

Зоологический журнал, 2021, T. 100, № 12, стр. 1422-1438
Наблюдения за морскими млекопитающими и птицами в Антарктике: “Экологические отпечатки” распределения в океане во время 79-го рейса НИС “Академик Мстислав Келдыш”
С. П. Харитонов a, *, А. В. Третьяков a, А. Л. Мищенко a, Н. Б. Конюхов a, А. Е. Дмитриев a, С. М. Артемьева b, Г. Ю. Пилипенко c
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
b Научно-исследовательский зоологический музей Московского государственного университета
имени М.В. Ломоносова
125009 Москва, Россия
c Географический факультет Московского государственного университета
имени М.В. Ломоносова
119991 Москва, Россия
* E-mail: serpkh@gmail.com
Поступила в редакцию 12.02.2021
После доработки 24.03.2021
Принята к публикации 26.03.2021
Аннотация
Работа научно-исследовательского судна “Академик Мстислав Келдыш” в водах к югу от южной оконечности Аргентины, в проливе Дрейка, окрестностях Антарктического полуострова, море Скоша и северной части моря Уэдделла проходила в два этапа: с 16 января по 6 февраля (далее – январь) и с 8 февраля по 3 марта (далее – февраль) 2020 г. Предложен новый метод анализа результатов экологических исследований, и на примере этого метода представлены новые сведения об экологии и о пространственном распределении морских млекопитающих и морских птиц Антарктики. “Экологические отпечатки” некоторых видов морских млекопитающих и птиц очень похожи, следовательно, разные виды (вне зависимости от их систематической принадлежности) характеризуются одинаковым отношением к условиям среды и одинаковым пространственным распределением. Финвал, горбатый кит, снежный буревестник, пингвин Адели, антарктический буревестник и антарктический глупыш в январе были похожи друг на друга по характеру экологических отпечатков, который указывает на приуроченность этих видов к акваториям с айсбергами и/или битым льдом. В феврале связь китов со льдами ослабла, что стало заметно по их экологическим отпечаткам, а у всех перечисленных выше видов морских птиц такая связь сохранилась. Экологические отпечатки хорошо демонстрируют ширину абиотической экологической ниши, которую занимают конкретные виды и группы видов. Экологические отпечатки могут показать изменение областей распределения в конкретные периоды (южный королевский альбатрос), смену стратегии использования акватории (антарктический морской котик), а также другие экологические особенности, в том числе пока нераскрытые.
В декабре 2019 г. по поручению правительства Российской Федерации начала выполняться Программа комплексных экспедиционных исследований экосистемы Атлантического сектора Южного океана. В программе участвует более 10 институтов, головным из которых является Институт океанологии им. П.П. Ширшова РАН (Морозов и др., 2019, 2020). В рамках этой программы есть проект Института проблем экологии и эволюции им. А.Н. Северцова РАН: “Ресурсные исследования экосистемы Южного океана (Атлантический сектор Антарктики)”, т.е. поставлена задача изучения биоты Антарктики в современных условиях изменения климата. Работа научно-исследовательского судна “Академик Мстислав Келдыш” в водах к югу от южной оконечности Аргентины, проливе Дрейка, окрестностях Антарктического п-ова, море Скоша и северной части моря Уэддела проходила в два этапа: с 16 января по 6 февраля и с 8 февраля по 3 марта 2020 г. С 6 по 8 февраля был осуществлен промежуточный заход в порт Ушуайя, Аргентина. Общая длина маршрута составила примерно 6000 км на каждом этапе, всего в Антарктических водах судно прошло примерно 12 000 км.
Исследований по экологии морских млекопитающих Антарктики, преимущественно китообразных, существует множество (Joiris, Dochy, 2013; Friedlaender at al., 2006; Kasamatsu et al., 2000; Mondreti et al., 2020 и др.). Также немало работ посвящено экологии птиц Антарктики (например, Lyver et al., 2011; Ainley et al., 2010; Santora et al., 2009; Mondreti et al., 2020; Serratosa et al., 2020 и др.) При этом отдельные экологические параметры среды обычно рассматриваются изолировано друг от друга. В данном исследовании нам хотелось бы подняться на более высокий уровень (метауровень) рассмотрения экологических факторов и сделать попытку обобщить оценку их воздействия, чтобы, сконцентрировав действие факторов среды в малое число метапараметров (в нашем случае – в два метапараметра), постараться лучше понять адаптации, экологические возможности, а в ряде случаев и экологические предпочтения отдельных видов морских млекопитающих и птиц.
Нами в работах Харитонова с соавторами (Kharitonov et al., 2021, 2021a, 2021b) изложены предварительные результаты исследований влияния разных экологических факторов на встречаемость морских млекопитающих и морских птиц путем факторного анализа. В данном сообщении мы вышли на следующий уровень обобщения результатов этого анализа. Предлагаемый нами здесь “метод экологических отпечатков”, по всей видимости, является новым подходом. Если мы имеем дело с большим количеством данных, их бывает полезно свернуть в 1–2–3 численных показателя, например – главные компоненты (Коросов, 1996). Однако хотелось бы в дополнение к численным характеристическим показателям иметь еще и визуальную информационную характеристику, нечто вроде “визуального индекса”, т.е. такой тип графика (диаграммы), который позволял бы быстро получить представление об экологических особенностях биологических объектов в рассматриваемое время и в рассматриваемом месте, включая и пространственное распределение этих объектов. Метод экологических отпечатков позволяет увидеть и оценить совокупность экологических факторов, связанных с теми или иными группами животных и временны́ми периодами. Цель данного сообщения – предложить новый метод анализа экологических результатов и на примере этого метода представить новые сведения об экологии морских млекопитающих и птиц Антарктики.
МАТЕРИАЛЫ И МЕТОДЫ
Полевые наблюдения. Наблюдения и учет встречаемости морских млекопитающих и птиц проводили с борта судна по маршрутам: 1) 16 января–6 февраля 2020, вход в пролив Бигль – пролив Дрейка – окрестности вершины Антарктического полуострова – Южные Шетландские о-ва – Южные Оркнейские о-ва – обратно к Антарктическому п-ову – пролив Дрейка – пролив Бигль, далее в статье это период будет называться “январь”; 2) 8 февраля–3 марта 2020, пролив Бигль – пролив Дрейка – вершина Антарктического п-ова – бассейн Пауэлла – вершина Антарктического п-ова – пролив Дрейка – пролив Бигль – Ушуайя, далее это период будет называться “февраль” (рис. 1).
Рис. 1.
Карта маршрутов НИС “Академик Мстислав Келдыш” в январе (фиолетовая линия) и феврале (красная линия).
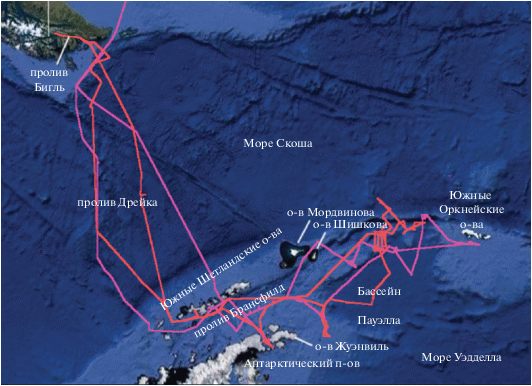
Наблюдения проводили в светлое время суток в основном с пеленгаторной палубы корабля два наблюдателя, которые стояли по правому и левому борту корабля. Продолжительность наблюдений была сопряжена со сменой вахт на корабле и для каждой пары наблюдателей составляла 4 ч, однако утром и вечером, в зависимости от времени восхода и захода солнца, периоды наблюдений могли быть короче. Наблюдатели, кроме биноклей, были оснащены современными фотоаппаратами с длиннофокусными объективами. Почти все встреченные морские млекопитающие были сфотографированы, снять удалось также подавляющее большинство встреченных птиц. Во время наблюдений фиксировали вид, число особей в группе, их поведение, наличие льда (айсберги) и общие показатели погоды. На протяжении всего маршрута судна три полевых GPS навигатора осуществляли запись координат, поэтому для каждой встречи с биологическими объектами в океане было известно ее точное место.
Всего по данному району атлантического сектора Южного океана собраны данные по 1142 встречам с морскими млекопитающими и 5425 встречам с морскими птицами. Встречены представители 23 видов или систематических групп морских млекопитающих и представители 42 видов или систематических групп морских птиц. Однако для экологического анализа годились только те виды, для которых собранный материал был достаточно обширным. Среди морских млекопитающих таких видов всего три: антарктический морской котик (Arctocephalus gazella), горбатый кит (Megaptera novaeangliae), финвал (Balaenoptera physalus). Среди птиц таким критериям соответствовали 18 видов или групп видов: пингвин Адели (Pygoscelis adeliae), антарктический пингвин (Pygoscelis antarcticus), вильсонова качурка (Oceanites oceanicus), чернобрюхая качурка (Fregetta tropica), странствующий альбатрос (Diomedea exulans), южный королевский альбатрос (Diomedea epomophora), светлоспинный альбатрос (Phoebetria palpebrata), чернобровый альбатрос (Thalassarche melanophris), сероголовый альбатрос (Thalassarche chrysostoma), южный гигантский буревестник (Macronectes giganteus), северный гигантский буревестник (Macronectes halli), антарктический глупыш (Fulmarus glacialoides), капский голубок (Daption capense), антарктический буревестник (Thalassoica Antarctica), снежный буревестник (Pagodroma nivea), прионы двух видов (Pachyptila sp.)11, белогорлый буревестник (Procellaria aequinoctialis), антарктический поморник (Stercorarius antarcticus).
Дополнительные инструментальные методы. Глубина океана во время маршрута записывалась по показаниям научного эхолота фирмы Kongsberg EA600, частота излучения 12 кГц. Однако не всегда было возможно снимать показания глубины для каждой точки, где были встречены птицы и млекопитающие. Эхолот не работал при сильном волнении, а также во время пребывания на “станциях” – в точках взятия гидрологических проб. В этих случаях мы вычисляли положение ближайшей точки, где глубина была определена, и эти данные использовали для анализа. Обычно ближайшая точка находилась на расстоянии менее 3 км от точек встреч биологических объектов.
Благодаря работе метеостанции Davis Vantage Pro 2 под управлением команды сотрудников Тихоокеанского океанографического института имени В.И. Ильичева ДВО РАН, каждые 5 мин фиксировались общие параметры погоды. Температура воды на разных глубинах фиксировалась только во время взятия гидрологических проб при помощи спускаемых аппаратов “Розетта” (SBE9 и SBE19).
Статистическая обработка производилась на программном пакете Statistica-12, StatSoft Inc. В дополнение к этому, для сравнения средних вместо общепринятых критериев Стьюдента и Манн-Уитни, мы использовали критерий Бейли (Плохинский, 1978). Большое достоинство этого критерия в том, что он годится для любых типов распределений, а не только для нормального распределения, и обладает большей статистической мощностью, чем критерий Маннa-Уитни, поскольку последний является непараметрическим (ранговым), а критерий Бейли – параметрический. Критерий Бейли обладает той же статистической мощностью, что и критерий Стьюдента, но не содержит требования обязательного нормального распределения данных. Для картографических работ использовалась программа MapInfo-12.5.
Описание метода экологических отпечатков. Для визуальной оценки и сравнения набора экологических параметров отдельных видов млекопитающих и птиц мы строили специальные лепестковые диаграммы, которые в данном случае можно назвать экологическими отпечатками объектов. Объектами могли служить отдельные виды, обнаруженные за весь период работы (январь–февраль), или те же виды, но за меньший отрезок времени: январь или февраль. Было выбрано 10 экологических параметров: широта, долгота, глубина, волнение моря, температура воздуха, влажность воздуха, скорость ветра, скорость ветра при порывах, фактор ветрового охлаждения, атмосферное давление. Все эти параметры равнозначно использовались нами ранее в факторном анализе (Kharitonov et al., 2021). В данном сообщении мы размещаем значения этих параметров на одной диаграмме с целью более удобной визуализации комплекса экологических факторов, связанных с разными биологическими объектами. Для каждого параметра выбирались общие минимум и максимум значений за весь (т.е. двухмесячный) период работы экспедиции, максимальному значению каждого параметра соответствовал самый внешний круг лепестковой диаграммы, минимум значений каждой оси (каждого параметра) находился в центре круга диаграммы. Эти минимумы и максимумы для каждого параметра определялись по принципу: какие минимальные и максимальные значения отмечены за рассматриваемый период, вне зависимости от того, наблюдался при таких параметрах рассматриваемый вид или нет. Поскольку наблюдения вели только в светлое время суток, общие минимум и максимум определяли только для светлого времени. Например, выясняли, какие минимальные и максимальные температуры воздуха наблюдались в рассматриваемый период, но только в светлое время суток. Каждый конкретный вид не обязательно наблюдали при минимальных и максимальных температурах рассматриваемого периода, он мог отмечаться в более узком диапазоне температур. И в этих диапазонах каждый вид имел свои реальные минимум и максимум температур, при которых он наблюдался. Соответственно, из набора наблюдений можно было стандартным образом подсчитать среднее значение параметра, при котором наблюдался каждый вид. Подобные вычисления производили для каждого параметра. Широта и долгота в данном исследовании рассматриваются исключительно в численном виде, т.е. не градусы и минуты с указанием полушария, а градусы с десятичными долями и со знаком минус. Знак минус для широты означает южную широту, для долготы – западную долготу. Поэтому более западные значения долготы и более южные значения широты на диаграммах расположены в области минимумов.
Учитывая, что разные параметры одинаковым образом используются в факторном анализе, мы решили увеличить уровень обобщения значения параметров: установить некие одинаковые условные единицы, в которые затем и перевести значения параметров. Для этого значения в интервале от общего минимума до общего максимума принимали равными 100 условным единицам. Таким образом, получалось, что в пределах каждого параметра каждая такая условная единица содержала разные реальные единицы (градусы географических координат, баллы шкалы Бофорта, градусы Цельсия, миллиметры ртутного столба и проч.), причем разное число единиц в зависимости от параметра. Однако теперь все параметры имели одинаковую шкалу измерения. После этого можно было находить минимум, максимум и среднее по всем параметрам в условных единицах. Таким образом, вместо 30 значений для 10 параметров, в нашем случае мы получаем только 3 значения, которые затем сравниваем между видами и между месяцами. В реальности оказалось, что от рассмотрения минимумов чаще всего можно отказаться, интерес для вида представляют средние и максимальные значения всех параметров, выраженные в условных единицах. Эти значения позволяют лучше понять, как какой-либо вид в рассматриваемый период встраивался в те пределы экологических параметров, которые были отмечены в данном месте и в данное время. Кроме того, для каждого такого случая имеется и вышеописанная диаграмма (“экологический отпечаток”), форма которой также позволяет судить о характере пребывания вида при наблюдавшихся в рассматриваемый период экологических условиях.
При разработке этого метода была написана специальная компьютерная программа на языке Visual Basic-3 (Харитонов, 2021). При помощи этой программы, на основании данных из входного текстового файла, содержащего информацию об экологических параметрах, общие минимумы и максимумы по каждому параметру и набор конкретных данных, создается рисунок экологического отпечатка, представленного во входном файле объекта, а также осуществляются все необходимые вычисления в реальных и условных единицах измерения для каждого параметра.
РЕЗУЛЬТАТЫ
В феврале район обследования был немного меньше распространен в долготном направлении на восток, чем в январе: в феврале судно не посетило окрестности Южных Оркнейских о-вов. Однако в феврале судно заходило более чем на 100 км западнее, чем в январе; широтное покрытие для обоих месяцев было практически одинаковым (рис. 1, табл. 1). Интервалы значений большинства экологических параметров, наоборот, – в январе были несколько у́же, чем в феврале (табл. 1). Если для глубин, волнения моря, скоростей ветра и порывов, а также атмосферного давления разница была невелика, то верхний предел температуры воздуха (и, соответственно, фактор ветрового охлаждения) в феврале был существенно выше, чем в январе (табл. 1). Максимальные показатели влажности в оба месяца совпадают, однако минимальные значения в январе были значительно выше, что означает в целом более высокую влажность воздуха в этом месяце (табл. 1). Таким образом, мы можем ожидать, что по своим параметрам январь в Антарктике более холодный и влажный месяц, чем февраль. Кроме того, северная граница плавучих льдов в январе проходила севернее, чем в феврале, когда эта граница сместилась ближе к Антарктиде (рис. 2, 3). Можно сказать, что январь в Антарктике соответствует июлю в высокой Арктике, а февраль – августу.
Таблица 1.
Общие минимумы и максимумы в значениях экологических параметров во время рейсов в Антарктику
| Месяц | Широта (DD.DDDD) | Долгота (DD.DDDD) | Глубина, м | Волнение, баллы Бофорта | Темп. воздуха,°C | Влажность, % | Скорость ветра, м/с | Скорость ветра при порывах, м/с | Фактор ветрового охлаждения, °C | Атмосферное давление, мм рт. столба | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Январь | Минимум | –63.48524 | –67.15015 | 58.82 | 0 | –2.0 | 65 | 0.0 | 0.4 | –7.9 | 734.8 |
| Максимум | –54.36893 | –45.06985 | 5570.57 | 7 | 12.7 | 97 | 22.4 | 27.3 | 12.4 | 771.2 | |
| Февраль | Минимум | –63.51173 | –68.10433 | 49.89 | 0 | –1.8 | 35 | 0.0 | 0.4 | –11.0 | 732.6 |
| Максимум | –54.80998 | –48.23468 | 5604.52 | 8 | 21.4 | 97 | 23.7 | 28.2 | 20.8 | 766.6 | |
| Январь + | Минимум | –63.51173 | –68.10433 | 49.89 | 0 | –2.0 | 35 | 0.0 | 0.4 | –11.0 | 732.6 |
| + февраль | Максимум | –54.36893 | –45.06985 | 5604.52 | 8 | 21.4 | 97 | 23.7 | 28.2 | 20.8 | 771.2 |
Рис. 2.
Встречи финвалов (символ “горизонтальный кит”) и горбатых китов (символ “наклонный кит”) в январе (A) и феврале (B). Бирюзовая линия – маршрут в январе, красная линия – маршрут в феврале. Бирюзовыми треугольниками обозначены области айсбергов и колотого льда.
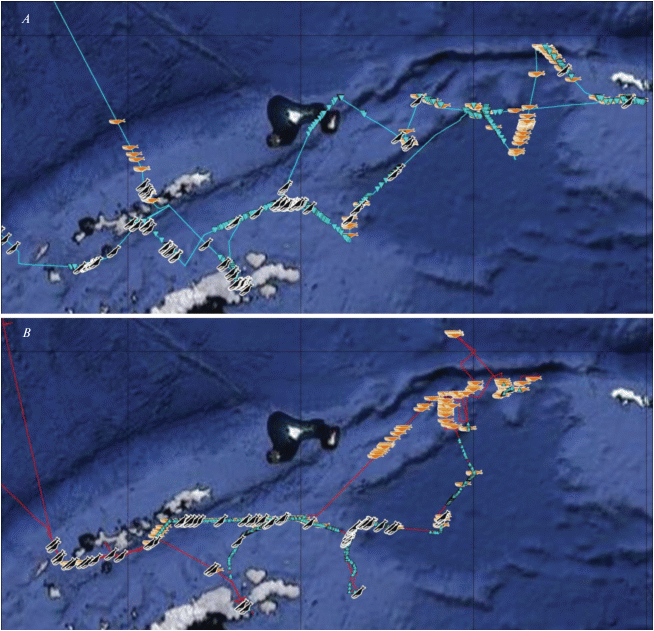
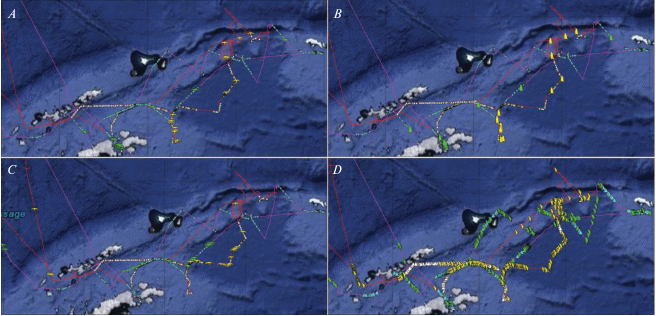
Рис. 3.
Встречи ряда видов птиц в связи с ледовой обстановкой: A – снежный буревестник, B – пингвин Адели, C – антарктический буревестник, D – антарктический глупыш. Фиолетовая линия – маршрут в январе, красная линия – маршрут в феврале. Бирюзовыми треугольниками обозначены области айсбергов и колотого льда в январе, белыми треугольниками – в феврале. Символы “птица” и “пингвин”, окрашенные зеленым, – встречи в январе, окрашенные желтым – встречи в феврале.
При построении экологических отпечатков для совместимости результатов января и февраля использовались только общие минимумы и максимумы за два месяца (табл. 1). Основной полученный результат, т.е. сами экологические отпечатки, средние значения факторов в условных единицах и средние максимальные значения факторов в тех же условных единицах представлены в табл. 2.
Таблица 2.
Экологические отпечатки морских млекопитающих и птиц с указанием средних и максимальных значений экологических параметров в условных единицах
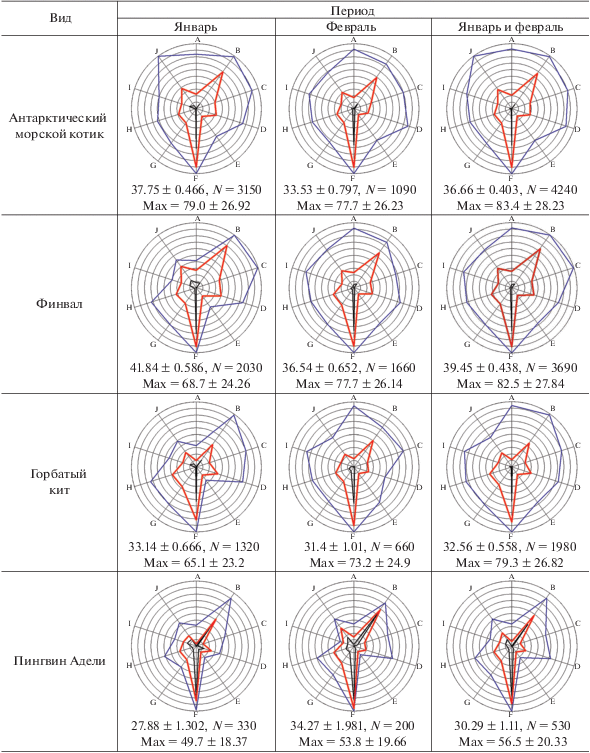
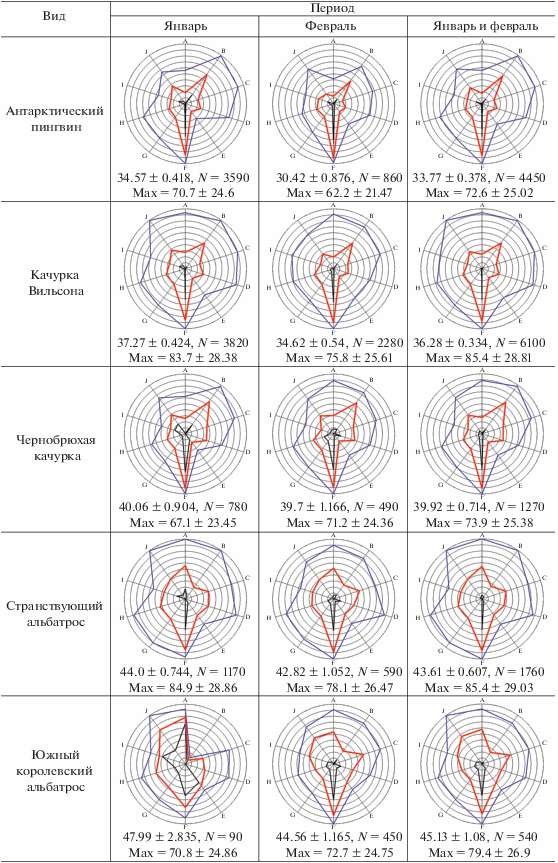
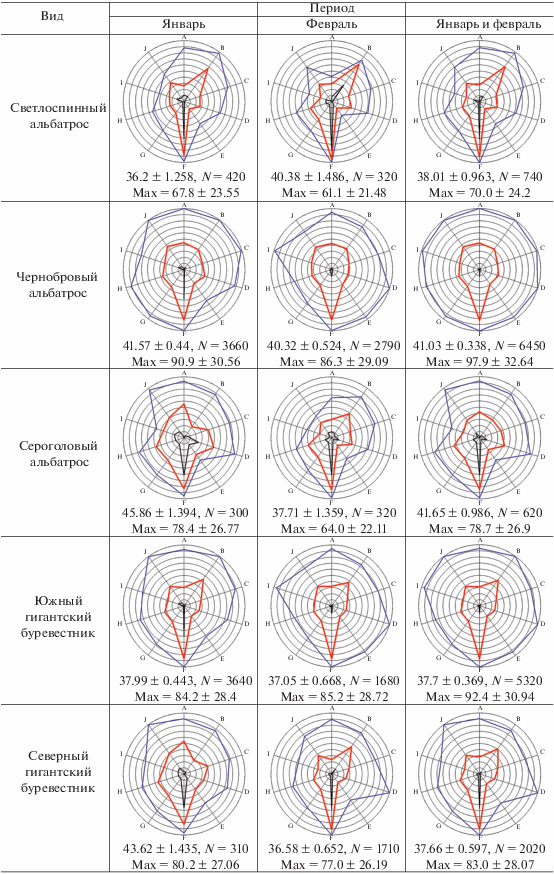
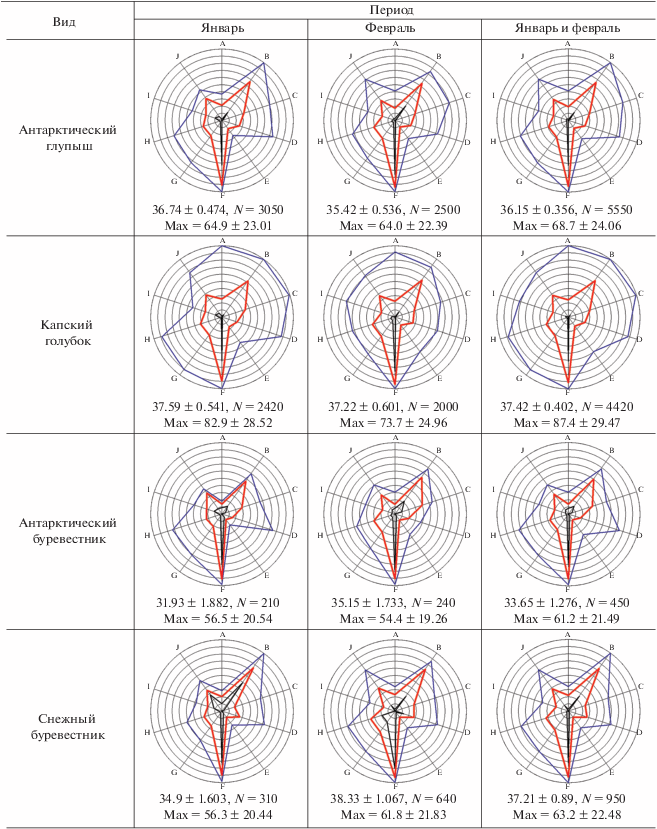
Таблица 2. Окончание
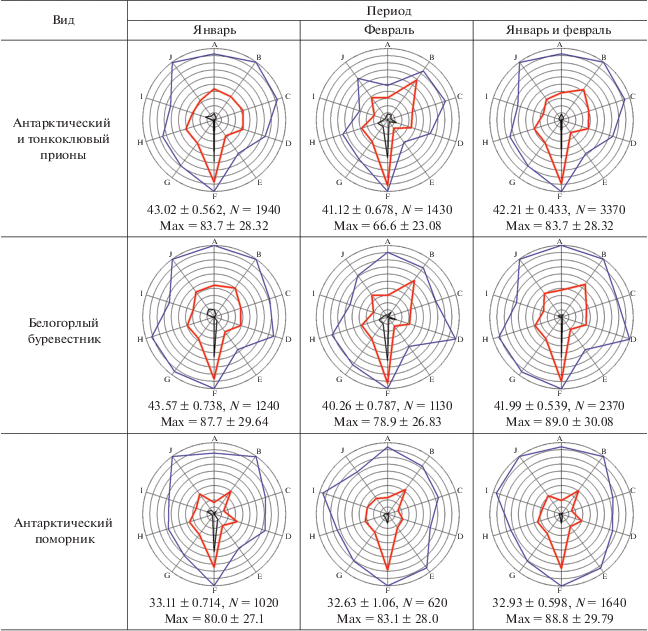
[i] Примечания. N для максимумов равно числу экологических параметров, в данном сообщении это 10. Обозначения осей: A–широта, B–долгота, C–глубина, D–волнение моря, E–температура воздуха, F–влажность, G–скорость ветра, H–скорость ветра при порывах, I–фактор ветрового охлаждения, J–атмосферное давление.
ОБСУЖДЕНИЕ
Замечания к применению экологических отпечатков. Следует иметь в виду, что экологический отпечаток не отражает все адаптации, которыми обладает вид или популяция. Отпечаток показывает, каким образом вид или популяция встраиваются в диапазон экологических параметров, отмечаемых в данном месте и в рассматриваемый промежуток времени. При этом представители одного и того же вида в разных местах, для которых в данный момент характерны другие значения экологических параметров, могут встраиваться в диапазон этих параметров и могут быть распределены по акватории совсем по-другому. Например, встречаемость чернобрового альбатроса в январе положительно связана с атмосферным давлением, а в феврале – отрицательно (Kharitonov et al., 2021а), что сказалось на падении максимума данного параметра у этого вида в феврале (табл. 2). Спутниковое прослеживание этого вида показало, что вид действительно предпочитает теплый воздух, тем не менее часто использует холодные районы Антарктики для добывания пищи (Wakefield et al., 2011).
При этом экологические параметры в ряде случаев могут быть взаимозаменяемы. Для данного района Антарктики важно выяснить встречаемость и пространственное распределение морских млекопитающих и птиц в зависимости от температуры воды. Однако температуру воды регистрировали не постоянно, а только при проведении гидрографических измерений (Морозов и др., 2020, 2020a). Данные этих измерений нам были предоставлены Е.Г. Морозовым. Эти измерения были неравномерно распределены во времени и в пространстве, связать их результаты со всеми наблюдениями за птицами и млекопитающими едва ли возможно. Однако анализ показал наличие почти функциональной связи между температурой воды и температурой воздуха в Антарктике во время наших работ: коэффициент корреляции между этими температурами равен 0.95 (N = 113 параллельных измерений температуры воздуха и поверхностного слоя воды). Кроме того, средняя разница между температурой воздуха и температурой воды в январе составляла 0.4403 ± 0.14778°C (N = 62), для февраля – 0.4431 ± ± 0.1714°C (N = 51). Между указанными месяцами этот показатель достоверно не различается (t = 0.0124, P = 0.99), т.е. абсолютно одинакова. Кроме того, за весь период наблюдений не обнаружено резких вторжений холодного или теплого воздуха в район исследований. Таким образом, вместо температуры воды можно использовать температуру воздуха, и то отношение к температуре воздуха у морских птиц и млекопитающих, которое мы выявили в данном исследовании, автоматически распространяется и на температуру поверхностного слоя воды.
Толкование экологических отпечатков. Для толкования экологических отпечатков их полезно разделить на группы по: 1) визуальной похожести экологических отпечатков; 2) сходству средних значений; 3) сходству максимальных значений; 4) сходству разницы между максимальным и средним значениями. Причем для пунктов 2), 3) и 4) имеются в виду не только численные значения, но, так же как и для пункта 1), форма самих графиков.
Первая экологическая особенность, на которую могут указывать экологические отпечатки, – это предпочтение районов моря с особыми условиями. При просмотре выявляется ряд сходных диаграмм, причем диаграммы китов похожи на диаграммы некоторых видов птиц. Экологические отпечатки являются ярким отражением такой экологической особенности видов, как отношение антарктических видов (птиц и млекопитающих) к ледовой обстановке и их пространственного распределения в зависимости от распределения плавающих льдов. Отпечатки финвала и горбатого кита за январь, пингвина Адели, антарктического буревестника, снежного буревестника и антарктического глупыша за оба месяца оказались “скошены” в левом верхнем секторе диаграммы, а также имеют прогиб графика на оси температур воздуха. Последнее указывает на приуроченность этих биологических объектов к низким температурам воздуха, а значит – и воды. Нанесение точек встреч птиц и китов на карту демонстрирует, что такой тип экологического отпечатка наблюдается у морских организмов (неважно, птиц или млекопитающих), если их присутствие в рассматриваемый период связано с ледовыми полями: айсбергами или полями битого льда.
Таблица 3.
Средние значения экологических параметров в январе и феврале в безледовых и ледовых областях маршрута судна
| Экологический параметр | Январь | Февраль | ||
|---|---|---|---|---|
| Безо льдов (2270 наблюдений) |
Со льдами (895 наблюдений) |
Безо льдов (1566 наблюдений) |
Со льдами (817 наблюдений) |
|
| Широта (DD.DDDDD) | –60.24733 ± 0.04930 |
–61.76117 ± 0.02956 |
–59.89386 ± 0.06485 |
–62.26812 ± 0.02029 |
| Долгота (DD.DDDDD) | –58.07083 ± 0.14591 |
–52.00558 ± 0.14311 |
–57.25356 ± 0.17949 |
–53.72052 ± 0.09645 |
| Глубина, м | 1781.8 ± 27.09 | 1420.8 ± 40.21 | 1734.0 ± 32.01 | 1436.9 ± 35.71 |
| Волнение, баллы Бофорта | 2.5 ± 0.04 | 2.2 ± 0.05 | 2.4 ± 0.03 | 1.5 ± 0.03 |
| Температура воздуха, °C | 3.7 ± 0.06 | 1.3 ± 0.04 | 4.4 ± 0.13 | 0.2 ± 0.04 |
| Влажность, % | 87.2 ± 0.16 | 90.0 ± 0.19 | 87.6 ± 0.32 | 92.0 ± 0.18 |
| Скорость ветра, м/с | 8.2 ± 0.09 | 6.4 ± 0.12 | 7.9 ± 0.09 | 7.0 ± 0.10 |
| Скорость ветра при порывах, м/с | 10.4 ± 0.11 | 8.1 ± 0.14 | 10.1 ± 0.10 | 8.7 ± 0.11 |
| Фактор ветрового охлаждения, °C | –1.4 ± 0.08 | –3.5 ± 0.07 | –0.7 ± 0.18 | –5.4 ± 0.08 |
| Атмосферное давление, мм рт. столба | 748.4 ± 0.18 | 745.9 ± 0.16 | 748.7 ± 0.22 | 744.9 ± 0.13 |
Экологические отпечатки в январе у горбатых китов и финвалов очень похожи. Нанесение точек встреч обоих видов на карту показало, что в этом месяце представители и того и другого вида встречались преимущественно в районе льдов (рис. 2). В районе плавучих льдов и в январе, и в феврале почти все показатели погоды (кроме влажности воздуха), а также волнение моря имели меньшее значение, чем в районах, где плавучие льды не встречались (табл. 3). Для этой таблицы средние значения параметров в ледовых и свободных ото льда областях в январе и феврале получены на основании данных по встречам только птиц, без учета встреч млекопитающих. Это сделано, чтобы избежать повторов при расчетах, поскольку моменты встречи с морскими млекопитающими часто совпадали по времени со встречами птиц. Влажность воздуха в районах плавучих льдов была выше, чем в областях безо льда. Хотя финвалы явно тяготели к восточному сектору района работ, а горбатые киты – к западному, льды “уравняли” эти виды, т.е. сделали их похожими друг на друга по экологическим отпечаткам. К февралю горбатые киты ушли с северо-востока, а финвалы продвинулись дальше на запад. При этом горбатые киты преимущественно остались в районе айсбергов, а финвалы встречались везде, в районе как со льдами, так и без них. Кроме того, горбатые киты в большинстве своем к февралю не только сократили ареал на востоке прежней области пребывания, но и увеличили свое присутствие в западной части области встреч, которая в феврале оказалась безо льда. В результате у обоих видов китов в феврале экологические отпечатки стали менее “ледовыми”. При этом у финвала, чье пребывание в этом месяце, скорее всего, не было приурочено именно к району льдов, отпечатки стали совсем другими. Февральские горбатые киты имеют в своих экологических отпечатках большее влияние льда: меньший максимум в области атмосферного давления и больший прогиб линии средних в области температур воздуха (хотя максимумы температур воздуха у обоих видов китов оказались близки) (табл. 2). Однако в экологических отпечатках у горбатых китов средняя влажность воздуха была значительно ниже именно в январе. Этот феномен, вероятно, связан с более западным распространением горбатых китов, а влажность воздуха на западе исследуемого района была меньше, чем на востоке (коэффициент корреляции Пирсона r = = 0.49, N = 3165, P = 0.0002). В этом плане, действительно, визуально общая ледовая обстановка на западе была не столь сложной, как на востоке, хотя детальной количественной оценки мощности ледовых полей провести не удалось.
Такой же “ледовый” характер экологических отпечатков имели и некоторые птицы: антарктический буревестник, пингвин Адели, снежный буревестник и, как ни странно, антарктический глупыш – широко распространенный вид, пространственное распределение которого при визуальных оценках в море никак не кажется связанным со льдами (рис. 3). Пингвины Адели в феврале были представлены преимущественно молодыми особями, которым, по-видимому, часто требуется отдых на льдинах. Про антарктического буревестника известно, что его распределение в море приурочено к районам с айсбергами, поскольку эти птицы предпочитают отдыхать не на воде, а на айсбергах (Delord et al., 2020). Однако, как видно по экологическим отпечаткам, эти птицы не всегда тяготеют к айсбергам, а могут находиться и довольно далеко от них: в феврале экологические отпечатки этого вида уже менее “ледовые”, чем в январе. Снежные буревестники вообще тяготели ко льдам, которые присутствовали в восточной части обследованного района, как в январе, так и в феврале (рис. 3). Что касается антарктического глупыша, то “ледовый” характер экологических отпечатков у этого вида был и в январе, и в феврале. Однако, судя по распределению в море (рис. 3), эти птицы не тяготеют именно ко льдам, видимо, они просто предпочитают (или при необходимости образуют кормовые скопления) холодные и с высокой влажностью воздуха районы океана.
Прионы, наоборот, именно в феврале, когда льдов было меньше, оказались более “ледовыми”. В норме прионы имеют обширную область кормовых разлетов (Quillfeldt et al., 2014). Светлоспинный альбатрос тоже оказался более “ледовым” в феврале, когда льдов стало меньше, подобная тенденция отмечается и у белогорлого буревестника. Возможно, это связано уже с кормовыми предпочтениями видов. Ранее мы предположили, что прионы, светлоспинный альбатрос и белогорлый буревестник, возможно, тяготеют к бассейну Пауэла и впадине Гесперид в феврале, поскольку там отмечен более “чистый” криль (Euphausia superba), с наименьшей примесью сальп (Salpa tompsoni) (Kharitonov et al., 2021а). А эти районы как раз и были более “ледовыми” в феврале (рис. 3).
Вторая экологическая особенность, на которую могут указывать экологические отпечатки – это широта области распространения. Для выяснения этого не обязательно наносить точки встреч на карту, что не всегда бывает удобно. Южные королевские альбатросы в январе были немногочисленны и встречены в узкой области на западе исследуемого района, о чем хорошо сигнализирует январский отпечаток этого вида: долгота встреч близка к общим минимальным значениям.
Третья особенность, о которой могут сигнализировать отпечатки, – это широта экологических условий, в которых может находиться вид. Максимумы показывают, где данный вид мог находиться, а средние значения показывают, в каких условиях он чаще всего находился. Например, антарктический поморник имеет по всем параметрам большие максимумы, значит адаптирован к широкому спектру условий, но средние значения значительно меньше максимумов (табл. 2), т.е. представители этого вида предпочитали гораздо более мягкие условия, далекие от крайних. Антарктические поморники – довольно редкие птицы в водах Антарктики вдали от берегов. Однако подобной особенностью обладают и некоторые широко распространенные океанические виды: качурка Вильсона, южный гигантский буревестник и чернобровый альбатрос. Видом с самыми широкими экологическими предпочтениями, судя по максимумам, является чернобровый альбатрос (табл. 2). Недаром представители этого вида временами встречаются в северном полушарии в окрестностях Новой Земли и Земли Франца-Иосифа (Покровская и др., 2018). Судя по экологическим отпечаткам, южный гигантский буревестник, тоже очень многочисленный и распространенный в Антарктике вид, хотя и является конкурентом чернобрового альбатроса (Kharitonov et al., 2021а), все же не является столь же “экологически широким” видом как последний.
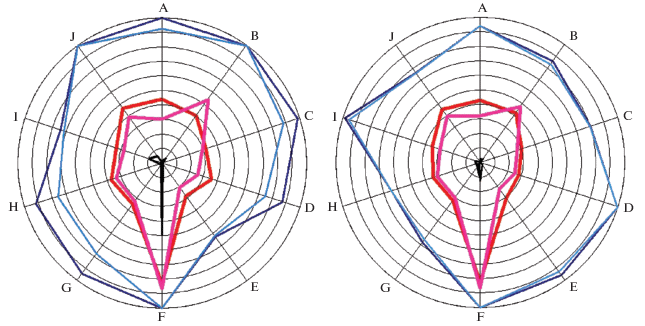
Что касается конкуренции между чернобровым альбатросом и южным гигантским буревестником, то и в этом случае метод экологических отпечатков позволил выявить интересную особенность. Согласно приведенным выше нашим данным в январе конкуренция между этими видами была не слишком велика и линейная плотность их была схожа – 10.6 и 9.5 встреч на 100 км маршрута судна, соответственно. В феврале конкуренция, очевидно, была более сильная, линейная плотность чернобрового альбатроса уменьшилась до 7.4 встреч на 100 км маршрута, у южного гигантского – сократилась почти втрое по сравнению с январем – до 3.85 встречи на 100 км (Kharitonov et al., 2021а). Экологические опечатки этих видов по месяцам дали следующую картину: в январе, когда конкуренция слабее, экологические отпечатки чернобрового альбатроса и южного гигантского буревестника были мало схожи между собой, а в феврале, при усилении конкуренции эти виды, казалось бы, должны были еще больше “разойтись”, однако они, наоборот, “сошлись” (рис. 4). Картины максимумов при этом практически совпали, а средние – значительно сблизились (табл. 2, рис. 4). Что здесь причина, а что – следствие, неясно, но задача для дальнейшего исследования интересная.
Рис. 4.
Изображение (наложение) экологических отпечатков чернобрового альбатроса (темные максимумы и средние) и южного гигантского буревестника (светлые максимумы и средние) в январе (слева) и феврале (справа). Обозначения осей как в табл. 2.
Из морских млекопитающих антарктический морской котик оказался самым экологически широким видом – шире, чем рассмотренные здесь два вида китов. В то же время, к февралю котики сузили свои экологические показатели по сравнению с январем (табл. 2). Видимо, это связано с разным характером использования акватории данным видом. В январе распределение этих животных было групповым, причем большие группы котиков отмечены у вершины Антарктического п-ова и соседних островов, возле Южных Шетландских и Южных Оркнейских о-вов, т.е. в январе котики были и далеко в море, и недалеко от земли. К февралю котики стали чаще встречаться далеко от земли, и их распределение в океане стало случайным (Kharitonov et al., 2021). Изменение в характере распределения и, соответственно, экологических отпечатках в эти месяцы весьма вероятно связано с циклом размножения этих животных: в январе они были еще недалеко от лежбищ, в феврале антарктические морские котики начинают мигрировать на север (Дикий, Пекло, 2012).
Северный и южный гигантский буревестники – близкие виды, которые имеют вполне сходные суммарные за два месяца экологические отпечатки. Правда, в основном это касается средних величин, экологический максимум у южного гигантского явно шире (табл. 2). Однако по отдельным месяцам эти виды могут разительно различаться. В январе северный гигантский буревестник вполне соответствует своему названию “северный”: по широтам он встречался севернее, но и западнее, чем южный гигантский (табл. 2), при этом предпочитая более высокие температуры воздуха, однако – то ли вытерпливая, то ли предпочитая более сильные ветер и порывы. В более теплый февраль северные гигантские буревестники распространились дальше на юг, в среднем даже несколько южнее южного гигантского буревестника. При этом, в отличие от южного, избегали крайне высоких температур, отчего средние температуры воздуха для мест встречи вида получились немного ниже, чем у южного. Южный оказался и более толерантен не только к размаху температур, но и к размаху атмосферного давления и влажности воздуха (табл. 2). Известно, что эти виды различаются по областям разлета и спектрам питания. Кроме планктона, оба вида питаются и падалью. Самцы южных гигантских буревестников даже хищничают, добывая живых птиц, для северных гигантских буревестников это нехарактерно (Cooper et al., 2001; Shirihai, 2008).
Хотя общие экологические пределы от января к февралю расширились (табл. 1), у многих птиц значения экологических параметров сузились. В основном это коснулось уменьшения максимумов. Данное явление отмечено у следующих видов: антарктического пингвина, качурки Вильсона, сероголового альбатроса, капского голубка, прионов и белогорлого буревестника. У трех последних сильно уменьшились максимумы при почти неизменных средних. Причина этих явлений пока не совсем ясна, требует дальнейшего изучения.
У странствующего альбатроса довольно широкие экологические пределы, вид широко распространен в Антарктике. Однако хорошо видно, что этот вид четко придерживается низких температур и, соответственно, более сильного действия фактора ветрового охлаждения. В области распространения он чем-то сходен с южным королевским альбатросом, тяготеет к западной части обследованной области Антарктики. Толерантен к волнению или предпочитает неспокойное море, видимо, поэтому большое количество встреч этого вида зафиксировано в проливе Дрейка. При этом отмечался при несколько меньшей влажности, чем “ледовые” виды.
Чернобрюхая качурка имеет интересные экологические отпечатки: почти по всем параметрам она встречалась в более узком спектре условий, чем близкий вид – качурка Вильсона. У чернобрюхой качурки меньшее значение максимумов и, что интересно, более высокие минимумы, чем у качурки Вильсона. Возможно, поэтому большинство средних значений параметров у чернобрюхой качурки выше, чем у качурки Вильсона. Если качурка Вильсона многочисленна, встречалась практически повсеместно, как в областях айсбергов, так и без них, то чернобрюхая качурка встречалась значительно реже, льдов этот вид явно избегает.
ЗАКЛЮЧЕНИЕ
Предложенный метод экологических отпечатков позволяет визуализировать трудно представляемые параметры среды. Это дает возможность точнее оценить экологические особенности видов или групп животных в разных местах и в разные сезоны, позволяет выявить и такие особенности, которые могут оставаться плохо выявляемыми другими методами, включая метод факторного анализа.
Некоторые виды морских млекопитающих и морских птиц имеют очень похожие экологические отпечатки, что говорит об одинаковом отношении к условиям среды у разных видов вне зависимости от их систематической принадлежности. Финвал, горбатый кит, снежный буревестник, пингвин Адели, антарктический буревестник и антарктический глупыш в январе имели специфический характер экологических отпечатков, указывающий на приуроченность этих видов к акваториям с айсбергами и битым льдом. В феврале связь китов со льдами ослабла, что стало видно по их экологическим отпечаткам, а у всех перечисленных выше видов птиц сохранилась. Экологические отпечатки хорошо демонстрируют ширину абиотической экологической ниши, которую занимают конкретные виды и группы. Они могут показать изменение областей распределения в конкретные периоды (южный королевский альбатрос), смену стратегии использования акватории (антарктический морской котик), а также другие, в том числе пока нераскрытые, экологические особенности.
Необходимо подчеркнуть, что данное сообщение является первым, где используется предлагаемый здесь же метод экологических отпечатков. Поэтому едва ли нам удалось извлечь всю информацию, которую могут дать экологические отпечатки. Дальнейшая разработка метода должна это позволить.
Список литературы
Дикий И.В., Пекло А.М., 2012. Тюлени Аргентинских островов (Антарктика) // Сборник трудов Зоологического музея. № 43. С. 104–116.
Коросов А.В., 1996. Экологические приложения компонентного анализа. Петрозаводск: Издательство Петрозаводского университета. 152 с.
Морозов Е.Г., Флинт М.В., Спиридонов В.А., Тараканов Р.Ю., 2019. Программа комплексных экспедиционных исследований экосистемы Атлантического сектора Южного океана (декабрь 2019–март 2020 г.) // Океанология. Т. 59. № 6. С. 1086–1088.
Морозов Е.Г., Спиридонов В.А., Молодцова Т.Н., Фрей Д.И., Демидова Т.А., Флинт М.В., 2020. Исследования экосистемы атлантического сектора Антарктики (79-й рейс научно-исследовательского судна “Академик Мстислав Келдыш”) // Океанология. Т. 60. № 5. С. 823–825.
Морозов Е.Г., Фрей Д.И., Полухин А.А., Кречик В.А., Артемьев В.А., Гавриков А.В., Касьян В.В., Сапожников Ф.В., Гордеева Н.В., Кобылянский С.Г., 2020а. Мезомасштабная изменчивость океана в северной части моря Уэддела // Океанология. Т. 60. № 5. С. 663–669.
Плохинский Н.А., 1978. Математические методы в биологии. М.: Издательство Московского университета. 265 с.
Покровская И.В., Похелон А., Гоммерштадт О.М., Вайсс И., 2018. Повторные регистрации чернобрового альбатроса Thalassarche melanophris в российских арктических водах // Русский орнитологический журнал. Т. 25. Вып. 1663. С. 4375–4378.
Харитонов С.П., 2021. Компьютерная программа “Spider graph”, исполняемый файл – spider_g.exe
Ainley D., Russell J., Jenouvrier S., Woehler E., Lyver P.O’B., Fraser W.R., Kooyman G.L., 2010. Antarctic penguin response to habitat change as Earth’s troposphere reaches 28C above preindustrial levels // Ecological Monographs. V. 80. № 1. P. 49–66.
Cooper J., Brooke M.L., Burger A.E., Crawford R.J.M., Hunter S., Williams T., 2001. Aspects of the breeding biology of the Northern Giant Petrel (Macronectes halli) and the Southern Giant Petrel (M. giganteus) at sub-Antarctic Marion Island // Int. J. Ornithol. V. 4. P. 53–68.
Delord K., Kato A., Tarroux A., Orgeret1 F., Cotté C., Ropert-Coudert Y., Cherell Y., Descamps S., 2020. Antarctic petrels ‘on the ice rocks’: wintering strategy of an Antarctic seabird. Royal Society Open Science. V. 7. Iss. 4. P. 1–8.
Friedlaender A.S., Halpin P.N., Qian S.S., Lawson G.L., Wiebe P.H., Thiele D., Read A.J., 2006. Whale distribution in relation to prey abundance and oceanographic processes in shelf waters of the Western Antarctic Peninsula // Marine ecology progress series. № 317. P. 297–310.
Joiris C.R., Dochy O., 2013. A major autumn feeding ground for fin whales, southern fulmars and grey-headed albatrosses around the South Shetland Islands, Antarctica // Polar Biology. V. 36. P. 1649–1658.
Kasamatsu F., Matsuoka K., Hakamada T., 2000. Interspecific relationships in density among the whale community in the Antarctic // Polar Biology. V. 23. № 7. P. 466–473.
Kharitonov S.P., Tretyakov A.V., Mischenko A.L., Konyukhov N.B., Artemyeva S.M., Pilipenko G.Yu., Mamayev M.S., Dmitriyev A.E., 2021. Spatial distribution, species composition, and number of marine mammals at Argentine Sea, Drake Passage, east of Antarctic Peninsula and the Powell Basin in January-March 2020 // Advances in Polar Ecology. Antarctic Peninsula Region of the Southern Ocean. Oceanography and Ecology. Morozov E.G., Flint M.V., Spiridonov V.A. (eds). P. 316–330.
Kharitonov S.P., Mischenko A.L., Konyukhov N.B., Dmitriyev A.E., Tretyakov A.V., Pilipenko G.Yu., Artemyeva S.M., Mamayev M.S., 2021a. Spatial distribution, species composition, and number of seabirds at Argentine Sea, Drake Passage, east of Antarctic Peninsula and the Powell Basin in January-March 2020 // Advances in Polar Ecology. Antarctic Peninsula Region of the Southern Ocean. Oceanography and Ecology. Morozov E.G., Flint M.V., Spiridonov V.A. (eds). P. 331–344.
Kharitonov S.P., Tretyakov A.V., Mischenko A.L., 2021b. Meat in the ocean: how much and who is to blame? // Advances in Polar Ecology. Antarctic Peninsula Region of the Southern Ocean. Oceanography and Ecology. Morozov E.G., Flint M.V., Spiridonov V.A. (eds). P. 345–355.
Lyver P.O’B., MacLeod C.J., Ballard G., Karl B.J., Barton K.J., Adams J., Ainley D.G., 2011. Intra-seasonal variation in foraging behavior among Ade’lie penguins (Pygocelis adeliae) breeding at Cape Hallett, Ross Sea, Antarctica // P.R. Wilson. Polar Biology. V. 34. P. 49–67.
Mondreti R., Davidar P., Ryan P.G., Thiebot J.B., Gremillet D., 2020. Seabird and cetacean occurrence in the Bay of Bengal associated with marine productivity and commercial fishing effort // Marine Ornithology. V. 48. P. 91–101.
Quillfeldt P., Phillips R.A., Marx M., Masello J., 2014. Colony attendance and at-sea distribution of thin-billed prions during the early breeding season // Journal of Avian Biology. V. 45. P. 315–324.
Santora J.A., Reiaa C.S., Cossio A.M., Veiti R.R., 2009. Interannual spatial variability of krill (Euphausia superba) influences seabird foraging behavior near Elephant Island, Antarctica // Fisheries Oceanography. V. 18. № 1. P. 20–35.
Serratosa J., Hyrenbach K.D., Miranda-Urbina D., Portflitt-Toro M., Luna N., Luna-Jorquera G., 2020. Environmental Drivers of Seabird At-Sea Distribution in the Eastern South Pacific Ocean: Assemblage Composition Across a Longitudinal Productivity Gradient // Frontiers in Marine Science. V. 6. I. 838. P. 1–13.
Shirihai H., 2008. The Complete Guide to Antarctic Wildlife. Birds and marine mammals of the Antarctic Continent and the Southern Ocean. Princeton and Oxford: Princeton University Press. 546 p.
Wakefield E.D., Phillips R.A., Trathan P.N., Arata J., Gales R., Huin N., Robertson G., Waugh S.M., Weimerskirch H., Matthiopoulos J., 2011. Habitat preference, accessibility, and competition limit the global distribution of breeding Black-browed Albatrosses // Ecological Monographs. V. 81. P. 141–167.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал