

Зоологический журнал, 2021, T. 100, № 3, стр. 333-343
Астрономическое ориентирование у птиц
А. Д. Золотарёва a, *, Н. С. Чернецов a, b, **
a Биологическая станция “Рыбачий”, Зоологический институт РАН
238535 Рыбачий, Калининградская обл., Россия
b С.-Петербургский государственный университет
199034 С.-Петербург, Россия
* E-mail: maytheforcebewithanna@gmail.com
** E-mail: nikita.chernetsov@gmail.com
Поступила в редакцию 07.05.2020
После доработки 02.06.2020
Принята к публикации 06.06.2020
Аннотация
Для успешного совершения миграций в сезонном и видоспецифичном направлении птицам необходимы компасные системы. Первыми были описаны солнечный и звездный компасы, однако с открытием магнитного компаса интерес исследователей в основном направлен на него. Солнечный и звездный компасы иногда объединяют в единую астрономическую компасную систему, противопоставляя ее магнитной, однако правомерность этого утверждения еще не доказана. Для ориентации по солнцу птица должна использовать систему компенсации его неравномерного движения в течение дня (т.н. “внутренние часы”). В связи с этой неравномерностью, а также региональными и сезонными различиями солнечной дуги, считается, что солнечный компас не используется при миграциях на большие расстояния. Птицы также используют солнечный поляризованный свет на закате и восходе для калибровки других компасов. В отличие от солнечного, звездный компас мигрирующих ночью птиц не зависит от чувства времени. Солнечный и звездный компасы птиц не являются врожденными. Считается, что птица обучается распознавать движение солнца по небу в течение первых недель жизни, а также находить центр вращения звездного неба, совпадающий с Полярной звездой, до первой миграции. Использование луны в качестве компасной системы для птиц маловероятно.
Многие птицы совершают миграции на огромные расстояния (сотни и тысячи километров), многократно превышающие размеры их индивидуальных участков. При этом они оказываются способными успешно находить видоспецифичные районы зимовок и возвращаться в районы рождения и предыдущего размножения с высокой точностью, которая свидетельствует о существовании у них высокоэффективных механизмов ориентации и навигации (Соколов, 1991; Чернецов, 2016).
Люди составляли календари, изготовляли часы, а также ориентировались в пространстве относительно сторон света по солнцу и звездам задолго до появления компаса и современных навигационных устройств. Неудивительно, что экспериментальное изучение ориентации и навигации животных было начато именно с проверки гипотезы использования ими астроориентиров. В настоящее время показано применение положения всевозможных небесных тел и их свойств как позвоночными, так и беспозвоночными животными в различных поведенческих контекстах, с использованием разнообразных ориентационных механизмов (см. обзоры Foster et al., 2018; Guilfor, Taylor, 2014). Среди позвоночных животных наиболее изученными в этом плане являются перелетные птицы (Wiltschko, Wiltschko, 2015; Mouritsen et al., 2016; Чернецов, 2016).
Способность птиц к нахождению верного направления может проявляться в двух формах – ориентации и навигации (Perdeck, 1958). В 50-х годах XX века немецкий ученый Густав Крамер предложил концепцию “карты и компаса” (Kramer, 1957, 1961), актуальную до сих пор. Согласно этой концепции, мигрирующая (или совершающая хоминг, т.е. возвращающаяся к гнезду, голубятне и т.п.) птица должна сначала определить собственное месторасположение относительно цели движения (этап карты), а затем выбрать и поддерживать направление относительно сторон света (этап компаса). В исследованиях по ориентации животных принято называть способность пользоваться картой, т.е. осознавать свое положение относительно цели перемещений, навигацией, а способность использовать компас, т.е. определять направление на стороны света, – ориентацией (Кишкинев, Чернецов, 2014; Чернецов, 2016). Данная модель исходит из представления о том, что система позиционирования (карта) и компасная система могут быть основаны на разных физических, а значит и сенсорных, механизмах (Wiltschko, Wiltschko, 2015). Это тесно функционально связанные, но отдельные системы.
В настоящее время показано не только использование разными видами птиц звездного и/или солнечного компаса, но и магнитного (Chernetsov, 2017), а также магнитной (Mouritsen, 2018) и запаховой карты (Gagliardo, 2013). Интересно, что открытие использования птицами магнитного поля Земли, совершенное в лаборатории немецкого ученого Фридриха Вильгельма Меркеля (Merkel, Fromme, 1958; Wiltschko, Wiltschko, 1972), сперва вызвало весьма скептическое отношение научной общественности (Дольник, 1973, 1975). Однако в дальнейшем признание существования магнитного компаса и все более и более нарастающий поток исследований этого феномена практически вытеснили интерес к астрономической ориентации птиц. Ориентация по астрономическим объектам чаще всего упоминается лишь в связи с иерархией и/или калибровкой компасных систем относительно друг друга у разных видов птиц. Это вряд ли можно считать оправданным, т.к. в области изучения астрономической ориентации до сих пор осталось большое количество нерешенных фундаментальных вопросов.
СОЛНЕЧНЫЙ КОМПАС
В середине прошлого века Густав Крамер обратил внимание на то, что активность содержащихся в неволе скворцов (Sturnus vulgaris) во время миграционного сезона носит направленный характер (Kramer, 1950). Крамер помещал птиц в круглые клетки и регистрировал их активность в разных секторах клетки. В результате этого эксперимента ученый доказал, что зафиксированное таким образом направление соответствует типичному направлению миграции птиц этого вида в природе в данное время года. При этом при сильной облачности птицы не ориентировались. Также Крамер обучил птиц выбирать одну из 12 кормушек с едой, расположенных на круговой арене, относительно определенного угла падения солнечных лучей. Изменяя этот угол при помощи зеркал, Крамер регистрировал выбор птицами кормушек уже относительно отраженных от зеркал лучей (Kramer, 1952).
Еще в первых работах ученым был поднят вопрос о неравномерности видимого движения солнца по небу и наличии у птиц возможности к компенсации этой неравномерности при помощи их внутренних часов (Kramer, 1952). На восходе и закате солнце быстро набирает высоту, а около полудня – медленно. При этом на восходе и закате солнечный азимут изменяется незначительно, а вот днем, когда солнце находится высоко, – наоборот. Соответственно, для успешной работы солнечного компаса внутренние часы птиц должны быть подстроены под эту закономерность.
Для проверки этого предположения немецким ученым Клаусом Шмидт-Кёнигом был предложен оригинальный метод (Schmidt-Koenig, 1958). Экспериментальных птиц (в данном случае почтовых голубей (Columba livia var. domestica)) некоторое время содержали при искусственном фотопериоде, который отличался от естественного на несколько часов. После синхронизации внутренних ритмов птиц этим искусственным фотопериодом с ними проводили эксперименты в естественном времени. Если “перевести” внутренние часы у голубей на 6 часов вперед, а затем утром в 6 утра местного солнечного времени их выпустить, птицы увидят Солнце не высоко над горизонтом и на юге, чего следовало бы ожидать, если бы их новый биоритм соответствовал действительности (голуби считают, что сейчас солнечный полдень), а существенно ниже и ближе к востоку. Экспериментальные птицы изменяли направление своего полета к голубятне, как если бы солнце действительно было на юге, при этом игнорируя тот факт, что оно низко над горизонтом. Это свидетельствует о том, что высота солнца над горизонтом птицами игнорируется, а важен азимут, т.е. направление на солнце (рис. 1). Для правильной интерпретации изменяющегося в течение дня азимута солнца птицы обладают “внутренними часами”, синхронизированными с астрономическим временем. Другими словами, солнечный компас птиц зависит от времени (Schmidt-Koenig, 1958; 1990; Able, Cherry, 1986).
Рис. 1.
Схема работы солнечного компаса, зависящего от времени: А – ориентация птицы при естественном фотопериоде; Б – ориентация птицы, чей фотопериод смещен на 6 часов вперед.
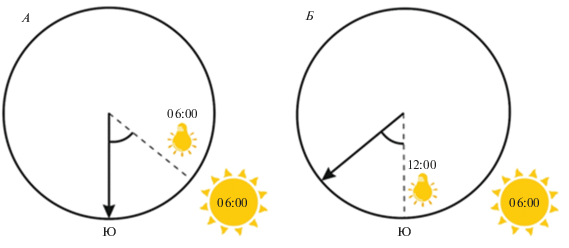
Этот вывод наглядно иллюстрирует работа группы ученых из лаборатории Вольфганга и Розиты Вильчко, выполненная в Новой Зеландии (Wiltschko et al., 2000). На 37° ю.ш. в течение первого часа после восхода солнца и часа после его заката азимут изменяется на 10°, тогда как в течение часа около полудня – на 40°. Голубей со смещенными на 4 часа вперед внутренними часами выпускали в разное время суток, при максимальной и минимальной разнице в солнечных азимутах. Голуби во всех случаях ошибались с выбором направления своего полета, исходя из изменения солнечного азимута в данные часы.
Помимо концепции солнечного компаса, Джеффри Мэтьюзом была выдвинута гипотеза полноценной навигации птиц (на примере почтовых голубей) по солнцу (Matthews, 1951, 1951a, 1953). В северном полушарии солнце восходит на восточной стороне и через юг в зените (здесь и далее в тексте мы условно называем так точку наиболее высокого положения солнца, поскольку за пределами широт, расположенных между северным и южным тропиками, солнце никогда не достигает истинного зенита – точки строго над головой наблюдателя) в полдень заходит на западной стороне. В южном полушарии восходит на восточной стороне и через север в зените в полдень заходит на западной стороне. Согласно идее Мэтьюза, голубь мог бы определять широту, т.е. к югу или северу от исходной точки (“дома”) он находится, сравнивая высоту солнца, которую он запомнил, в зените дома с высотой солнца в зените в новой точке (точке выпуска). Долготу птица могла бы определить, оценивая изменение солнечного азимута относительно своего чувства времени. Дома в полдень по внутренним часам птицы солнце было в зените, а в новой точке в это же время по ее внутренним часам оно либо прошло зенит (перемещение птицы на восток), либо не дошло до него (перемещение на запад) (рис. 2).
Рис. 2.
Иллюстрация гипотезы навигации по Солнцу (по: Matthews, 1953). Птица выпущена к северу и западу от дома. Ярким желтым цветом, снизу – высота солнца в текущем месте (в месте выпуска) в полдень. В северном полушарии солнце в зените расположено на юге. Тусклым цветом, сверху – высота солнца в полдень в предыдущем месте (дома), после чего – расположение солнца дома при солнце в зените в месте выпуска. Крупный пунктир – направление движение солнца. а – Разница в высоте солнца над горизонтом между домом и точкой выпуска, широта. b – Разница азимутов между домом и точкой выпуска, долгота. Рисунок выполнен не в масштабе.
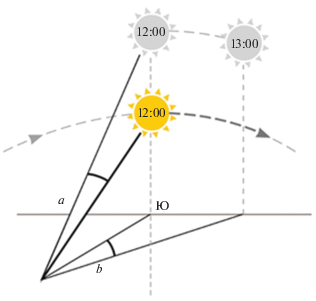
Таким образом, для определения долготы по солнцу птице было бы необходимо иметь два датчика времени: один, настроенный на время дома, а другой – измеряющий местное время. На данный момент такой механизм установления смещения, основанный на определении разницы времени по неподстраиваемым часам (независимым от внешнего времязадатчика (цайтгебера)), не описан ни для каких животных, в т.ч. и для птиц (Albus et al., 2005; Kishkinev et al., 2010; Cassone, Westneat, 2012). Однако накапливается все больше данных о том, что в гипоталамусе млекопитающих, в определенной группе нейронов, в т.н. супрахиазматических ядрах, разные популяции нейронов имеют разную скорость синхронизации к внешнему фотопериоду (Piggins, Loudon, 2005). Это гипотетически могло бы служить физиологической основой для вышеописанного механизма определения смещения по долготе.
Солнечная дуга зависит не только от времени суток, но и от географической широты и времени года. Чем дальше от экватора, тем по более плоской дуге мы видим двигающееся солнце. В разное время года солнце восходит не строго на восходе, а заходит не строго на западе. Например, на широте Москвы, 56° с.ш., восход летом на северо-востоке происходит примерно на 53° (закат – на 305°, СЗ), а зимой на юго-востоке, на 120° (закат – на 238°, ЮЗ).
Это означает, что птицы должны иметь механизмы компенсации к суточному неравномерному движению солнца, адаптированные к данному географическому району и данному времени года. Что, в свою очередь, делает крайне затруднительным использование солнечного компаса в качестве основной компасной системы во время миграций. При этом подобная компасная система применима для выполнения других, более локальных задач (Wiltschko, Wiltschko, 2015).
Большинство исследований солнечного компаса птиц проведено на почтовых голубях, которые, пусть и не совершают миграций, однако имеют высокую мотивацию к возвращению к своей голубятне и проявляют эту мотивацию круглый год (а не только в течение короткого периода размножения). К тому же эти птицы являются удобными лабораторными животными (Schmidt-Koenig, 1958; Keeton, 1979; Wiltschko et al., 1976, 1984; и др.). Для голубей в лаборатории рассчитана и точность работы солнечного компаса – она варьирует от 3.4° до 5.1° (McDonald, 1972). Авторы данного исследования отмечают невысокий показатель такой точности, однако компьютерная симуляция полетов выявила что этого будет достаточно для успешного хоминга с тех расстояний, с которых обычно возвращаются почтовые голуби (в большинстве случаев не более 100 км).
Солнечный компас может использоваться птицами и для поиска спрятанной ими еды. Такие опыты проводили, например, на скворцах (Kramer, 1952), на голубых кустарниковых сойках (Aphelocoma coerulescens) (Wiltschko, Balda, 1989), на черношапочных гаичках (Parus atricapillus) (Duff et al., 1998). Голуби также способны обучиться поиску еды по солнечному компасу, однако неясно, какой в этом биологический смысл, поскольку голуби не запасают пищу и не должны искать ранее сделанные запасы (Bingman, Gagliardo, 2006).
В отличие от магнитного компаса, который является врожденным, птицам, вероятнее всего, необходимо учиться пользоваться солнечным компасом. На это указывает эксперимент с молодыми голубями в возрасте 8–10 недель, которые не могли найти верное направление в условиях изменения их внутренних часов, в то время как птицы 12-недельного возраста с заданием справились (Wiltschko et al., 1981). Для того, чтобы компас сформировался, птицам необходимо наблюдать бóльшую часть видимого движения солнца по небу: наблюдений в течение лишь половины дня оказалось недостаточно (Budzynski et al., 2000; Wiltschko, Wiltschko, 1980).
ПОЛЯРИЗОВАННЫЙ СОЛНЕЧНЫЙ СВЕТ
Благодаря атмосферному рассеиванию, синее дневное небо является линейно поляризованным. Плоскость поляризации перпендикулярна направлению распространения волны от ее источника, т.е. в нашем случае – световым волнам от cолнца в природе. Таким образом, в природе животные могут использовать поляризацию световых лучей для навигационных целей, особенно если солнце находится за горизонтом (на закате или рассвете) или скрыто за облаками или деревьями (Brines, Gould, 1982).
В отличие от самого cолнца, траектория которого обычно пересекает горизонт не строго под углом в 90° и может быть сокращена для наблюдателя особенностями местности, во время истинного захода и восхода солнца на небе появляется полоса максимальной поляризации света, проходящая через зенит и перпендикулярная линии горизонта. Во время весеннего и осеннего равноденствия пересечение полосы максимальной поляризации от закатного и от рассветного солнца с горизонтом совпадает с осью север–юг, так что информацию о положении этих полос на восходе и на закате поляризованного света можно было бы использовать как независимую компасную систему. Во все остальное время года пересечение полосы максимальной поляризации с горизонтом на восходе происходит симметрично аналогичной полосе максимальной поляризации на закате относительно оси север – юг (рис. 3). Рахель Мухайм с соавторами (Muheim et al., 2006) предположили, что если бы птицы могли находить биссектрису угла между этими полосами, то они обладали бы отдельной компасной системой, которая бы не зависела ни от других компасов, ни от географического положения, ни от условий среды (рис. 3). В настоящее время эта гипотеза не подтверждена. Несмотря на ее простоту и логичность, данных о том, что птицы действительно пользуются этим механизмом определения сторон света, нет (Wiltschko et al., 2008).
Рис. 3.
Схематичная иллюстрация гипотезы использования мигрирующими птицами в качестве компасной системы поляризованного солнечного света на восходе и закате (по: Muheim et al., 2006). А – расположение полосы максимальной поляризации света на восходе; Б – расположение полосы максимальной поляризации света на закате; В – гипотетическое получение оси север–юг путем вычисления средней между полосой максимальной поляризации на восходе и на закате; Г, Д – гипотеза использования в качестве компасного направления расположения полосы максимальной гибридизации на восходе (Г) и закате (Д) относительно оси север–юг, выученной ранее (как на рис. В); Е – предполагаемые пути миграции саванных овсянок (P. s. sandwichensis), использующих разные стратегии калибровки своих компасов. Синяя центральная линия – путь птицы, использующей калибровку, полученную путем вычисления средней между полосой максимальной поляризации на восходе и на закате (как на рис. 3В). Фиолетовая нижняя линия – путь птицы, использующей калибровку только от восхода cолнца (как на рис. 3Г). Оранжевая верхняя линия – путь птицы, использующей калибровку только от захода солнца (как на рис. Д); с – географический север.
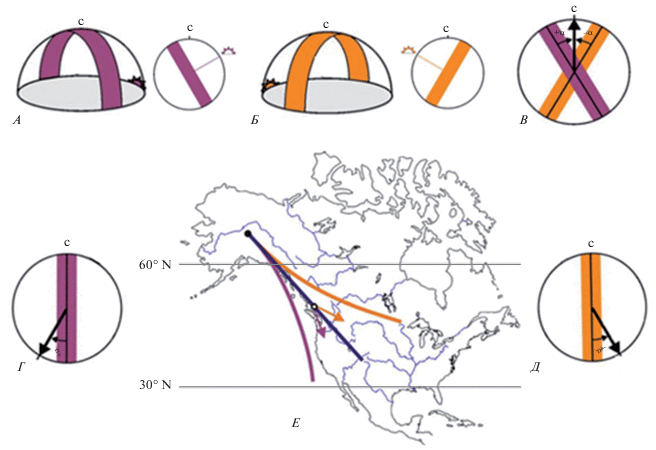
Несмотря на то, что возможность ориентации по солнцу даже в пасмурную погоду была описана на пчелах еще в середине прошлого века (Frisch, 1967), первые опыты, доказывающие использование поляризованного света птицами, появились значительно позже – сначала на голубях (Kreithen, Keeton, 1974), затем и на мигрирующих птицах (Moore, 1985; Able, 1989). Если первые опыты представляли собой обучение голубей распознаванию поляризованного света, то в случае с мигрирующими птицами использовались либо поляризационные фильтры (Muheim et al., 2006), либо конфликт магнитной информации, создаваемой внутри т.н. магнитных колец (Mouritsen, 2013) и зрительной информации от закатного неба (Bingman, Wiltschko, 1988; Cochran et al., 2004; Chernetsov et al., 2011). На сегодняшний день имеется большой объем противоречивых данных по конфликту этих компасов (см. обзор Liu, Chernetsov, 2012; Pakhomov, Chernetsov, 2020). Согласно имеющимся данным, информация по приоритету магнитного или солнечно-поляризационного компасов является видоспецифичной, если не популяционно-специфичной (Чернецов, 2016). Так, например, американские дрозды из рода Catharus калибруют свой магнитный компас по информации от закатного солнца (Cochran et al., 2004), в то время как певчие дрозды (Turdus philomelos) – нет (Chernetsov et al., 2011).
ЗВEЗДНЫЙ КОМПАС
Вдохновленный открытием Крамера солнечного компаса у птиц и методологией самой работы, другой немецкий исследователь, Эдгар Густав Франц Зауэр, в конце 1950-х годов начинает проводить ориентационные эксперименты с мигрирующими ночью птицами. Зауэр работал сначала под естественным ночным звездным небом (Sauer, 1957), а затем в планетарии, открыв таким образом саму возможность изучения влияния различных параметров ночного неба на поведение птиц в контролируемых и регулируемых условиях (Sauer, Sauer, 1960). Нельзя не отметить, что работы Зауэра выполнены на единичных особях (эксперименты 1958 г. – всего на двух садовых славках (Sylvia borin) и трех славках-черноголовках (Sylvia atricapilla)), без учета возможного влияния магнитного поля на ориентацию, и, следовательно, содержат ряд недоказанных допущений и утверждений. Так, он предполагал, что отсутствие ночной миграционной активности птиц в беззвездные ночи доказывает исключительную важность звезд. После того, как было установлено влияние геомагнитного поля на ориентацию птиц (Wiltschko, Merkel, 1966; Wiltschko, Wiltschko, 1972) и, в частности, значение света для работы магнитного компаса (Mouritsen, 2018) стало ясно, что общая картина намного сложнее.
Работы Зауэра вызвали критику и у современников (Wallraff, 1960; Emlen, 1967, 1967а со ссылкой на сообщения P. H. Klopfer, K. Schmidt-Koenig, A. Wolfson), в результате которой стало понятно, что эксперименты под звездным небом нуждаются в тщательной перепроверке. С середины 1960-х гг. американец Стивен Эмлен начинает ряд кропотливых исследований, многие из которых стали классическими. Для начала автор усовершенствовал модель круглой клетки Крамера (Emlen, Emlen, 1966). Клетка была уменьшена в размерах и приобрела коническую форму, на ее дне была установлена подушечка с чернилами. Птица, находящаяся в миграционном состоянии, прыгала на стенки конуса, покрытые бумагой, и оставляла на ней следы от лап, полученные при этом результаты потом можно было статистически обработать. Большинство лабораторных экспериментов по ориентации и навигации птиц до сих пор проводятся в этих клетках, так называемых конусах Эмлена, с различными модификациями – использование вместо чернил побеленной или иной бумаги, видео регистрации активности птицы в конусе, вариации размеров конуса и другие (Bianco et al., 2016).
Эмлен первым провел эксперимент, показавший существование звездного компаса у индиговых овсянковых кардиналов (Passerina cyanea) (Emlen, 1967, 1967а). Отловленных во время осенней миграции птиц на время тестов сажали в конуса Эмлена, расположенные под звездным небом планетария. В результате были получены контрольные направления птиц, соответствующие их ориентации под естественным звездным небом, после чего птицам было предъявлено “перевернутое небо”, с измененной на 180° осью север–юг (т.е. Полярная звезда, которая обычно находится над северной точкой горизонта в северном полушарии, была расположена на юге). Птицы изменили направление своей ориентации также на 180°, несмотря на то, что магнитное поле осталось без изменений.
В дальнейшем наличие у птиц звездного компаса было неоднократно подтверждено в различных лабораториях при разнообразных магнитных условиях. Важнейшим доказательством существования звездного компаса на сегодняшний день являются тесты в вертикальном магнитном поле, т.е. не несущем магнитной компасной информации. В вертикальном поле под звездами ночного неба или планетария эксперименты проводили на черноголовых славках (Viehmann, 1982), садовых славках (Wiltschko et al., 1987), саванных овсянках (Passerculus sandwichensis) (Bingman, 1983), мухоловках-пеструшках (Ficedula hypoleuca) (Bingman, 1984), обыкновенных горихвостках (Phoenicurus phoenicurus) (Mouritsen, 1998), зарянках (Erithacus rubecula) (Pakhomov et al., 2017).
Следующий вопрос, вставший перед исследователями, – каков механизм работы звездного компаса. Видимым следствием вращения Земли является движение всех звезд и созвездий ночного неба со скоростью 15° в час, кроме Полярной звезды, расположенной над осью вращения Земли и потому кажущейся с Земли неподвижной. В связи с этим есть три гипотетических варианта работы звездного компаса у птиц. Первый – птицы ориентируются относительно конкретных созвездий и, в связи с изменением их положения на небе в течение ночи, изменяют направление своей ориентации вслед за этими звездами или же мигрируют строго в определенный промежуток времени. Впрочем, путем проверки ориентации птиц в разное время ночи под неподвижным (не вращающимся) небом в планетарии было показано, что птицы во всех случаях выбирают направление, соответствующее сезонному направлению миграции свободно мигрирующих птиц (Emlen, 1967).
Второй вариант логически связан с первым – птицы используют звездный компас, зависимый от времени. Т.е. предполагается, что птица неким образом соотносит время и характерное для данного времени местоположение звезды/созвездия на небе, а затем делает “поправку” на время в курсе своего полета, аналогично описанному выше механизму работы солнечного компаса.
Однако полностью доказанным является третий вариант работы звездного компаса птиц, согласно которому ориентация мигрирующих птиц по звездам не зависит от времени ночи (Pakhomov et al., 2017). Еще ранние эксперименты со смещением звездного неба планетария на разное количество часов показывали способность индиговых овсянковых кардиналов, независимо от транслируемого им часа ночи, ориентироваться в направлении, характерном для данного сезона миграции (Emlen, 1967). Такой же результат был получен позднее на большем количестве мухоловок-пеструшек и славок-черноголовок. Эти птицы в течение 11 часов наблюдали неподвижное небо планетария, настроенного на полтретьего ночи, и все равно сохраняли способность ориентироваться в естественном направлении миграции (Mouritsen, Larsen, 2001). Наконец, зарянки, суточный ритм которых в условиях искусственного фотопериода был смещен на 4 часа вперед, успешно выбирали направление осенней миграции под естественным ночным небом без доступа к компасной магнитной информации (в вертикальном магнитном поле, Pakhomov et al., 2017).
Какую именно ориентационную информацию получают птицы от звездного неба? Для ориентации по звездам человек находит Полярную звезду, неподвижно расположенную над осью вращения Земли и, следовательно, предоставляющую ось север–юг. Неудивительно поэтому, что первый исследователь звездного компаса у птиц тоже начал с теста гипотезы ориентации птиц по Полярной звезде (Emlen, 1967). Однако уже первые эксперименты показали, что взрослые птицы, находящиеся в планетарии с выключенной Полярной звездой, прекрасно ориентируются. Включение и выключение в планетарии разных созвездий по отдельности (Большой Медведицы и Кассиопеи), а также Млечного пути не повлияло на возможность птиц использовать звездный компас (Emlen, 1967). При этом описанные эксперименты не подчеркивают незначимую роль этих созвездий и звезд в работе звездного компаса, а говорят о том, что даже в отсутствие упомянутых звезд птицы могут получать информацию, необходимую для успешной ориентации.
К тому же стало очевидным, что птицам важна возможность воспринимать рисунок созвездий относительно оси вращения звездного неба (совпадающей в настоящее время с Полярной звездой). Эмлен показал, что если птенцов индиговых овсянковых кардиналов выращивать под звездным небом планетария, вращающимся вокруг звезды Бетельгейзе (α Ориона), то с наступлением осеннего миграционного состояния они будут ориентироваться на юг именно от Бетельгейзе даже под естественным ночным небом, вращающимся уже вокруг Полярной звезды (Emlen, 1970, 1975). Уже сформированный под вращающимся звездным небом звездный компас взрослых птиц больше не нуждается во вращении, т.е. птицы могут ориентироваться и под неподвижными звездами планетария (Emlen, 1970; Wiltschko et al., 1987, Weindler et al., 1997, Mouritsen, Larsen, 2001).
В дальнейшем в ряде работ было продемонстрировано, что для успешного функционирования звездного компаса птицам вообще не обязательно наблюдать вращение именно созвездий – вполне достаточно 16 светящихся точек на черном фоне, причем даже без центральной точки, выполняющей роль “Полярной звезды” (Wiltschko et al., 1987; Alert et al., 2015). Соответственно, у молодых особей нет никакого врожденного знания про то, как должны выглядеть звезды, важно именно расположение звезд (или светящихся точечных объектов) относительно центра вращения и, возможно, относительно друг друга. Для птиц это имеет огромное эволюционное значение, т.к. один раз в течение нескольких тысяч лет происходит очередная смена одной звезды прецессионного круга, указывающей на север, на другую. Сейчас на север указывает Полярная звезда, а через 2000 лет будет Альраи (γ Цефея), что не помешает птицам успешно совершать свои ночные миграции.
Несколько кропотливых исследований было проведено по изучению деталей способности птиц к обучению распознавать ось вращения неба. Работы были выполнены на голубях, не являющихся мигрирующими птицами, зато способных к эффективному обучению в лабораторных условиях (Alert et al., 2015). После соответствующей тренировки голуби были в состоянии выбирать центр вращения экрана со светящимися точками, статистически достоверно отличающийся от центра массы изображения. Такие факторы, как разная плотность светящихся точек, различные комбинации точечных кластеров, точки рядом с центром вращения и дополнительные неподвижные точки, в конечном счете, не влияли на способность правильного выбора птицами центра вращения. Однако во всех экспериментах голуби справлялись с заданием только при скорости вращения экрана, не превышающей 5.6° в секунду (естественное вращение звездного неба составляет 360° в час, или 0.0042° в секунду). В настоящее время у голубей не известны зрительные нейроны, регистрирующие скорости больше 0.25° в секунду (Wolf-Oberhollenzer, Kirschfeld, 1994), да и те распознают не движения объектов относительно тела птицы, а движение самой птицы (Frost, 2010). Не исключено, что голуби могли бы использовать серию последовательных мгновенных снимков (snapshot), сопоставляя которые они вычисляли бы центр вращения экрана (Alert et al., 2015). Есть исследование, показывающее наличие зрительной памяти у голубей для пространственной ориентации (Pecchia et al., 2011), однако не понятно, можно ли экстраполировать эти результаты на звездное небо и функционирование звездного компаса. К тому же и сами эксперименты, выполненные на удобном для лаборатории модельном объекте, голубе, все же не равнозначны работе с мигрирующими птицами. Таким образом, в настоящее время нейрофизиологические основы работы звездного компаса остаются неизвестными не только у голубей, но и у диких птиц (Mouritsen et al., 2016).
Также остаются без ответа и некоторые вопросы онтогенеза обучения мигрирующих птиц ориентации по звездам. Не проверено утверждение, кочующее из статьи в статью еще со времен классических работ Эмлена, что молодые особи учатся распознавать вращение звездного неба только до своей первой миграции. Неизвестно, могут ли они научиться этому на более поздних этапах онтогенеза, например, весной следующего года. Если мигрирующие птицы могут научиться звездной ориентации весной, значит, это именно обучение, а не импринтинг, который может успешно пройти только в определенном возрасте. До конца не ясно и то, сколько времени нужно наблюдать это вращение птенцам. В большинстве экспериментов птенцы наблюдали вращающееся ночное небо две недели и больше – 1 месяц (Emlen, 1967, 1970; Wiltschko et al., 1987), 22 ночи (Able, Able, 1990), 7 недель (Prinz, Wiltschko, 1992) и 14 ночей (Weindler et al., 1995). Была предпринята только одна попытка установить минимальный срок экспозиции под звезды (Michalik et al., 2014). Зарянкам для формирования и успешной работы звездного компаса было недостаточно наблюдать вращение имитации звездного неба в течение одной или 7 ночей. Однако в случае, когда им предоставляли возможность наблюдать вращение имитации звездного неба в течение 21 ночи, они показывали нормальное ориентационное поведение.
Интересный момент заключается в том, что видимое вращение звездного неба предоставляет птицам лишь ось, относительно которой они могут ориентироваться, но непонятно, каким именно образом они выбирают направление этой оси, полярно меняющееся в следующий сезон миграции.
На сегодняшний момент имеется большой объем противоречивых данных по конфликту звездного и магнитного компасов (обзор Liu, Chernetsov, 2012), видоспецифичных, как и в случае с калибровкой магнитного и солнечного компасов.
ОРИЕНТАЦИЯ ПО ЛУНЕ И ДРУГИМ АСТРОНОМИЧЕСКИМ ТЕЛАМ
Гипотетическое использование Луны в качестве компаса осложняется тем, что во время лунного месяца она не всегда видна на небе, восходит и заходит каждый день позднее и восточнее и изменяет свою форму. В настоящее время показано использование луны и лунного поляризованного света для ориентации и навигации только у насекомых и ракообразных. Так, навозный жук Scarabaeus zambesianus использует очень слабо поляризованный свет от луны для построения ровной траектории катящегося навозного шарика (Dacke et al., 2003), а морские блохи Talitrus saltator – саму луну для нахождения кратчайшего пути от воды к суше (Papi, 1960). Упоминания об использовании луны в качестве ориентира у птиц встречаются лишь в самых ранних работах по изучению ориентации и навигации (Matthews, 1963), когда методологический аппарат разделения магнитного компаса и других компасных систем был еще не разработан. Более того, эксперименты по изучению ориентации и навигации птиц, наоборот, не проводятся в лунные ночи, в связи с описанным Эмленом явлением фототаксиса у ночных мигрантов, т.е. птица прыгает в более освещенный сектор клетки, а не туда, куда ее побуждало бы ночное миграционное беспокойство (Emlen, 1967 со ссылкой на эксперименты 1964 года). Наш опыт также подтверждает существование фототаксиса, который в разной степени развит у разных видов ночных мигрантов.
Для одного вида жуков из рода скарабеи – Scarabaeus satyrus – известна ориентация по Млечному пути (Dacke et al., 2013). Относительно него скарабеи катят навозные шарики по прямой траектории в разные стороны от навозной кучи, чтобы избежать конкуренции. Для птиц использование Млечного пути не доказано.
ЗАКЛЮЧЕНИЕ
На сегодняшний день принято считать ориентационную систему мигрирующих птиц избыточной (Чернецов, 2016; Chernetsov, 2017). Одних только компасных систем у птиц три – солнечная, звездная и магнитная.
Некоторыми исследователями солнечный компас, вместе со звездным, противопоставляется магнитному компасу (Wiltschko, Wiltschko, 2015). Действительно, магнитный компас является врожденным и пригодным для круглосуточного использования (Mouritsen, 2013), в то время как ориентации по солнцу и звездам птицы должны учиться в онтогенезе, наблюдая вращение небесных светил и некоторым образом обрабатывая эту информацию (Emlen, 1970; Wiltschko et al., 1981). Для успешной работы солнечного и звездного компасов необходим лишь зрительный анализатор, а для магнитного компаса нужна отдельная магниторецепторная сенсорная система (Кишкинёв, Чернецов, 2014). Несмотря на все это, правомерность такого объединения солнечного и звездного компасов в единую небесную компасную систему не подтверждена и не опровергнута. Существенным аргументом против этого слияния служит тот факт, что солнечный компас зависит от чувства времени птицы, а звездный независим от него и может успешно использоваться, даже если внутренние часы рассинхронизированы с внешним фотопериодом (Pakhomov et al., 2017).
Понятен все более возрастающий интерес к теме использования птицами магнитного поля Земли – помимо того, что у человека не выражена сама возможность к магниторецеции, у нас до сих пор нет четкого понимания механизма работы рецептора птиц для компасной магнитной информации и не описан рецептор для магнитной карты как таковой (Mouritsen, 2013; 2018). Однако на сегодняшний день все еще ожидает своего исследователя ряд фундаментальных вопросов об устройстве астрономических компасных систем птиц. Так как все работы по ориентации птиц по звездам проводились в Северном полушарии, наличие и механизмы работы звездного компаса в Южном полушарии не описаны. Есть еще множество белых пятен в наших знаниях об онтогенезе звездного компаса птиц; отсутствие навигации по звездам до конца не доказано. Вероятно, для понимания когнитивных механизмов, лежащих в основе восприятия птицами информации от небесных светил, нам потребуется более глубокое понимание устройства всего мозга птицы.
Список литературы
Дольник В.Р., 1973. Астрономическая ориентация птиц // Ориентация и территориальные связи популяций птиц. Рига: Зинатне. С. 14–61.
Дольник В.Р., 1975. Миграционное состояние птиц. М.: Наука. 398 с.
Кац Е.Б., Михельсон Х.А., 1978. Влияние изменения направления звездного и магнитного меридианов на ориентацию зарянок в круглых клетках в планетарии // Ориентация птиц. Рига: Зинатне. С. 180–193.
Кишкинёв Д.А., Чернецов Н.С., 2014. Магниторецепторные системы у птиц: обзор современных исследований // Журнал общей биологии. Т. 75. № 2. С. 104–123.
Соколов Л.В., 1991. Филопатрия и дисперсия птиц. Л. 233 с. (Труды Зоологического института АН СССР. Т. 230).
Чернецов Н.С., 2016. Ориентация и навигация мигрирующих птиц // Зоологический журнал. Т. 95. № 2. С. 128–146.
Able K.P., 1989. Skylight polarization patterns and the orientation of migratory birds // Journal of Experimental Biology. V. 141. P. 241–256.
Able K.P., Able M.A., 1990. Ontogeny of migratory orientation in the Savannah sparrow, Passerculus sandwichensis: Calibration of the magnetic compass // Animal Behaviour. V. 39. № 5. P. 905–913.
Able K.P., Cherry J.D., 1986. Mechanisms of dusk orientation in white-throated sparrows (Zonotrichia albicollis): clock-shift experiments // Journal of Comparative Physiology. V. 159. P. 107–113
Albus H., Vansteensel M.J., Michel S., Block G.D., Meijer J.H., 2005. A GABAergic mechanism is necessary for coupling dissociable ventral and dorsal regional oscillators within the circadian clock // Current Biology. V. 15. P. 886–893.
Alert B., Michalik A., Helduser S., Mouritsen H., Gunturkun O., 2015. Perceptual strategies of pigeons to detect a rotational center – a hint for star compass learning? // PLoS ONE. V. 10. № 3. P. e0119919.
Bianco G., Ilieva M., Veiback C., Ofj K., Gadomska A., Hendeby G., Felsberg M., Gustafsson F., Åkesson S., 2016. Emlen funnel experiments revisited: methods update for studying compass orientation in songbirds // Ecology and Evolution. V. 6. № 19. P. 6930–6942.
Bingman V.P., 1983. Importance of Earth’s magnetism for the sunset orientation of migratory naïve savannah sparrows // Monitoro Zoologico Italiano. V. 17. P. 395–400.
Bingman V.P., 1984. Night sky orientation of migratory pied flycatchers raised in different magnetic fields // Behavioral Ecology and Sociobiology. V. 15. P. 77–80.
Bingman V.P., Gagliardo A., 2006. Of birds and men: convergent evolution in hippocampal lateralization and spatial cognition // Cortex. V. 42. P. 99–100
Bingman V.P., Wiltschko W., 1988. Orientation of dunnocks (Prunella modularis) at sunset // Ethology. V. 77. P. 1–9.
Brines M.L., Gould J.L., 1982. Skylight polarization patterns and animal orientation // Journal of Experimental Biology. V. 96. P. 69–91.
Budzynski C.A., Dyer F.C., Bingman V.P., 2000. Partial experience with the arc of the sun is sufficient for all-day sun compass orientation in homing pigeons, Columba livia // Journal of Experimental Biology. V. 203. P. 2341–2348.
Cassone V.M., Westneat D.F., 2012. The bird of time: cognition and the avian biological clock // Frontiers in Molecular Neuroscience. V. 5. P. 32.
Chernetsov N., 2017. Compass systems // Journal of Comparative Physiology. V. 203. P. 447–453.
Chernetsov N., Kishkinev D., Kosarev V., Bolshakov C.V., 2011. Not all songbirds calibrate their magnetic compass from twilight cues: a telemetry study // Journal of Experimental Biology. V. 214. № 15. P. 2540–2543.
Cochran W.W., Mouritsen H., Wikelski M., 2004. Migrating songbirds recalibrate their magnetic compass daily from twilight cues // Science. V. 304. № 5669. P. 405–408.
Dacke M., Baird E., Byrne M., Scholtz CH., Warrant E.J., 2013. Dung beetles use the Milky Way for orientation // Current Biology. V. 23. P. 298–300.
Dacke M., Nordström P, Scholtz C.H., 2003. Twilight orientation to polarised light in the crepuscular dung beetle Scarabaeus zambesianus // Journal of Experimental Biology. V. 206. P. 1535–1543.
Duff S.J., Brownlie L.A., Sherry D.F., Sangster M., 1998. Sun compass and landmark orientation by black-capped chickadees (Parus atricapillus) // Journal of Experimental Biology. V. 24. P. 243–253.
Emlen S.T., 1967. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part I: Evidence of use of celestial cues // Auk. V. 84. № 3. P. 309–341.
Emlen S.T., 1967a. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part II: Mechanism of celestial orientation // Auk. V. 84. № 4. P. 463–489.
Emlen S.T., 1970. Celestial rotation: its importance in the development of migratory orientation // Science. V. 170. № 3963. P. 1198–1201.
Emlen S.T., 1975. The stellar-orientation system of a migratory bird // Scientific American. V. 233. P. 102–111.
Emlen S.T., Emlen J.T., 1966. A technique for recording migratory orientation in captive birds // Auk. V. 84. P. 300–342.
Foster J.J., Smolka J., Nilsson D-E., Dacke M., 2018. How animals follow the stars // Proceedings of the Royal Society B. V. 285. P. 20 172 322.
Frisch K., 1967. The Dance Language and Orientation of Bees. Cambridge, Harvard University Press.566 p.
Frost B.J., 2010. A taxonomy of different forms of visual motion detection and their underlying neural mechanisms // Brain Behavior and Evolution. V. 75. P. 218–35.
Gagliardo A., 2013. Forty years of olfactory navigation in birds // Journal of Experimental Biology. V. 216. № 12. P. 2165–2171.
Guilford T., Taylor GK., 2014. The sun compass revisited // Animal Behaviour. V. 97. P. 135–143.
Keeton W.T., 1979. Avian orientation and navigation: a brief overview // British Birds. V. 72. P. 451–470.
Kishkinev D., Chernetsov N., Mouritsen H., 2010. A double-clock or jetlag mechanism is unlikely to be involved in detection of east–west displacements in a long-distance avian migrant // Auk. V. 127. № 4. P. 773–780.
Kramer G., 1950. Weitere Analyse der Faktoren, welche die Zugaktivität des gekäfigten Vogels orientieren // Naturwissenschaften. № 37. S. 377–378.
Kramer G., 1952. Experiments on bird orientation // Ibis. V. 94. P. 265–285.
Kramer G., 1957. Experiments in bird orientation and their interpretation // Ibis. V. 99. № 2. P. 196–227.
Kramer G., 1961. Long-distance orientation // Biology and Comparative Physiology of Birds. New York, London: Academic Press. P. 341–371.
Kreithen M.L., Keeton W.T., 1974. Detection of polarized light by the homing pigeon, Columba livia // Journal of Comparative Physiology. V. 89. P. 83–92
Liu X., Chernetsov N., 2012. Avian orientation: multi-cue integration and calibration of compass systems // Chinese Birds. V. 3. № 1. P. 1–8.
Matthews G.V.T., 1951. The experimental investigation of navigation in homing pigeons // Journal of Experimental Biology. V. 28. № 4. P. 508–536.
Matthews G.V.T., 1951a. The sensory basis of bird navigation // Journal of Institute of Navigation. V. 4. P. 260–75.
Matthews G.V.T., 1953. Sun navigation in homing pigeons // Journal of Experimental Biology. V. 30. № 2. P. 243–267.
Matthews G.V.T., 1963. The astronomical basis of “nonsense” orientation // Proceedings of XIIIth International Ornithological Congress. Ithaca, N.Y. P. 415–429.
McDonald D.L., 1972. Some aspects of the use of visual cues in directional training of homing pigeons // Animal orientation and navigation. Galler S.R., Schmidt-Koenig K., Jacobs G.J., Belleville R.E. (eds). NASA Technical Report SP-262. P. 293–304.
Merkel F.W., Fromme H.G., 1958. Untersuchungen über das Orientierungsvermögen nächtlich ziehender Rotkehlchen (Erithacus rubecula) // Naturwissenschaften. Bd. 45 S. 499–500.
Michalik A., Alert B., Engels S., Lefeldt N., Mouritsen H., 2014. Star compass learning: how long does it take? // Journal of Ornithology. V. 155. № 1. P. 225–234.
Moore F.R., 1982. Sunset and the orientation of a nocturnal bird migrant: a mirror experiment // Behavioral Ecology and Sociobiology. V. 10. P. 153–155.
Moore F.R., 1985. Integration of environmental stimuli in the migratory orientation of the Savannah sparrow, Passerculus sandwichensis // Animal Behaviour. V. 33. № 2. P. 654–663.
Mouritsen H., 1998. Redstarts, Phoenicurus phoenicurus, can orient in a true-zero magnetic field // Animal Behaviour. V. 55. P. 1311–1324
Mouritsen H., 2013. The magnetic senses // Neurosciences – from molecule to behavior. A university textbook. Berlin, Heidelberg: Springer. P. 427–443.
Mouritsen H., 2018. Long-distance navigation and magnetoreception in migratory animals // Nature. V. 558. № 7708. P. 50–59.
Mouritsen H., Heyers D., Güntürkün O., 2016. The neural basis of long-distance navigation in birds // Annual Review of Physiology. V. 78. P. 133–154.
Mouritsen H., Larsen O.N., 2001. Migrating songbirds tested in computer-controlled Emlen funnels use stellar cues for a time-independent compass // Journal of Experimental Biology. V. 204. № 22. P. 3855–3865.
Muheim R., Phillips J.B., Akesson S., 2006. Polarized light cues underlie compass calibration in migratory songbirds // Science. V. 313. № 5788. P. 837–839.
Papi F., 1960. Orientation by night: the moon // Cold Spring Harbor Symposium on Quantitative Biology. V. 25. P. 475–480.
Pakhomov A., Anashina A., Chernetsov N., 2017. Further evidence of a time-independent stellar compass in a night-migrating songbird // Behavioral Ecology and Sociobiology. V. 71. P. 48.
Pakhomov A., Chernetsov N., 2020. A hierarchy of compass systems in migratory birds // Biological Communications. V. 65. № 3. P. 262–276.
Pecchia T., Gagliardo A, Vallortigara G., 2011. Stable panoramic views facilitate snap-shot like memories for spatial reorientation in homing pigeons // PLoS ONE. V. 6. № 7. P. e22657.
Perdeck A.C., 1958. Two types of orientation in migrating starlings, Sturnus vulgaris L., and chaffinches, Fringilla coelebs L., as revealed by displacement experiments // Ardea. V. 46. № 1. P. 1–37.
Piggins H.D., Loudon A., 2005. Circadian biology: clocks within clocks // Current Biology. V. 15. № 12. P. 455–457.
Prinz K., Wiltschko W., 1992. Migratory orientation of pied flycatchers: interaction of stellar and magnetic information during ontogeny // Animal Behaviour. V. 44. P. 539–545.
Sauer F., 1957. Die Sternenorientierung nächtlich ziehender Grasmucken (Sylvia atricapilla, borin und curruca) // Zeitschrift für Tierpsychologie. Bd. 14. H. 1. S. 29–70.
Sauer E.G.F., Sauer E.M., 1960. Star navigation of nocturnal migrating birds. The 1958 planetarium experiments // Cold Spring Harbor Symposia on Quantitative Biology. V. 25. P. 463–473.
Schmidt-Koenig K., 1958. Experimentelle Einflußname auf die 24-Stunden-Periodik bei Brieftauben und deren Auswirkungen unter besonderer Berücksichtigung des Heimfindevermögens // Zeitschrift für Tierpsychologie. Bd. 15. H. 3. S. 301–331.
Schmidt-Koenig K., 1990. The sun compass // Experientia. V. 46. № 4. P. 336–342.
Viehmann W., 1982. Interrelation of magnetic compass, star orientation, and the sun in the orientation of Blackcaps and Robins // Avian Navigation. P. 59–67. Proc. Life Science. Berlin, Heidelberg. New York: Springer Verlag.
Wallraff H.G., 1960. Does celestial navigation exist in animals? // Cold Spring Harbor Symposia on Quantitative Biology. V. 25. P. 451–461.
Weindler P., Baumetz M., Wiltschko W., 1997. The direction of celestial rotation influences the development of stellar orientation in young garden warblers (Sylvia borin) // Journal of Experimental Biology. V. 200. P. 2107–2113.
Weindler P., Beck W., Liepa V., Wiltschko W., 1995. Development of migratory orientation in pied flycatchers in different magnetic inclinations // Animal Behaviour. V. 49. P. 227–234.
Wiltschko W., Balda R.P., 1989. Sun compass orientation in seed-caching scrub jays (Aphelocoma coerulescens) // Journal of Comparative Physiology A. V. 164. P. 717–721.
Wiltschko W., Daum P., Fergenbauer-Kimmel A., Wiltschko R., 1987. The development of the star compass in garden warblers, Sylvia borin // Ethology. V. 74. P. 285–292.
Wiltschko W., Merkel F.W., 1966. Orientierung zugunruhiger Rotkehlchen im statischen Magnetfeld // Verhandlungen der deutschen zoologischen Gesellschaft. P. 362–367.
Wiltschko R., Nohr D., Wiltschko W., 1981. Pigeons with a deficient sun compass use the magnetic compass // Science. V. 214. P. 343–345.
Wiltschko W., Wiltschko R., 1972. Magnetic compass of European Robins // Science. V. 176. № 4030. P. 62–64.
Wiltschko R., Wiltschko W., 1980. The process of learning sun compass orientation in young homing pigeons // Naturwissenschaften. V. 67. P. 512–514.
Wiltschko R., Wiltschko W., 2015. Avian navigation: a combination of innate and learned mechanisms // Advances in the Study of Behavior. V. 47. P. 229–310.
Wiltschko R., Munro U., Ford H., Wiltschko W., 2008. Response to the comments by R. Muheim, S. Akesson, and J.B. Phillips to our paper “Contradictory results on the role of polarized light in compass calibration in migratory songbirds” // Journal of Ornithology. V. 149. № 4. P. 663–664.
Wiltschko R., Walker M., Wiltschko W., 2000. Sun-compass orientation in homing pigeons: compensation for different rates of change in azimuth? // Journal of Experimental Biology. V. 203. P. 889–894.
Wiltschko W., Wiltschko R., Keeton W.T., 1976. Effects of a “permanent” clock-shift on the orientation of young homing pigeons // Behavioral Ecology and Sociobiology. V. 1. P. 229–243.
Wiltschko W., Wiltschko R., Keeton W.T., 1984. The effect of a “permanent” clock-shift on the orientation of experienced homing pigeons in experiments in Ithaca, New York. Behavioral Ecology and Sociobiology. V. 15. P. 263–272.
Wolf-Oberhollenzer F., Kirschfeld K., 1994. Motion sensitivity in the nucleus of the basal optic root of the pigeon // Journal of Neurophysiology. V. 71. P. 1559–1573.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал