

Зоологический журнал, 2021, T. 100, № 3, стр. 299-306
Морфометрические и репродуктивные особенности полосатого гологлаза (Ablepharus bivittatus (Menetries 1832), Reptilia, Scincidae) в Талышских горах
А. А. Кидов a, *, Т. Э. Кондратова a
a Российский государственный аграрный университет – МСХА им. К.А. Тимирязева
127550 Москва, Россия
* E-mail: kidov_a@mail.ru
Поступила в редакцию 12.02.2020
После доработки 24.05.2020
Принята к публикации 30.05.2020
Аннотация
Приведены результаты изучения морфометрических и репродуктивных показателей полосатого гологлаза в горно-ксерофитном поясе Талышских гор. Животные были собраны в мае 2018 и мае 2019 гг. в окрестностях селений Ханегях-е-Олийа, Минаабад, Мирзанек и Анбаран около города Намин (шахрестан Намин, провинция Ардебиль, Исламская Республика Иран). Часть самок содержали в лабораторных условиях для получения яиц. Произведены семь промеров и рассчитаны их индексы. Длина тела взрослых самцов в изученной популяции составляла 39.1–47.3 мм, самок – 42.4–54.4 мм, молодых ящериц (годовиков) – 30.5–39.0 мм. Половой диморфизм отмечен по двум промерам и пяти индексам. Беременные самки имели длину тела 42.4–53.7 мм. В яйцеводах самок содержалось от 1 до 4 яиц. Отловленные самки в лабораторных условиях откладывали яйца с 20 мая по 11 июня. Кладки содержали 2–4 яйца. Яйца имели длину 8.2–11.9 мм, ширину 3.9–5.9 мм, массу 0.165–0.210 г. Выживаемость яиц за период инкубации 72%. Молодь в условиях лаборатории вылуплялась в период с 9 по 18 июля. Длительность инкубации 43–56 суток. Длина тела (SVL) молоди сразу после вылупления 20.6–24.0 мм, длина хвоста – 26.8–38.0 мм. Масса новорожденных особей 0.185–0.256 г. Полученные данные сравниваются с результатами других исследований.
Полосатый гологлаз (Ablepharus bivittatus (Menetries 1832)) является одним из наиболее широко распространенных представителей рода Ablepharus Lichtenstein 1823 (Еремченко, Щербак, 1986). Вид был описан с крайней северной границы ареала (в Талышских горах из окрестностей села Пирембель в современном Ярдымлинском р-не Азербайджана), а большая часть его ареала лежит в пределах северного и западного Ирана (провинции Западный Азербайджан, Восточный Азербайджан, Курдистан, Ардебиль, Гилян, Мазендаран, Казвин, Тегеран, Керманшах, Фарс, Хамедан, Семнан, Северный Хорасан, Хорасан Разави) (Anderson, 1999; Safaei-Mahroo et al., 2015; Sanchooli, 2016; Karamiani et al., 2017), и лишь на периферии вид проникает в Туркмению, Азербайджан, Армению (Банников и др., 1977; Еремченко, Щербак, 1986; Ананьева и др., 1998; Arakelyan et al., 2011) и Турцию (Baran, Atatür, 1998; Ilgaz et al., 2007). В условиях пессимума краевые популяции нередко изолированы друг от друга и находятся в угнетенном состоянии (Ananjeva et al., 2006; Arakelyan et al., 2011; Бунятова и др., 2012; Ahmadov, 2013). На большей же части видового ареала в Иране полосатый гологлаз обычен, а местами многочислен (Ahmadzadeh et al., 2008). В частности, это справедливо и для горно-ксерофитного пояса южных и юго-западных склонов Талышского хребта в пределах иранской провинции Ардебиль, где этот вид в весенний период является самым встречаемым представителем герпетофауны (Кидов, 2019).
Несмотря на локально высокую численность, A. bivittatus трудно назвать хорошо изученным видом. Например, сведения об изменчивости морфометрических показателей полосатого гологлаза базируются на результатах изучения лишь десятков особей, зачастую единичных экземпляров из каждого локалитета (Еремченко, Щербак, 1986; Ilgaz et al., 2007; Karamiani et al., 2017). Репродуктивная биология A. bivittatus также остается недостаточно исследованной, информация ограничивается сведениями о числе и размерах яиц в яйцеводах единичных вскрытых самок, и о степени развития семенников у самцов (Банников и др., 1977; Джафарова, 1984; Еремченко, Щербак, 1986).
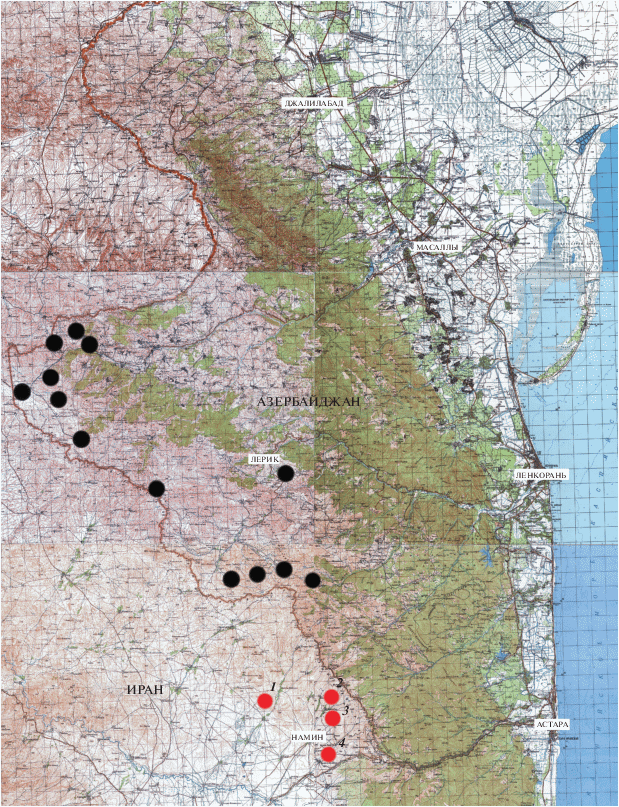
В азербайджанской части Талышских гор полосатый гологлаз к настоящему времени известен из 13 точек в Ярдымлинской, Деманской и Диабарской (Зувандской) внутригорных котловинах на территории Ярдымлинского и Лерикского р-нов (рис. 1). Так, в июле 1901 г. в окрестностях Дыманского поста (село Деман в Ярдымлинском р-не) собрал A. bivittatus А.М. Завадский, позднее передавший этот экземпляр в Зоологический музей МГУ. Соболевский (1929), на основании сборов Э.П. Менетриэ, Г. Ледера, А.Б. Шелковникова и А.Н. Кириченко, приводит этот вид для истоков р. Виляжчай на границе с Ираном, для окрестностей селений Пирембель и Розанова (часто упоминается также под названиями Розанов и Розано), для пограничных постов Аваш и Дыманского, около горы Мара-Юрт (сейчас на картах обычно обозначается как Кюмюр-Кѐй или Кямяр-Куа, 2492.8 м над ур. м.). В фондах отделения орнитологии и герпетологии Зоологического института РАН хранится кадастр находок земноводных и пресмыкающихся юго-восточного Азербайджана. В этом кадастре, составленном советским орнитологом А.А. Кузнецовым, в период с мая 1973 г. по октябрь 1978 г. отмечены три точки находок полосатого гологлаза в Ярдымлинском р-не – слева от грунтовой дороги между селами Аваш и Деман, между селами Деман и Арвана, а также на повороте дороги у Родниковой балки. И.С. Даревский в мае 1974 г. отловил экземпляр A. bivittatus в окрестностях Кельвяза в Лерикском р-не, этот экземпляр хранится в коллекции Зоологического института РАН. Еремченко и Щербак (1986) перечисляют большинство из вышеупомянутых точек, а также, ссылаясь на работу Соболевского (1929), отмечают полосатого гологлаза для села Джанги (вероятно, Джангамиран или Джони Лерикского р-на). Отметим, что в монографии Н.И. Соболевского этот локалитет не упоминается, а вышеупомянутые авторы имели в виду другой источник. Ахмедов (1985) нашел A. bivittatus в окрестностях сел Деман, Аваш и Пирембель Ярдымлинского р-на, а также в окрестностях сел Джони, Пирасура, Хилдара и Мистан Лерикского р-на. Позднее тот же автор (Ahmadov, 2013) отмечает, что полосатый гологлаз в Азербайджане редок и находится под охраной. Таким образом, изучение его морфометрических и репродуктивных характеристик представляет определенные трудности. Вероятно, по этой причине Еремченко и Щербак (1986) в своих исследованиях A. bivittatus искусственно объединяли в одну выборку (состоящую всего из двух самцов и десяти самок) ящериц из двух удаленных друг от друга участков ареала в пределах Азербайджана – с Малого Кавказа и Талыша. Нами была предпринята попытка охарактеризовать особенности размерных характеристик и размножения полосатого гологлаза на территории Иранского Талыша, где этот вид является массовым.
Рис. 1.
Распространение Ablepharus bivittatus в Талышских горах. Места сбора материала в провинции Ардебиль, Иран: 1 – Анбаран, 2 – Мирзанек, 3 – Минаабад, 4 – Ханегях-е-Олийа.
МАТЕРИАЛ И МЕТОДИКА
Материалом для исследований послужили ящерицы, собранные в мае 2018 и 2019 гг. Животных отлавливали в пригороде Намина: в окрестностях селений Анбаран (на некоторых картах также Энберан или Эмберан) (38°27′ с.ш., 48°27′ в.д., 1565 м над ур. м.), Мирзанек (также Мирзанаг или Мызна) (38°27′ с.ш., 48°30′ в.д., 1605 м над ур. м.), Минаабад (также Мейнабад, Минавар, или Минаво) (38°27′ с.ш., 48°30′ в.д., 1600 м над ур. м.) и Ханегях-е-Олийа (также Ашагы-Ханегах) (38°25′ с.ш., 48°34′ в.д., 1460 м над ур. м.) в шахрестане Намин (провинция (остан) Ардебиль, Исламская Республика Иран) (рис. 1). Максимальное расстояние между пунктами сбора ящериц 11 км (между селами Анбаран и Ханегях-е-Олийа), в других случаях 2–9 км, эти пункты не разделены зоогеографическими преградами (реками, хребтами) и не имеют принципиальных различий по ландшафту и высоте (различия по этому показателю составляют 5–160 м). Это позволило нам считать животных из этих точек принадлежащими к одной популяции и рассматривать их как одну выборку.
Ящериц, собранных в 2018 г. (16 самцов, 32 самки и 16 молодых), непосредственно после поимки зафиксировали в этиловом спирте; они были использованы для изучения морфометрической изменчивости и плодовитости.
Все отловленные особи по результатам вскрытия подразделялись на три половозрастные группы – взрослые самцы, взрослые самки и молодь, вероятно, годовики после первой зимовки.
У всех особей при помощи электронного штангенциркуля марки Solar Digital Caliper (производитель – “Xueliee”, КНР) с погрешностью 0.1 мм оценивали промеры, предложенные ранее для изучения гологлазов (Еремченко, Щербак, 1986): SVL – длина тела, TL – длина хвоста, HW – ширина головы, HL – длина головы, LP1 – длина передней конечности, LP2 – длина задней конечности, GA – расстояние между передними и задними конечностями. Длину хвоста измеряли у животных, не имеющих заметных следов его повреждения или регенерации.
Также рассчитывали индексы: SVL/TL – отношение длины тела к длине хвоста, (GA1) – отношение расстояния между передними и задними конечностями к длине туловища, (HL1) – отношение длины головы к длине туловища, (HI) – отношение ширины головы к длине туловища, (LP1a) – отношение длины передней конечности к длине туловища, (LP2a) – отношение длины задней конечности к длине туловища.
В 2019 г. отловили девять взрослых самок, у которых прижизненно измерили длину тела (SVL), хвоста (TL) и массу. Их поместили в стеклянные контейнеры и содержали до получения кладок яиц по методике, многократно апробированной в работах по изучению репродуктивных показателей ящериц (Кидов, 2018; 2020; Кидов и др., 2018; 2019). Субстратом служила увлажняемая смесь торфяной крошки и крупного песка, уложенная слоем 2–3 см, в которую гологлазы откладывали яйца. Локальный обогрев дна контейнеров осуществляли при помощи нагревательных шнуров марки Repti Zoo RS7050 (КНР) мощностью 50W. Освещение и ультрафиолетовое облучение проводили люминесцентными лампами марки Sylvania Reptistar T8 (Германия) мощностью 20W. Животных кормили через день нимфами двупятнистого сверчка (Gryllus bimaculatus De Geer, 1773) и личинками большого мучного хруща (Tenebrio molitor Linnaeus, 1758).
Ежедневно в субстрате искали кладки. У найденных яиц при помощи штангенциркуля измеряли длину и ширину, а электронными весами Massa-K-BK-300 (производитель – “Масса-К”, Россия) с погрешностью до 0.005 г – массу. Поврежденные, деформированные или жировые яйца учитывали лишь при оценке плодовитости самок, однако в дальнейшем анализе не использовали. Сразу после измерений яйца помещали в открытые пластиковые контейнеры объемом 200 мл, наполненные смесью увлажненного торфа с песком слоем 3 см. Инкубацию осуществляли при температуре 27°C в инкубационном аппарате для рептилий “Herp Nursery II” (производитель – Lucky Reptile, КНР). У вылупляющейся молоди измеряли длину тела (SVL) и хвоста (TL), а также массу. Часть рожденных в лаборатории гологлазов была выпущена в природу, а часть оставлена для дальнейшего содержания и разведения.
Для статистической обработки материала использовали пакет программ Microsoft Excel и Statistica 6.0. Рассчитывали среднее арифметическое (M), стандартное отклонение (SD) и пределы варьирования признака (min–max). В связи с малым объемом выборки и тем, что разные половозрастные группы содержали разное количество изученных животных, статистическую значимость наблюдаемых различий оценивали при помощи непараметрического U-критерия Манна-Уитни (Uэмп), т.к. при этом не требуется наличия нормального распределения сравниваемых совокупностей.
РЕЗУЛЬТАТЫ
Гологлазы в Иранском Талыше, как и в азербайджанской части этой горной системы (Ахмедов, 1985), населяют участки горно-ксерофитного пояса, поросшие Acantholimon hohenackeri (Jaub. et Spach) Boiss., Astracantha aurea (Willd.) Podlech и A. meyeri (Boiss.) Podlech, реже – Onobrychis cornuta (L.) Desv., колючие подушки которых служат для них убежищами. Реже эти ящерицы попадаются на каменистых осыпях, обычно – на окраинах распаханных полей. Из синтопических видов в окрестностях Намина нами отмечены Bufotes sitibundus (Pallas 1771), Rana pseudodalmatina Eiselt et Schmidtler 1971, Paralaudakia caucasia (Eichwald 1831), Eremias strauchi Kessler 1878, Darevskia raddei (Boettger 1892), Iranolacerta brandtii (De Filippi 1863), Lacerta strigata Eichwald 1831 и Hemorrhois ravergieri (Ménétries 1832). В период проведения исследований гологлазы встречались на поверхности все светлое время суток, включая вечерние сумерки. Соотношение полов (самцы: самки) среди взрослых особей в изученной выборке составляло 1 : 2.
Гологлазы различных половозрастных групп различались по многим изученным морфометрическим характеристикам (табл. 1). Взрослые самцы статистически значимо отличались от самок по длине тела (SVL) (p ≤ 0.01), расстоянию между передними и задними конечностями (GA) (p ≤ 0.01), отношению расстояния между передними и задними конечностями к длине туловища (GA1) (p ≤ 0.01), отношению длины туловища к длине хвоста (SVL/TL) (p ≤ 0.01), отношению длины головы к длине туловища (HL1) (p ≤ 0.01), отношению длины передней конечности к длине туловища (LP1a) (p ≤ 0.01) и отношению длины задней конечности к длине туловища (LP2a) (p ≤ 0.01). Также взрослые животные, как самцы, так и самки, демонстрировали достоверные различия с молодью по большинству анализируемых морфометрических признаков. Так, самцы значимо отличались от молодых ящериц по длине тела (SVL) (p ≤ 0.01), длине хвоста (TL) (p ≤ 0.05), ширине головы (HW) (p ≤ 0.01), длине головы (HL) (p ≤ 0.01), длине передней конечности (LP1) (p ≤ 0.01), длине задней конечности (LP2) (p ≤ 0.01), расстоянию между передними и задними конечностями (GA) (p ≤ 0.01) и отношению длины передней конечности к длине туловища (LP1a) (p ≤ 0.05), а самки – по длине тела (SVL) (p ≤ 0.01), ширине го-ловы (HW) (p ≤ 0.01), длине головы (HL) (p ≤ 0.01), длине передней конечности (LP1) (p ≤ 0.01), длине задней конечности (LP2) (p ≤ 0.01), расстоянию между передними и задними конечностями (GA) (p ≤ 0.01), отношению расстояния между передними и задними конечностями к длине туловища (GA1) (p ≤ 0.01), отношению длины туловища к длине хвоста (L/TL) (p ≤ 0.01), отношению длины головы к длине туловища (HL1) (p ≤ 0.01), отношению ширины головы к длине туловища (HI) (p ≤ 0.05), отношению длины передней конечности к длине туловища (LP1a) (p ≤ 0.01) и отношению длины задней конечности к длине туловища (LP2a) (p ≤ 0.01).
Таблица 1.
Морфометрические показатели Ablepharus bivittatus в Иранском Талыше
| Показатель, мм | Самцы (n = 16) | Самки (n = 32) | Молодь (n = 16) |
|---|---|---|---|
| SVL | $\frac{{44.71 \pm 2.66}}{{39.1{\kern 1pt} - {\kern 1pt} 47.3}}$ | $\frac{{49.30 \pm 2.99}}{{42.4{\kern 1pt} - {\kern 1pt} 54.4}}$ | $\frac{{33.96 \pm 2.42}}{{30.5{\kern 1pt} - {\kern 1pt} 39.0}}$ |
| TL | $\frac{{68.75 \pm 49.70}}{{58.9{\kern 1pt} - {\kern 1pt} 81.7}}$ | $\frac{{61.64 \pm 8.93}}{{46.4{\kern 1pt} - {\kern 1pt} 74.7}}$ | $\frac{{50.84 \pm 3.12}}{{45.5{\kern 1pt} - {\kern 1pt} 53.5}}$ |
| HW | $\frac{{5.81 \pm 0.43}}{{4.8{\kern 1pt} - {\kern 1pt} 6.3}}$ | $\frac{{5.71 \pm 0.41}}{{4.6{\kern 1pt} - {\kern 1pt} 7.1}}$ | $\frac{{4.49 \pm 0.28}}{{3.9{\kern 1pt} - {\kern 1pt} 5.1}}$ |
| HL | $\frac{{8.76 \pm 0.39}}{{7.7{\kern 1pt} - {\kern 1pt} 9.2}}$ | $\frac{{8.51 \pm 0.38}}{{7.9{\kern 1pt} - {\kern 1pt} 9.5}}$ | $\frac{{7.00 \pm 0.47}}{{6.1{\kern 1pt} - {\kern 1pt} 7.8}}$ |
| LP1 | $\frac{{11.71 \pm 0.74}}{{10.4{\kern 1pt} - {\kern 1pt} 12.9}}$ | $\frac{{11.60 \pm 0.92}}{{9.6{\kern 1pt} - {\kern 1pt} 13.7}}$ | $\frac{{9.35 \pm 0.57}}{{8.6{\kern 1pt} - {\kern 1pt} 10.2}}$ |
| LP2 | $\frac{{16.62 \pm 0.95}}{{14.8{\kern 1pt} - {\kern 1pt} 18.2}}$ | $\frac{{16.80 \pm 1.08}}{{13.7{\kern 1pt} - {\kern 1pt} 18.5}}$ | $\frac{{12.82 \pm 0.70}}{{11.6{\kern 1pt} - {\kern 1pt} 14.0}}$ |
| GA | $\frac{{23.30 \pm 1.98}}{{19.9{\kern 1pt} - {\kern 1pt} 25.4}}$ | $\frac{{27.99 \pm 2.36}}{{20.8{\kern 1pt} - {\kern 1pt} 31.9}}$ | $\frac{{17.64 \pm 1.64}}{{15.2{\kern 1pt} - {\kern 1pt} 20.2}}$ |
| (GA1) | $\frac{{52.09 \pm 2.41}}{{47.66{\kern 1pt} - {\kern 1pt} 56.73}}$ | $\frac{{56.21 \pm 3.97}}{{46.32{\kern 1pt} - {\kern 1pt} 68.16}}$ | $\frac{{51.90 \pm 2.15}}{{49.67{\kern 1pt} - {\kern 1pt} 56.78}}$ |
| SVL/TL | $\frac{{0.65 \pm 0.10}}{{0.58{\kern 1pt} - {\kern 1pt} 0.76}}$ | $\frac{{1.27 \pm 0.28}}{{0.97{\kern 1pt} - {\kern 1pt} 1.86}}$ | $\frac{{0.64 \pm 0.08}}{{0.57{\kern 1pt} - {\kern 1pt} 0.75}}$ |
| (HL1) | $\frac{{19.63 \pm 0.71}}{{18.30{\kern 1pt} - {\kern 1pt} 21.23}}$ | $\frac{{17.61 \pm 1.53}}{{15.27{\kern 1pt} - {\kern 1pt} 22.66}}$ | $\frac{{20.85 \pm 1.08}}{{18.97{\kern 1pt} - {\kern 1pt} 22.67}}$ |
| (HI) | $\frac{{66.29 \pm 4.40}}{{59.52{\kern 1pt} - {\kern 1pt} 73.49}}$ | $\frac{{67.34 \pm 4.86}}{{56.79{\kern 1pt} - {\kern 1pt} 78.89}}$ | $\frac{{64.27 \pm 3.97}}{{56.41{\kern 1pt} - {\kern 1pt} 71.43}}$ |
| (LP1a) | $\frac{{26.24 \pm 1.67}}{{22.13{\kern 1pt} - {\kern 1pt} 29.01}}$ | $\frac{{23.90 \pm 2.50}}{{18.43{\kern 1pt} - {\kern 1pt} 31.28}}$ | $\frac{{27.73 \pm 1.68}}{{25.28{\kern 1pt} - {\kern 1pt} 31.80}}$ |
| (LP2a) | $\frac{{37.27 \pm 2.40}}{{33.40{\kern 1pt} - {\kern 1pt} 42.22}}$ | $\frac{{33.49 \pm 3.17}}{{25.18{\kern 1pt} - {\kern 1pt} 43.85}}$ | $\frac{{37.96 \pm 2.59}}{{34.10{\kern 1pt} - {\kern 1pt} 44.11}}$ |
Примечания. Над чертой M ± SD, под чертой min–max. SVL – длина тела, TL – длина хвоста, HW – ширина головы, HL – длина головы, LP1 – длина передней конечности, LP2 – длина задней конечности, GA – расстояние между передними и задними конечностями, (GA1) – отношение расстояния между передними и задними конечностями к длине туловища, (HL1) – отношение длины головы к длине туловища, (HI) – отношение ширины головы к длине туловища, (LP1a) – отношение длины передней конечности к длине туловища, (LP2a) – отношение длины задней конечности к длине туловища.
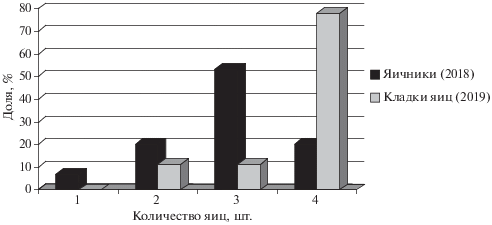
Вскрытые самки, содержавшие в яйцеводах яйца (n = 15), имели длину тела (SVL) 42.4–53.7 мм, в среднем 48.8 ± 3.18. Длина хвоста (TL) этих животных (n = 7) составила 50.8–69.4 мм, в среднем 61.9 ± 6.25. Расстояние между передними и задними конечностями (GA) у беременных самок (n = 15) варьировало в пределах 24.4–31.2 мм, в среднем 28.1 ± 1.99. В яйцеводах вскрытых самок находили 1 (1 самка), 2 (3 самки), 3 (8 самок) и 4 (3 самки) яйца, в среднем 2.9 ± 0.83.
Отловленные в 2019 г. беременные самки (n = 9) в лабораторных условиях откладывали яйца с 20 мая по 11 июня включительно. Из них 2 и 5 кладок были получены во II и III декадах мая соответственно, а по одной кладке – в I и II декадах июня. Кладки содержали 2 (1 кладка), 3 (1 кладка) и 4 (7 кладок) яйца, в среднем 3.7 ± 0.70. В первые сутки после откладки яйца (n = 33) имели длину 8.2–11.9 мм (10.8 ± 1.04), ширину (n = 33) – 3.9–5.9 мм (5.3 ± 0.56), массу (n=3) – 0.165–0.210 г (0.19 ± 0.023). Зависимости между репродуктивными и размерными показателями самок выявлено не было. Выживаемость яиц за период инкубации составила 72% (18 из 25 отложенных). Молодь в условиях лаборатории вылуплялась в период с 9 по 18 июля. В целом, длительность инкубации (n = 18) варьировала от 43 до 56 суток, в среднем 50 ± 3.8. Молодь в пределах одной кладки выходила из яиц с интервалом 0–3 суток. Длина тела (SVL) молоди сразу после вылупления (n = 18) составляла 20.6–24.0 мм (22.4 ± 0.96), а длина хвоста – 26.8–38.0 мм (32.5 ± 2.69). Масса новорожденных варьировала в пределах 0.185–0.256 г (0.242 ± 0.0256).
ОБСУЖДЕНИЕ
Таким образом, самок в изученной выборке было в два раза больше, чем самцов. Существенное преобладание самок неоднократно было отмечено в некоторых популяциях и других видов гологлазов (Еремченко, Щербак, 1986). Вероятно, это явление может быть объяснено не реальной диспропорцией в половом составе популяции, а большей доступностью самок для коллектора. По нашим наблюдениям, в мае самки демонстрируют более высокую наземную активность, чем самцы, обнаруживаемые преимущественно в убежищах (пустоты под камнями). Видимо, беременные самки имеют высокую потребность в прогреве и поэтому чаще встречаются на поверхности.
Ранее отмечалось (Еремченко, Щербак, 1986), что длина тела (SVL) A. bivittatus в целом по ареалу составляет 41.0–50.0 мм (M = 45.40, SD = 0.40) для самцов и 40.5–61.0 мм (M = 49.55, SD = 0.70) для самок. Самцы полосатого гологлаза из Иранского Талыша имели меньшие минимальные и средние значения этого показателя (табл. 1), тогда как оценки длины тела самок лежали в пределах изменчивости, отмеченной для вида.
Как и в других изученных популяциях (Еремченко, Щербак, 1986; Ilgaz et al., 2007; Karamiani et al., 2017), у гологлазов из Иранского Талыша хорошо выражен половой диморфизм по размерным признакам: самки крупнее самцов. Считается (Еремченко, Щербак, 1986), что самцы A. bivittatus становятся половозрелыми при достижении длины тела 41 мм, реже – 39–40 мм, а самки – при длине 43 мм. В нашей выборке самый мелкий самец с хорошо выраженными признаками полового диморфизма в окраске имел длину 39.1 мм, а самая мелкая самка с яйцами в яйцеводах – 42.4 мм.
При сравнении по длине тела гологлазов из Талыша с ящерицами из других локалитетов также отмечены некоторые различия. Так, они в среднем крупнее A. bivittatus из иранских провинций Восточный Азербайджан и Хамедан: по данным Карамиани с соавторами (Karamiani et al., 2017), средняя длина тела самцов в этих регионах 42.7 мм (SD = 0.52), а самок – 48.1 мм (SD = 0.65). В то же время гологлазы талышской популяции мельче ящериц из восточной Турции: Ильгаз с соавторами (Ilgaz et al., 2007) приводят значения этого показателя 37.9–47.9 мм (M = 45.16; SD = 0.81) для самцов и 46.6–56.0 (M = 50.9; SD = 0.93) для самок.
Размножение полосатого гологлаза описано по-разному в работах разных авторов. Так, Терентьев и Чернов (1949) отмечают, что откладка 3–5 яиц длиной 10.0–11.5 мм происходит в июне и июле. Алекперов (1978) цитирует эти данные без изменений, однако добавляет, что молодые ящерицы появляются в конце августа и имеют длину тела 23.2–27.0 мм. Банников с соавторами (1977) пишут, что откладка яиц у этого вида начинается с последних чисел мая и длится до второй половины июня. В кладках (которых, по их мнению, может быть две в сезоне) содержится обычно 4, реже – 5 яиц размером 9.0 × 5.5 мм, и молодые гологлазы с длиной тела 36.0–40.0 мм вылупляются в конце августа и сентябре. Еремченко и Щербак (1986) приводят сведения Банникова с соавторами (1977), однако уточняют, что яйца после откладки имеют размеры 9.0–11.0 × 5.0–5.5 мм, а длина тела у молоди после вылупления 22.0–38.0 мм.
Согласно нашим результатам изучения репродуктивных возможностей самок A. bivittatus в горах Иранского Талыша, плодовитость ящериц в исследуемой популяции варьирует в пределах от 1 до 4 яиц, причем наиболее обычны кладки с 3–4 яйцами (рис. 2). Интересно, что более половины вскрытых беременных самок имели в яйцеводах по три яйца (53%), тогда как подавляющее большинство полученных в лабораторных условиях кладок (80%) насчитывали по 4 яйца. Вероятно, наблюдаемые различия можно объяснить влиянием климатических или кормовых условий конкретного года, так как в первом случае ящериц отлавливали в 2018 г., а во втором – в 2019 г.
Рис. 2.
Распределение по числу яиц в яичниках (2018 г., n = 15) и кладках (2019 г., n = 9) у Ablepharus bivittatus.
Различия по размерам яиц в наших исследованиях (8.2–11.9 × 3.9–5.9 мм) были больше, чем различия, приводимые в литературе (Еремченко, Щербак, 1986).
Длина тела полученной нами молоди была в целом меньше отмеченной в более ранних исследованиях (Алекперов, 1978; Банников и др., 1977; Еремченко, Щербак, 1986). Очевидно, измеренные другими авторами молодые особи были пойманы в природе, и остается неизвестным, сколько времени прошло с момента их вылупления.
В то же время, можно предположить, что упомянутые выше различия данных, приводимых в работах разных исследователей, отражают высокую географическую изменчивость репродуктивных показателей этого вида, населяющего обширную территорию в Передней Азии в широких пределах высот.
Список литературы
Ахмедов С.Б., 1985. Распространение и биология полосатого гологлаза (Ablepharus bivittatus) в Азербайджане и прилегающих районах Армении // Вопросы герпетологии. Шестая Всесоюзная герпетологическая конференция (Ташкент, 18–20 сентября 1985 г.). Авторефераты докладов. Л.: Наука, Ленинградское отделение. С. 16–17.
Алекперов А.М., 1978. Земноводные и пресмыкающиеся Азербайджана. Баку: Элм. 264 с.
Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л., 1998. Земноводные и пресмыкающиеся. Энциклопедия природы России. М.: ABF. 576 с.
Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н., 1977. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение. 415 с.
Бунятова С.Н., Ахмедов С.Б., Джафаров А.Р., 2012. Экологический анализ герпетофауны (Testudinidae, Sauria, Serpentes) Талыша // Известия Самарского научного центра Российской академии наук. Т. 14. № 1. С. 144–149.
Джафарова С.К., 1984. Фауна, экология и охрана пресмыкающихся Малого Кавказа в пределах Азербайджана. Дис. … канд. биол. наук. Баку. 149 с.
Еремченко В.К., Щербак Н.Н., 1986. Аблефаридные ящерицы фауны СССР и сопредельных стран. Фрунзе: Илим. 171 с.
Кидов А.А., 2018. К репродуктивной биологии гирканской луговой ящерицы, Darevskia praticola hyrcanica (Lacertidae, Reptilia) // Современная герпетология. Т. 18. № 3–4. С. 118–124. https://doi.org/10.18500/1814-6090-2018-18-3-4-118-124
Кидов А.А., 2019. Весенний аспект герпетофауны Иранского Талыша // Известия высших учебных заведений. Поволжский регион. Естественные науки. № 1 (25). С. 50–60. https://doi.org/10.21685/2307-9150-2019-1-6
Кидов А.А., 2020. К изучению репродуктивной биологии скальной ящерицы (Darevskia saxicola, Reptilia, Lacertidae) // Зоологический журнал. Т. 99. № 11. С. 1293–1297. https://doi.org/10.31857/S004451342008005X
Кидов А.А., Иванов А.А., Кондратова Т.Э., Столярова Е.А., Немыко Е.А., 2019. О повторной откладке яиц у зеленобрюхих ящериц комплекса Darevskia (chlorogaster) (Reptilia, Lacertidae) // Современная герпетология. Т. 19. № 2–3. С. 153–159. https://doi.org/10.18500/1814-6090-2019-19-3-4-153-159
Кидов А.А., Немыко Е.А., Иванов А.А., Пыхов С.Г., 2018. О случаях позднего размножения у понтийской ящерицы, Darevskia pontica (Lantz et Cyren, 1919) на Северо-Западном Кавказе // Вестник Чувашского государственного педагогического университета имени И.Я. Яковлева. № 2 (98). С. 44–49.
Соболевский Н.И., 1929. Герпетофауна Талыша и Ленкоранской низменности (опыт зоогеографической монографии) // Мемуары зоологического отделения Общества любителей естествознания, антропологии и этнографии. Вып. 5. М. 143 с.
Терентьев П.В., Чернов С.А., 1949. Определитель пресмыкающихся и земноводных. М.: Советская наука. 340 с.
Ahmadov S.B., 2013. Zolaqlı çılpaqgöz (Two-streaked snake-eyed skink), Ablepharus bivittatus Menetries, 1832. Red Data Book of Azerbaijan Republic. Second Edition. Baku. S. 240–241.
Ahmadzadeh F., Kiabi B.H., Kami H.G., Hojjati V., 2008. A preliminary study of the lizard fauna and their habitats in Northwestern Iran // Asiatic Herpetological Research. V. 11. P. 1–9.
Ananjeva N.B., Orlov N.L., Khalikov R.G., Darevsky I.S., Ryabov S.A., Barabanov A.V., 2006. The Reptiles of Northern Eurasia. Sofia–Moscow: Faunistica Pensoft Series, Pensoft. 232 p.
Anderson S.C., 1999. The Lizards of Iran. Oxford, Ohio: Society for the Study of Amphibians and Reptiles. 442 p.
Arakelyan M.S., Danielyan F.D., Corti C., Sindaco R., Leviton A.E., 2011. Herpetofauna of Armenia and Nagorno-Karabakh. Salt Lake City: Society for the Study of Amphibians and Reptiles. 154 p. https://doi.org/10.13128/Acta_Herpetol-11380
Baran I., Atatür M.K., 1998. Turkish Herpetofauna. Amphibians and Reptiles. Ankara: T.C. Çevre Bakanligi. 214 p.
Ilgaz Ç., Kumlutas Y., Olgun K., Baran I., 2007. The morphology and distribution of Ablepharus bivittatus (Ménétriés, 1832) (Reptilia: Sauria: Scincidae) in Turkey // Russian Journal of Herpetology. V. 14. № 2. P. 91–97.
Karamiani R., Rastegar-Pouyani N., Rastegar-Pouyani E., Dehghani A., Banan-Khojasteh S.M., Mahmoodi M., 2017. Phenotypical variation and range extension distribution of the Snake-eyed Skink, Ablepharus bivittatus (Sauria: Scincidae) in Iran // Iranian Journal of Animal Biosystematics (IJAB). V. 13. № 2. P. 289–294. https://doi.org/10.22067/ijab.v13i2.59994
Safaei-Mahroo B., Ghaffari H., Fahimi H., Broomand S., Yazdanian M., Majd E.N., Saeed S., Yousefkhani H., Rezazadeh E., Hosseinzadeh M.S., Nasrabadi R., Rajabizadeh M., Mashayekhi M., Motesharei A., Naderi A., Kazemi S.M., 2015. The herpetofauna of Iran: checklist of taxonomy, distribution and conservation status // Asian Herpetological Research. V. 6. № 4. P. 257–290. https://doi.org/10.16373/j.cnki.ahr.140062
Sanchooli N., 2016. Modeling the potential distribution of Ablepharus bivittatus (Ménétriés, 1832), in Iran (Squamata: Sauria: Scincidae) // Herpetozoa. V. 29. № 1–2. P. 63–68.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал