

Зоологический журнал, 2021, T. 100, № 12, стр. 1371-1387
Связь лейкоцитарной формулы с энергетическим состоянием, гематокритом и метаболитами крови на примере большой синицы (Parus major)
А. А. Волкова a, *, М. С. Березанцева b, К. И. Афонов c, А. В. Спиридонов b, Д. Р. Поликарпова b, Е. А. Жукова d, И. В. Демина a, А. Л. Цвей a
a Биологическая станция Рыбачий Зоологического института РАН
238535 Калининград, Россия
b Российский государственный педагогический университет имени А.И. Герцена
191186 Санкт-Петербург, Россия
c Санкт-Петербургский государственный университет ветеринарной медицины
196084 Санкт-Петербург, Россия
d Русский музей, филиал “Летний сад, Михайловский сад и зеленые территории музея”
191186 Санкт-Петербург, Россия
* E-mail: 10a.a.volkova@gmail.com
Поступила в редакцию 19.11.2020
После доработки 18.01.2021
Принята к публикации 09.02.2021
Аннотация
Состояние природных популяций животных может служить индикатором изменений мест обитания. Для оценки состояния популяций часто используются различные физиологические параметры. Эти параметры могут быть связаны между собой или могут быть независимы, позволяя организму гибко реагировать на изменения окружающей среды. На примере городской популяции большой синицы (Parus major) мы исследовали изменчивость лейкоцитарной формулы (общего количества лейкоцитов и процентного отношения разных типов лейкоцитарных клеток в крови) в течение осенней миграции и зимовки. Также мы проанализировали связь между параметрами лейкоцитарной формулы и индексами энергетического состояния, гематокритом и уровнем метаболитов в крови (глюкоза, β-гидроксибутират). Общее количество лейкоцитов и количество лимфоцитов были выше в осеннюю миграцию по сравнению с зимовкой, что свидетельствует о более активном состоянии иммунной системы больших синиц во время миграции. Повышенное количество лимфоцитов (специфический иммунитет) может объясняться высокой вероятностью встречи с уже знакомыми инфекциями при миграции на относительно короткие дистанции, характерные для данного вида. Отношение гетерофилов и лимфоцитов (отношение H : L, характеристика уровня долговременного стресса) в исследованной популяции синиц было выше в зимовку, чем в осеннюю миграцию, что может быть связано с низкими температурами и повышенной конкуренцией за кормовые ресурсы у зимующих птиц. Общее количество лейкоцитов было связано положительно с энергетическим состоянием и отрицательно – с уровнем глюкозы в крови, что позволяет предположить согласованную реакцию этих физиологических параметров на изменения условий среды. Таким образом, синицы с достаточным количеством энергетических запасов и низкой концентрацией глюкозы в крови (что характеризует индивидуальное благополучие) поддерживали иммунную систему в более активном состоянии. Уменьшение количества энергетических резервов, например, в связи с неблагоприятными кормовыми условиями, увеличением энергетических затрат на локомоторную активность и термогенез, приводило к увеличению концентрации глюкозы в крови и снижению числа лейкоцитов. Отношение H : L не было связано с другими физиологическими показателями, т.е. уровень долговременного стресса в исследованной популяции больших синиц не зависел от степени их энергетической обеспеченности и уровня метаболизма в текущих экологических условиях. Совокупность параметров лейкоцитарной формулы отражает низкий уровень физиологического стресса в исследованной популяции больших синиц.
В связи с глобальным изменением окружающей среды особую значимость приобретают методы оценки состояния природных популяций животных как индикатора изменений мест обитания (Hebert et al., 2000; Вершинин, 2008; Соколов, 2010; Колодезников, 2017; Cooke et al., 2020). При этом особое внимание уделяется синантропным видам птиц (Liker et al., 2008). В урбанизированных ландшафтах птицы подвержены воздействию различных факторов. Например, беспокойство людьми во время размножения вызывает нарушение кормовой активности птиц и снижение репродуктивного успеха (Kaliński et al., 2014). Воздействие токсинов приводит к уменьшению вероятности вылупления и снижению темпа роста у птенцов (Scheuhammer et al., 2007; Seewagen, 2018), а также может привести к гибели взрослых особей и потомства (Artacho et al., 2007). В то же время, урбанизированные территории могут представлять собой благоприятную среду обитания, например, за счет лучшего обеспечения птиц пищевыми ресурсами (Becker et al., 2019; Гашков, 2007; Evans, Gawlik, 2020). Оценка состояния популяций на основании прямых демографических параметров, таких как продолжительность жизни, репродуктивный успех или выживаемость, является трудоемким процессом. В качестве альтернативного метода могут использоваться параметры оценки физиологического состояния отдельных особей, дающие представление об условиях существования популяции (Willmer et al., 2000; Carey, 2005; Müller et al., 2011; Milenkaya et al., 2013; Cooke et al., 2020).
Одним из простых и доступных методов оценки физиологического состояния, как индивидуального, так и на уровне популяции, является анализ показателей крови, в частности лейкоцитарной формулы (Davis et al., 2008). Лейкоцитарная формула – это общее количество лейкоцитов и процентное соотношение разных типов лейкоцитарных клеток в крови (Campbell, Ellis, 2007). У птиц основными анализируемыми параметрами лейкоцитарной формулы являются общее количество лейкоцитов и соотношение самых многочисленных типов лейкоцитов – гетерофилов и лимфоцитов (отношение H : L) (Davis et al., 2008). На количество и процентное соотношение лейкоцитов в крови влияет концентрация глюкокортикоидов – гормонов, выделяющихся в ответ на различные стрессовые воздействия (Sapolsky et al., 2000; Romero, 2004).
При увеличении концентрации кортикостерона, главного глюкокортикоида у птиц (Harvey et al., 1984), происходит перераспределение гетерофилов и лимфоцитов в тканях, в результате чего возрастает отношение гетерофилов к лимфоцитам в крови (Davis et al., 2008). Считается, что изменения лейкоцитарной формулы под действием стрессовых факторов происходят значительно медленнее, чем изменение концентрации кортикостерона (Davis et al., 2008). Это позволяет использовать отношение H : L в качестве показателя долговременного стресса. Так, у видов с высоким отношением H : L сокращалась продолжительность жизни и снижалась годовая выживаемость на индивидуальном и популяционном уровнях (Minias et al., 2018). Повышение отношения H:L может быть связано с неблагоприятными погодными явлениями, например в сильные морозы у больших синиц были отмечены высокие значения отношения H : L (Krams et al., 2011). Повышенные отношения H : L были зафиксированы у птенцов пищухи (Certhia familiaris) и сероголовой гаечки (Poecile cinctus) на территориях с низкой обеспеченностью пищевыми ресурсами (Suorsa et al., 2004; Krams et al., 2010). Увеличение отношения H : L также может быть связано с присутствием эктопаразитов (Müller et al., 2011) и паразитов крови (Fokidis et al., 2008).
Общее количество лейкоцитов может свидетельствовать о состоянии иммунной системы (Campbell, 1995; Norris, Evans, 2000). Так, увеличение числа лейкоцитов в городской популяции серых юнко (Junco hyemalis) авторы связали с кормовыми условиями в городе, оптимальными для этого вида (Becker et al., 2019). Кроме того, общее количество лейкоцитов может увеличиваться в ответ на инфекции (Ots et al., 1998). У мигрирующих воробьиных было отмечено повышение общего количества лейкоцитов при заражении паразитами крови (Emmenegger et al., 2018).
Состояние иммунной системы в целом и лейкоцитарной формулы в частности меняется в течение года в зависимости от условий окружающей среды (температура, наличие инфекций и паразитов и т.д.) и количества доступных ресурсов (Raberg et al., 2000; Nava et al., 2001; Owen, Moore, 2008; Pap et al., 2010; Demina et al., 2019). Следовательно, при использовании лейкоцитарной формулы для оценки состояния популяции важно определить пределы ее вариаций в разные стадии годового цикла.
Для оценки состояния популяций используются и другие физиологические параметры. В первую очередь, это оценка жировых запасов и массы тела как показателей энергетического состояния (Labocha, Hayes, 2012). Эти параметры положительно связаны с выживаемостью и успехом размножения (Newton, 1993; Blums et al., 2005; Guy Morrison et al., 2007; Paquette et al., 2014).
Помимо этого, для оценки физиологического состояния используется гематокрит – относительный объем эритроцитов в общем объеме крови. Этот показатель отражает способность крови переносить кислород и связан с уровнем метаболизма (Kostelecka-Myrcha, Jaroszewicz, 1993; Saino et al., 1997; Bech, Klaassen, 1996). Высокие значения гематокрита положительно связаны с массой тела, отражают отсутствие инфекций и паразитов, а также хорошую обеспеченность пищевыми ресурсами (Averbeck, 1992; Potti, 2007; Minias et al., 2018). Более высокий гематокрит был отмечен у больших синиц (Parus major) в сельской местности по сравнению с городской популяцией, что могло быть связано с увеличением энергетических затрат на размножение из-за большего размера кладки. Снижение гематокрита у городских птиц могло быть вызвано также подавлением гемопоэза соединениями свинца из выхлопных газов (Ots et al., 1998).
Также для оценки физиологического состояния применяются показатели уровня метаболитов в крови, например концентрации глюкозы и β-гидроксибутирата. Глюкоза является основным и наиболее универсальным источником энергии для обеспечения метаболических процессов (Braun, Sweazea, 2008). У птиц концентрация глюкозы значительно выше, чем у млекопитающих (Braun, Sweazea, 2008; Lill, 2011). У птенцов лазоревки (Cyanistes caeruleus) и у взрослых зебровых амадин (Taeniopygia guttata) смертность положительно коррелировала с уровнем глюкозы (Kaliński et al., 2014; Montoya et al., 2018). Концентрация глюкозы увеличивается с уменьшением массы тела, снижением температуры воздуха и при повышенной физической нагрузке (Lill, 2011; Kaliński et al., 2014; Montoya et al., 2018).
β-гидроксибутират – это кетон, синтезируемый из свободных жирных кислот и замещающий глюкозу при отрицательном энергетическом балансе (Ramenofsky, 1990). Низкий уровень β-гидроксибутирата указывает на преобладание процессов анаболизма при увеличении массы тела и/или наличие энергетических резервов (Seaman et al., 2006). Низкая концентрация β-гидроксибутирата была обнаружена у птиц при восстановлении энергетических ресурсов во время миграционной остановки на территории с более благоприятными кормовыми условиями (Guglielmo et al., 2005). Уровень β-гидроксибутирата в крови увеличивается даже после непродолжительного периода голодания (Jenni-Eiermann, Jenni, 1997), но быстро снижается при восстановлении доступа к пище (Zajac et al., 2006).
Однако все эти параметры описывают разные компоненты физиологического состояния, и зачастую неизвестно, есть ли между ними взаимосвязь, или их можно использовать как независимые характеристики. Разные физиологические системы организма могут быть функционально связаны между собой, что уменьшает степень их пластичности в ответ на изменение экологических условий (Piersma, van Gils, 2011; Buehler et al., 2012). В то же время, отсутствие строгой функциональной связи может позволить организму более гибко реагировать на изменения окружающей среды, особенно во время энергетически затратных стадий годового цикла. Для выявления истинного характера связи между различными физиологическими параметрами необходимо оценить ее в течение нескольких стадий годового цикла, которые характеризуются различными условиями (Buehler et al., 2012).
Целью нашей работы было изучение связи лейкоцитарной формулы с энергетическими показателями физиологического состояния, гематокритом и метаболитами крови в городской популяции большой синицы во время осенней миграции и зимовки.
На основании анализа литературы мы предполагаем:
1) Общее количество лейкоцитов отражает состояние иммунной системы и положительно связано с энергетическим состоянием птиц. Т.е. при наличии достаточного количества ресурсов увеличивается потенциальная возможность организма противостоять инфекциям.
2) Отношение H : L, как показатель долговременного стресса, отрицательно связано с энергетическим состоянием птиц. Т.е. при недостаточной обеспеченности ресурсами уровень физиологического стресса возрастает.
3) Гематокрит как показатель, связанный с обеспеченностью ресурсами и уровнем метаболизма, положительно коррелирует с общим количеством лейкоцитов и отрицательно – с отношением H : L.
4) Поскольку концентрация глюкозы повышается при усилении долговременного стресса (с ростом энергетических затрат и снижением массы тела), этот показатель положительно связан с отношением H : L и отрицательно – с общим количеством лейкоцитов.
5) Общее количество лейкоцитов и отношение H : L не связаны с уровнем β-гидроксибутирата в крови. Концентрация β-гидроксибутирата может меняться несколько раз в течение суток, в то время как изменение лейкоцитарной формулы происходит гораздо медленнее.
В качестве объекта исследования была выбрана большая синица – синантропный вид, широко распространенный на территории Ленинградской обл. и превосходящий по численности другие виды синиц (Мальчевский, Пукинский, 1983; Гашков, 2007; Носков и др., 2015). Большая синица может быть отловлена в городской черте на всех стадиях годового цикла (Мальчевский, Пукинский, 1983; Гашков, 2007), что позволило собрать материал для исследования в достаточном объеме. Помимо этого, большая синица является модельным видом в экофизиологических исследованиях в России и Европе (Kerimov, Ivankina, 1997; Керимов, Иванкина, 1999; Ots et al., 2001; Pap et al., 2010; Хозина и др., 2015; Гаврилов, 2018), что позволяет сравнивать наши данные с результатами, полученными в других экологических условиях.
МАТЕРИАЛ И МЕТОДЫ
Отлов и кольцевание
Исследования проводили в 2018–2020 гг. на территории Летнего сада в г. Санкт-Петербурге. Летний сад – парковый ансамбль, памятник садово-паркового искусства, основанный в 1704 г. На территории площадью 11.7 га произрастают 26 видов деревьев и 24 вида кустарника. Древесные виды преимущественно представлены липами, кленами, а также ясенями и дубами с преобладанием старовозрастных деревьев (Черданцева и др., 2016). Администрацией Русского музея с 2013 г. организована ежегодная подкормка птиц в холодное время года (с октября-ноября по апрель). Широкий спектр укрытий и регулярная подкормка привлекают в Летний сад много видов птиц, в том числе большую синицу. В Летнем саду исследуемый вид встречается в течение всего года (Жукова и др., 2018).
Отловы синиц проводили с сентября по май, один раз в неделю, в первой половине дня. Синицы были отловлены на следующих стадиях годового цикла: линька (послебрачная линька у взрослых особей, постювенильная – у молодых), осенняя миграция, зимовка и весенняя миграция. Временны́е границы стадий определяли в соответствии с литературными данными (Смирнов, Носков, 1975; Bojarinova et al., 2002; Носков, Смирнов, 2010, 2020). Линька у большой синицы начинается в июле и длится до конца сентября, отдельные линяющие особи могут попадаться и позже (Носков, Смирнов, 1981, 2020). Осенняя миграция проходит со второй половины сентября и до конца октября, зимовка – с декабря по январь, весенняя миграция – вторая половина февраля–март. В ноябре и первой половине февраля одновременно могут быть отловлены птицы, находящиеся на разных стадиях годового цикла (осенняя миграция/зимовка и зимовка/весенняя миграция, соответственно). Поэтому для птиц, пойманных в эти промежутки времени, не указывали точную стадию годового цикла. Во время миграций синицы подвижны и могут преодолеть большие расстояния в поисках кормных мест, а во время зимовки – в основном оседлы. Осенью на северо-западе России встречаются птицы, совмещающие линьку и миграцию (Мальчевский, Пукинский, 1983; Носков, Смирнов, 2020), поэтому в нашем исследовании к мигрирующим мы относили только особей, завершивших линьку. Большие синицы могут быть как мигрирующими, так и оседлыми, при этом наибольшую степень оседлости проявляют взрослые самцы. Кольцевать синиц на территории Летнего сада начали в 2017 г., поэтому мы предположили, что большинство местных оседлых птиц должны быть уже окольцованы. За весь период исследования окольцованные птицы встречались в отловах в незначительном количестве (13 птиц из 82, т.е. 16%) и, в основном, в период зимовки. В течение миграционного периода было отловлено всего 3 (8% от числа синиц, пойманных за этот период) птицы, окольцованные в предыдущие сезоны. Поэтому все птицы, отловленные в период осенней миграции, были объединены нами в одну выборку. Применение вышеописанных критериев позволило классифицировать точные стадии годового цикла у большинства отловленных синиц.
Птиц начинали отлавливать обычно спустя час после восхода солнца и заканчивали около полудня. Отлов проводили с помощью автоматических ловушек – бойков, которые устанавливали поверх стационарной кормушки. Для привлечения птиц использовали прикормку (семена подсолнечника) и запись голоса большой синицы. За ловушкой велось постоянное наблюдение. После отлова у птицы быстро брали пробу крови, затем ее кольцевали и отпускали. При этом фиксировали время отлова птицы (с точностью до часа) и время от отлова до окончания взятия пробы (мин : с).
Каждую синицу подвергали прижизненным измерениям согласно методике, принятой в программе ESF (Bairlein et al., 1995). У птицы определяли:
– возраст (Виноградова и др., 1976; Svensson, 1992) (sad – молодые особи, от окончания постювенильной линьки до первой послебрачной линьки, и ad – взрослые, особи в дефинитивном оперении, начиная с первой послебрачной линьки);
– пол (Виноградова и др., 1976; Svensson, 1992);
– длину крыла с точностью до 0.5 мм (Виноградова и др., 1976);
– жирность по 9-балльной шкале (Kaiser, 1993; Bairlein et al., 1995);
– состояние грудного мускула по 4-балльной шкале (Bairlein et al., 1995);
– массу тела с точностью до 0.1 г.
Взятие крови, приготовление мазков и измерение гематологических показателей
Взятие крови производили из плечевой вены в гепаринизированный микрокапилляр. Объем собранной крови варьировал от 5 до 75 мкл. Сразу после взятия крови измеряли концентрацию глюкозы и β-гидроксибутирата с помощью портативного глюкометра FreeStyle Optium (Abbott diagnostics, USA). Этот глюкометр был валидирован и рекомендован для исследований на птицах (Breuner et al., 2013; Tomasek et al., 2019). Диапазон измеряемых концентраций для глюкозы составляет 1.1–27.8 ммоль/л, для β-гидроксибутирата – 0–8.0 ммоль/л, что охватывает диапазон концентраций, известных для птиц (Guglielmo et al., 2005; Braun, Sweazea, 2008). Измерение метаболитов крови проводили по инструкции использования глюкометра: тест-полоску помещали в прибор, наносили на нее образец крови, ожидали отображения информации о концентрации. Для измерения концентрации каждого метаболита требовалось по 4 мкл крови.
После измерения концентрации метаболитов на предметных стеклах изготовляли парные мазки крови. Мазки высушивали на воздухе, после чего фиксировали в 100% метаноле в течение 4 мин. Зафиксированные мазки окрашивали по Романовскому-Гимзе в течение часа (Болотников, Соловьев, 1980).
Оставшуюся в капилляре кровь центрифугировали в гематокритной центрифуге (CM-70, Elmi, Latvia) в течение 5 мин с последующим измерением гематокрита. Гематокрит (отношение объема эритроцитов к общему объему крови, выраженное в процентах) определяли с помощью гематокритной линейки.
Микроскопический анализ мазков
Микроскопический анализ мазков проводился на базе СПбГУ: 1) в учебной лаборатории кафедры зоологии позвоночных на микроскопе Leica DM2500 (×1000, масляная иммерсия), 2) на кафедре прикладной экологии на микроскопе с масляной иммерсионной линзой Leica DM750 P (×1000, масляная иммерсия). Для идентификации лейкоцитов использовали руководства по гематологии птиц (Campbell, 1995; Clark et al., 2009). Относительные пропорции каждого типа лейкоцитов оценивали при подсчете 100 лейкоцитов на один мазок. Также оценивали общее количество эритроцитов в каждом просмотренном поле зрения. Общее количество лейкоцитов каждого типа выражали относительно 10 000 эритроцитов. Большинство мазков проанализировано А.А. Волковой и небольшое количество – Д.Р. Поликарповой, которые в ходе исследования взаимно корректировали идентификацию лейкоцитов и подсчет числа эритроцитов.
Выборка
Всего в течение двух полевых сезонов (2018/ 2019 и 2019/2020) было собрано 110 проб крови. Наибольшее количество проб было собрано в две стадии годового цикла: в осеннюю миграцию (n = 37) и зимовку (n = 45). Соответственно, мазки, сделанные в эти стадии, были отобраны для микроскопического анализа. Суммарная выборка составила 82 мазка, из которых в сезоне 2018/2019 было сделано 46 мазков, в сезоне 2019/ 2020 – 36. Большинство отловленных птиц были первогодками (sad, n = 70), 11 птиц были старше одного года (ad), у одной птицы возраст не был определен. В выборке было поровну самцов и самок, по 41 особи соответственно. В каждом сезоне отлова выборка была равномерно распределена по стадиям годового цикла и полу. В течение сезона 2018/2019 осенью была собрана 21 проба крови (самки 13, самцы 8), зимой – 25 проб (самки 13, самцы 12). В течение сезона 2019/2020 было собрано 16 проб крови осенью (самки 7, самцы 9) и 20 проб зимой (самки 8, самцы 12). В связи с тем, что объема собранной крови не всегда хватало для одновременного измерения метаболитов и определения гематокрита, выборка для каждого параметра имеет определенный размер (табл. 1).
Таблица 1.
Размер выборки, лимиты и средние значения для параметров лейкоцитарной формулы, энергетического состояния и метаболитов крови у больших синиц, отловленных в Летнем саду г. Санкт-Петербурга во время осенней миграции и зимовки
| Стадия годового цикла | Показатель | Количество лейкоцитов | Отношение H : L | Индекс состояния | Гематокрит, % | Глюкоза, ммоль/л |
β-гидроксибутират, ммоль/л |
|---|---|---|---|---|---|---|---|
| Осенняя миграция | n | 37 | 37 | 37 | 28 | 7 | 7 |
| lim | 6.8–41.4 | 0.01–0.91 | 0.223–0.278 | 44–56 | 18.4–19.3 | 0.9–4.9 | |
| Среднее ± SD | 17.8 ± 8.5 | 0.23 ± 0.16 | 0.242 ± 0.012 | 49.3 ± 3.2 | 18.8 ± 0.3 | 2.8 ± 1.5 | |
| Зимовка | n | 42 | 42 | 42 | 41 | 31 | 22 |
| lim | 4.5–31.9 | 0.05–0.91 | 0.218–0.286 | 41–59 | 16.8–25.0 | 0.8–4.3 | |
| Среднее ± SD | 13.5 ± 5.9 | 0.31 ± 0.23 | 0.258 ± 0.014 | 50.2 ± 3.6 | 19.6 ± 1.6 | 2.0 ± 1.1 |
Статистический анализ
Статистический анализ проводили в среде R 3.5.0 (R Core Team, 2018). Форму распределения и наличие выбросов проверяли на гистограммах и одномерных диаграммах рассеяния (графические функции hist и dotchart базового пакета R). Мазки с количеством лейкоцитов 71.4 на 10 000 эритроцитов и отношением гетерофилов к лимфоцитам 1.49 и 1.53 были исключены из анализа, т.к. эти значения отстояли от соответствующих средних как минимум на 3.8 стандартных отклонения, являясь выбросами (табл. 1). Таким образом, итоговая выборка для статистического анализа составила 79 мазков. Согласно рекомендации Zuur et al. (2010) был проведен предварительный анализ данных с целью проверки формы зависимостей между переменными и влияния взаимодействия независимых факторов на зависимую переменную. Это позволило исключить из основного анализа заранее незначимые переменные и их взаимодействия. В тексте в качестве средних тенденций приведены арифметические средние и величина стандартного отклонения (среднее ± SD).
Для анализа зависимости между общим количеством лейкоцитов и параметрами физиологического состояния были использованы Пуассоновские смешанные линейные модели (Poisson linear mixed models, Korner-Nievergelt et al., 2015). Количество лейкоцитов (размах 100–105) выступало в качестве зависимой переменной. Поскольку нас интересовало количество лейкоцитов, выраженное относительно 10 000 эритроцитов, сумма количества эритроцитов во всех полях зрения была учтена в модели при помощи функции оффсет (offset = lg10(количество эритроцитов/10 000), Korner-Nievergelt et al., 2015). Отношение гетерофилов к лимфоцитам было проанализировано с использованием нормальных смешанных моделей (Linear mixed effects models, Korner-Nievergelt et al., 2015). В качестве зависимой переменной выступали lg10 трансформированные значения отношения H : L.
Для анализа зависимости между параметрами лейкоцитарной формулы (общим количеством лейкоцитов и отношением H : L) и компонентами физиологического состояния (индекс состояния, балл жирности, гематокрит, глюкоза, ß-гидроксибутират) были построены отдельные модели, где параметры лейкоцитарной формулы выступали в качестве зависимых переменных и один компонент физиологического состояния – в качестве независимой переменной (ковариаты). Пример моделей для индекса состояния приведен ниже. Индекс состояния (ИС, масса тела, разделенная на длину крыла) представляет собой индекс массы тела, учитывающий структурный размер птицы, и отражает, таким образом, энергетическое состояние (Labocha, Hayes, 2012; Titov, Chernetsov, 1999).
Помимо одного компонента физиологического состояния, в каждой модели учитывали влияние стадии годового цикла (СГЦ, осенняя миграция и зимовка) и пола птиц (пол, самцы и самки), которые входили в модели в качестве независимых категориальных факторов. Предварительный анализ не выявил различий между взрослыми и первогодками, поэтому возраст не был включен в окончательные модели. Чтобы учесть возможную динамику лейкоцитарной формулы в зависимости от времени отлова в течение суток (время1, lim 0.94–5.37, среднее 2.4 ± 0.9 ч относительно восхода солнца) и времени от поимки до момента окончания взятия пробы крови (время2, lim 1.07–7.0, среднее 2.2 ± 1.1 мин), эти факторы были включены в модели в качестве независимых переменных (ковариат). На основе результатов предварительного анализа время отлова в течение суток (время1) учитывали только в моделях для отношения H : L. Все ковариаты были z-трансфомированы перед анализом, что упрощает сходимость моделей и позволяет сравнивать коэффициенты между собой (Schielzeth, 2010). Z-трансформация представляет собой разницу между каждым значением и средним по выборке, разделенную на стандартное отклонение по выборке (Schielzeth, 2010). Предварительный анализ данных показал, что наклоны зависимостей между параметрами лейкоцитарной формулы и компонентами физиологического состояния не различаются у особей разного пола и в разные стадии годового цикла, поэтому в модели включены только главные факторы, без учета взаимодействий.
Отлов птиц проводили один раз в неделю. Поскольку в одну дату могли быть сделаны мазки от 1 до 7 птиц, для учета возможного влияния даты отлова, дата была включена в модели в качестве случайного (random) фактора. Предварительный анализ также показал, что отношение H : L, но не количество лейкоцитов, различается между сезонами. В связи с этим сезон отлова (2018/2019 и 2019/2020) был включен в качестве случайного фактора в модели, исследующие отношение H : L. Чтобы учесть возможные различия в подсчете количества эритроцитов разными людьми, ID исследователя был включен в качестве случайного фактора в модели для общего количества лейкоцитов. При помощи функции dispersion_glmer из пакета blmeco (Korner-Nievergelt et al., 2015) было определено, что все модели для общего количества лейкоцитов характеризовались значительной величиной необъясненной дисперсии (overdispersion). Согласно рекомендации Korner-Nievergelt et al. (2015), чтобы получить устойчивые коэффициенты, в случайную часть моделей был добавлен специальный фактор – ORLF (observation level random factor). Этот фактор представляет собой порядковый номер каждого наблюдения в выборке, т.е. если в выборке n наблюдений, то уровнями этого фактора будут числа от 1 до n (Korner-Nievergelt et al., 2015).
Ниже, в качестве примера, приведены модели, оценивающие влияние индекса состояния (ИС) на общее количество лейкоцитов (а) и отношение H : L (б). СГЦ – стадия годового цикла; пол – пол птиц; время1 – время отлова относительно восхода солнца, время2 – время от поимки до момента окончания взятия пробы крови; (1|сезон) – случайный фактор, учитывающий влияние полевого сезона, в котором была отловлена птица; (1|дата) – случайный фактор, учитывающий влияние конкретной даты отлова; (1|ID) – случайный фактор, учитывающий влияние исследователя на подсчет эритроцитов; (1|ORLF) – случайный фактор, корректирующий величину необъясненной дисперсии в моделях для общего количества лейкоцитов, offset – функция, позволяющая выразить количество лейкоцитов относительно 10 000 эритроцитов (см. выше). Индекс z означает, что ковариаты были z‑трансформированы (см. выше)
(а) общее количество лейкоцитов ~ ИС.z + пол + + СГЦ + время2.z + (1|дата) + (1|ORLF) + (1|ID), offset = lg(количество эритроцитов/10 000).
(б) lg10(отношение H:L) ~ ИС.z + пол + СГЦ + + время1.z + время2.z + (1|сезон) + (1| дата).
Модели строили с использованием пакета lme4 и функций glmer (для общего количества лейкоцитов) и lmer (для отношения H : L, Bates et al., 2015). Достоверность коэффициентов модели оценивали с использованием пакета lmerTest (Kuznetsova et al., 2017). Достоверность влияния категориальных факторов (стадии годового цикла и пола птиц), а также эффекты ковариат (времени отлова относительно восхода солнца и времени от момента отлова до окончания взятия пробы крови) проверяли в моделях, включающих индекс состояния (показателя энергетического состояния). Эти модели использовали потому, что масса тела была измерена у всех птиц, у которых были взяты пробы крови и сделаны мазки, в отличие от гематокрита и метаболитов, выборка которых была неравномерно распределена по стадиям годового цикла. При анализе связи между параметрами лейкоцитарной формулы и энергетическим состоянием мы приводим результаты только для индекса состояния. Несмотря на достоверную корреляцию между баллом жирности и индексом состояния (корреляция Спирмена, r = 0.55, p < < 0.0001, n = 79), оценка балла жирности более субъективна, и действительное содержание жира даже при одном балле жирности может сильно варьировать (Åkesson et al., 1992; Zimin, 2003).
Проверку адекватности моделей проводили на основе графического анализа стандартных графиков (Zuur et al., 2009; Korner-Nievergelt et al., 2015), который не выявил серьезных отклонений от основных допущений линейных моделей.
РЕЗУЛЬТАТЫ
Общее количество лейкоцитов
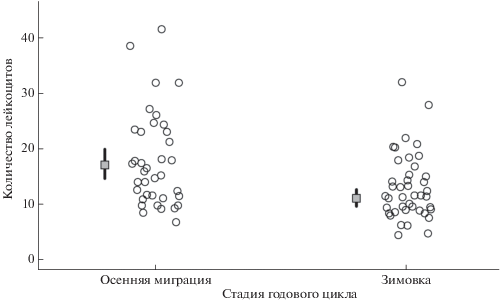
Общее количество лейкоцитов варьировало от 4.5 до 41.4 клеток относительно 10 000 эритроцитов и в среднем составляло 15.3 ± 7.5 клеток. В осеннюю миграцию количество лейкоцитов (17.8 ± 8.5 клеток относительно 10 000 эритроцитов) было достоверно больше, чем в зимовку (13.0 ± 5.9; табл. 2, рис. 1). Самцы и самки имели одинаковое количество лейкоцитов (табл. 2). Время от момента отлова до окончания сбора проб крови не оказывало влияния на количество лейкоцитов (табл. 2), т.е. количество лейкоцитов не изменялось вплоть до 7 мин после отлова. Анализ вариации, связанной со случайными факторами, показывает, что дата отлова объясняет часть вариации общего количества лейкоцитов (табл. 2), т.е. мазки, сделанные в одну дату, по количеству лейкоцитов более сходны между собой по сравнению с мазками, сделанными в другие, даже соседние, даты. Это с большой вероятностью связано с тем, что состояние иммунной системы зависит от конкретных экологических условий и меняется довольно синхронно у разных особей. Вариация, связанная с ID исследователя, анализировавшего мазки, была минимальна (табл. 2), что указывает на адекватность дифференциации клеток и подсчета количества эритроцитов разными исследователями.
Таблица 2.
Результаты анализа источников вариации лейкоцитарной формулы (количества лейкоцитов (WBC) и отношения H : L) у больших синиц, отловленных в Летнем саду во время осенней миграции и зимовки в сезоны 2018/2019 и 2019/2020
| Показатель | WBC | Отношение H : L | ||||||
|---|---|---|---|---|---|---|---|---|
| β | SE | z-value | p | β | SE | t-value | p | |
| Фиксированные факторы | ||||||||
| Интерсепта | 2.92 | 0.10 | 29.7 | <0.0001 | –0.80 | 0.10 | –8.5 | <0.0001 |
| ИС | 0.15 | 0.06 | 2.4 | 0.02 | 0.01 | 0.05 | 0.2 | 0.82 |
| СГЦ (зимовка) | –0.46 | 0.11 | –4.1 | <0.0001 | 0.24 | 0.10 | 2.4 | 0.02 |
| Пол (самцы) | –0.13 | 0.10 | –1.2 | 0.21 | –0.04 | 0.08 | –0.4 | 0.66 |
| Время1 | – | – | – | – | –0.03 | 0.04 | –0.8 | 0.45 |
| Время2 | 0.08 | 0.05 | 1.8 | 0.08 | 0.10 | 0.05 | 2.0 | 0.05 |
| Случайные факторы (SD) | ||||||||
| Сезон | – | 0.064 | ||||||
| Дата | 0.0001 | <0.0001 | ||||||
| ORLF | 0.391 | – | ||||||
| ID | 0 | – | ||||||
| Residual | – | 0.324 | ||||||
Примечания. Для общего количества лейкоцитов представлены результаты Пуассоновской смешанной модели. Для отношения гетерофилов к лимфоцитам – результаты нормальной смешанной модели. Приведены результаты моделей, где главной ковариатой был индекс состояния (ИС), т.е. для выборки, куда входили все проанализированные мазки. Достоверные эффекты выделены жирным шрифтом. Для категориальных факторов в скобках приведен сравниваемый уровень этого фактора. Результаты моделей, анализирующих влияние гематокрита и метаболитов крови, приведены в тексте.
Рис. 1.
Общее количество лейкоцитов у больших синиц, отловленных во время осенней миграции и зимовки в Летнем саду г. Санкт-Петербурга. Показаны предсказанные Пуассоновской смешанной моделью средние значения (серые квадраты) с доверительным 95% интервалом (вертикальные линии), а также исходные данные (отдельные точки).
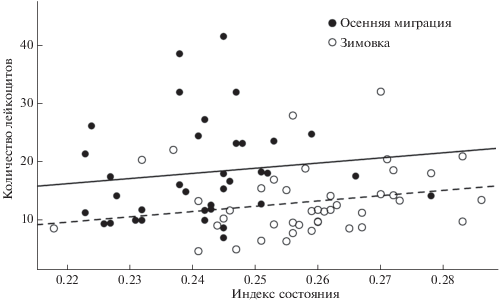
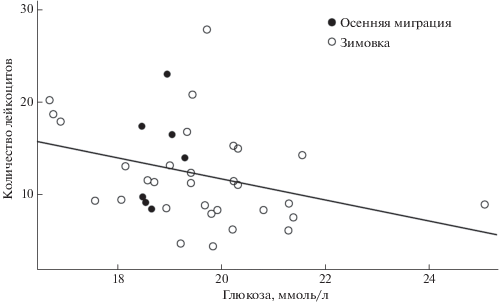
Общее количество лейкоцитов было положительно связано с индексом состояния (табл. 2, рис. 2), т.е. количество иммунных клеток увеличивалось у особей, находившихся в лучшем энергетическом состоянии. Количество лейкоцитов не было связано с гематокритом (Пуассоновские смешанные модели: β = 0.06 ± 0.05 (SE), z = 1.1, p = 0.25) и β-гидроксибутиратом (β = –0.02 ± 0.06 (SE), z = –0.4, p = 0.70). Концентрация глюкозы в плазме крови была отрицательно связана с количеством лейкоцитов (β = –0.15 ± 0.07 (SE), z = –2.3, p = 0.02, рис. 3), т.е. количество иммунных клеток было низким у синиц с высокой концентрацией глюкозы.
Рис. 2.
Зависимость между общим количеством лейкоцитов и индексом состояния (масса тела/длина крыла) у больших синиц, отловленных во время осенней миграции и зимовки в Летнем саду г. Санкт-Петербурга. Сплошная линия – линейный тренд для осенней миграции. Пунктирная линия – линейный тренд для зимовки.
Отношение гетерофилов к лимфоцитам
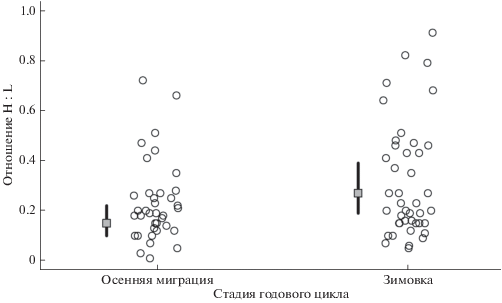
Отношение H : L больших синиц, отловленных в Летнем саду, варьировало от 0.01 до 0.91 и в среднем составило 0.27 ± 0.20. Между отношением H : L и общим количеством лейкоцитов взаимосвязи не наблюдалось (корреляция Спирмена: r = –0.11, p = 0.35, n = 79). В осеннюю миграцию отношение H : L было достоверно ниже (0.23 ± 0.16), чем во время зимовки (0.31 ± 0.23, табл. 2, рис. 4). Индекс состояния, пол и время отлова относительно восхода солнца не влияли на отношение H : L (табл. 2). В то же время, наблюдался положительный тренд отношения H : L со временем окончания взятия пробы крови (табл. 2), т.е. отношение H : L имело тенденцию к увеличению уже в течение семи минут после отлова. Анализ вариации, связанной со случайными факторами, показывает, что конкретная дата отлова не объясняет вариацию отношения H : L у синиц, т.е. у разных особей этот параметр меняется асинхронно (табл. 2). Сезон отлова объяснял часть вариации отношения H : L (табл. 2), что было обнаружено при предварительном анализе данных (см. Методы). В то же время, значительная часть вариации (residual) осталась необъясненной (табл. 2). Отношение гетерофилов к лимфоцитам не было связано с гематокритом (смешанные линейные модели: β = –0.03 ± 0.04 (SE), t = –0.7, p = 0.51) и метаболитами крови (глюкоза: ß = = 0.06 ± 0.05 (SE), t = 1.3, p = 0.22; β-гидроксибутират: β = –0.07 ± 0.06 (SE), t = –1.1, p = 0.28 соответственно).
Рис. 4.
Отношение гетерофилов к лимфоцитам у больших синиц, отловленных во время осенней миграции и зимовки в Летнем саду г. Санкт-Петербурга. Показаны предсказанные Нормальной смешанной моделью средние значения (серые квадраты) с доверительным 95% интервалом (вертикальные линии), а также исходные данные (отдельные точки).
ОБСУЖДЕНИЕ
Состояние иммунной системы меняется на протяжении годового цикла, поэтому для корректного использования лейкоцитарной формулы в качестве параметра оценки состояния популяции важно определить пределы ее вариаций на разных стадиях. Мы оценивали изменчивость лейкоцитарной формулы в зависимости от пола больших синиц, стадии годового цикла и сезона отлова, а также изучали связь параметров лейкоцитарной формулы птиц с другими показателями их физиологического состояния.
Различий в параметрах лейкоцитарной формулы у синиц разного пола не было выявлено, что согласуется с данными ряда исследователей (Norte et al., 2009; Pap et al., 2010). Однако Ots et al. (1998) и Krams et al. (2011) отмечали повышенное отношение H : L у самок большой синицы в сравнении с самцами во время размножения и зимовки. Krams et al. (2011) предположили, что обнаруженные половые различия в отношении H : L определяются у синиц разным социальным статусом самок и самцов: в условиях экстремально низких температур самцы испытывают меньший уровень стресса из-за большей доступности для них ресурсов. Возможно, отсутствие половых различий в параметрах лейкоцитарной формулы у синиц в исследованной нами популяции было связано с большей доступностью и предсказуемостью ресурсов в условиях постоянной подкормки в парке, а также с достаточно мягкими температурными условиями зимовки.
Общее количество лейкоцитов у больших синиц в исследованной популяции было значительно ниже, чем у синиц во время размножения в Эстонии (Ots et al., 1998), но при этом выше, чем отмечалось у синиц в Португалии в течение всего года (Norte et al., 2009). Данный параметр лейкоцитарной формулы обладает широкой индивидуальной и видовой изменчивостью (Jain, 1986; Buehler et al., 2010), что усложняет сравнение абсолютных значений между разными популяциями.
Изменение лейкоцитарной формулы между периодами осенней миграции и зимовки и различия между сезонами отлова
Во время осенней миграции птицы сталкиваются с повышением энергетических затрат в связи с увеличением локомоторной активности (Sapir et al., 2011). Для стадии миграции характерны непредсказуемость кормовых ресурсов (Bibby, Green, 1980; Moore et al., 1995), а также усиление конкуренции за них (Rappole, Warner, 1976; Moore, Yong, 1991). Кроме того, в период миграции увеличивается риск заражения новыми для организма патогенами (Buehler et al., 2008). Большие синицы во время осенней миграции перемещаются на относительно короткие дистанции. Лишь небольшая часть особей (около 2% от всей численности вида) мигрирует на сотни и тысячи километров (Носков, Смирнов, 2020).
Во время зимовки стаи синиц постоянно держатся на одной и той же территории при условии достаточного количества пищевых ресурсов или перемещаются на небольшие расстояния (не больше нескольких километров, реже – десятков километров) (Носков, Смирнов, 2020). На зимовку синицы часто перемещаются из лесных массивов в населенные пункты (Мальчевский, Пукинский, 1983; Гашков, 2007), где увеличивается их плотность и возрастает конкуренция за пищевые ресурсы (Krams et al., 2011). Низкие температуры зимой приводят к росту энергетических затрат на термогенез (Dawson, Marsh, 1989; Swanson, 2010).
Общее количество лейкоцитов в осеннюю миграцию у синиц из исследованной популяции было выше по сравнению с зимовкой. Повышенное количество лейкоцитов может свидетельствовать о текущем инфекционном процессе или отражать активность иммунной системы, т.е. потенциальные возможности организма предотвратить инфицирование (Ots et al., 1998; Norris, Evans, 2000; Campbell, Ellis, 2007). Прямых (истощение, конъюнктивит, присутствие кровепаразитов в мазках и т.д.) и косвенных (повышенное количество эозинофилов в крови) признаков инфекции у птиц в период осенней миграции мы не обнаружили. Кроме того, у синиц с высокими индексами энергетического состояния было отмечено увеличение количества лейкоцитов (см. ниже), что также косвенно свидетельствует об отсутствии у них инфекции. Мы предполагаем, что повышенное количество лейкоцитов в осеннюю миграцию связано с активным состоянием иммунной системы птиц, направленным на предотвращение заражения во время активных перемещений (Møller, Erritzoe, 1998; Hasselquist, 2007).
В связи с сезонной динамикой ресурсов, а также с различными функциями иммунных клеток и затратами на их образование, в течение годового цикла может меняться активность отдельных ветвей иммунной системы (Buehler et al., 2008; Hegemann et al., 2012; Demina et al., 2019). Согласно Buehler et al. (2008), для сезонных миграций характерно развитие энергетически затратного, неспецифического иммунитета (гетерофилы), поскольку он оптимален в условиях повышенной вероятности инфицирования новыми для организма патогенами. В немиграционные периоды преобладает специфический иммунитет (лимфоциты), оптимальный при нахождении в пределах ограниченной территории, где высока вероятность повторного столкновения с уже известными патогенами (Buehler et al., 2008). У синиц из исследованной популяции изменение общего количества лейкоцитов определялось, в основном, изменением количества лимфоцитов: в осеннюю миграцию было отмечено повышенное количество лимфоцитов в сравнении с зимовкой, что противоречит данным других исследователей (Buehler et al., 2008; Pap et al., 2010). Возможно, это связано с особенностями перемещения данной популяции на короткие дистанции во время миграции и высокой вероятностью повторной встречи с инфекциями.
Во время зимовки синицы ведут относительно оседлый образ жизни и встречаются с меньшим количеством новых патогенов, чем в остальные стадии годового цикла, по крайней мере, в умеренном поясе. В условиях ограниченных ресурсов и возрастания энергетических затрат на термогенез активность иммунной системы может снижаться, что, возможно, и проявляется в обнаруженном нами уменьшении общего количества лейкоцитов у синиц во время зимовки. При этом количество гетерофилов сохраняется примерно на том же уровне, что и во время миграции, обеспечивая потенциальный ответ на неспецифические инфекции.
Отношение H : L в исследованной популяции синиц в зимовку было выше, чем в осеннюю миграцию, что может свидетельствовать о более высоком уровне стресса (Davis et al., 2008). По данным Pap et al. (2010), соотношение гетерофилов и лимфоцитов у синиц, напротив, снижалось во время зимовки, что исследователи связывали с пониженной активностью иммунной системы из-за низких температур и отсутствия паразитов в зимний период. Однако условия зимовки исследованной нами популяции синиц отличались от данных Pap et al. (2010): птицы зимовали в условиях города и при более низких температурах, что, возможно, и повлияло на разницу в результатах. Низкие температуры, ограниченные пищевые ресурсы и обострение конкуренции из-за локального увеличения плотности популяции – все это может характеризовать стадию зимовки как более сложную в отношении выживания для исследованной популяции больших синиц. Есть данные, согласно которым, отношение H : L может возрастать в период миграции в связи с повышением физических нагрузок при полете (Owen, Moore, 2006, 2008). Однако следует принимать во внимание сравнительно небольшую дистанцию и относительную легкость (отсутствие экологических барьеров и т.д.) миграционных перемещений большой синицы.
Соотношение гетерофилов и лимфоцитов в сезон отлова 2019/2020 гг. было ниже, чем в сезон 2018/2019 гг. Погодные условия зим 2018/2019 и 2019/2020 годов на территории Санкт-Петербурга сильно различались: зима 2019/2020 гг. была значительно теплее. Мы предполагаем, что пониженный уровень стресса, испытываемого синицами в более теплый сезон отлова, может быть связан со снижением энергозатрат на термогенез. При этом вариации общего количества лейкоцитов между сезонами отлова не были обнаружены. Это свидетельствует о том, что уровень активности иммунной системы на разных стадиях годового цикла мало подвержен сезонным колебаниям, по крайней мере, в том диапазоне экологических условий, в котором были собраны наши данные.
Значения отношения H : L у больших синиц в нашем исследовании находились на сравнительно невысоком уровне по сравнению с данными других авторов (Norte et al., 2009; Pap et al., 2010). Это показывает относительно низкий уровень долговременного стресса у больших синиц, отловленных в Летнем саду Санкт-Петербурга во время осенней миграции и зимовки. При этом мы не исключаем, что обнаруженные вариации отношения H : L между стадиями годового цикла и сезонами отлова не были напрямую связаны с развитием физиологического стресса, а отражают эндогенную сезонную динамику состояния иммунной системы (Demina et al., 2019). В то же время наши результаты показывают, что отношение H : L увеличивается уже в течение 7 мин после отлова. Принято считать, что изменения этого параметра происходят не раньше чем через час после наступления стрессового воздействия (отлова) или значительно позже (Davis, 2005; Cırule et al., 2012). Низкая скорость изменений отношения H : L позволяет использовать этот параметр как показатель долговременного стресса (Davis et al., 2008). Нам трудно объяснить быстрое увеличение отношения H : L. Мы не исключаем, что это может быть артефактом конкретной выборки, который не влиял на остальные результаты, т.к. время от момента отлова до окончания взятия пробы крови было учтено в статистических моделях.
Таким образом, для оценки изменений физиологического состояния особей и популяций большой синицы необходимо учитывать стадию годового цикла, а также влияние условий сезона отлова. При этом отношение H : L и общее количество лейкоцитов отражают различные аспекты индивидуального физиологического состояния, что объясняет противоположные тренды изменений этих показателей в разные стадии годового цикла.
Связь лейкоцитарной формулы с энергетическим состоянием, гематокритом и метаболитами крови
Физиологическое состояние зависит от действия факторов среды и от потребностей организма в каждую стадию годового цикла. Различные параметры физиологического состояния могут меняться в течение годового цикла как синхронно, так и асинхронно, независимо друг от друга. В условиях повышенных энергетических затрат, а также ограниченности ресурсов, в первую очередь, поддерживается активность физиологических систем, которые на данной стадии имеют для организма первостепенное значение. В результате характер связи физиологических параметров на разных стадиях может быть разным. Существует предположение, согласно которому об истинной функциональной связи между физиологическими параметрами можно говорить лишь в случае, если эта связь выявляется и имеет одинаковое направление (положительная/отрицательная) на разных стадиях годового цикла (Buehler et al., 2012). При этом отсутствие строгой функциональной связи между разными физиологическими системами дает возможность организму более гибко реагировать на воздействие факторов среды, особенно в условиях повышенных энергетических затрат и ограниченных ресурсов.
Согласно нашим данным, изменение общего количества лейкоцитов у большой синицы положительно связано с индексом состояния в обе стадии годового цикла, т.е. особи, находившиеся в лучшем энергетическом состоянии, в среднем, имели больше лейкоцитов. Это свидетельствует о том, что при наличии достаточного количества ресурсов организм может направлять их на поддержание иммунной системы в более активном состоянии. В условиях ограниченных ресурсов и повышенных энергозатрат (в связи с увеличением локомоторной активности во время миграции или расходов энергии на терморегуляцию во время зимовки) организм может столкнуться с необходимостью распределения ресурсов между энергозатратной активностью и поддержанием иммунной системы. В этом случае тощие птицы будут вынуждены снижать активность иммунной системы ради решения более важных для выживания задач (Svensson et al., 1998).
Связь между отношением H : L и индексом энергетического состояния не была выявлена в обе стадии годового цикла. На основании этого мы предполагаем, что величина долговременного стресса в исследованной популяции больших синиц не связана со степенью энергетической обеспеченности птиц в текущих экологических условиях. Другие исследователи (Suorsa et al., 2004; Krams et al., 2010) отмечали более высокий уровень стресса у тощих птиц. Однако, как отмечалось выше, в исследуемой популяции синиц уровень стресса был относительно низким, возможно из-за предсказуемости условий городской среды и регулярной подкормки птиц на территории Летнего сада.
Мы не обнаружили связь параметров лейкоцитарной формулы с гематокритом во время осенней миграции и зимовки. Работы, в которых исследовалась бы связь этих параметров, отсутствуют. Наши результаты позволяют предположить, что лейкоцитарная формула и гематокрит меняются независимо, поскольку отражают различные аспекты физиологического состояния птиц.
Нами была выявлена отрицательная связь общего количества лейкоцитов и концентрации глюкозы в крови в обе стадии годового цикла, т.е. у птиц с высоким уровнем глюкозы было снижено количество лейкоцитов. Нам не известны работы, в которых исследована связь общего количества лейкоцитов с концентрацией глюкозы в крови. Повышенная концентрация глюкозы в крови отражает неблагоприятные кормовые условия и/или увеличение энергетических затрат. При этом активность иммунной системы в условиях дефицита ресурсов может снижаться, что находит отражение в отрицательном характере зависимости между этими физиологическими параметрами.
Связь между концентрацией глюкозы и отношением H : L не была обнаружена. По данным других исследователей, отношение гетерофилов и лимфоцитов может положительно коррелировать с уровнем глюкозы в крови (Montoya et al., 2018; Lill, 2011; Kaliński et al., 2014). Так, у птенцов лазоревки из парковой зоны был отмечен повышенный уровень глюкозы и увеличенное отношение H : L по сравнению с лесной популяцией (Kaliński et al., 2014). Исследователи предположили, что это может быть следствием увеличения беспокойства птиц в гнездовой период и относительно низкого количества пищевых ресурсов в городе (Kaliński et al., 2014). Возможно, отсутствие взаимосвязи глюкозы и отношения H : L можно объяснить общим низким уровнем стресса в исследованной нами популяции.
Связь между параметрами лейкоцитарной формулы и уровнем β-гидроксибутирата в крови не была обнаружена в обе стадии годового цикла. Отсутствие связи подтверждает наше предположение о разной скорости изменения этих физиологических показателей. Концентрация β-гидроксибутирата в крови меняется при питании и голодании достаточно быстро, до нескольких раз в течение суток (Zajac et al., 2006; Johnson, 2007). В то же время считается, что лейкоцитарная формула меняется значительно медленнее (Davis, 2005; Davis et al., 2008).
Таким образом, нами была выявлена связь общего количества лейкоцитов с индексами энергетического состояния и с уровнем глюкозы в крови, позволяющая предположить согласованную реакцию со стороны этих физиологических систем на изменения условий среды. Однако для уточнения характера этой связи необходимо в дальнейшем провести ее оценку на остальных стадиях годового цикла, различающихся энергетическими потребностями и экологическими условиями. Отсутствие связи параметров лейкоцитарной формулы с гематокритом и уровнем β-гидроксибутирата в крови свидетельствует о том, что они меняются под действием разных факторов окружающей среды и/или с разной скоростью. Отношение H : L не было связано ни с одним из исследованных физиологических показателей, т.е. уровень долговременного стресса в исследованной популяции больших синиц не был связан со степенью энергетической обеспеченности птиц и уровнем их метаболизма в текущих экологических условиях.
Использование лейкоцитарной формулы наряду с другими параметрами физиологического состояния отдельных особей позволяет получить комплексное представление об условиях существования популяции. Низкие значения соотношения гетерофилов и лимфоцитов и положительная связь общего количества лейкоцитов с энергетическим состоянием птиц показывают хорошее состояние популяции модельного вида в Летнем саду. Полученные данные могут быть использованы для сравнения с другими популяциями этого вида. При использовании лейкоцитарной формулы для оценки физиологического состояния популяций большой синицы необходимо учитывать стадию годового цикла, а также условия сезона отлова.
Список литературы
Болотников И.А., Соловьев Ю.В., 1980. Гематология птиц. Л.: Наука, Ленинградское отделение. 116 с.
Вершинин В.Л., 2008. Амфибии как индикаторы состояния урбанизированных экосистем // Материалы ІІІ междунар. Науч.-практ. конф. “Урбоэкосистемы. Проблемы и перспективы развития”. С. 21–25.
Виноградова Н.В., Дольник В.Р., Ефремов В.Д., Паевский В.А., 1976. Определитель пола и возраста воробьиных птиц фауны СССР // М.: Наука. 189 с.
Гаврилов В.В., 2018. Экологическая энергетика синиц, зимующих в западном Подмосковье // Орнитология: история, традиции, проблемы и перспективы. Материалы Всероссийской конференции, посвященной 120-летию со дня рождения профессора Г.П. Дементьева. М.: Товарищество научных изданий КМК. 421 с.
Гашков С.И., 2007. Биология большой синицы (Parus major L.) южной тайги Западной Сибири. Автореф. дис. … канд. биол. наук. Томск: Томский госуниверситет. 24 с.
Жукова Е.А., Березанцева М.С., Шалакитская О.В., 2018. Орнитофауна садов Русского музея // Первый Всероссийский орнитологический конгресс. Тезисы докладов. Тверь. С. 111–112.
Керимов А.Б., Иванкина Е.В., 1999. Социодемографические факторы изменчивости уровня базального метаболизма в зимующих группировках большой синицы (Parus major) // Зоологический журнал. Т. 78. № 3. С. 358–371.
Колодезников В.Е., 2017. Использование мелких млекопитающих в качестве индикаторов состояния среды в Якутии // Вестник Северо-Восточного федерального университета им. МК Аммосова. Т. 59. № 3. С. 13–24.
Мальчевский А.С., Пукинский Ю.Б., 1983. Птицы Ленинградской области и сопредельных территорий: история, биология, охрана // Л.: Изд-во ЛГУ. Т. 1. 480 с.
Носков Г.А., Рымкевич Т.А., Гагинская А.Р., 2015. Орнитофауна Санкт-Петербурга: история изучения, современный состав, задачи охраны // Биосфера. Т. 7. № 1. С 80–95.
Носков Г.А., Смирнов О.П., 1981. Территориальное поведение и миграции большой синицы (Parus major L.) // Экология птиц Приладожья. Л.: Изд-во ЛГУ. С. 100–130.
Носков Г.А., Смирнов О.П., 2010. Некоторые особенности территориального поведения и миграции большой синицы Parus major и их адаптивное значение // Русский орнитологический журнал. Т. 19. № 575. С. 995–998.
Носков Г.А., Смирнов О.П., 2020. Большая синица Parus major // Миграции птиц Северо-Запада России. Воробьиные. Отв. ред. Носков Г.А., Рымкевич Т.А., Гагинская А.Р. СПб.: “Реноме”. С. 346–355.
Смирнов О.П., Носков Г.А., 1975. Структура популяции большой синицы в Ленинградской области // Экология. № 6. С. 79–83.
Соколов Л.В., 2010. Климат в жизни растений и животных. СПб.: Изд-во ТЭССА. 344 с.
Хозина В.М., Якименко Н.Н., Пономарев В.А., Клетикова Л.В., 2015. Гемато-биохимический профиль модельного вида птиц на примере большой синицы (Parus major L.), обитающей в урбанизированной среде // Современные проблемы науки и образования. № 3. С. 557–563.
Черданцева О.А., Лукмазова Е.А., Дмитриева А.Г., Дубровина Т.А., 2016. Сохранение старовозрастных деревьев и мониторинг их состояния в Летнем и Михайловском садах Санкт-Петербурга // Вестник Российского университета дружбы народов, Серия агрономия и животноводство. № 4. С. 45–50.
Åkesson S., Karlsson L., Pettersson J., Walinder G., 1992. Body composition and migration strategies: a comparison between Robins (Erithacus rubecula) from two stopover sites in Sweden // Die Vogelwarte. V. 36. P. 188–195.
Artacho P., Soto-Gamboa M., Verdugo C., Nespolo R.F., 2007. Using haematological parameters to infer the health and nutritional status of an endangered black-necked swan population // Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology. V. 147. № 4. P. 1060–1066.
Averbeck C., 1992. Haematology and blood chemistry of healthy and clinically abnormal great black-backed gulls (Larus marinus) and herring gulls (Larus argentatus) // Avian Pathology. V. 21. № 2. P. 215–223.
Bairlein F., Jenni L., Kaiser A., Karlsson L., van Noordwijk A. et al., 1995. Manual of field methods // European-African songbird migration network. Institut für Vogelforschung, Wilhelmshaven. 32 p.
Bates D., Martin M., Bolker B., Walker S., 2015. Fitting linear mixed-effects models using lme4 // Journal of Statistical Software. V. 67. P. 1–48.
Bech C., Klaassen M., 1996. Blood hemoglobin content and metabolic performance of Arctic Tern chicks Sterna paradisaea // Journal of Avian Biology. P. 112–117.
Becker D.J., Schultz E.M., Atwell J.W., Ketterson E.D., 2019. Urban residency and leukocyte profiles in a traditionally migratory songbird // Animal Migration. V. 6. № 1. P. 49–59.
Bibby C.J., Green R.E., 1980. Foraging behaviour of migrant pied flycatchers, Ficedula hypoleuca, on temporary territories // Journal of Animal Ecology. V. 49. P. 507–521.
Blums P., Nichols J.D., Hines J.E., Lindberg M.S., Mednis A., 2005. Individual quality, survival variation and patterns of phenotypic selection on body condition and timing of nesting in birds // Oecologia. V. 143. № 3. P. 365–376.
Bojarinova J.G., Rymkevich T.A., Smirnov O.P., 2002. Timing of autumn migration of early and late-hatched Great Tits Parus major in NW Russia // Ardea. V. 90. № 3. P. 401–409.
Braun E.J., Sweazea K.L., 2008. Glucose regulation in birds // Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology. V. 151. № 1. P. 1–9.
Breuner C.W., Delehanty B., Boonstra R., 2013. Evaluating stress in natural populations of vertebrates: total CORT is not good enough // Functional Ecology. V. 27. № 1. P. 24–36.
Buehler D.M., Piersma T., Matson K., Tieleman B.I., 2008. Seasonal redistribution of immune function in a migrant shorebird: annual-cycle effects override adjustments to thermal regime // The American Naturalist. V. 172. № 6. P. 783–796.
Buehler D.M., Tieleman B.I., Piersma T., 2010. Indices of immune function are lower in red knots (Calidris canutus) recovering protein than in those storing fat during stopover in Delaware Bay // The Auk. V. 127. P. 394–401.
Buehler D.M., Vézina F., Goymann W., Schwabl I., Versteegh M., Tieleman B.I., Piersma T., 2012. Independence among physiological traits suggests flexibility in the face of ecological demands on phenotypes // Journal of Evolutionary biology. V. 25. № 8. P. 1600–1613.
Campbell T.W., 1995. Avian hematology and cytology // Iowa State University Press. Ed. 2. 104 p.
Campbell T.W., Ellis C.K., 2007. Hematology of birds // Avian and Exotic animal hematology and cytology. P. 3–50.
Carey C., 2005. How physiological methods and concepts can be useful in conservation biology // Integrative and Comparative Biology. V. 45. № 1. P. 4–11.
Cīrule D., Krama T., Vrublevska J., Rantala M. J., Krams I., 2012. A rapid effect of handling on counts of white blood cells in a wintering passerine bird: a more practical measure of stress? // Journal of Ornithology. V. 153. № 1. P. 161–166.
Clark P., Boardman W., Raidal S., 2009 Atlas of clinical avian hematology // John Wiley & Sons. 184 p.
Cooke S.J., Madliger C.L., Cramp R.L., Beardall J., Burness G. et al., 2020. Reframing conservation physiology to be more inclusive, integrative, relevant and forward-looking: reflections and a horizon scan // Conservation physiology. V. 8. № 1. P. 1–19.
Davis A.K., 2005. Effect of handling time and repeated sampling on avian white blood cell counts // Journal of Field Ornithology. V. 76. № 4. P. 334–338.
Davis A.K., Maney D.L., Maerz J.C., 2008. The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists // Functional Ecology. V. 22. № 5. P. 760–772.
Dawson W.R., Marsh R.L., 1989. Metabolic acclimatization to cold and season in birds // Physiology of cold adaptation in birds. P. 83–94.
Demina I., Tsvey A., Babushkina O., Bojarinova J., 2019. Time-keeping programme can explain seasonal dynamics of leukocyte profile in a migrant bird // Journal of Avian Biology. V. 50. № 7. P. 1–13.
Emmenegger T., Bauer S., Hahn S., Müller S.B., Spina F., Jenni L., 2018. Blood parasites prevalence of migrating passerines increases over the spring passage period // Journal of Zoology. V. 306. № 1. P. 23–27.
Evans B.A., Gawlik D.E., 2020. Urban food subsidies reduce natural food limitations and reproductive costs for a wetland bird // Scientific Reports. V. 10. № 1. P. 1–12.
Fokidis B.H., Greiner E.C., Deviche P., 2008. Interspecific variation in avian blood parasites and haematology associated with urbanization in a desert habitat // Journal of Avian Biology. V. 39. № 3. P. 300–310.
Guglielmo C.G., Cerasale D.J., Eldermire C., 2005. A field validation of plasma metabolite profiling to assess refueling performance of migratory birds // Physiological and Biochemical Zoology. V. 78. № 1. P. 116–125.
Guy Morrison R.I., Davidson N.C., Wilson J.R., 2007. Survival of the fattest: body stores on migration and survival in red knots Calidris canutus islandica // Journal of Avian Biology. V. 38. № 4. P. 479–487.
Harvey S., Phillips J.G., Rees A., Hall T.R., 1984. Stress and adrenal function // Journal of Experimental Zoology. V. 232. № 3. P. 633–645.
Hasselquist D., 2007. Comparative immunoecology in birds: hypotheses and tests // Journal of Ornithology. V. 148. № 2. P. 571–582.
Hebert C.E., Norstrom R.J., Weseloh D.C., 2000. A quarter century of environmental surveillance: the Canadian Wildlife Service’s Great Lakes herring gull monitoring program // Environmental Reviews. V. 7. № 4. P. 147–166.
Hegemann A., Matson K.D., Both C., Tieleman B.I., 2012. Immune function in a free-living bird varies over the annual cycle, but seasonal patterns differ between years // Oecologia. V. 170. № 3. P. 605–618.
Jain N.C., 1986. Schalm’s veterinary hematology // Philadelphia, Pa: Lea & Febiger. Ed. 4. 1221 p.
Jenni-Eiermann S., Jenni L., 1997. Diurnal variation of metabolic responses to short-term fasting in passerine birds during the postbreeding, molting and migratory period // The Condor. V. 99. № 1. P. 113–122.
Johnson M.D., 2007. Measuring habitat quality: a review // The Condor. V. 109. № 3. P. 489–504.
Kaiser A., 1993. A new multi-category classification of subcutaneous fat deposits of songbirds // Journal of Field Ornithology. V. 64. P. 246–255.
Kaliński A., Bańbura M., Glądalski M., Markowski M., Skwarska J. et al., 2014. Landscape patterns of variation in blood glucose concentration of nestling blue tits (Cyanistes caeruleus) // Landscape ecology. V. 29. № 9. P. 1521–1530.
Kerimov A.B., Ivankina E.V., 1997. The Energetics and Social Status of a Specimen: Relationship between the Basal Metabolism Rate and the Social Position of a Specimen in Wintering Groups of Great Tit (Parus major, Passeriformes, Aves) // In doklady biological sciences section C/C of doklady- Akademiia Nauk SSSR. V. 357. P. 565–568.
Korner-Nievergelt F., Roth T., Von Felten S., Guélat J., Almasi B., Korner-Nievergelt P., 2015. Bayesian data analysis in ecology using linear models with R, BUGS, and Stan // Academic Press. 328 p.
Kostelecka-Myrcha A., Jaroszewicz M., 1993. The changes in the values of red blood indices during the nestling development of the House Martin Delichon urbica // Acta Ornithologica. V. 28. № 1. P. 39–46.
Krams I., Cīrule D., Krama T., Hukkanen M., Rytkönen S. et al., 2010. Effects of forest management on haematological parameters, blood parasites, and reproductive success of the Siberian tit (Poecile cinctus) in northern Finland // Annales Zoologici Fennici. – Finnish Zoological and Botanical Publishing Board. V. 47. № 5. P. 335–346.
Krams I., Cīrule D., Krama T., Vrublevska J., 2011. Extremely low ambient temperature affects haematological parameters and body condition in wintering great tits (Parus major) // Journal of Ornithology. V. 152. № 4. P. 889–895.
Kuznetsova A., Brockhoff P.B., Christensen R.H., 2017. lmerTest package: tests in linear mixed effects models // Journal of Statistical software. V. 82. № 13. P. 1–26.
Labocha M.K., Hayes J.P., 2012. Morphometric indices of body condition in birds: a review // Journal of Ornithology. V. 153. № 1. P. 1–22.
Liker A., Papp Z., Bókony V., Lendvai A.Z., 2008. Lean birds in the city: body size and condition of house sparrows along the urbanization gradient // Journal of Animal Ecology. V. 77. № 4. P. 789–795.
Lill A., 2011. Sources of variation in blood glucose concentrations of free-living birds // Avian Biology Research. V. 4. № 2. P. 78–86.
Milenkaya O., Weinstein N., Legge S., Walters J.R., 2013. Variation in body condition indices of crimson finches by sex, breeding stage, age, time of day, and year // Conservation physiology. V. 1. № 1. P. 1–14.
Minias P., Włodarczyk R., Meissner W., 2018. Leukocyte profiles are associated with longevity and survival, but not migratory effort: a comparative analysis of shorebirds // Functional Ecology. V. 32. № 2. P. 369–378.
Møller A.P., Erritzøe J., 1998. Host immune defence and migration in birds // Evolutionary Ecology. V. 12. № 8. P. 945–953.
Montoya B., Briga M., Jimeno B., Moonen S., Verhulst S., 2018. Baseline glucose level is an individual trait that is negatively associated with lifespan and increases due to adverse environmental conditions during development and adulthood // Journal of Comparative Physiology B. V. 188. № 3. P. 517–526.
Moore F.R., Gauthreaux S.A. Jr., Kerlinger P., Simons T.R., 1995. Habitat requirements during migration: important link in conservation // Ecology and management of neotropical migratory birds. P. 121–144.
Moore F.R., Yong W., 1991. Evidence of food-based competition among passerine migrants during stopover // Behavioral Ecology and Sociobiology. V. 28. № 2. P. 85–90.
Müller C., Jenni-Eiermann S., Jenni L., 2011. Heterophils/Lymphocytes-ratio and circulating corticosterone do not indicate the same stress imposed on Eurasian kestrel nestlings // Functional Ecology. V. 25. № 3. P. 566–576.
Nava M.P., Veiga J.P., Puerta M., 2001. White blood cell counts in house sparrows (Passer domesticus) before and after moult and after testosterone treatment // Canadian Journal of Zoology. V. 79. № 1. P. 145–148.
Newton S.F., 1993. Body condition of a small passerine bird: ultrasonic assessment and significance in overwinter survival // Journal of Zoology. V. 229. № 4. P. 561–580.
Norris K., Evans M. R., 2000. Ecological immunology: life history trade-offs and immune defense in birds // Behavioral Ecology. V. 11. № 1. P. 19-26.
Norte A.C., Ramos J.A., Sousa J.P., Sheldon B.C., 2009. Variation of adult Great Tit Parus major body condition and blood parameters in relation to sex, age, year and season // Journal of Ornithology. V. 150. № 3. P. 651–660.
Ots I., Kerimov A.B., Ivankina E.V., Ilyina T.A., Hõrak P., 2001. Immune challenge affects basal metabolic activity in wintering Great tits // Proceedings of the Royal Society B: Biological Sciences. V. 268. № 1472. P. 1175–1181.
Ots I., Murumägi A., Horak P., 1998. Haematological health state indices of reproducing great tits: methodology and sources of natural variation // Functional Ecology. V. 12. № 4. P. 700–707.
Owen J.C., Moore F.R., 2006. Seasonal differences in immunological condition of three species of thrushes // The Condor. V. 108. № 2. P. 389–398.
Owen J.C., Moore F.R., 2008. Relationship between energetic condition and indicators of immune function in thrushes during spring migration // Canadian Journal of Zoology. V. 86. № 7. P. 638–647.
Pap P.L., Vágási C.I., Tökölyi J., Czirják G.Á., Barta Z., 2010. Variation in haematological indices and immune function during the annual cycle in the Great Tit Parus major // Ardea. V. 98. № 1. P. 105–112.
Paquette S.R., Pelletier F., Garant D., Bélisle M., 2014. Severe recent decrease of adult body mass in a declining insectivorous bird population // Proceedings of the Royal Society B: Biological Sciences. V. 281. № 1786. P. 1–9.
Piersma T., van Gils J.A., 2011. The flexible phenotype: a body-centred integration of ecology, physiology, and behaviour. Oxford: Oxford University Press. 248 p.
Potti J., 2007. Variation in the hematocrit of a passerine bird across life stages is mainly of environmental origin // Journal of Avian Biology. V. 38. № 6. P. 726–730.
Råberg L., Nilsson J.Å., Ilmonen P., Stjernman M., Hasselquist D., 2000. The cost of an immune response: vaccination reduces parental effort // Ecology Letters. V. 3. № 5. P. 382–386.
Ramenofsky M., 1990. Fat storage and fat metabolism in relation to migration // Bird migration. P. 214–231.
Rappole J.H., Warner D.W., 1976. Relationships between behavior, physiology and weather in avian transients at a migration stopover site // Oecologia. V. 26. № 3. P. 193–212.
Romero L.M., 2004. Physiological stress in ecology: lessons from biomedical research // Trends in Ecology and Evolution. V. 19. № 5. P. 249–255.
Saino N., Cuervo J.J., Krivacek M., de Lope F., Møller A.P., 1997. Experimental manipulation of tail ornament size affects the hematocrit of male barn swallows (Hirundo rustica) // Oecologia. V. 110. № 2. P. 186–190.
Sapir N., Butler P.J., Hedenström A., Wikelski M., 2011. Energy gain and use during animal migration // Animal Migration: A Synthesis. P. 52–67.
Sapolsky R.M., Romero L.M., Munck A.U., 2000. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions // Endocrine reviews. V. 21. № 1. P. 55–89.
Scheuhammer A.M., Meyer M.W., Sandheinrich M.B., Murray M.W., 2007. Effects of environmental methylmercury on the health of wild birds, mammals, and fish // AMBIO: A Journal of the Human Environment. V. 36. № 1. P. 12–19.
Schielzeth H., 2010. Simple means to improve the interpretability of regression coefficients: Interpretation of regression coefficients // Methods in Ecology and Evolution V. 1. P. 103–113.
Seaman D.A., Guglielmo C.G., Elner R.W., Williams T.D., 2006. Landscape-scale physiology: site difference in refueling rates indicated by plasma metabolite analysis in free-living migratory sandpipers // The Auk. V. 123. № 2. P. 563–574.
Seewagen C.L., 2018. The threat of global mercury pollution to bird migration: potential mechanisms and current evidence // Ecotoxicology. P. 1–14.
Suorsa P., Helle H., Koivunen V., Huhta E., Nikula A., Hakkarainen H., 2004. Effects of forest patch size on physiological stress and immunocompetence in an area-sensitive passerine, the Eurasian treecreeper (Certhia familiaris): an experiment // Proceedings of the Royal Society of London. Series B: Biological Sciences. V. 271. № 1537. P. 435–440.
Svensson E., Raberg L., Koch C., Hasselquist D., 1998. Energetic stress, immunosuppression and the costs of an antibody response // Functional Ecology. V. 12. № 6. P. 912–919.
Svensson L., 1992. Identification guide to European passerines. // Stockholm: Lars Svensson. 368 p.
Swanson D.L., 2010. Seasonal metabolic variation in birds: functional and mechanistic correlates // Current Ornithology. V. 17. P. 75–129.
Titov N., Chernetsov N., 1999. How body mass should be compared to structural size when calculating condition index // Avian Ecology and Behaviour. V. 3. P. 111–113.
Tomasek O., Bobek L., Kralova T., Adamkova M., Albrecht T., 2019. Fuel for the pace of life: Baseline blood glucose concentration co-evolves with life-history traits in songbirds // Functional Ecology. V. 33. № 2. P. 239–249.
Willmer P., Johnston I., Stone G., 2000. Environmental Physiology of Animals. Malden, MA: Blackwell. 644 p.
Zajac R., Cerasale D., Guglielmo C., 2006. The rapid response of plasma metabolites to changes in feeding rate in a small passerine Wilsonia pusilla // Journal of Avian Biology. V. 37. № 4. P. 405–408.
Zimin V.B., 2003. Body mass variability in juvenile Robins Erithacus rubecula in the Ladoga area // Avian Ecology and Behaviour. V. 10. P. 1–31.
Zuur A., Ieno E.N., Walker N., Saveliev A.A., Smith G.M., 2009. Mixed effects models and extensions in ecology with R. New York, NY: Springer Science and Business Media. 574 p.
Zuur A.F., Ieno E.N., Elphick C.S., 2010. A protocol for data exploration to avoid common statistical problems // Methods in ecology and evolution. V. 1. № 1. P. 3–14.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал