Самара. Вып. 12. С. 59–66.
Гасилин В.В., 2013. Крупные млекопитающие Приморья в голоцене // Зоологический журнал. Т. 92. № 9. С. 1055–1063.
Гасилин В.В., Косинцев П.А., 2009. Динамика структуры фауны копытных Урало-Поволжья в голоцене // Вестник Оренбургского университета. № 6. С. 108–110.
Гвоздецкий Н.А., Михайлов Н.И., 1978. Физическая география СССР. Азиатская часть. М.: Мысль. С. 448.
Горбаненко С.А., Журавльов О.П., Пашкевич Г.О., 2008. Сільське господарство жителів Пастирського городища. Київ: Академперіодика. 188 с.
Горчыца К., Чубур А., 2016. Рекогносцировочные раскопки средневекового Конина (поселение Старое Място 5, Конинский повят Великопольского воеводства) // Город Средневековья и раннего Нового времени: археология, история. Материалы V Всерос. семинара. Ноябрь 2013 г. Тула. С. 162–171.
Громова В.И., 1930. Тур и древнейшая история домашнего быка в СССР // Природа. № 7–8. С. 755–770.
Громова В.И., 1931. Первобытный бык или тур (Bos primigenius Воj.) в СССР // Ежегодник Зоологического музея АН СССР. Т. 32. С. 293–364.
Громова В.И., 1935. Первобытный зубр (Bison priscus Bojanus) в CCCР // Труды ЗИН АН СССР. Т. 2. № 2–3. С. 77–204.
Давид А.И., 1982. Формирование териофауны Молдавии в антропогене. Кишинев: Штиинца. 151 с.
Даниленко В.М., 1969. Неолит Украины. Киев: Наукова думка. 173 с.
Данилкин А.А., 1999. Млекопитающие России и сопредельных регионов. Оленьи (Cervidae) М.: ГЕОС. 552 с.
Данилкин А.А., 2002. Млекопитающие России и сопредельных регионов. Свиные (Suidae). М.: ГЕОС. 309 с.
Данилкин А.А., 2018. Дикие копытные России: тренд динамики населения // Поволжский экологический журнал. № 3. С. 357–367.
Девяшин М.М., Косинцев П.А., 2013. Копытные млекопитающие среднего и позднего голоцена юго-востока Западной Сибири // Вестник Томского государственного университета. Сер. Биология. Вып. 2. № 2. С. 127–140.
Динамика ландшафтных компонентов и внутренних морских бассейнов Северной Евразии за последние 130 000 лет, 2002. Под ред. А.А. Величко. М.: ГЕОС. Вып. 2. 232 с.
Динесман Л.Г., 1982. Изменение численности копытных в степях европейской части СССР в голоцене // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 87. № 2. С. 3–14.
Ермолова Н.М., 1978. Териофауна долины Ангары в позднем антропогене. Новосибирск: Наука. 222 с.
Журавлев О.П., 1989. Животноводство и охота в украинской лесостепи // Комплексные методы исследования археологических источников. М.: Изд-во МГУ. С. 11.
Журавлев О.П., 1990. О животноводстве и охоте племен Трипольской культуры Среднего Поднепровья и Побужья // Раннеземледельческие поселения-гиганты Трипольской культуры на Украине: тез. докл. Тальянки-Киев. С. 134–138.
Журавлев О.П., 1998. Животноводство у славянского населения восточноевропейской лесостепи во второй половине I тыс. нашей эры // Вопросы истории славян. № 12. С. 34–43.
Журавлев О.П., 2000. Животноводство и охота на трипольских поселениях Черкасской области // Археометрія та охорона історико-культурної спадщини. № 4. С. 62–65.
Журавльов О.П., 2000а. Остеологічні матеріали з поселення Діброва // АДУ 1994-1996. Київ: Прайм. С. 43–45.
Журавлев О.П., 2001. Остеологические материалы из памятников эпохи бронзы лесостепной зоны Днепро-Донского междуречья Киев: ИА НАНУ. 200 с.
Журавлев О.П., 2009. Животноводство и охота на раннеславянском поселении Семенки // Stratum plus. № 5. С. 632–639.
Журавлев О.П., Лысенко С.Д., 2000. Археозоологические исследования поселения Малополовецкое-3 эпохи поздней бронзы // Археология и древняя архитектура Левобережной Украины и смежных территорий. Донецьк: Східний видавничий дім. С. 81–82.
Журавлев О.П., Маркова Е.В., 2000а. Остеологические материалы из поселения Молюхов бугор // Археологічні дослідження в Україні в 1994-1996 роках. Киев: Прайм. С. 40–43.
Журавльов О.П., Котова Н.В., 1998. Свійські та дикі ссавці з давньоруського Пастирського городища // Археологія. № 3. С. 31–40.
Журавльов О.П., Котова Н.С., 1996. Тваринництво неолітичного населення України // Археологія. № 2. С. 3–17.
Журавльов О.П., Терпиловський Р.В., Шекун О.В., 1996. Остеологічні матеріали з поселення Олександрівка-1 // Олександрівка-1 – багатошарове ранньослов’янське поселення біля Чернігова. Чернігів: Сiверяньска думка. С. 103–125.
Збенович В.Г., 1980. Поселение Бернашевка на Днестре: (к происхождению трипольской культуры). Киев: Наукова Думка. 179 с.
Иллеш Е., 2006. Характерные признаки растительности лесостепи Венгрии и ее исследование // Материалы IV Международного симпозиума “Степи Северной Евразии”. Оренбург: ИПК “Газпромпечать”. С. 300–302.
Кабанов Д.О., 2014. Животноводство и охота населения Курского Посемья в эпоху раннего железного века // Исторические чтения – материалы научной конференции с междун. уч. Курск: Изд-во Курск. гос. ун-та. С. 86–91.
Кирюшин Ю.Ф., Гальченко А.В., Удодов В.С., Шамшин А.Б., 1988. Хозяйственно-культурные типы поздней бронзы лесостепного Алтая // Хронология и культурная принадлежность памятников каменного и бронзового веков Южной Сибири. Тез. докл. и сообщ. науч. конф. (23–25 марта 1988 г.). Барнаул: Изд-во ИИФиФ; АГУ. С. 138–142.
Колиснык Б.И., 1990. Динамика численности и промысел копытных в угодьях Волынского Полесья // Вестник зоологии. Т. 24. № 3. С. 65–67.
Колода В.В., Горбаненко С.А., 2012. Сельское хозяйство населения славяно-хазарской контактной зоны (на примере городища Водяное) // Дивногорский сборник. Воронеж: Научная книга. Вып. 3. С. 163–176.
Колода В.В., Кройтор Р.В., 2012. Остеология как отражение поэтапного развития скотоводства славяно-русского поселения Городное // Славяне Восточной Европы накануне образования Древнерусского государства. Материалы междун. конф., посвященной 110-летию со дня рожд. И.И. Ляпушкина (1902–1968) 3-5 дек. 2012 г. С.-Пб.: СОЛО. С. 139.
Колода В.В., Фоменко Г.Н., 2004. Остеологические материалы из славянских жилищ на городище Мохнач // Проблеми iсторii та археологii Украiни / Матерiали V МНК, присвяченої 350 річчю ім. Харкова і 200 річчю ХНУ ім. В.Н. Каразіна. Харків. С. 82–84.
Корнеев А.П., 1969. Домашние и дикие животные из раскопок древнего города Родни на Княжьей Горе // Вестник зоологии. Киев: Наукова думка. № 1. С. 32–38 с.
Косинцев П.А., 1988. Голоценовые остатки крупных млекопитающих Западной Сибири // Современное состояние и история животного мира Западносибирской низменности. Свердловск: Изд-во УрО АН СССР. С. 32–51.
Косинцев П.А., 2002. Типология териокомплексов лесостепного Приобья из поселений эпохи бронзы // Фауна Урала в плейстоцене и голоцене. Сб. науч. трудов. Екатеринбург: Университет. С. 162–166.
Косинцев П.А., Бачура О.П., 1999. Крупные млекопитающие волго-уральских степей в голоцене // Экология древних и современных обществ. Тез. докл. конф., 18–19 нояб. 1999. Тюмень. С. 91–94.
Косинцев П.А., Варов А.И., 1996. Ранние этапы животноводства в Волго-Уральском регионе. Взаимодействие человека и природы на границе Европы и Азии. Самара. С. 29–31.
Косинцев П.А., Кисагулов А.В., 2018. Костные остатки тура и крупного рогатого скота из голоценовых местонахождений южного Зауралья // Вестник Оренбургского государственного педагогического университета. Вып. 25. № 1. С. 44–55.
Косинцев П.А., Пластеева Н.А., Васильев С.К., 2013. Дикие лошади (Equus (Equus) s. l.) Западной Сибири в голоцене // Зоологический журнал. Т. 92. № 9. С. 1107–1116.
Косинцев П.А., Явшева Д.А., 2009. Промысловые млекопитающие степной зоны Урала и Западной Сибири в голоцене // Вестник Оренбургского государственного педагогического университета. № 6. С. 164–167.
Котова Н.С., 2002. Неолитизация Украины. Луганск: Шлях. 268 с.
Кройтор Р.В., Колода В.В., 2014. Животноводство поздних северян на славяно-хазарском пограничье. Ч. 1 // Древности. Харьковск. историко-археологический ежегодник. Вып. 13. С. 212–227.
Кузьмина И.Е., 1988. Млекопитающие Северного Прикаспия в голоцене // Археологические культуры Северного Прикаспия: сб. науч. трудов. Куйбышев. С. 173–188.
Кузьмина И.Е., Каспаров А.К., 1987. Остатки животных их неолитических стоянок Копанище и Черкасская в Воронежской области // Труды ЗИН АН СССР. Т. 168. С. 87–99.
Максимов Е.В., 1972. Среднее Поднепровье на рубеже нашей эры. Киев: Наукова думка. С. 184.
Мильков Ф.Н., 1950. Лесостепь Русской равнины. М.: АН СССР. 296 с.
Мильков Ф.Н., Гвоздецкий Н.А., 1986. Физическая география СССР. Общий обзор. Европейская часть СССР. Кавказ. М.: Высшая школа. 376 с.
Молодин В.И., 1981. Памятники одинцовского типа в Барабинской лесостепи // Проблемы западносибирской археологии. Эпоха камня и бронзы. Новосибирск. С. 63–75.
Молодин В.И., 1985. Бараба в эпоху бронзы. Новосибирск: Наука. 200 с.
Молодин В.И., Парцингер Г., 2009. Чича – городище переходного от бронзы к железу времени в Барабинской лесостепи. Т. 3. Новосибирск-Берлин: Изд-во Института археологии и этнографии СО РАН. 246 с.
Молодин В.И., Соболев В.И., 1986. Скотоводство населения Барабинской лесостепи в 14–16 вв. // Палеоэкономика Сибири. Новосибирск: Наука. С. 120–124.
Молодин В.И., Соболев В.И., Соловьёв А.И., 1990. Бараба в эпоху позднего средневековья. Новосибирск: Наука. 262 с.
Моргунова Н.Л., Васильева И.Н., Кулькова М.А., Рослякова Н.В., Салугина Н.П. и др., 2017. Турганикское поселение в Оренбургской области. Оренбург: Издательский центр ОГАУ. С. 221–232.
Мордкович В.Г., 2006. О статусе лесостепи в системе биомов // Материалы IV Междунар. симп. “Степи Северной Евразии”. Оренбург: ИПК “Газпромпечать”. С. 490–492.
Мосин В.С., 1996. Стоянка Бурли II и некоторые вопросы энеолита Южного Зауралья // Новое в археологии Южного Урала. Челябинск: Рифей. 265 с.
Пiдоплiчко I.Г., 1956. Матерiали до вивчення минулих фаун УРСР. Вип. 2. Киiв. 189 с.
Паавер К.Л., 1965. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту. 449 с.
Петренко А.Г., 1984. Древнее и средневековое животноводство Среднего Поволжья и Предуралья. М.: Наука. 176 с.
Петренко А.Г., 2003. Исследования остеологических материалов из древнейших археологических памятников Среднего Поволжья и Предуралья методами естественных наук, анализ проблем становления животноводческих основ в крае // Археология и естественные науки Татарстана. Казань. Кн. 1. С. 5–63.
Пластеева Н.А., Клементьев А.М., 2018. Крупные млекопитающие Прибайкалья в голоцене // Зоологический журнал Т. 97. № 5. С. 599–612.
Потемкина Т.М., 1985. Бронзовый век лесостепного Притоболья. М.: Наука. С. 375.
Пряхин А.Д., 1976. Поселения абашевской общности. Воронеж: Изд-во Воронежск. ун-та. 167 с.
Пряхин А.Д., 2008. Доно-донецкая степь-лесостепь в эпоху бронзы. История изучения (вторая половина XIX–начало 90-х гг. XX в.). Кн. 1. Воронеж: Издательско-полиграфический центр Воронежск. гос. ун-та. 257 с.
Пуголовок Ю.О., Володарець-Урбанович Я.В., Горбаненко С.А., Сергєєва М.С., Яніш Є.Ю., 2015. Міждисциплінарні дослідження Глинського археологічного комплексу в 2014 році // Археологічні дослідження Більсьского городища – 2014. Київ: Котельва. С. 107–128.
Сиволап М.П., 2009. Новые данные о хозяйстве племен ямной культуры центральной Украины // Материалы междун. науч. конф. “Маргулановские чтения-2008”. Караганда. С. 168–173.
Сидоров Е.А., 1989. Присваивающие виды хозяйственной деятельности населения лесостепного Приобья в 1 тыс. до н. э. // Экономика и общественный строй древних и средневековых племен Западной Сибири: Межвуз. сб. науч. трудов Новосибирск: Изд-во НГПИ. С. 16–41.
Сидоров Е.А., 1989а. Скотоводство лесостепного Приобья в I тыс. до н. э. // Советская Археология. № 3. С. 141–153.
Ситников С.М., Васильев С.К., Кирюшин К.Ю., 2007. Анализ фаунистических остатков с поселения Новоильинка III // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 11. С. 363–366.
Состояние охотничьих ресурсов в Российской Федерации в 2008–2010 гг. Информационно-аналитические материалы, 2011. Охотничьи животные России. Биология, охрана, ресурсоведение, рациональное использование. М.: Физическая культура. Вып. 9. Под ред. Н.В. Ломановой. 219 с.
Старобогатов А.М., 2018. Стоянка Черкасская 5 и ее место в раннем неолите Среднего Дона // Самарский научный вестник. Изд-во Самарск. гос. социально-педагогического ун-та. Т. 7. № 3. С. 176–189.
Тимків С.А., 2015. Остеологічні матеріали з давньоруського міста Губин // Гуманітарний журнал. № 3–4. С. 166–171.
Тимченко Н.Г., 1970. О фауне Диких и домашних животных средневековых памятников Среднего Поднепровья // Природная обстановка и фауны прошлого. Киев: Наукова Думка. № 5. С. 166–174.
Тимченко Н.Г., 1972. К истории охоты и животноводства в Киевской Руси (Среднее Поднепровье). Київ: Наукова Думка. 212 с.
Третьяков П.Н., 1959. Чаплинское городище // Материалы и исследования по археологии. М.-Л.: Изд-во АН СССР. № 70. С. 119–153.
Троицкая Т.Н., 1976. Развитие скотоводства у племен Новосибирского Приобья в 1 тысячелетии до н. э.– 5 в. н. э. // Из истории Сибири. Вып. 21. Томск. С. 155–164.
Хотинский Н.А., Алешинская З.В., Гуман М.А., Климанов В.А., Черкинский А.Е., 1991. Новая схема периодизации ландшафтно-климатических изменений в голоцене // Изв. АН СССР. Сер. геогр. № 3. С. 30–42.
Цалкин В.И., 1958. Фауна из раскопок археологических памятников Среднего Поволжья: (материалы для истории скотоводства и охоты в СССР) // Материалы и исследования по археологии СССР. М. № 61. Т. 2. С. 221–281.
Цалкин В.И., 1963. К истории млекопитающих восточноевропейской лесостепи // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 68. № 2. С. 48–62.
Цалкин В.И., 1966. Древнее животноводство племен Восточной Европы и Средней Азии. М.: Наука. 159 с.
Цалкин В.И., 1967. Фауна из раскопок поселений культуры гумельницы в СССР // Краткие сообщения о докладах и полевых исследованиях Института археологии. М.: Наука. Вып. 111. С. 43–49.
Цалкин В.И., 1969. Фауна из раскопок боршевских и роменских городищ // Советская археология. № 4. С. 91–101.
Цалкин В.И., 1970. Древнейшие домашние животные Восточной Европы. М.: Наука. 280 с.
Цалкин В.И., 1972. Фауна из раскопок андроновских памятников Приуралья // Труды Московского общества испытателей природы. Т. 48. С. 66–81.
Цвек Е.В., 1997. Новые исследования многослойного поселения трипольской культуры Березовка на Южном Буге // Новые исследования археологов России и СНГ. С.-Пб.: Ин-т истории материальной культуры РАН. С. 77–81.
Чубур А.А., 2014. Археозоология римского времени (I–IV вв. до н.э.) бассейна Нижней Судости по материалам многослойного поселения Курово 7 // Ежегодник НИИ фундаментальных и прикладных исследований. Брянск: Изд-во Брянск. гос. ун-та. Т. 5. № 1. С. 113–120.
Чубур А.А., 2018. Археозоология и обработанная кость городища раннего железного века Кудеярка (Хотялёво) // Деснинские древности: материалы IХ межгосударственной научной конференции “История и археология Подесенья”, посвященной памяти Ф.М. Заверняева. Брянск: Изд-во Брянск. гос. ун-та. С. 61–77.
Чубур А.А., 2018а. Охота и животноводство в раннем железном веке Трубчевского ополья по остеологическим материалам города Лбище // Ежегодник НИИ фундаментальных исследований Брянского гос. ун-та. Брянск: Изд-во Брянск. гос. ун-та. Т. 10. № 1. С. 59–62.
Чубур А.А., Новик Т.А., 2010. Остеологические материалы древнерусского времени из раскопок на территории Елецкого монастыря в Чернигове // Чернiгiвськi старожитностi. Вип. 3. Чернигов: Чернігів. обереги. С. 121–126.
Чубур А.А., Радюш О.А., 2013. К зооархеологической характеристике Новосильского городища // Материалы XV Всерос. науч. конф. Калуга, 2–4 апреля 2013 г. Калуга: Изд-во “Фридгельм”. С. 53–57.
Чубур А.А., Тропин Н.А., 2017. Археозоологические исследования на многослойном поселении Целыковка-2 в округе города Ельца // История: факты и символы. Елец: Изд-во Елецкого государственного университета. Т. 11. № 2. С. 86–94.
Чубур А.А., Шинаков Е.А., 2006. О современном состоянии археозоологической изученности роменско-древнеруссих поселений Брянского Подесенья // Археология юго-востока Руси. Материалы IV науч. конф. Елец: ЕГУ им. И.А. Бунина. С. 247–263.
Шамшин А.Б., 1986. Поселение Мыльниково – памятник финальной бронзы и переходного времени от эпохи бронзы к эпохе железа // Скифская эпоха Алтая (тез. докл. к конф.). Барнаул. С. 100–103.
Шамшин А.Б., Гальченко А.В., 1997. Хозяйство населения Барнаульско-Бийского Приобья в эпоху поздней бронзы и в переходное время от бронзы к железу // Источники по истории республики Алтая. Горно-Алтайск: ГАИГИ. С. 90–117.
Шрамко И.Б., Бондаренко В.Л., Задников С.А., 2006. Археозоологические материалы с поселения скифского времени у с. Барчаны // Археологическое изучение центральной России. Тез. Междун. науч. конф., посвященной 100-летию со дня рожд. В.П. Левенка (13–16 нояб. 2006 г.). Липецк: Изд-во Липецк. гос. пед. ун-та. С. 227–229.
Юго-восток европейской части СССР, 1971. Отв. ред. А.Г. Доскач, К.В. Долгополов. М.: Наука. 455 с.
Явшева Д.А., 2006. Крупные млекопитающие лесостепи Западной Сибири в голоцене // Динамика современных экосистем в голоцене. Материалы Российской научной конференции. М.: Товарищество научных изданий КМК. С. 271–276.
Яниш Е.Ю., Смольянинов Р.В., Шеменёв С.В., Желудков А.С., Юркина Е.С., Бессуднов А.Н., 2018. Проявление свидетельств охоты и рыбной ловли в материалах энеолитического поселения и могильника Васильевский кордон 27 // Стратегии жизнеобеспечения в каменном веке, прямые и косвенные свидетельства рыболовства и собирательства. Материалы междун. конф., посвященной 50-летию В.М. Лозовского. С.-Пб.: Ин-т истории материальной культуры РАН. С. 76–80.
Anderson M.J., 2001. A new method for non-parametric multivariate analysis of variance // Austral Ecology. V. 26. P. 32–46.
Bachura O.P., Kosintsev P.A., Lobanova T.P., 2019. Large mammal fauna of the West Siberian forest-tundra zone in the Late Holocene // Russian Journal of Theriology. V. 18. № 1. P. 43–50.
Bartosiewicz L., 1995. Cattle trade across the Danube at Vác (Hungary) // Animal in man’s space, man in the animal space. Proceedings of the 5th international meeting of HASRI, Genova, 23-25 Nov. 1994. P. 189–195.
Bartosiewicz L., 1999. Turkish Period bone finds and cattle trade in south-western Hungary // Historia Animalium Ex Ossibus: Beitrage zur Palaoanatomie, Archaologie, Agyptologie, Ethnologie und Geschichte der Tiermedizin. Leidorf. P. 47–56.
Bartosiewicz L., 2003. A millennium of migrations: Protohistoric mobile pastoralism in Hungary // Bulletin of the Florida Museum of Natural History. V. 44 (1). P. 101–130.
Bartosiewicz L., Gál E., 2010. Living on the frontier: “Scythian” and “Celtic” animal exploitation in Iron Age Northeastern Hungary // Anthropological approaches to zooarchaeology: complexity, colonialism, and animal transformations. Oxford: Oxbow. P. 113–128.
Benecke N., 2005. The Holocene distribution of European bison – the archaeozoological record // Munibe (Antropologia-Arkeologia). V. 57. P. 421–428.
Bokonyi S., 1974. History of domestic mammals in central and eastern Europe. Budapest: Akademiai Kiado. 598 p.
Choyke A.M., Vretemark M., Sten S., 2004. Levels of social identity expressed in the refuse and worked bone from the Middle Bronze Age Százhalombatta-Földvár, Vatya culture, Hungary. // S. Jones O’Day, W. van Neer, A. Ervynck eds: Behaviour Behind Bones. The Zooarchaeology of Ritual, Religion. Status and Identity. Oxford: Oxbow Books. P. 177–189.
Crees J.J., Carbone C., Sommer R.S., Benecke N., Turvey S.T., 2016. Millennial-scale faunal record reveals differential resilience of European large mammals to human impacts across the Holocene // Proceedings of the Royal Society B: Biological Sciences 283 (1827). P. 1–9.
Csány S., Lehoczki R., 2010. Ungulates and their management in Hungary // European ungulates and their management in the 21st century. Cambridge: Cambridge University Press. P. 219–318.
Erdős L., Ambarlı D., Anenkhonov O., Bátori Z., Cserhalmi D. et al., 2019. Where forests meet grasslands: Forest-stepes in Eurasia // Palaearctic Grasslands. № 40. P. 22–26.
Fekete G., Molnar Zs., Kun A., Botta-Dukat Z., 2002. On the structure of the Pannonian forest stepe: Grasslands on sand // Acta zoologica Academiae Scientiarum Hungaricae. № 48. P. 137–150.
Gál E., 2009. Animal bone offerings from the Baden Cemetery at Budakalász // Bondár M., Raczky P. (eds). The Coper Age cemetery of Budakalász, Budapest, Pytheas. P. 371–379.
Horváth T., 2012. Animal deposits in the Late Coper Age Settlement of Balatonőszöd-Temetői dűlő, Hungary // The ritual killing and Burial of Animals by A. Pluskowski. Oxbow. P. 115–136.
Legendre P., Gallagher E.D., 2001. Ecologically meaningful transformations for ordination of species data // Oecologia. V. 129. P. 271–280.
Legendre P., Legendre L., 2012. Numerical Ecology. V. 24. 3th ed. 1006 p.
Lister A.M., Stuart A.J., 2019. The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence // Quaternary International. V. 500. P. 185–203.
Lorenzen E.D., Nogue’s-Bravo D., Orlando L., Weinstock J., Binladen J. et al., 2011. Species-specific responses of Late Quaternary megafauna to climate and humans // Nature. V. 479. P. 359–365.
Lyublyanovics K., 2015. The Socio-Economic Integration of Cumans in Medieval Hungary. An Archaeozoological Aproach. Unpublished PhD-dissertation defended at CEU, Budapest. 675 p.
Oksanen J., Blanchet F.G., Friendly M., Kindt R., Legendre P. et al., 2019. vegan: Community Ecology Package. R package version 2.5-6. [Электронный ресурс] Режим доступа: https://CRAN.R-project.org/package=vegan
Paradis E., Schliep K., 2019. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R // Bioinformatics. V. 35. № 3. P. 526–528.
QGIS Geographic Information System. QGIS Association. [Электронный ресурс] Режим доступа:: http:// www.qgis.org/
R Core Team, 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [Электронный ресурс] Режим доступа: https://www.r-project.org/
Rosvold J., Andersen R., Linnell J.D.C., Hufthammer A.K., 2013. Cervids in a dynamic northern landscape: Holocene changes in the relative abundance of moose and red deer at the limits of their distributions // Holocene. V. 8. № 23. P. 1143–1150.
Sommer R.S., Zachos F.E., Street M., Jöris O., Skog A., Benecke N., 2008. Late Quaternary distribution dynamics and phylogeography of the red deer (Cervus elaphus) in Europe // Quaternary Science Reviews. V. 27. № 7–8. P. 714–733.
Sommer R.S., Zachos F.E., 2009. Fossil evidence and phylogeography of temperate species: 'Glacial refugia' and post-glacial recolonization // Journal of Biogeography. V. 36. № 3. P. 2013–2020.
Sommer R.S., Benecke N., Lõugas L., Nelle O., Schmölcke U., 2011. Holocene survival of the wild horse in Europe: a matter of open landscape? // Journal of Quaternary Science. V. 26. P. 805–812.
Sommer R.S., Kalbe J., Ekström J., Benecke N., Liljegren R., 2014. Range dynamics of the reindeer in Europe during the last 25.000 years // Journal of Biogeography. № 41. P. 298–306.
Schmolcke U., Zachos F.E., 2005. Holocene distribution and extinction of the moose (Alces alces, Cervidae) in Central Europe // Mammalian Biology. V. 70. № 6. P. 329–344.
The Early Neolithic on the Great Hungarian Plain: investigations of the Körös culture site of Ecsegfalva 23, County Békés. 2007. Varia archaeologica Hungarica. V. 21. Budapest: Institute of Archaeology, Hungarian Academy of Sciences. 808 p.
Wickham H., 2016. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag. 211 p.
Wickham H., Averick M., Bryan J., Chang W., McGowan L.D’A. et al., 2019. Welcome to the tidyverse // Journal of Open Source Software. V. 4. № 43. P. 1–6.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал