

Зоологический журнал, 2021, T. 100, № 3, стр. 288-298
Гнездование и биология Eustenancistrocerus amadanensis (Hymenoptera, Vespidae, Eumeninae)
А. В. Фатерыга a, *, М. В. Ончуров b, **
a Карадагская научная станция имени Т.И. Вяземского – природный заповедник РАН –
филиал Института биологии южных морей имени А.О. Ковалевского РАН
298188 Курортное, Феодосия, Россия
b Таврическая академия Крымского федерального университета имени В.И. Вернадского
295007 Симферополь, Россия
* E-mail: fater_84@list.ru
** E-mail: larinioides@mail.ru
Поступила в редакцию 17.12.2019
После доработки 09.01.2020
Принята к публикации 15.02.2020
Аннотация
Изучено строение гнезд Eustenancistrocerus amadanensis в Крыму. Самки гнездились в земле на горизонтальных участках или пологих склонах с разреженной растительностью. Гранулометрический состав почвы, в которой осы устраивали свои гнезда, варьировал в широких пределах: от супесей до тяжелых суглинков, близких по свойствам к глинам. Исследовано 11 гнезд, пять из которых были брошены самками до откладки яйца. Остальные гнезда содержали 1–4 ячейки. Они имели глубину 5.0–7.5 см; диаметр гнездового хода составлял около 3 мм. Ячейки не отличались от гнездового хода по диаметру и имели длину 9–13 мм. Многоячейковые гнезда имели линейно-ветвистую структуру. Гнезда также содержали пустые боковые ходы, функция которых до конца не ясна. У входа в гнездо выстраивались наклонные трубки из земли высотой 5–10 мм; они были сплошными в основании и ажурными в дистальной части. Для строительства входной трубки самки извлекали из гнездового хода небольшие комочки земли. После того, как трубка была готова, самки продолжали углублять гнездовой ход, вынося из него более крупные комочки земли и выбрасывая их. Самки разбирали трубки при запечатывании гнезд. Осы использовали для размягчения земли воду, которую добывали из влажной земли у ручьев или на берегу моря, а также из луж, садясь прямо на их поверхность и удерживаясь за счет сил поверхностного натяжения. Самки охотились на гусениц двух видов молей-чехлоносок. Темп гнездования был очень низким (1 ячейка за 2–8 дней). Питание имаго нектаром цветков было отмечено на 10 видах растений из восьми семейств. Вид дает два поколения в году; зимовка происходит на стадии предкуколки. В гнездах найден кокон неидентифицированной осы-блестянки. Репродуктивный успех составил 81.1%. Обнаружена агрегация спящих самцов. Обсуждается биология ос рода Eustenancistrocerus и близких родов.
Eustenancistrocerus Blüthgen 1938 (Hymenoptera, Vespidae, Eumeninae) – небольшой род одиночных складчатокрылых ос, насчитывающий 19 видов, распространенных в аридных регионах Палеарктической и Афротропической зоогеографических областей. В составе рода выделяют три подрода. Номинативный подрод Eustenancistrocerus s. str. включает девять палеарктических видов, три афротропических вида и один вид, общий для Палеарктической и Афротропической зоогеографических областей. Подрод Hemistenancistrocerus Blüthgen 1938 включает один вид общий для Палеарктической и Афротропической зоогеографических областей. Подрод Parastenancistrocerus Blüthgen 1938 включает пять палеарктических видов (Carpenter et al., 2010; Gusenleitner, 2013; Fateryga et al., 2019). В фауне России известно два вида: Eustenancistrocerus (Eustenancistrocerus) jerichoensis (von Schulthess 1928), обнаруженный только в Дагестане, и Eustenancistrocerus (Parastenancistrocerus) amadanensis (de Saussure 1855) [= E. transitorius (Morawitz 1867)], распространенный на юге европейской части страны, Северном Кавказе и в Крыму (Antropov, Fateryga, 2017; Fateryga, Mokrousov, 2019). Общее распространение E. amadanensis включает Южную и Восточную Европу, о-в Кипр, Северную Африку, Аравийский п-ов, Малую Азию, Закавказье, Среднюю Азию, а также Казахстан, Иран и Пакистан (Antropov, Fateryga, 2017). Ареал E. jerichoensis включает Кавказ, Малую Азию, Аравийский полуостров, а также Иран (Fateryga, Mokrousov, 2019).
Биология представителей рода Eustenancistrocerus изучена слабо. В литературе имеются сведения о гнездовании типового вида рода – Eustenancistrocerus (Eustenancistrocerus) blanchardianus de Saussure 1855 – в Алжире (Ferton, 1914). Самки этого вида самостоятельно выгрызают гнездовой ход в земле, используя для ее размягчения отрыгиваемую воду. Для пополнения запасов воды в зобике они посещают ближайшие открытые источники, садясь при этом прямо на поверхность воды и удерживаясь за счет сил поверхностного натяжения. У входа в гнездо самка выстраивает входную трубку из земли длиной около 15 мм. Когда трубка готова, самка углубляет гнездовой ход, вынося из него комочки земли и выбрасывая их в 1–2 м от гнезда. Внутри гнездового хода E. blanchardianus обнаружены ячейки, разделенные поперечными земляными перегородками. В качестве провизии зарегистрированы гусеницы молей-чехлоносок (Lepidoptera, Coleophoridae) (Ferton, 1914). Второй вид, о биологии которого сообщалось ранее, – E. amadanensis. Имеется информация о его гнездовании в Иране, согласно которой E. amadanensis гнездится в замочных скважинах, залепляя их глиной (Bingham, 1897). Однако в Крыму было обнаружено гнездо E. amadanensis в земле (Fateryga et al., 2013). При этом самка самостоятельно выгрызала гнездовой ход и выстраивала входную трубку около 5 мм длиной. К сожалению, гнездо было не окончено и не содержало ячеек. В качестве объектов охоты E. amadanensis зарегистрированы гусеницы молей-чехлоносок Perygra maritimella (Newman 1873) (Fateryga et al., 2013); питание имаго зарегистрировано на цветках шести видов растений (Kostylev, 1928; Фатерыга, 2010; Fateryga et al., 2013). Этим сведения о биологии ос рода Eustenancistrocerus исчерпываются.
Цель настоящей работы – изучить гнездовую активность, строение гнезд, трофические связи и другие особенности биологии E. amadanensis на материале, собранном после публикации предыдущей заметки (Fateryga et al., 2013).
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в Крыму в 2011–2019 гг. Гнездование E. amadanensis изучали в Лисьей бухте (городской округ Феодосия) в 2011 и 2014 гг., на берегу соленого оз. Бараколь (также городской округ Феодосия) в 2014 и 2016 гг. и в балке Кипчак на Тарханкутском п-ове (Черноморский р-н) в 2019 г. Всего было найдено 11 гнезд (табл. 1). Дополнительные наблюдения за особями этого вида (прежде всего, во время питания имаго на цветках) были также проведены в некоторых других районах полуострова с 2011 по 2017 гг.
Таблица 1.
Характеристика участков гнездования Eustenancistrocerus amadanensis и число обнаруженных гнезд
| № | Описание | Координаты | Год | Число гнезд | |
|---|---|---|---|---|---|
| широта | долгота | ||||
| 1 | Лисья бухта, терраса у подножия бедленда | 44°54′02″ с.ш. | 35°09′28″ в.д. | 2011 | 1 |
| 2 | Лисья бухта, терраса у подножия бедленда | 44°53′58″ с.ш. | 35°09′29″ в.д. | 2014 | 1 |
| 3 | Лисья бухта, приморский склон | 44°54′04″ с.ш. | 35°10′00″ в.д. | 2014 | 3 |
| 4 | Лисья бухта, приморская терраса | 44°54′02″ с.ш. | 35°09′57″ в.д. | 2014 | 2 |
| 5 | Берег оз. Бараколь, солончак | 44°59′52″ с.ш. | 35°14′40″ в.д. | 2014 | 1 |
| 6 | Берег оз. Бараколь, солончак | 45°00′00″ с.ш. | 35°14′40″ в.д. | 2016 | 1 |
| 7 | Балка Кипчак, грунтовая дорога на склоне | 45°28′47″ с.ш. | 32°35′46″ в.д. | 2019 | 2 |
Гнезда были обнаружены, главным образом, случайно. За гнездящимися самками устанавливали наблюдение. Гнездовую активность ос регистрировали визуально и с помощью фотокамер Canon PowerShot A570 IS и Canon PowerShot SX160 IS. Гнезда раскапывали после того как самки их запечатывали либо после того как убеждались, что самки их бросили. Раскопку гнезд проводили с помощью небольшой лопаты и ножа, стараясь, чтобы разрез почвы был параллелен гнездовому ходу. Для этого вначале на расстоянии 20–30 см от входа в гнездо выкапывали ямку в форме полумесяца глубиной 15–20 см. Затем последовательно срезали землю слой за слоем по направлению к гнезду. В ряде случаев гнезда были извлечены из субстрата вместе с комом земли с помощью большой лопаты, а затем вскрыты в лаборатории. Однако иногда наличие крупных камней и обилие корней вблизи гнезд не позволяли это сделать. Вскрытые гнезда фотографировали и зарисовывали на миллиметровой бумаге, используя циркуль для измерения параметров гнездового хода и ячеек. Строение ячеек изучали под бинокулярным микроскопом МБС-9, снабженным окулярной сеткой, и также рисовали на миллиметровой бумаге. Рисунки гнезд и ячеек, представленные в работе, выполнены путем прорисовки фотографий и черновых рисунков, сделанных на миллиметровой бумаге. Преимагинальные стадии ос, найденные в ячейках гнезд, помещали в короткие отрезки стеклянных трубок, закрытые с обеих сторон плотными ватными тампонами и снабженные этикетками с номером гнезда и ячейки. До выхода имаго их хранили в неотапливаемом помещении.
С каждого из участков гнездования E. amadanensis отбирали пробы почвы для анализа содержания физических фракций по методу Рутковского и определения гранулометрического класса по методу Качинского (Вадюнина, Корчагина, 1986).
Фенологию лёта имаго изучали по материалам коллекций Таврической академии Крымского федерального университета им. В.И. Вернадского (Симферополь), Зоологического института РАН (Санкт-Петербург), Зоологического музея Московского государственного университета им. М.В. Ломоносова (Москва), Института зоологии им. И.И. Шмальгаузена НАН Украины (Киев), Одесского государственного зоопарка (Одесса), а также частных коллекций Д.В. Пузанова (Евпатория) и А.В. Амолина (Донецк). Всего было исследовано 342 экз. E. amadanensis, собранных в Крыму в 1887–2017 гг. (из них 336 имели полные даты сборов на этикетках).
РЕЗУЛЬТАТЫ
Субстрат гнездования
Самки E. amadanensis гнездились в земле на горизонтальных участках террас или пологих склонах холмов и балок. Участки гнездования вида в Лисьей бухте представляли собой полупустыню с участием Artemisia lercheana Weber ex Stechm., A. taurica Willd., Galatella villosa (L.) Rchb. f. (Asteraceae) и различных видов семейств Chenopodiaceae и Poaceae (рис. 1a). Чаще всего осы выбирали место для гнезда под этими растениями, маскируя таким образом вход в него. На берегу оз. Бараколь участки гнездования представляли собой солончак с растрескавшейся почвой. Растительность здесь была сформирована сообществом Halimione verrucifera (M. Bieb.) Aellen, Petrosimonia oppositifolia (Pall.) Litv. (Chenopodiaceae) и Puccinellia fominii Bilyk (Poaceae). В балке Кипчак гнезда ос были обнаружены на грунтовой дороге, почти лишенной растительности.
Рис. 1.
Биология Eustenancistrocerus amadanensis: a – терраса у подножия бедленда – типичный участок гнездования и охоты; b – самка, добывающая воду из влажной земли; c – самка, добывающая воду из лужи; d – самка, сооружающая входную трубку в гнездо; e – полностью отстроенная входная трубка в гнездо; f – самка, возвращающаяся в гнездо после полета за водой; g – самка, извлекающая крупный комочек земли из гнездового хода. Масштаб: e – 5 мм.

Гранулометрический состав почвы, в которой осы устраивали свои гнезда, варьировал в широких пределах: от супесей до тяжелых суглинков, близких по свойствам к глинам (табл. 2). Почва на участках гнездования, расположенных в Лисьей бухте на террасах под бедлендами (1 и 2), характеризовалась высокой долей илистой фракции, смываемой с этих бедлендов, и обладала свойствами супеси. На приморском склоне (участок 3) было обнаружено абсолютное преобладание в почве пылеватой фракции; свойства этой почвы были промежуточными между средним и тяжелым суглинком. Участок, расположенный на приморской террасе (4), характеризовался наибольшим содержанием в почве песчаной фракции и относительно большой долей илистой фракции; эта почва обладала свойствами супеси, близкой к легкому суглинку. Почва на участках, расположенных на берегу оз. Бараколь (5 и 6), характеризовалась наибольшим содержанием глинистой фракции и обладала свойствами тяжелых суглинков, близких к глинам. В балке Кипчак (участок 7) обнаружено примерно равное содержание пылеватой и илистой фракций при средней доле песчаной и глинистой; эта почва обладала свойствами среднего суглинка.
Таблица 2.
Характеристика почвы на участках гнездования Eustenancistrocerus amadanensis
| № | Удельный вес твердой фазы, г/см3 | Содержание физических фракций по методу Рутковского, % | Гранулометрический класс по методу Качинского | |||
|---|---|---|---|---|---|---|
| >0.5 мм (песчаная) | 0.5–0.05 мм (пылеватая) | 0.05–0.005 мм (илистая) | <0.005 мм (глинистая) | |||
| 1 | 2.75 | 7.49 | 8.56 | 83.95 | 0.00 | Супесь |
| 2 | 2.67 | 14.00 | 17.2 | 66.05 | 2.75 | Супесь |
| 3 | 2.10 | 3.96 | 87.08 | 1.63 | 7.33 | Средний суглинок – тяжелый суглинок |
| 4 | 2.87 | 27.03 | 19.25 | 52.14 | 1.58 | Супесь – легкий суглинок |
| 5 | 2.24 | 11.66 | 15.86 | 58.73 | 13.75 | Тяжелый суглинок |
| 6 | 2.48 | 10.53 | 22.37 | 56.56 | 10.54 | Тяжелый суглинок – глина |
| 7 | 2.43 | 8.9 | 42.05 | 41.26 | 7.79 | Средний суглинок |
Примечание. Нумерация участков соответствует табл. 1.
Гнездовая активность
Самки E. amadanensis начинали гнездование с поиска места для гнезда, однако продолжительность поиска измерить не удалось. После того как место для гнезда было выбрано, самка отрыгивала из зобика на поверхность земли каплю воды и приступала к формированию небольших комочков грязи. Эти комочки она укладывала по кругу, выстраивая, таким образом, кольцо вокруг будущего входа в гнездо. Через некоторое время она улетала для пополнения запасов воды. Самки посещали открытые источники, такие как небольшие ручьи и лужи, реже берег моря. Для сбора воды им было достаточно высосать ее из влажной земли на дне пересыхающего ручья или на берегу моря (рис. 1b). Однако при наличии луж осы предпочитали добывать воду из них, садясь при этом прямо на поверхность воды и удерживаясь на ней за счет сил поверхностного натяжения (рис. 1c). Вернувшись в гнездо с запасом воды в зобике, самка продолжала извлекать из гнездового хода небольшие комочки земли, формируя из них входную трубку. При этом голова и передние ноги осы всегда находились внутри трубки, а средние и задние ноги – снаружи (рис. 1d). Таким образом, по мере углубления гнездового хода входная трубка увеличивалась в высоту. Готовые трубки имели высоту 5–10 мм и располагались под углом примерно 30°–45° по отношению к поверхности земли. Они были сплошными в основании и имели ажурную структуру в дистальной части (рис. 1e).
После завершения строительства трубки самка продолжала углублять гнездовой ход. При этом она также регулярно улетала за водой, за исключением тех дней, когда земля была влажная после дождя. Вернувшись с запасом воды, она садилась на входную трубку гнезда (рис. 1f) и затем заходила в него вентральной стороной тела кверху. После этого она извлекала из гнездового хода более крупные комочки земли (рис. 1g). Самка взлетала с каждым таким комочком в воздух и выбрасывала его на расстоянии 20–50 см от входа в гнездо. Одной порции воды хватало для формирования 3–4 таких комочков. Иногда самка на время прерывала вынос крупных комочков земли и опять выносила несколько небольших комочков, прикрепляя их к вершине входной трубки, дополнительно удлиняя ее. Затем она возобновляла вынос крупных комочков. В случае повреждения входной трубки гнезда людьми или дождем самка восстанавливала ее, а затем также возобновляла вынос крупных комочков.
Темп гнездостроительной активности самок E. amadanensis был довольно низким. В одном из отмеченных случаев самка выгрызала гнездовой ход глубиной 4 см, как минимум, два дня. Другая самка делала перерывы в работе и по часу и более находилась внутри гнездового хода головой к выходу. Общее время ее работы неизвестно, однако в итоге она бросила гнездо на стадии гнездового хода глубиной 3.5 см.
Закончив выгрызание гнездового хода, самка откладывала яйцо и приступала к провиантированию. После этого она запечатывала ячейку пробкой из земли и либо откладывала следующее яйцо в том же гнездовом ходе, либо выгрызала новый гнездовой ход в том же гнезде, либо запечатывала все гнездо и приступала к следующему. В случае разрушения входной трубки во время провиантирования ячейки, самка не восстанавливала ее долгое время, до тех пор, пока процесс провиантирования не завершался.
Процесс гнездования двух самок наблюдали в течение четырех дней, еще одной самки – пяти дней и еще двух самок – в течение восьми дней. Однако в каждом из этих случаев начало гнездования не было зафиксировано, так что реальная продолжительность их деятельности могла быть больше. В итоге гнезда двух первых самок были брошены на стадии провиантирования третьей ячейки во втором гнездовом ходе и на стадии выгрызания второго гнездового хода после запечатывания первой ячейки. Гнездо самки, которую наблюдали в течение пяти дней, было в итоге запечатано и содержало всего одну ячейку. Гнезда самок, которых наблюдали в течение восьми дней, также были в итоге запечатаны; одно из них содержало всего одну ячейку и пустой второй гнездовой ход, второе – четыре ячейки. Таким образом, общий темп гнездования E. amadanensis был также очень низким (1 ячейка за 2–8 дней).
Строение и состав гнезд
Из 11 раскопанных гнезд E. amadanensis пять были брошены самками до откладки яйца, то есть содержали только пустой гнездовой ход без ячеек. Оставшиеся шесть гнезд были успешными. Они имели глубину 5.0–7.5 см; диаметр гнездового хода составлял около 3 мм. Ячейки не отличались от гнездового хода по диаметру и имели длину 9–13 мм. Три гнезда с ячейками также были брошены самками до их запечатывания. Они содержали 1–3 ячейки (рис. 2a–2c). Многоячейковое гнездо имело линейно-ветвистую структуру: ячейки располагались как последовательно в одном гнездовом ходе, так и в разных гнездовых ходах (рис. 2a). Два одноячейковых гнезда содержали также пустые боковые ходы (рис. 2b–2c). Основной ход часто был отделен земляной перегородкой, сходной с перегородками, выполняющими функцию пробок ячеек (рис. 2a и 2c).
Рис. 2.
Строение гнезд Eustenancistrocerus amadanensis: a–с – брошенные гнезда, d–e – полностью отстроенные гнезда. 1–3 – номера ячеек по порядку закладки. Масштаб 1 см.
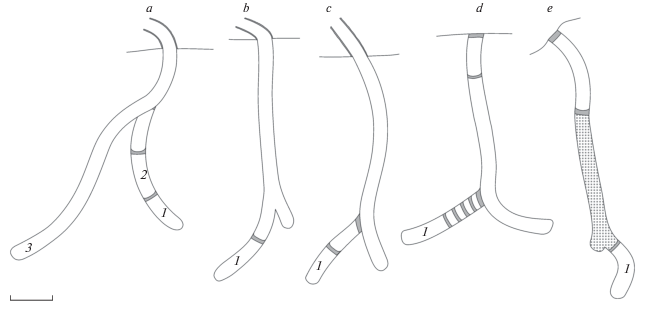
Запечатанные гнезда содержали 1–4 ячейки. Два одноячейковых гнезда также содержали пустые боковые ходы (рис. 2d–2e). В одном из этих гнезд имелись три дополнительные перегородки между пробкой ячейки и пробкой гнездового хода (рис. 2d). Во всех трех запечатанных гнездах также имелась дополнительная перегородка в гнездовом ходе на расстоянии 1–2 см от поверхности земли. В одном из гнезд пространство между этой перегородкой и ячейкой было засыпано рыхлой землей (рис. 2e). Происхождение этой земли неизвестно. Во время наблюдений за построившей это гнездо самкой было отмечено, что она выносила из гнездового хода мелкие комочки сухой земли после того, как входная трубка была разрушена дождем (рис. 3a). Гнездо, содержавшее четыре ячейки, имело ветвистую структуру. К сожалению, зарисовать его строение не удалось, поскольку оно располагалось в земле с большим количеством корней и развалилось при раскопке. Гнезда были запечатаны конечной пробкой вровень с поверхностью земли. Входные трубки запечатанных гнезд были полностью разобраны самками ос.
Рис. 3.
Биология Eustenancistrocerus amadanensis: a – самка, извлекающая мелкий комочек сухой земли из гнезда, входная трубка которого была разрушена дождем; b – ячейка вскрытого гнезда с предкуколкой в коконе; c – самка, извлекающая гусеницу из чехлика (показан стрелкой) на побеге Galatella villosa; d – самка, питающаяся нектаром на цветке Limonium scoparium; e – самцы, ночующие на соцветии Halimione verrucifera. Масштаб: b – 5 мм.

Трофические связи
Самки E. amadanensis охотились на гусениц двух видов молей-чехлоносок (Lepidoptera, Coleophoridae), обитающих на Artemisia spp. и Galatella villosa. Вывести из гусениц имаго и идентифицировать эти виды не удалось, однако обитающий на G. villosa вид определенно относился к роду Casignetella Strand 1928. Помимо A. lercheana, A. taurica и G. villosa, самки ос во время охоты обследовали и побеги различных видов семейства Chenopodiaceae (прежде всего, Camphorosma monspeliaca L.), а также соплодия Petrorhagia prolifera (L.) P.W. Ball et Heywood (Caryophyllaceae) и Sanguisorba minor subsp. balearica (Nyman) Muñoz Garm. et C. Navarro (Rosaceae). Обнаружив чехлик с гусеницей, оса сразу же приступала к ее извлечению и парализации, не перемещая чехлик с кормового растения (рис. 3c).
Питание имаго нектаром цветков было отмечено на 10 видах растений из восьми семейств (табл. 3). При этом основную роль в питании ос играли цветки Limonium spp. (Plumbaginaceae). Осы посещали не только свежераспустившиеся цветки этого рода, но и отцветшие цветки с увядшим венчиком. На цветках других растений были отмечены лишь единичные особи ос (1–3 экз.). Большинство кормовых видов растений имели открытые нектарники. Лишь у Thymus tauricus Klokov et Des.-Shost. и Limonium spp. нектар был расположен внутри трубки венчика. Однако венчики этих цветков неглубокие, что позволяло осам E. amadanensis легко извлекать из них нектар хоботком (рис. 3d).
Таблица 3.
Видовой состав растений, на цветках которых отмечено питание Eustenancistrocerus amadanensis в Крыму
| Вид растения | Самки | Самцы |
|---|---|---|
| Apiaceae | ||
| Eryngium campestre L. | + | − |
| Pimpinella tragium Vill. | − | + |
| Apocynaceae | ||
| Cynanchum acutum L. | − | + |
| Butomaceae | ||
| Butomus umbellatus L. | + | − |
| Chenopodiaceae | ||
| Halimione verrucifera (M. Bieb.) Aellen | + | − |
| Convolvulaceae | ||
| Cuscuta sp. | + | − |
| Lamiaceae | ||
| Thymus tauricus Klokov et Des.-Shost. | + | − |
| Plumbaginaceae | ||
| Limonium platyphyllum Lincz. | + | + |
| Limonium scoparium (Pall. ex Willd.) Stank. (= L. gmelinii auct.) | + | + |
| Ranunculaceae | ||
| Nigella arvensis L. | − | + |
Развитие потомства, фенология и соотношение полов
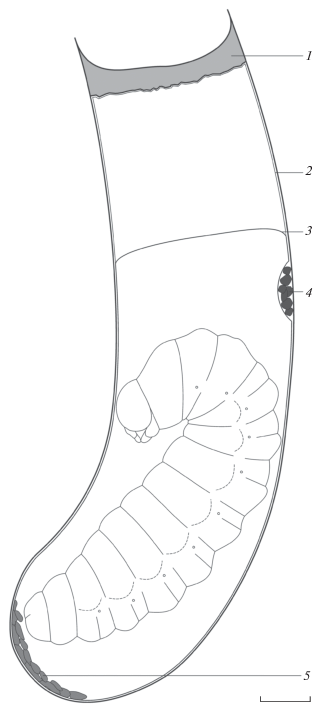
Ячейки вскрытых гнезд E. amadanensis содержали предкуколок (рис. 3b) или куколок в коконах; яиц и питающихся личинок обнаружено не было. Кокон состоял из двух слоев (рис. 4). Наружный слой представлял собой многочисленные тонкие нити, полностью покрывающие внутреннюю полость ячейки, за исключением небольшого участка, где были собраны остатки провизии (экскременты гусениц). Наружный слой кокона плотно прилегал к стенкам ячейки и не отделялся от них, однако при его изготовлении не была использована жидкость, которая могла проникнуть в стенки ячейки и скрепить их. Внутренний слой кокона имел вид коричневатой пергаментообразной пленки, так же плотно прилегающей к стенкам ячейки, но отделяющейся от них в случае применения силы. В базальных двух третях ячейки оба слоя кокона были слиты между собой, однако на некотором расстоянии от пробки ячейки внутренний слой кокона отходил от ее стенок и замыкался в виде выпуклого мениска. Меконий находился внутри внутреннего слоя кокона на дне ячейки. Предкуколки ос имели изогнутую форму и бледно-желтый цвет. Размеры коконов несколько превышали размеры предкуколок, которые располагались внутри внутреннего слоя кокона довольно свободно (рис. 3b и 4).
Рис. 4.
Строение ячейки Eustenancistrocerus amadanensis: 1 – земляная пробка, 2 – наружный слой кокона (для наглядности показан на расстоянии от стенок ячейки, в действительности же он плотно прилегает к ним), 3 – свободная часть внутреннего слоя кокона, 4 – экскременты жертв, 5 – меконий. Масштаб 1 мм.
Три гнезда, которые наблюдали в июне 2014 г. и затем раскопали 29 числа, содержали куколок. Вывести из них имаго не удалось, однако само присутствие куколок в гнездах, отстроенных в начале лета, свидетельствует о наличии двух поколений в году, а не одного, как предполагалось ранее (Фатерыга, 2009; Fateryga et al., 2013). Одно гнездо, которое наблюдали в августе 2016 г. и затем раскопали 1 сентября, содержало предкуколку, которая окуклилась на следующий год. Самка осы вышла из куколки 19 июля. Предкуколки из двух гнезд, которые наблюдали в июне 2019 г. и затем раскопали 1 июля, впали в зимнюю диапаузу, не окуклившись. Все это свидетельствует о том, что вид дает два поколения в году, но второе поколение факультативное: потомство первого поколения может перезимовать, не дав второго поколения, кроме того, потомство второго поколения может выйти на следующий год во второй половине лета, “пропустив”, таким образом, период гнездования первого поколения.
По данным исследованных коллекций, лёт самок E. amadanensis в Крыму был отмечен с 7 июня по 28 августа, лёт самцов – с 5 июня по 29 августа. Фенология лёта вида, представленная на диаграмме (рис. 5), в целом подтверждает вывод о наличии в году двух поколений, которым соответствуют два пика в середине июня и в начале августа. Однако следует иметь в виду, что как таковой границы между сроками лёта поколений нет, и в середине лета одновременно могут встречаться особи как первого, так и второго поколений.
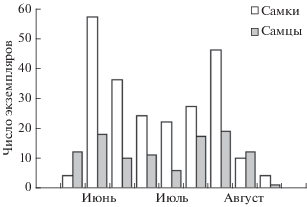
В коллекционных сборах E. amadanensis обнаружено более чем в два раза больше самок (232), чем самцов (110). Такое соотношение полов в коллекциях не обязательно отражает реальное соотношение полов в природе, которое может быть установлено лишь по результатам исследования большого числа гнезд (см. Иванов, Фатерыга, 2006), чего у данного вида сделать не удалось.
Гнездовые паразиты и репродуктивный успех
В одной из 11 ячеек шести гнезд был обнаружен кокон осы-блестянки (Hymenoptera, Chrysididae). К сожалению, предкуколка из этого кокона погибла, и установить ее видовую принадлежность не удалось. Еще в одной ячейке E. amadanensis погибло яйцо и развилась плесень. Остальные девять ячеек были успешными (репродуктивный успех составил 81.1%), хотя все потомство, находившееся во время раскопки гнезд на стадии куколки, погибло при транспортировке в лабораторию.
Поведение самцов
Самцы E. amadanensis были обнаружены на участках гнездования и вблизи источников воды. Чаще всего они патрулировали территорию, совершая более или менее регулярные полеты вдоль русел ручьев (в том числе пересохших) и вдоль грунтовых дорог. Иногда они прерывали патрулирование для питания на цветках. Один раз (2 августа 2014 г.) в 18.20 (солнечное время) удалось наблюдать двух самцов, спящих на соцветии Halimione verrucifera (рис. 3e). При этом к ним пытался присоединиться третий самец, но затем он улетел. Ухаживание и копуляцию наблюдать не удалось.
ОБСУЖДЕНИЕ
Особенности биологии E. amadanensis сходны с известными особенностями биологии E. blanchardianus (Ferton, 1914). Оба вида самостоятельно выгрызают гнездовой ход в земле, используя для ее размягчения отрыгиваемую воду, и даже одинаково умеют добывать воду, садясь прямо на ее поверхность. Оба вида выстраивают у входа в гнездо трубку из земли с той лишь разницей, что у E. blanchardianus она несколько длиннее (однако и сами осы этого вида крупнее, чем E. amadanensis). Также оба вида охотятся на гусениц молей-чехлоносок. Таким образом, сообщение о гнездовании E. amadanensis в замочных скважинах (Bingham, 1897) следует признать ошибочным. Возможно, сообщение на самом деле относится к представителю другого рода, например Stenancistrocerus de Saussure 1863, для которого достоверно зарегистрировано гнездование в готовых полостях (Fateryga, 2020).
Биология ос рода Eustenancistrocerus в общих чертах сходна с биологией некоторых видов родов Stenodynerus de Saussure 1863, например Stenodynerus microstictus (Viereck 1906) (Evans, 1956; Hackney, Barrows, 1977), Antepipona de Saussure 1855, например Antepipona deflenda S. Saunders 1853 (= Odynerus parvulus Lepeletier de Saint-Fargeau 1841), A. orbitalis Herrich-Shäffer 1839 (Móczár, 1939) и A. scutellaris Giordani Soika 1985 (Gess, Gess, 1991), а также Jucancistrocerus Blüthgen 1938, например J. caspicus Giordani Soika 1970 (Фатерыга, Амолин, 2013). В то же время, необычными чертами строения гнезд E. amadanensis являются довольно длинные ходы и наличие пустых боковых ходов в запечатанных гнездах. Функция пустых ходов до конца не ясна. Возможно, они используются для добычи стройматериала (комочков земли), необходимого для запечатывания ячеек и основного хода или для восстановления входной трубки при ее частых поломках. Также может быть, что в этих ходах самки ос должны были устроить ячейки, но затем не стали этого делать по какой-либо причине (например, из-за бедной кормовой базы). Большая длина основного хода, не занятого ячейками, может быть связана с тем, что почва, в которой способен гнездиться E. amadanensis, менее плотная, по сравнению с той, где гнездятся представители других родов. Например, J. caspicus гнездится только в глине, а E. amadanensis – в суглинках различной плотности и даже супесях. Более глубокие ходы в менее плотной почве необходимы для лучшей защиты ячеек от случайных внешних механических воздействий, а также от дождевой воды.
Еще одна интересная особенность, обнаруженная у E. amadanensis, – два типа комочков земли, которые самки извлекают из гнездового хода при его выгрызании: небольшие, используемые для строительства входной трубки, и крупные, которые самка осы выбрасывает. У гнездящихся в почве ос родов Odynerus Latreille 1802, Paragymnomerus Blüthgen 1938 и Paravespa Radoszkowski 1886 такого не наблюдается: комочки, используемые для строительства трубки, и выбрасываемые комочки имеют приблизительно одинаковые размеры (Fateryga, 2013, 2018; Fateryga, Ivanov, 2013). Для родов, родственных Eustenancistrocerus, данных о размерах этих комочков недостаточно, однако известно, что самки Stenancistrocerus obstrictus (Morawitz 1895), гнездящиеся в готовых ходах в земле, используют для строительства входной трубки крупные комочки, из которых изготовляют ажурный узор (Fateryga, 2020). E. amadanensis, напротив, строит ажурную часть трубки из цельных небольших комочков, просто укладывая их один к другому.
Несмотря на низкий темп гнездования, E. amadanensis является широко распространенным и довольно многочисленным видом. Такому его успеху могла способствовать, в частности, адаптация к гнездованию в разнообразных типах почвы. Для сравнения J. caspicus гнездится только в глинистых обрывах (Фатерыга, Амолин, 2013), а представители рода Brachyodynerus Blüthgen 1938 выгрызают боковые ходы внутри трещин суглинистой почвы, что было показано на примере B. magnificus (Morawitz 1867) (Fateryga et al., 2017). Оба типа субстрата очень специфичны и встречаются не везде, и, как следствие, представители родов Jucancistrocerus и Brachyodynerus довольно редки в природе. E. amadanensis, напротив, гнездится на относительно горизонтальных поверхностях и в почве с широким диапазоном гранулометрического состава. Этот вид встречается гораздо чаще, чем представители родов Jucancistrocerus и Brachyodynerus. При этом B. magnificus также охотится на гусениц молей-чехлоносок, хотя охота у него протекает немного иначе, чем у E. amadanensis. Самка B. magnificus вначале отрывает чехлик от субстрата и перелетает с ним на соседнее соцветие кормового растения, а уже затем начинает извлекать из чехлика гусеницу (Fateryga et al., 2017). Однако такие различия кажутся несущественными для биологического успеха вида.
Род Eustenancistrocerus относят к одной из терминальных клад ос-эвменин, которая включает также Stenodynerus и Antepipona (Bank et al., 2017). Роды Jucancistrocerus и Stenancistrocerus, вероятно, также следует относить к этой кладе, хотя они и не были включены в филогенетические реконструкции подсемейства (Bank et al., 2017; Piekarski et al., 2018). Для этой клады характерно наибольшее разнообразие гнезд (в готовых полостях, в земле и на поверхности субстрата). Предполагается, что исходным способом гнездования в этой кладе было гнездование в готовых полостях, а самостоятельное выгрызание ходов в земле возникло вторично (см. Fateryga, 2020). Одним из аргументов в пользу такой гипотезы может быть, в частности, строение коконов E. amadanensis, сходное с коконами некоторых видов, гнездящихся в готовых полостях, например рода Euodynerus Dalla Torre 1904 (Фатерыга, 2012), также относимого к этой кладе (Bank et al., 2017; Piekarski et al., 2018). Коконы всех этих ос заметно отличаются от таковых у представителей клады Odynerini s. str., у которых кокон пропитывает боковые стенки и дно ячейки (Fateryga, 2013, 2018; Fateryga, Ivanov, 2013).
Список литературы
Вадюнина А.Ф., Корчагина З.А., 1986. Методы исследования физических свойств почв. Третье издание. М.: Агропромиздат. 416 с.
Иванов С.П., Фатерыга А.В., 2006. Биология гнездования одиночной складчатокрылой осы Syneuodynerus egregius (Hymenoptera: Vespidae: Eumeninae) в Крыму // Вестник зоологии. Т. 40. № 4. С. 341–349.
Фатерыга А.В., 2009. Фенология лета складчатокрылых ос (Hymenoptera: Vespidae) в Крыму // Известия Харьковского энтомологического общества. 2008. Т. 16. № 1/2. С. 57–63.
Фатерыга А.В., 2010. Трофические связи складчатокрылых ос (Hymenoptera, Vespidae) с цветковыми растениями в Крыму // Энтомологическое обозрение. Т. 89. № 2. С. 380–389.
Фатерыга А.В., 2012. Строение гнезд четырех видов ос рода Euodynerus Dalla Torre (Hymenoptera, Vespidae: Eumeninae) // Энтомологическое обозрение. Т. 91. № 3. С. 520–532.
Фатерыга А.В., Амолин А.В., 2013. Гнездование и биология Jucancistrocerus caspicus (Hymenoptera, Vespidae, Eumeninae) // Зоологический журнал. Т. 92. № 8. С. 935–940.
Antropov A.V., Fateryga A.V., 2017. Family Vespidae // Annotated Catalogue of the Hymenoptera of Russia. V. 1. Symphyta and Apocrita: Aculeata. Eds Lelej A.S., Proshchalykin M.Yu., Loktionov V.M. Saint-Petersburg: Zoological Institute RAS. P. 175–196.
Bank S., Sann M., Mayer C., Meusemann K., Donath A., Podsiadlowski L., Kozlov A., Petersen M., Krogmann L., Meier R., Rosa P., Schmitt T., Wurdack M., Liu S., Zhou X., Misof B., Peters R.S., Niehuis O., 2017. Transcriptome and target DNA enrichment sequence data provide new insights into the phylogeny of vespid wasps (Hymenoptera: Aculeata: Vespidae) // Molecular Phylogenetics and Evolution. V. 116. P. 213–226.
Bingham C.T., 1897. The Fauna of British India, Including Ceylon and Burma: Hymenoptera. V. 1. Wasps and Bees. London: Taylor and Francis. xxix + 579 p. + 4 pls.
Carpenter J.M., Gusenleitner J., Madl M., 2010. A catalogue of the Eumeninae (Hymenoptera: Vespidae) of the Ethiopian Region excluding Malagasy Subregion. Part II: Genera Delta de Saussure 1885 to Zethus Fabricius 1804 and species incertae sedis // Linzer Biologische Beiträge. Bd. 42. H. 1. S. 95–315.
Evans H.E., 1956. Notes on the biology of four species of ground-nesting Vespidae // Proceedings of the Entomological Society of Washington. V. 58. № 5. P. 265–270.
Gess F.W., Gess S.K., 1991. Some aspects of the ethology of five species of Eumenidae (Hymenoptera) in southern Africa // Annals of the Cape Provincial Museums Natural History. V. 18. № 12. P. 245–270.
Fateryga A.V., 2013. Nesting biology of Odynerus albopictus calcaratus (Morawitz, 1885) and Odynerus femoratus de Saussure, 1856 (Hymenoptera: Vespidae: Eumeninae) // Journal of Insects. Article ID 597 583. P. 1–8.
Fateryga A.V., 2018. Nesting biology of Paragymnomerus signaticollis tauricus (Kostylev, 1940) (Hymenoptera: Vespidae: Eumeninae) // Zootaxa. V. 4378. № 3. P. 429–441.
Fateryga A.V., 2020. Nesting biology and distribution of Stenancistrocerus (Paratropancistrocerus) obstrictus (Morawitz, 1895) (Hymenoptera: Vespidae: Eumeninae) // Zootaxa. V. 4718. № 4. P. 591–600.
Fateryga A.V., Budashkin Yu.I., Puzanov D.V., 2013. On the ecology of the wasp Eustenancistrocerus amadanensis (Hymenoptera, Vespidae, Eumeninae) in the Crimea // Vestnik Zoologii. V. 47. № 2. P. 166.
Fateryga A.V., Ivanov S.P., 2013. Nesting biology of Paravespa rex (von Schulthess 1924) (Hymenoptera: Vespidae: Eumeninae) in the Crimea, Ukraine // Zootaxa. V. 3721. № 6. P. 589–600.
Fateryga A.V., Ivanov S.P., Budashkin Yu.I., 2017. First data on the bionomics of Brachyodynerus magnificus magnificus (Morawitz, 1867) (Hymenoptera: Vespidae: Eumeninae) // Zootaxa. V. 4312. № 2. P. 343–354.
Fateryga A.V., Mokrousov M.V., 2019. New records of eumenine wasps (Hymenoptera: Vespidae: Eumeninae) from Russia with description of a new species of Leptochilus de Saussure, 1853 // Zootaxa. V. 4612. № 3. P. 412–422.
Fateryga A.V., Proshchalykin M.Yu., Aliyev Kh.A., Maharramov M.M., 2019. To the knowledge of eumenine wasps (Hymenoptera: Vespidae: Eumeninae) of Nakhchivan Autonomous Republic of Azerbaijan // Far Eastern Entomologist. № 379. P. 25–32.
Ferton C., 1914. Notes détachées sur l’instinct des Hyménoptères mellifères et ravisseurs (8e Série) avec la description de quelques espèces nouvelles // Annales de la Société Entomologique de France. V. 83. P. 81–119.
Gusenleitner J., 2013. Die Gattungen der Eumeninae im Nahen Osten, in Nordafrika und in Arabien (Hymenoptera: Vespidae: Eumeninae) // Linzer Biologische Beiträge. Bd. 45. H. 1. S. 5–107.
Hackney J.M., Barrows E.M., 1977. Biological notes on Stenodynerus microstictus (Hymenoptera: Eumenidae) // Journal of the Washington Academy of Science. V. 67. № 1. P. 34–37.
Kostylev G., 1928. Materialien zur Kenntnis der Vespidenfauna der östlichen Krim // Entomologische Mitteilungen. Bd. 17. H. 6. S. 398–407.
Móczár L., 1939. Beobachtungen über den Nestbau einiger Odynerus-Arten // Zoologischer Anzeiger. V. 125. № 3/4. P. 70–80.
Piekarski P.K., Carpenter J.M., Lemmon A.R., Moriarty Lemmon E., Sharanowski B.J., 2018. Phylogenomic evidence overturns current conceptions of social evolution in wasps (Vespidae) // Molecular Biology and Evolution. V. 35. № 9. P. 2097–2109.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал