

Зоологический журнал, 2021, T. 100, № 4, стр. 462-480
Использование участка обитания самцом кабарги (Moschus moschiferus L.) в Центральном Сихотэ-Алине
В. А. Зайцев a, *, Д. А. Максимова b, Ю. В. Смирнов c, Н. В. Белотелов d, e
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
b Тихоокеанский институт географии ДВО РАН
690041 Владивосток, Россия
c Российский Новый университет
105005 Москва, Россия
d Вычислительный центр имени А.А. Дородницына Федерального исследовательского центра
“Информатика и управление” РАН
119333 Москва, Россия
e Московский государственный технический университет имени Н.Э. Баумана
105005 Москва, Россия
* E-mail: zvit09@mail.ru
Поступила в редакцию 15.12.2019
После доработки 23.04.2020
Принята к публикации 25.04.2020
Аннотация
В 2014–2015 гг. в хвойном лесу Сихотэ-Алинского заповедника изучали перемещения кабарог с целью определения связи использования участка обитания с факторами среды и социальным окружением. Методика включала многосуточные тропления, визуальные наблюдения за привыкшими к человеку кабаргами, радиослежение, методы изучения среды обитания. Применяли ГИС технологии, математическую обработку данных. Выделен вариант использования участка обитания, в котором самец кабарги длительное время поддерживал его внешние размеры (1.02–1.1 км2) и форму при перераспределении своей активности в общем центре активности участка обитания (39% площади). Наложение участков самцов-соседей на участок резидента достигло 64.7–65.5%, центра активности – 53.5%. Количество основных кормов (эпифитных лишайников) на участках обитания в разное время в 60–300 раз превышало потребности кабарог. В условиях обеспеченности пищей на всей площади распределение активности и ольфакторных меток, порядок переходов самца были, прежде всего, согласованы с социальным окружением: распределением самок и посещением самцами-соседями ядер активности. Среди контактов между самцами преобладали дистанционные. Распределение переходов кабарги включало широкие “полосы” обходов и центральную систему, которая объединяла локальные ядра активности. Данная структура согласована с формами рельефа и границами растительных сообществ. В центре активности участка обитания самец провел более 84% времени. Это связано с нередкими заходами самцов-соседей, что сопровождалось формированием резидентом модулей – функциональных элементов структуры, в которых кабарга обеспечивала свою жизнедеятельность и охрану в форме “дежурства”, что создавало заслоны от проникновения самцов-соседей.
Исследования пространственных структур, которые формируют животные во время своих переходов, актуальны в связи с выяснением отношений между видами, экологических связей в сообществах видов, механизмов, формирующих поведение особей в группировках. Среди немногих публикаций, содержащих сведения о пространственной и социальной структуре популяции, об использовании обитаемого пространства (участков обитания и др.) кабаргой в разных частях ареала, выделяются работы, выполненные с использованием разных способов тропления, в некоторых случаях дополнительно с визуальной регистрацией и наблюдениями (Устинов, 1967; Green, 1985; Зайцев, 1991, 2006, и др.; Приходько, 2003; Доманов, 2013; Zaitsev et al., 2015; и др.), радиослежения (Kattel, 1990; Harris, Cai, 1993; Максимова и др., 2014, 2014а; 2015; и др.). Единичны публикации об использовании кабаргой участка обитания в связи с условиями местообитания и социального окружения.
С целью изучения структуры популяции кабарги в разных условиях с 1975 г. организуются исследования на нескольких стационарных участках в Сихотэ-Алинском заповеднике и его окрестностях. С 1970 гг. мы применяли комплексную методику, состоящую из троплений и визуальных наблюдений за привыкшими к нам особями на подготовленных ключевых (стационарных) участках. С 2012 г. применяется также радиослежение (Максимова и др., 2014, 2014а; 2015; и др.).
С 2004 г. мы регулярно ведем исследования на участке “Нечетовкий” в бассейне р. Таежная у северной окраины Сихотэ-Алинского заповедника. В связи с замеченными особенностями использования местообитания, в сезон 2014–2015 гг. особое внимание было уделено изучению участка обитания одного из трех радиомеченных самцов кабарги. Данный участок находился в плотном окружении участками обитания других особей.
Цель данного исследования – изучить использование кабаргой участка обитания с несколькими, по нашему предположению, основным условиями среды обитания, включая и социальные, определяющие структуру участка обитания. Для того чтобы выяснить, как возникает адаптация пространственной и социальной структуры популяции к условиям обитания, необходимо определить: 1) параметры участка обитания (площади, форму) в разные периоды времени и 2) связи посещений кабаргой разных частей участка обитания с факторами среды, социальным окружением и характером контактов между особями, 3) изменения площади, осваиваемого особью участка с начала слежения за ней с целью выявления некоторой перманентной смещаемой площади (участка); 4) структурные элементы, объединенные переходами зверя, ядра активности разного пространственного масштаба на основе анализа частот регистрации, а также функциональных зон, модулей.
МАТЕРИАЛ И МЕТОДИКА
Описание природных условий участка “Нечетовский” приведено в более ранних публикациях (Зайцев, 2019 и др.). На части этого участка (11–12 км2) у окончания водоразделов при впадении нескольких ключей в р. Таежная, где проведены основные работы в 2014–2015 гг., вся площадь покрыта лесом с преобладанием кедровников (Pinus koraiensis) со значительным участием пихты (Abies nephrolepis). Кедровники в верхней части крутых (до 30°–50°) склонов южных экспозиций включают значительное количество дуба (Quercus mongolica). Часть северных склонов занята лиственничниками (Larix cajánderi) и кедрово-лиственничниковыми ассоциациями. На северо-восточных окончаниях водоразделов с крутизной склона до 35°–50°, занимающих не больше 10–11% общей площади, содержится повышенное количество пихты и ели (Picea jezoensis) с подростом, лиственницы. Вдоль нижней или средней части склонов, особенно, на южных экспозициях водоразделов простираются древние, более пологие, чем склоновые поверхности, террасы.
При общем снижении численности кабарги в заповеднике (Зайцев, 2006) на примыкающей к р. Таежная части “Нечетовского” участка, где проводили основные работы, к 2004–2015 гг., по данным учета троплением с окладом (Зайцев и др., 2013), поддерживалась повышенная концентрация ее населения – 14–18 особей на 10 км2.
Способы сбора данных заключаются в многосуточном троплении и радиопрослеживании привыкших и непривыкших к нам особей кабарги. Каждая кабарга имела особенности внешнего вида, их принадлежность к половозрастным группам определялась визуально по индивидуальным и известным групповым признакам, особенностям оставленных следов (Зайцев, 1991 и др.). В статье использованы номера радиомеченных самцов кабарги из публикации Максимовой с соавторами (2015). Самец кабарги М3 – “Гусар” (группа возраста 2–3 г.), чей участок обитания в 2014–2015 гг. (рис. 1) изучали особенно тщательно, был приучен к нам сеголетком (возраст 0–1 г.) в сезон 2012–2013 г. В апреле 2013 г. М3 и его соседи М6 (2–3 г.) и М5 (1–2 г.) были помечены ошейниками с радиопередатчиками Telonics, USA (150–152 МГц). Данные радиослежения использованы согласно программе совместных исследований Сихотэ-Алинского биосферного заповедника, Тихоокеанского института географии ДВО РАН, Общества сохранения диких животных (WCS, Нью-Йорк, США), Института проблем экологии и эволюции РАН.
Рис. 1.
Схема переходов самца М3 и заходов самцов-соседей на участок М3, схема расположения участков обитания самок F1, F2, F3 (штриховка) по данным тропления в декабре 2014 г.; stream – русло и днище долины ключей; wsh2–wsh7 – водоразделы; SS и NS – склоны южной и северной экспозиций; цифры и буквы в прямоугольниках: A–C – группы лежек и переходов (локальные ядра активности) на участке обитания М3; 1 – зона контактов М3 и М7; 2 и 3 в прямоугольниках – крутые северо-восточные окончания водоразделов, посещаемые многими особями (3 – у узкого днища двух ключей).
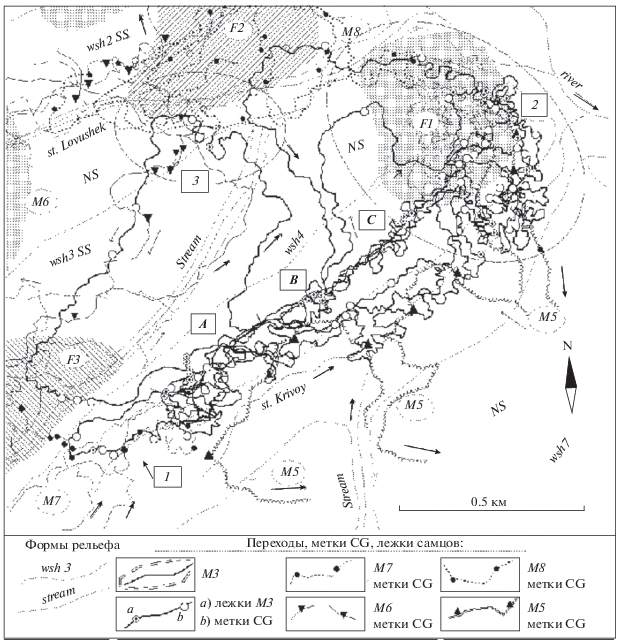
Организация системы слежения за кабаргами соответствовала базовой методике (Зайцев, Зайцева, 1980; Зайцев, 1991 и др.). С маршрутов, пролегающих не далее 200–500 м друг от друга вдоль ручьев, гребней и склонов водоразделов, вели поиск и учет следов зверей, заходили к месту, где в предыдущий день закончили тропление кабарги и продолжали тропить далее. С этих маршрутов обычно дважды в день: с 8–9 до 10 ч утра и в 16–17 ч определяли место расположения кабарог при радиослежении.
Мы тропили М3 в течение 5–6 дней, затем делали перерыв на один или несколько суток. М3, как и другие привыкшие кабарги, подпускал к себе после длительного (месяцы) перерыва в наблюдениях. В течение светлого времени суток, обычно, следили не только за М3, но еще за 1–3 соседними кабаргами, встречая следы этих соседних особей при троплении М3 и на маршрутах, окаймляющих их участки обитания. Соседей М3, которые частично привыкли к нашему многолетнему присутствию на стационаре, неоднократно встречали визуально после тропления. С близкого расстояния мы не наблюдали зимой лишь радиомеченного самца М5. Однако обширный северный склон, на котором, по данным радиослежения, в основном обитал М5, в сезоне 2014–2015 гг. другие кабарги не посещали, что было установлено контрольными обследованиями. После троплений какой-либо особи ее локализацию на выявленном участке и заходы на участок М3 контролировали при встрече следов на маршрутах, проложенных в окрестностях, с последующим троплением.
При троплении с GPS-Глонас-регистраторами и компасом следовали непосредственно по ходу кабарог. 16 троплений суточных ходов и их фрагментов совмещали съемку GPS-Глонас с регистрацией компасом смен направлений, измерений расстояний выверенными шагами по естественным составляющим траектории – векторам Va (по средним от 1.3 до 4.0 м на разных переходах) (Зайцев, 1991, 2002 и др.). Отмечали следы деятельности зверей, поеди лишайника, хвои и др., смены форм рельефа и другие особенности участка. Общий период слежения троплением охватывал снежный период с начала декабря (3.12.2014 г.) до марта (5.03. 2015 г.). За это время за кабаргами следили 38 дней.
Учеты лишайника (воздушно-сухая биомасса и число экземпляров (штук) обрывков талломов в пересчете на 100 м2) в доступной кабарге зоне до высоты 1–1.2 м над поверхностью снега или земли выполнены на площадках, трансектах 400 м2 или 900 м2. В конце января и в феврале 2015 г. проведен учет на 22 площадках, удаленных одна от другой на расстояние не менее 200 м. Учитывали также уклон склона, проективное покрытые (в долях от 1) верхнего древесного яруса и отдельно хвойного подроста по известной методике (Воронов, 1973).
Обработка и анализ данных. Данные слежения (с GPS-регистраторов и др.) кабарог транспортированы на космический снимок в программе NexQGIS. Затем схемы траекторий, полигонов частот посещений конвертировали в программу MapInfo 7.2, в которой проведена обработка и анализ пространственных данных.
При троплении М3, в том числе 21 суточного перехода и 11 их фрагментов, проведен учет следов деятельности (меток, лежек и др.) на 48 км хода в ноябре–декабре и около 55 км в январе–марте. Кроме того, в этом сообщении приведена часть данных троплений и радиослежения по перемещениям соседей М3.
Способы определения границ, соответственно размеров, формы участка обитания, выделение зональных областей в его пределах относятся к обычным методическим проблемам при применении разных способов слежения и анализа данных (Hayne, 1949; Powell, 2000; Kie et al., 2010; Powell, Mitchell, 2012; и др.). При использовании троплений контуры участка обитания, наложения участков соседних особей друг на друга проведены по внешним траекториям особей, окаймляющим участок, со сглаживанием интервалами не меньше 50–100 м небольших их изломов (Зайцев, 1991, 2014). В 2014–2015 гг. мы не отметили случаев, когда М3 покидал свой участок при бегстве от хищников (эпизодические, случайные выходы). Перемещение самца после встречи с наблюдателем, обычно, укладывалось в интервал не более 150 м. В масштабе участка обитания это не имело значения в выяснении его внутренней структуры. Такие же переходы самец совершал и при появлении кабанов, которые перерывали кедровника с дубом в верхней части южного склона, и которые проходящих (особенно в конце февраля 2015 г.) по средней части склона.
Измерение площади, окаймленной переходами М3, проведены на схемах после каждого тропления за два периода зимы 2014–2015 г. В первые день-два с начала слежения переходы кабарги не всегда замыкались конкретной площадью. При отсутствии протяженных обходов участка прирост площади в эти дни определен в качестве среднего значения ее изменения обычно за два дня.
При зонировании участка обитания мы использовали представления о центрах или ядрах активности (“centre of activity” – Burt, 1943; фактически, синонимы: ядро – “kernel of home range” – Worton, 1989; центр, ядро, сердцевинная зона – “core area” – Kaufmann, 1983), которые в общем случае выделяются на основе более частых посещений или долгого пребывания животного в определенном месте, наличия убежищ, лежек. Для исключения излишней терминологии мы использовали данное определение как для выделения “общего центра (ядра)” активности” на участке обитания, так и для обозначения “локальных ядер” – мест с наибольшей частотой посещений в общем центре или вне его. В рамках статьи термин “центр активности” применяется, в основном, к общей зоне повышенного числа посещений на участке обитания, “ядро” – к местам локального увеличения частот.
Определение ядер, центров активности предполагает использование в основном статистических способов для их выделения, основанных на построении полигонов частот регистрации зверя, плотности траекторий в разных местах и др. В случае радиослежения данные полигоны определены нами на основе параметрической модели Вортона (Worton, 1989) с обработкой данных ГИС в программе R Statistical Software (Максимова и др., 2014 и др.). Было получено 39 точек локации М3 в ноябре–декабре и 69 точек в январе–марте. Полигоны модели с частотами 95% принимали за размер участка обитания, 50, 64 или 35% – за ядра активности. Выбор процента для полигона согласовывался с данными троплений, как способа наиболее подробной и точной регистрации многосуточных переходов. При анализе данных тропления, на схемы переходов кабарги накладывалась сеть с ячейками размером 100 × 100 м (1 га) и подсчитывалось число заходов кабарги в каждую ячейку. Для анализа и визуализации данных инструментами “wafer” в программе Statistica 8. Software моделировали поверхности полигонов частот посещений.
Для интерпретации связи поведения особи с распределением частот посещений мы используем определение “модуль”. В отличие от ядер модуль имеет значение функциональной части, элемента структуры участка обитания, сочетающего разные активности поведения, обеспечивающие жизнедеятельность особей вместе с эффективной охраной локальных территорий. Выделение модулей проведено этологическим способом по характеристике траекторий и поведения особи.
Мы выделили основные паттерны поведения, включающие сочетание нескольких поведенческих активностей с доминированием обычно одной из них на разных фрагментах траектории. К ним относятся паттерны: а) сбора пищи (по поедям) и отдыха (по лежкам), нередко на непротяженных траекториях со множеством поворотов. На переходах по периферийной части участка обитания у взрослых самцов преобладают: б) активное патрулирование при большом числе меток экскретом комплекса хвостовых и надхвостовых желез (метки CG и небольшие кучки экскрементов) с регистрацией немногих поедей; в) комплексные обходы (патрулирование), сочетающее активный сбор пищи с маркировкой. При активном сборе пищи кабарга движется часто коротким шагом (до 25–35 см) по траектории со множеством поворотов. Комплексное и, тем более, активное патрулирование происходит длинным шагом (до 40–45 см), иногда переходящим на рысь. Кроме этого, выделяются фрагменты г) обходов длинным шагом, рысью и прыжками мест, редко посещаемых кабаргами, с очень небольшой частотой маркировки (Зайцев, 1991).
Для выделения свойств переходов зверей в разных местах и режимах деятельности использовали векторный анализ, основанный на выделении групп векторов, обобщающих естественные фрагменты пути разной протяженности (Зайцев, 1991, 2002 и др.). По схемам определены расстояния |Vsl| между дальними точками каждого суточного перехода особи вдоль основной линии пролонгации хода. Размеры модулей установлены по длинам радиус-векторов Rmod из геометрического центра группы лежек или от конкретной лежки до наиболее удаленных от них точек замкнутых переходов.
Данные с 16 площадок учета (9 в центре активности участка обитания) использованы для определения связи факторов среды с переходами. Среди факторов и условий, вероятно, влияющих на частоты (fi) посещений кабаргой разных частей участка и ячеек 1 га в их сети, взяты следующие показатели: 1) число лежек в ближнем к ячейке учета локальном ядре активности (показатель регулярности посещений); 2) количество основной пищи в зимний период – кустистого лишайника (родов Usnea, Evernia и др., г/100 м2; воздушно-сухая биомасса); 3) число объектов пищи (штук/100 м2) – обрывков лишайника и веточек с лишайником; 4) угол уклона склона (град) в месте учета; 5) проективное покрытие (в долях от 1) и особенности хвойного подроста у площади учета. Проведена также 6) оценка влияния социального окружения по характеру посещений соседями мест наложения участков обитания. Определяли показатель синхронизации Сс = fvis1/fvis2, где fvis1 – число совпадений заходов двух самцов в сутки в зону наложения участков (Сс рассчитывался для первых, вторых суток после захода соседа); fvis2 – число заходов изучаемого самца.
Для характеристики сезонных изменений в структуре группировок и участка обитания кабарги мы использовали периодизацию, основанную на изменении подвижности особей и других активностей в разные периоды годового цикла (Зайцев, 1991; Максимова и др., 2014). Для снежного периода года, когда было применено тропление, выделено два периода: 1) ноябрь–декабрь – период гона и его окончания, повышенной двигательной активности; 2) январь–март – период пониженной двигательной активности (с января к февралю), небольшой частоты мечения участка обитания специфическими метками.
Математическая обработка. Статистические расчеты и анализ траекторий проведены средствами программ Statistica 8, Statgraphycs Plus, Python Numpy Scipy, EasyTrace. Использовали регрессионный, факторный анализ методом главных компонент (PC), для переменных в центре активности участка обитания с нормализованной варимаксной ротацией. Исключение влияния разных шкал осуществлено Z-преобразованием: ${{Z}_{i}} = ({{x}_{i}} - \bar {x})$/Sd; где ${{x}_{i}}$ – значение члена i переменной; $\bar {x}$ – среднее переменной; Sd – среднее квадратичное отклонение. Другие обозначения в тексте: rs – коэффициент корреляции Спирмена; Me – медиана; CV% – коэффициент вариации; W-test – Wald-Wolfowitz Runs test.
Анализ траекторий в модулях и между ними проведен методом непараметрического оценивания, который используется в задачах восстановления плотности неизвестного распределения, в том числе в анализе изображений (Shen et al., 2017; Lychkov et al., 2018; Maltsev et al., 2018; Dmitriev et al., 2019). Мы использовали один из вариантов данного оценивания – метод оценки ядерной плотности. Суть метода заключается в том, что для выборки случайных величин $\left( {{{x}_{1}},{{x}_{2}},{{x}_{3}}, \ldots {{x}_{n}}} \right)$ определяется ядерная функция $K\left( x \right)$ и на ее основе строиться непрерывная функция плотности ${{f}_{h}}\left( x \right)~$ (Parzen, Rosenblatt, 1956). Под “ядром” в данном случае подразумевается вещественная неотрицательная функция, обладающая следующими свойствами: $\int_{ - \infty }^{ + \infty } {K(x)dx} = 1;~K( - x) = K(x).$ В качестве ядерной функции мы использовали Гауссово распределение. Параметр h, как ключевой свободный параметр “сглаживания” в построении итогового распределения, определен, исходя из эмпирического критерия Скота (Scott, 2010): $h \approx \frac{1}{{\sqrt[{d\, + \,4}]{N}}}$, где d – количество размерностей случайной величины, N – размер выборки.
Вначале проводилась векторизация схем треков в программе EasyTrace с переводом в табличные данные формата csv. Один пиксель изображения соответствовал 0.85 м реального хода зверя. Каждый трек-вектор, входящий в общий массив данных, описывался упорядоченной парой координат $(x_{i}^{j},y_{i}^{j})$, где верхний индекс j – номер трека, нижний i изменяется от 1 до N j – числа узлов, формирующих j-трек. Траектории представлены совокупностью векторов: длин $\Lambda _{i}^{j}~$ и углов $\varphi _{i}^{j}~$ отклонения между ними. $\Lambda _{i}^{j}$ соответствовали реальным длинам векторов |Va| между пунктами – узлами с заметными поворотами пути.
РЕЗУЛЬТАТЫ
В сезон 2014–2015 гг., по результатам тропления, участок М3 соседствовал с местами обитания семи кабарог-резидентов (рис. 1, 2): самцами М7 (старше 3 лет), М6 (возрастная группа 2–3 года), М8 и М5 (группа возраста 1–2 года), самок F1 (старше 1.5 лет), F2 (>1.5 лет), F3 (1–2 года). Трое суток (27–29.12.2014 г.) у места соседства М3 и М7 (1 на рис. 1) обитала восьмая кабарга – самка, которая затем перешла вверх по водоразделу. В течение всего снежного сезона состав особей-резидентов в окрестностях участка М3 не менялся.
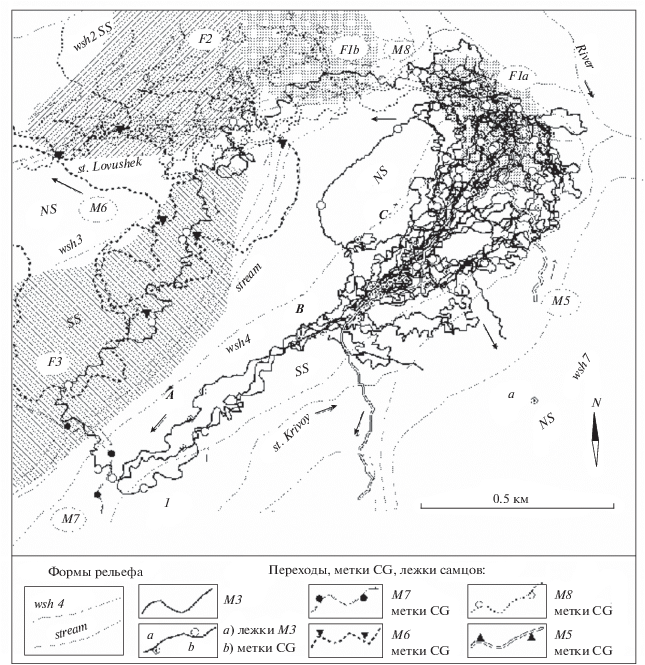
Рис. 2.
Схема переходов самца М3 и кабарог-соседей, расположения участков обитания самок F1, F2, F3 (штриховка) по данным тропления с января до 05.03.2015 г.; а – пункт регистрации М3 при посещении соседнего водораздела вслед за заходом М5.
Внешние параметры участка обитания. Общая площадь участка обитания М3, определенная троплением за два периода зимы (рис. 1, 2; табл. 1), составила Sshr = 1.21 км2, при радиослежении – 1.3 км2. 82% площади в декабре и 79.5% в январе–марте располагалось на горных склонах, из них 6 и 8%, соответственно, на северо-восточном склоне, 32.4 и 31.6% – на южном склоне водораздела wsh4 (рис. 1, 2), 16.2 и 16.3% – wsh3, 18.2 и 19.5% – на днищах долин ключей. Северные склоны водораздела wsh4 (у М3 27.2% в декабре и 24.6% площади в январе–феврале) в эту зиму все кабарги посещали редко. Их переходы пролегали обычно у небольшого ключа, куда кабарог привлекал опад лишайника. Участок М3 простирался вдоль водораздела при соотношении наибольших расстояний вдоль склона (1.55 и 1.56 км) и поперек склона (0.84 и 0.8 км) 1: 1.87 в декабре и 1: 1.95 в феврале–марте.
Таблица 1.
Параметры участка обитания М3 (площадь SM3(Month), км2, периметр PM3(m) (км) и наложение (% OVi, км2) на участок участков кабарог-соседей в зимний сезон 2014–2015 г.
| Период | Показатель для М3 | OV♂-перекрытие (Sov, P, %) с самцами | OV♀ с самками | ||||||
|---|---|---|---|---|---|---|---|---|---|
| М7 | М6 | М8 | М5 | ΣOV♂ км2 | F1 | F3 | |||
| Декабрь–середина января | SM3(D) км2 | 1.10 | 0.19 | 0.17 | 0.15 | 0.21 | 0.72 | 0.17 | 0.04 |
| PM3(D) км | 4.25 | 1.84 | 2.13 | 2.01 | 2.49 | 1.42 (1.43) | (0.83) | ||
| % OVhr | 65.5* | 17.3 | 15.5 | 13.6 | 19.1 | (15.5%) | (≈4%) | ||
| Середина января–март | SM3(F) км2 | 1.02 | 0.02 | 0.18 | 0.17 | 0.29 | 0.66 | 0.11 | 0.13 |
| PM3(F) км | 4.45 | 0.3 | 2.0 | 2.6 | 2.4 | 1.22 | 2.13 | ||
| % OVhr | 64.7* | 1.8 | 17.6 | 16.7 | 28.4 | 11% | 12.7% | ||
Общий процент наложения (% OVmales) на участок М3 участков самцов-соседей (табл. 1) мало менялся в первую и вторую половину снежного периода. Участок самки F1, а также F2 и F3 имел характер “вложения” в участок М3 и самцов-соседей. Участок F1 (0.17 км2; рис. 1) перекрывался заходами М3 в середине декабря на 88%, в конце декабря – на 93% (участок F1a в 0.1 км2; рис. 2). Со второй декады января самка ушла на соседний водораздел, где также в основном на северо-восточном склоне обосновалась на участке F1b (≈0.15 км2), расположенном на участке М8. В этот период при редких заходах М3 встреч его с F1, как и с F2, нередко посещающей днище ключа в декабре и феврале, не отметили. К месту обитания F3 самцы М3, М7, М6 заходили зимой нечасто (отмечено 3 захода М3 за 38 сут), но пересекали ее участок (наложение 0.21 км2) в декабре и феврале–марте в разных местах.
При значительных показателях перекрытия площадей участков самцов, в некоторых местах происходило пересечение переходов трех самцов. Такие места расположены на северо-восточных окончаниях водоразделов с прилегающими площадями (эллипсы с цифрами 2 и 3 на рис. 1). Также в окрестностях седловины 1 наблюдалось перекрытие участков М3, М7 и М6. Зону 3 наиболее часто посещали М8, М6, а также самка F2, затем и F3. Присутствие и постоянное обитание самок отмечено во всех этих местах.
Изменение размера участка обитания М3 при последовательном троплении. Измерения площади, обходимой М3 после каждого тропления, показали на ее увеличение до предельного значения в каждый период зимы (табл. 1) по зависимости, приближающейся к логарифмической (рис. 3). Также оказалось возможным выделить интервалы времени в сутках, в течение которых обойденная кабаргой площадь после короткого периода значительного прироста $\left( {IA} \right)~$ достигала сравнительно стабильных размеров.
Рис. 3.
Изменение размера участка обитания самца М3 при последовательных троплениях в декабре 2014 г. (А) и в январе (с 11.01) по 5 марта 2015 г. (В).
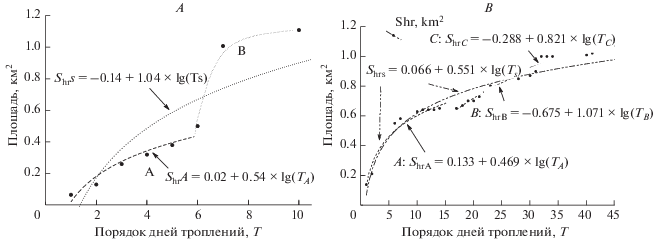
В декабре 2014 г. после семи дней слежения участок достиг больших размеров (волны А и В; рис. 3А), которые, в основном, поддерживались всю остальную часть снежного периода. В январе–марте значительный прирост участка М3 произошел в течение 10 суток с начала слежения за ним (рис. 3В). Затем произошла стабилизация площади, но еще через 6–7 сут вновь прирост и относительная стабилизация к 24–27 суткам (т.е. еще через 8–10 сут) с начала слежения. Затем площадь вновь увеличилась за 2–3 суток и стабилизировалась вплоть до 40–41 сут, считая перерывы в троплении, которые, однако, не маскировали основные этапы изменения площади. Среднесуточный ее прирост за весь период составил 0.023 км2 (Me = 0.02, Sd = 0.025, n = 20). При общем увеличении площади с начала наблюдений до Shr = 1.21 км2 в марте, происходило ее смещение и перераспределение активности (посещений) самца.
Распределение посещений разных частей участка и ядра активности. Обработка инструментами “wafer” (Statistica) моделирует связь числа посещений fi…j ячеек сети площадью 1 га c процентом площадей полигонов частот Sf % от общей площади участка (Shr) за всю зиму по зависимости: Sf % = = 96.27 – (94.74 lg(fi)) – 5, r2 = 0.852, p = 0.00002. Полученная карта поверхности частот (рис. 4) свидетельствует об очень неравномерном использовании М3 участка обитания. На большей части (>65%) ячеек отмечено не больше 1–2 заходов за зиму. На части северного склона водораздела wsh4 (8 на рис. 4) с произрастающими здесь лиственничниками и кедровниками с лиственницей, с глубоким (до 50 см и выше) снежным покровом, заходы М3 не были замечены, как и заходы других особей.
Рис. 4.
Распределение частот посещений М3 участка обитания зимой 2014–2015 гг.; на рис. вверху слева – шкала частот и проценты частот для площади полигонов; а – ядра активности участка (64%) по данным радиослежения в декабре, b – ядро (35%) в январе–марте, c – контуры участка М3 за весь период наблюдения; А–I – локальные ядра активности, формируемые М3 с декабря по март; 1 – место заходов самца М7 в декабре; 2 – узкая полоса переходов М3 по северному склону с маркировкой; 3, 4 – места, посещаемые М5 со стороны ключа Кривой; 5, 6 – место расположения в декабре участка F1 и нередких заходов М8; 7 – участки протяженных патрулирований М3, а также М6 южного склона; 8 – редко посещаемый кабаргами в сезон 2014–2015 гг. северный склон.
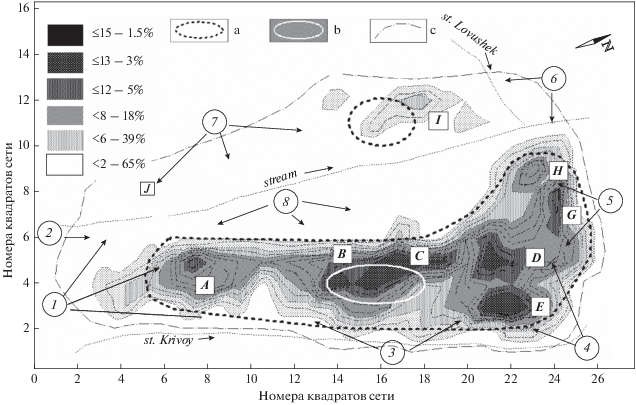
Основные переходы, места отдыха и сбора пищи М3 располагались вдоль южного склона и на части днища долины ключа Кривой (от А до E на рис. 4), а также на северо-восточном склоне и примыкающей к нему части водораздела (E–H–6). Эти места, каждую ячейку сети которых, по данным тропления, М3 посетил не менее 4–6 раз за зиму, составили общий центр (ядро) активности, занимающий около 39% (0.43 км2) площади участка обитания в декабре–январе. Приблизительно сходную конфигурацию имел и 64% полигон частот радиорегистрации М3 (0.42 км2; рис. 4) в ноябре–январе. Из 38 суток общего времени слежения М3 провел в общем центре 32 полных суток и в течение 6 суток совершал обходы периферии, возвращаясь в центр активности. Таким образом, общее время нахождения М3 в центре активности превысило 84%.
Полигон с частотой посещений 50%, по данным троплений, в этой модели занимал 17–18% площади участка. Площади с бóльшей частотой имели мозаичное распределение, указывающее на формирование локальных ядер в общем центре участка обитания. Локальные ядра активности В–С кабарга посещала наиболее часто всю зиму.
Влияние факторов среды на распределение посещений разных мест на участке обитания. Среди переменных, используемых в анализе (табл. 2, 3), лишь биомассу лишайника (“Лишайник, г/100 м2”) и число его обрывков в доступной зоне (“Лишайник, штук/100 м2”) можно отнести к существенно связанным между собой. Однако эта взаимосвязь относительна, так как лишайник может распределяться в мелких обрывках по подстилающей поверхности или сосредоточиваться на упавших деревьях. Тем не менее, на площадках учета корреляция между этими переменными достигала rs > 0.99. В пределах участка обитания высокие значения соотношений отмечены между числом заходов в ячейки 1 га и проективным покрытием хвойных – rs = 0.71 (р < 0.05). Корреляция между другими переменными была меньше (0.05–0.4; р > 0.05). В общем центре активности число заходов в ячейки значительно согласовалось с числом лежек в ближнем локальном ядре (rs = 0.80, р < 0.05), с показателем проективного покрытия (rs = 0.81). С последним связано и число лежек в локальном ядре (rs = 0.67). Как на всем участке (rs = 0.22, р < 0.05), так и в центре активности (rs = –0.05, р < 0.05) отмечена лишь локальная связь числа посещений ячеек с биомассой и числом объектов пищи. Данная связь выявлялась в случаях, когда кабарга начинала более активно посещать упавшие деревья, например, вдоль ключа Кривой, где биомасса лишайника достигала 560–590 г/га. Также М3 и его соседи нередко посещали северо-восточные склоны и днища малых ключей с большим количеством лишайника (5.8 г/100 м2, n = 3), сбитого ветрами. В кедровнике и лиственничнике северного склона, в которые кабарги заходили нечасто, лишайника было лишь немного меньше, чем в ядре активности М3: 3.1 г/100 м2, CV = = 27.4%, n = 5.
Таблица 2.
Число заходов кабарги в ячейки 1 га, количество доступного лишайника в январе–феврале 2015 г., значения других переменных и их Z-преобразований (в скобках) на площадях учета в пределах участка обитания М3
| № | Значение переменных и Z-преобразования | |||||
|---|---|---|---|---|---|---|
| число заходов (NV) в ячейку | лежек в ближнем ядре | лишайник, г/100 м2 | лишайник, штук/100 м2 | угол склона, град | проективное покрытие (от 1) | |
| 1 | 7 ( 0.13) | 10 (1.01) | 7 (1.87) | 63 (1.85) | 18 (–0.32) | 0.9 (0.88) |
| 2 | 4 (–0.55) | 0 (–0.75) | 4.8 (0.22) | 43 (0.19) | 2 (–1.61) | 0.7 (–0.29) |
| 3 | 5 (–0.32) | 9 (0.84) | 3.9 (–0.45) | 35 (–0.47) | 12.5 (–0.76) | 0.6 (–0.88) |
| 4 | 14 (1.70) | 21 (2.94) | 3.7 (–0.61) | 33 (0.63) | 20 (–0.16) | 1 (1.46) |
| 5 | 10 ( 0.80) | 4 (–0.04) | 4.9 (0.29) | 44 (0.28) | 16.5 (–0.44) | 1 (1.46) |
| 6 | 7 (0.13) | 2 (–0.39) | 5.2 (0.52) | 47 (0.53) | 28 (0.49) | 1 (0.29) |
| 7 | 13 (1.48) | 2 (–0.39) | 5.4 (0.67) | 49 (0.69) | 5 (–1.36) | 0.8 (0.88) |
| 8 | 12 (1.25) | 9 (0.84) | 4.3 (–0.6) | 39 (–0.14) | 8 (–1.12) | 0.9 (1.46) |
| 9 | 12 (1.25) | 6 (0.31) | 5.2 (0.52) | 47 (0.53) | 38 (1.29) | 1 (0.29) |
| 10 | 4 (–0.55) | 1 (–0.57) | 2 (–1.89) | 18 (–1.87) | 18 (–0.32) | 0.8 (–1.46) |
| 11 | 5 (–0.32) | 1 (–0.57) | 2.6 (–1.44) | 23 (–1.46) | 17 (–0.39) | 0.5 (–0.29) |
| 12 | 2 (–1) | 0 (–0.75) | 3.8 (–0.53) | 34 (–0.55) | 34 (0.97) | 0.7 (–1.46) |
| 13 | 1 (–1.23) | 0 (–0.75) | 3.2 (–0.99) | 29 (–0.96) | 38 (1.13) | 0.5 (–0.88) |
| 14 | 1 (–1.23) | 0 (–0.75) | 4.1 (–0.31) | 37 (–0.30) | 43 (1.69) | 0.7 (–0.29) |
| 15 | 2 (–1) | 0 (–0.75) | 5.7 (0.89) | 51 (0.86) | 35 (1.05) | 0.6 (–0.88) |
| 16 | 4 (–0.549) | 3 (–0.22) | 6.4 (1.42) | 58 (1.44) | 20 (–0.156) | 0.7 (–0.29) |
Таблица 3.
Значения Z-преобразований переменных (табл. 2) на площадях учета в общем центре активности участка обитания самца кабарги
| № | Значение переменных и Z-преобразования | |||||
|---|---|---|---|---|---|---|
| число заходов (NV) в ячейку | лежек в ближнем ядре | лишайник, г/100 м2 | лишайник, штук/100 м2 | угол склона, град | проективное покрытие | |
| 1 | –0.64 | 0.47 | 2.11 | 1.94 | 0.14 | 0.28 |
| 2 | –1.45 | –1.11 | –0.13 | –0.95 | –1.27 | –1.13 |
| 3 | (–1.19 | 0.32 | –1.05 | –1.67 | –0.35 | –1.83 |
| 4 | 1.27 | 2.21 | –1.25 | 0.35 | 0.31 | 0.99 |
| 5 | 0.18 | –0.47 | –0.03 | –0.37 | 0.01 | 0.99 |
| 6 | –0.64 | –0.79 | 0.28 | 0.21 | 1.01 | –0.42 |
| 7 | 1 | –0.79 | 0.48 | 0.06 | –1.00 | 0.28 |
| 8 | 0.73 | 0.32 | –0.64 | –0.08 | –0.74 | 0.99 |
| 9 | 0.73 | –0.16 | 0.28 | 0.49 | 1.89 | –0.42 |
Факторный анализ (PC) в пределах всего участка обитания (табл. 2) показал, что число заходов кабарги в ячейки 1 га, число лежек в ближнем ядре активности и проективное покрытие темнохвойных связаны с общим фактором, объясняющим 54.5% общей дисперсии (рl < 0.70) (табл. 4). Анализ для тех же переменных (табл. 3) в общем центре активности (от А к Н; рис. 4) указывает на связь с двумя факторами (табл. 4). Первый из них обусловил 42.9% общей дисперсии, второй – 24.3%. Общие условия связывали число заходов в ячейки, число лежек в ближнем ядре с проективным покрытием (рl < 0.70). Второй фактор определял распределение количества лишайника на площадках учета.
Таблица 4.
Факторные нагрузки для пяти переменных в пределах всего участка обитания (без варимакса) и в общем центре активности (нормализованный варимакс)
| Переменные | На всем участке | В общем центре активности | |
|---|---|---|---|
| фактор | фактор 1 | фактор 2 | |
| Число заходов | 0.921* | 0.905* | 0.057 |
| Лишайник, г/100 м2 | 0.369 | –0.221 | 0.892* |
| Лежки | 0.775* | 0.713* | –0.337 |
| Угол склона | –0.540 | 0.299 | 0.502 |
| Проективное покрытие | 0.920* | 0.824* | 0.220 |
Связь переходов по участку обитания с формами рельефа. Контуры участка обитания, определенные по внешним траекториям М3, во многом соотносились с основными линейными ориентирами рельефа и растительности. Вдоль подножия северо-восточного склона (Е–H; рис. 4), граничащего с лиственным лесом у реки, переходы далее пролегали вдоль днища долины ключа и подножия водораздела wsh2 (от H до I), в кедровнике с пихтовым подростом и лиственницей нижней части южного склона притока этого ключа (от I до J; рис. 1, 2, 4). Здесь обходы охватывали в разное время полосу шириной до 200–250 м. В западной части участка обитания у седловины водораздела wsh4 формируется узкая полоса переходов по северному склону (2 на рис. 4). М3 каждый раз оставлял в этой зоне перекрытия с участком М7 2–4 метки экскретом хвостовой железы (CG) на 80–110 м хода. На южном склоне wsh4 расширенная полоса переходов охватывала небольшое пятно лиственничника и далее кедровник вдоль русла ключа Кривой и террас.
Вдоль террасы южного склона через весь участок обитания пролегала полоса переходов – центральная их система (рис. 1, 2), которая объединяла локальные ядра А–G (рис. 4) в общем центре активности участка обитания. В кедровнике вдоль террасы почти сплошь произрастает плотный (проективное покрытие 0.8–1) высокий подрост, “жердняк” пихты, который распространен и ниже по более крутому склону с участками ветровала. В просторный кедровник с дубом, редкой пихтой и лещиной (Córylus sp.) верхней части склона М3 заходил, в основном, со второй половины февраля. В это время у гребня появились проталины, со 2.03–5.03.2015 г. обширные покопки кабанов, на которых кабарга собирала пищу. Вблизи более явно формируются ядра D и Е, с лежек которых самец имел возможность контролировать крутой склон, на который заходили соседи.
От ядра С к окончанию водораздела терраса постепенно исчезает, и на схеме (рис. 1, 2) сумма переходов М3 приобретает веерообразный вид, заходя на северо-восточный склон и выходя из него. От А, особенно В и в феврале–марте от С к Е, самец следовал обычно по выступам склона в долину ключа Кривой и обратно, сформировав широкое направление, сужающееся по выступу склона в феврале-марте у С до ≈30–40 м. С января выделяются ядра G и H, куда чаще стал заходить М8, на месте которых в декабре обитала самка F1.
Организация переходов в связи с социальным окружением. Наиболее протяженные суточные переходы М3 представлены обходами периферии или контролированием части участка, который начали активнее посещать соседи. Из шести его суточных переходов в декабре 2014 г., два – 4.5 км и 3.3 км – представляли собой подобные обходы с возвращением в общий центр активности в те же сутки. Обход в ночь с 27.12 на 28.12.2014 г. в ответ на заход М5 со стороны ключа Кривой достиг 3.78 км. В центре активности за сутки М3 проходил 2.5 км, 1.45 км, 2.14 км ($\bar {l}~$ = 2.0, Sd = 0.534), меньше, чем при дальних обходах: 3.87, Sd = = 0.590, n = 3. С середины января до начала марта длина суточного хода составила 2.08 км, Sd = 0.927, n = 15. С января эта длина (2.86 км, n = 4) уменьшилась в феврале, когда М3 ограничил заходы в зону контактов с М7, до 1.54 км, d = 0.43, n = 9. Но в марте с таянием снега наблюдали переходы на расстояние до 2.6 и 4.01 км.
В декабре самцы-соседи нередко посещали (отмечено 12 за 10 дней) две противоположные окраины участка обитания М3 вдоль водораздела (рис. 1, 2, 4). Из шести суточных переходов М3 один (3–4.12.2014 г.) охватывал только восточную часть участка от С почти до I с возвращением в ядро С на дневные лежки. Этот обход М3 совершил после захода М8, посетившего место обитания самки F1 прошлой ночью и повторившего подобный заход 5–6.12 и 28–29.12. В другие сутки М3 проходил вдоль всего участка между зонами контактов с М7, М8, а также М5 со стороны ключа Кривой. Наиболее часто (4 из 6) он начинал и заканчивал суточные переходы в группах лежек А и В, вблизи которых присутствовал М7. Однако за ночь М3 успевал обойти и места обычного присутствия М8 и М5, 27–28.01.2014 г. – дважды за сутки, переходя и днем. Длины векторов |Vsl| вдоль южного склона и ключа между дальними точками суточного хода составили от 740 до 1460 м ($\overline {{{V}_{{sl}}}} = 1080~\,{\text{м}}$; Sd = 310.6, n = 6).
С середины января 2015 г. М3 еще посещал обычную зону контактов с М7. Однако при отсутствии последнего всю остальную часть зимы, сместил свою активность (табл. 5; χ2 = 42.23, p < < 0.001) в восточную часть участка (рис. 2). Длины векторов |Vsl| составили от 230 м при многократных переходах между В и С 12.02, 18.02 (сбор пищи и отдых) до 773 м ($\overline {{{V}_{{sl}}}} ~$ = 625 м; Sd = 163.6, n = 10) и отличались от |Vsl| в декабре (W-test): z = 2.02, p = = 0.04. Обходы периферии со стороны участков М6 и F3 стали редки. Один из них отмечен ночью 24–25.01.2015 г., когда М3 от ядра Н прошел к I и J и вернулся к А и В (|Vsl| = 1430 м). На следующие сутки он обитал в окрестностях ядер А и В, посетив часть участка, на которую заходил ночью и М7. Обычно суточный ход в этот период начинался от ядра С (8 из 13), В (4), D (1), заканчиваясь также в этих ядрах.
Таблица 5.
Посещения (%) самцом М3 локальных ядер активности (рис. 4), заходы к группам лежек, в разные периоды зимы 2014–2015 гг.
| Период | Посещения ядер | n | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| А | В | С | D | Е | G | H | I | J | ||
| Декабрь–январь | 13.6 | 21.6 | 13.4 | 10.9 | 13.5 | 10.7 | 10.8 | 2.8 | 2.8 | 35 |
| Февраль–март | 3.8 | 14.1 | 23.1 | 11.5 | 19.2 | 17.9 | 7.7 | 1.4 | 1.3 | 79 |
Показатель синхронности Cc = 0.6 для посещений М3 зоны перекрытия в туже ночь, что и М7, определен в декабре, на вторые сутки после захода М7 – Cc = 0.2. Самцы ложились на дневные лежки иногда (29.12) на расстоянии 140 м друг от друга, каждый со стороны своего участка. М7 заходил и к ядру А. Следования вдоль следа соседа одним из них не отмечали. Посещения М7 данных мест были согласованы с присутствием самки, которая затем ушла выше по водоразделу. В январе-марте М7 редко посещал эти места. Тем не менее, из трех обходов M3 один ночью совпал с заходом М7 (Сс = 0.3).
Синхронность обходов М3 с М8 и М5 в декабре была меньше – Сс = 0.5, и только для вторых суток после их заходов. Однако в январе–марте “дежурство” М3 в окрестностях ядер С, D, E, G способствовало увеличению Сс до 0.6 для вторых суток и 0.3 для той же ночи при заходах М8, 0.5 и 0.3 для М5, соответственно. Для М3 и М6, чьи основные центры активности располагались друг от друга через водораздел (рис. 1, 2), различие в сроках посещений южного склона водораздела wsh3 достигали трех суток и больше.
Между всеми самцами мы не отметили явно агрессивных действий. Обычный дистанционный контакт М3 с М8 показывает случай, произошедший в ночь с 25 на 26.02.2015 г. В начале ночи М8 пересек прыжками и длинным шагом северо-восточный склон (в области 2 на рис. 2). Зайдя ночью с южного склона, М3 прошел его след (у G и H; рис. 4), но через ≈150 м вновь вышел к нему и многократно пересекая след шагом со сбором пищи и маркировкой (2 метки GC на 180 м), свернул вверх по склону и лег в 160 м от лежки соседа. Следующей ночью М8 ушел обратно по лиственничнику северного склона.
Между дневными лежками М3 и М5 в зоне перекрытия расстояния были не меньше 100 м. Преследуя этого соседа, М3 шел шагом, иногда прыжками, многократно пересекая след, иногда непосредственно по следу. М8 и М5 уходили при приближении М3. После их ухода М3 нередко еще сутки-двое посещал окрестности этого места, инспектировал периферию после заходов М8 (3–4.12.2014 г. и др.) или при заходах М5 сериями обходил с комплексным патрулированием периферии, переходил через ключ Кривой к водоразделу wsh7 (рис. 1, 2).
Посещая место обитания самки F1, М3 неоднократно следовал вдоль ее следа. Самка также переходила вдоль следа самца. Они устраивались недалеко друг от друга на лежки, часть из которых в разное время использовали обе кабарги. На ночных переходах 5–6.12. были замечены следы их встречи с “топтанием”, обходами, четкими отпечатками копыт одной особи за отпечатками копыт другой, что с большой вероятностью указывает на копуляцию. При обходе периферии М3 задерживался в месте обитания F3 (у J, рис. 4, 5), лежал и собирал пищу.
Рис. 5.
Модульная структура участка обитания самца М3 в 2014–2015 г.; 1 – модули A–K (на рис.): группы лежек с “лепестковыми” переходами (изображены согласно реальным траекториям, рис. 1, 2), Rj – радиус-векторы, L – область слияния модулей в феврале–марте; 2 – метки (GC) экскретом хвостовых желез; 3 – экскременты, встреченные на суточных переходах М3; 4 – центральная система переходов М3 вдоль юго-западного склона wsh 4; 5 – область отдаленных переходов М3 с преобладанием комплексного патрулирования.

Модульная структура участка обитания. Полигоны повышенных частот (A–I на рис. 4) совпадали с местами концентрации лежек (рис. 1, 2, 5). Встав с лежки, М3 затем обходил окрестности, и нередко возвращался к прежней лежке или ложился недалеко от нее, в том числе и днем. В других случаях самец, проходя недалеко от лежек, но не заходя к ним, менял траекторию, совершал частые повороты, активно собирал пищу. В результате формировались траектории разнообразной “лепестковой” формы, для которых лежки и их группы представляли собой центры ориентации, в том числе и в случаях, когда кабарга, совершив переход, не ложилась (рис. 5). Подобные структуры названы нами модулями, в которых на небольших площадях кабарга совмещает многие активности своего поведения. Естественно, что модули приурочены к локальным ядрам активности.
“Лепестковые” переходы хорошо заметны у А и В, Н и I, между лежками которых расстояния были не меньше 200 м (рис. 5). При уменьшении этих расстояний до 100 м и меньше (между С к D, E и далее) заметно слияние модулей, распределение лежек при множестве удобных для лежания мест становится не концентрированным. Присутствие соседей: самцов и самок в зонах наложения участков обитания отражалось на формировании модулей H и E, J и расширении G, их длительное отсутствие (М7 с января) – на почти полном исчезновении модуля А. Вследствие этого переходы самца в начале марта были ограничены площадью 0.5 км2.
Средние размеры модуля, определенные по длине радиус-вектора ∣Rmod∣ до наиболее удаленной от них точки замкнутых переходов, составили: $\overline {{{R}_{{{\text{mod}}}}}} $ = 178 м (Sd = 70.87; Cv = 39.9%, n = 15). $\overline {{{R}_{{{\text{mod}}}}}} $ модулей на склоне (у А, В, С), когда кабарга не выходила на днище долины, равнялись 143 м, Sd = 43.0, CV = 25.9%, n = 11. Наибольшие |Rmod| совпадали с включением фрагментов патрулирования c мечением экскретом хвостовых желез (CG). Расстояния вдоль склона от лежек М3 к периферии у места соседства с М7 составляли не больше 300 м, в месте соседства с М8 и М5 у Е и F – 200–300 м.
В модулях у окраины участка обитания кабарга лежала, активно собирала пищу, совмещая этот паттерн с маркировкой и обходом небольших площадей, находясь в окрестности лежек не менее нескольких часов. В присутствие поблизости соседа использовала несколько соседних модулей (“дежурство”) до суток и дольше. Различие параметров траекторий модулей и переходов между ними показывают карты плотностей параметров (рис. 6). Наибольшие значения двумерной плотности при параметре сглаживания h × 1 находились в модулях в интервале от 2.5 до 5.5 м, между ними от 4.3 до 6.1 м. Средняя длина ∣Va∣ в модуле при съемке компасом – 3.1 м (Sd = 2.08, min – 0.5, max – 16, n = 1422) соответствует этим значениям. Для углов отклонения $\varphi _{{\text{i}}}^{{\text{j}}}$ в модулях происходит расслоение наибольшей плотности на две почти симметричных области. максимумы которых находятся, приблизительно, на схожем расстоянии от оси х (0)– в окрестности ±2 рад. Между модулями данный интервал составил ±1 рад. Обходы периферии участка обитания (свыше 1 км; n = 29) против хода часовой стрелки преобладали над правосторонними – 1: 1.4, в том числе, при патрулировании через H, I, J (рис. 1, 2) – 1: 2, на переходах вдоль ключа Кривой – 1: 1.5. По склону М3 переходил в разных направлениях.
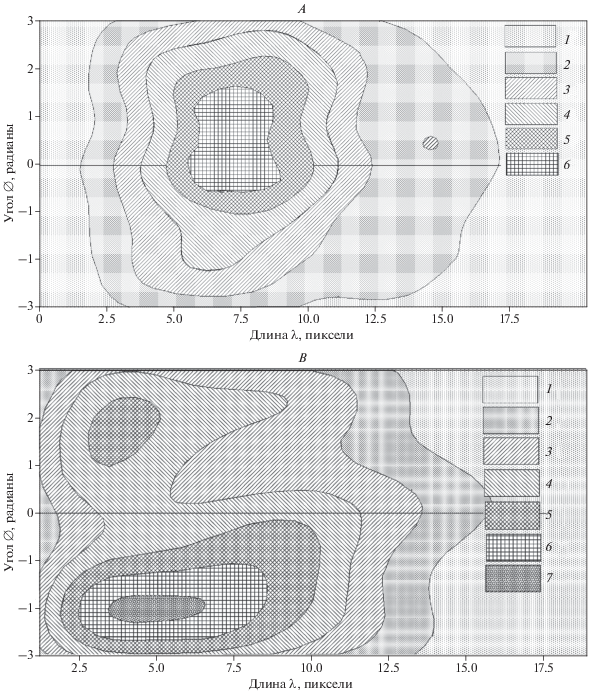
Рис. 6.
Контурная карта распределения двумерной плотности параметров для длин $\Lambda _{i}^{j}~$ и углов $\varphi _{i}^{j}$ векторов Vα (по оценке параметра Скотта с максимальным сглаживанием – h × 1) для векторов Va, их групп самца кабарги на переходах между модулями A и на “лепестковых” треках модулей B; на рис. А значения плотности: 1 – 0.0 < x ≤ 0.004, 2 – 0.004 < < x ≤ 0.008, 3 – 0.008 < x ≤ 0.012, 4 – 0.012 < x ≤ 0.016, 5 – 0.016 < x ≤ 0.02, 6 – 0.02 < x ≤ 0.024; на рис. В: 1 – 0.0 < x ≤ 0.003, 2 – 0.003 < x ≤ 0.006, 3 – 0.006 < x ≤ 0.009, 4 – 0.009 < x ≤ 0.012, 5 – 0.012 < x ≤ 0.015, 6 – 0.015 < x ≤ 0.018, 7 – 0.018 < x ≤ 0.021.
При преобладании комплексного патрулирования на всем участке обитания, активное патрулирование происходило на фрагментах пути, обычно не превышающих 0.2–0.3 км. Только один протяженный обход по северному склону ночью 18–19.02.2015 г. можно отнести к паттерну активного патрулирования. При частых заходах самцов, в зонах перекрытия с их участками и в их окрестностях М3 оставил значительное количество меток экскретом хвостовой железы (CG) (рис. 1, 2, 5), и 54.1% меток (от 98) располагалось в общем центре активности участка обитания, в том числе вблизи лежек модулей. Перекрытие центра активности заходами самцов-соседей достигло за зиму 53.5%.
ОБСУЖДЕНИЕ
Результаты применения комплексной методики характеризуют один из вариантов использования самцом кабарги участка обитания. Данный вариант согласован с условиями местообитания и активным проникновением самцов-соседей к центральной области участка обитания. В результате в центре активности на 39% площади участка обитания самец находился более 84% времени. Особенность состоит в том, что самец М3, как и часть его соседей, поддерживали приблизительную конфигурацию и размеры своих участков весь период слежения, совершая протяженные обходы, патрулирования. Но после периода гона (с января) они ограничили число обходов периферии. При этом происходило перераспределении пространственной активности в связи с заходами соседей, с формированием локальных ядер и менее частом использовании части прежних. В других условиях было достаточно обычным уменьшение размера участков обитания, установленного по внешним обходам, у взрослых самцов кабарги после периода гона (Зайцев, 1991). При этом размеры участка обитания М3 в декабре и январе-феврале (табл. 1) и другие внешние параметры соответствовали вариациям размеров участков обитания самцов со второго года жизни.
Данные об изменении площади участка, которую самец обходил с начала слежения за ним, характеризуют периодичность проявления пространственной активности особи, вследствие чего прирост данной площади происходил через определенные интервалы времени. Также данные позволяют определить минимальное число дней тропления (приблизительно, около 10 дней), необходимое для репрезентативной оценки размера участка. Несколько схожую с этими результатами (рис. 3) зависимость демонстрируют данные о приросте площади участка у лося (Alces alces) (Зайцев, 1994), а также результаты радиослежения койота (Canis latrans) о связи средней площади со средним числом локаций (кроме данных о периодичности изменения участка) (Bekoff, Mech, 1984).
При смещении пространственной активности кабарги в разные периоды возможно определить размеры более-менее стабильного участка (трансформируемой смещаемой площади) в конкретных экологических и социальных условиях, связанных с нормой двигательной и других активностей поведения. Для определения такой “перманентной” площади предложена формула: Sst = Sshr ‒ ($\overline {IA} $)Т; где Sshr ‒ площадь участка за все время наблюдений (в данном случае с начала декабря по март), Т (сут) – период стабилизации размера участка от окончания одного до начала следующего заметного прироста площади; ($\overline {IA} $) ‒ средний суточный ее прирост (Зайцев, 2014). При Sshr = 1.21 км2 (площадь участка за два периода зимы), $\overline {IA} $ = 0.023 км2; Т, меняющемся от 6 до 10 сут (рис. 3) размеры перманентной площади находятся в пределах 0.98–1.07 км2. Эти значения близки к реально наблюдаемым размерам участка в каждый период зимы (табл. 1).
Представление участка обитания в качестве статистически вероятностной системы находится в основе методов расчета его размера, других параметров и конфигурации в моделях, использующих данные радио- и спутникового GPS-слежения (Worton, 1989; Powell, 2000; Moorcroft, Barnett, 2008; Kie et al., 2010; etc.). Это исходит и из результатов многосуточных троплений кабарог (Зайцев, 1991). При уникальности каждой траектории в пограничной области участка обитания М3 и его соседей, пути кабарги в своей совокупности формировали определенные “полосы” обходов, которые только в некоторых местах с заметными ориентирами: выступами склона, ложбинами и др., изменениями растительного покрова – сужались до двух-четырех десятков метров (рис. 1, 2, 5). Данные направления обходов вместе с переходами вдоль общего центра активности (центральная система переходов) составили каркас структуры участка обитания, объединяющий локальные ядра активности и другие элементы.
В общем центре активности участка обитания, включающего места с частотами посещений не менее 4–6 в сети ячеек 1 га за зиму (рис. 4), 50% частот посещений охватывали всего 17–18% площади участка обитания, формируя мозаику локальных ядер активности. Как и 64%-полигон частот при радиорегистрации, центр протянулся вдоль водораздела. Линии перегиба поверхностей рельефа (гребни водоразделов, выступы склона, днища долин ключей и их русел), границы растительных сообществ определяли основные направления переходов кабарог, намечали рубежи между участками самцов-соседей. Концентрация переходов кабарги с формированием хорошо выраженной центральной системой обусловлена в данном варианте сочетанием рельефного ориентира (террасы) с благоприятными для кабарги условиями в сложном по составу кедровнике с сомкнутым пихтовым подростом, “жердняком” на крутом южном склоне, прогреваемом днем, защищенном зимой от муссонного ветра. Не явная связь посещений с углом наклона склона (табл. 4) не противоречит формированию конструктивной системы основных переходов, во многом пролегающих вдоль более пологих форм рельефа (террас и др.).
Неравномерное посещение разных частей участка обитания, однако, не было прямо связано с распределением биомассы основных трофических ресурсов – опада кустистых лишайников в доступной кабарге зоне. По результатам учета, можно приблизительно определить количество доступного лишайника, которое регулярно возобновлялось его опадом. Это количество каждые сутки достигало на участке обитания в феврале 50–70 кг, и в общем центре активности 20–27 кг, что, соответственно, не менее чем в 280–300 и 60–150 раз (без учета другой обычной пищи) превышало биомассу суточного потребления пищи кабаргой – 180–370 г (Холодова, Приходько, 1984; Зайцев, 1991).
Соответственно, распределение биомассы и числа объектов основной пиши имело только локальный эффект для распределения активности самца и его соседей. В учетах на площадках (табл. 2, 3) отсутствуют данные о посещении кабаргой мест у крупных упавших деревьев с концентрацией лишайника (до 1–3.5 кг на дереве; Зайцев, 1991). В пограничных областях участков обитания упавшие деревья привлекали в разное время многих соседей. Поэтому влияние распределения пищи, вероятно, несколько больше, чем отражено в корреляционном и факторном анализах.
Общее число посещений разных мест на участке обитания, количества лежек в локальных ядрах согласовано с комплексом экологических факторов, среди которых наибольшее значение имело проективное покрытие хвойных, прежде всего, подроста пихты с высоким расположением крон, что создает для кабарги защитные условия (Зайцев, 2019). Многочисленные кормовые объекты, рассредоточенные на поверхности снега, обеспечивали чередование отдыха на лежках с активным сбором пищи в их окрестностях, повторным посещением ячеек учета, что и способствовало формированию локальных ядер.
Имея возможность сбора рассредоточенных кормовых объектов на всей площади, самец, прежде всего, совмещал посещение разных частей своего участка с присутствием самок и посещением самцами-соседями его части. На крутом северо-восточном склоне, на днищах долин малых ключей (эллипс 2 на рис. 1) и в других подобных местах (пункты 1, 3 на рис. 1), которые кабарги посещали наиболее часто в декабре нередко встречалось до четырех кабарог, в январе–марте – трех кабарог на площади 6–10 га. Самцов привлекало обычное присутствие в этих местах самок. Нередкое посещение части данных зон самцами-соседями не позволяет отнести их только к участку обитания того или иного самца. В некоторых случаях самцы задерживались на такой площади на 5–6 сут и дольше, например, при заходе бурого медведя (Ursus arctos) 17–23.03.2015 г. в центр активности участка обитания М6 (в окрестности I) (Середкин и др., 2017).
Самцы приближались друг к другу при встрече обычно на расстояния не менее чем несколько десятков метров. Такие дистантные контакты между ними происходили без явной агрессии и стычек. Однако, было обычным преследование соседа по следу, иногда и прыжками. В переплетении многих следовых трасс самец устанавливал принадлежность свежего следа соседу (явно по запаху) не всегда сразу. Конкретные случаи контактов указывают на разные отношения между соседями. Эти отношения менялись от поддержания некоторого паритета (с М7) до неактивного преследования, что приводило к избеганию соседями (М8 и М5) доминанта М3. Проникновение соседей в обширную, редко посещаемую периферию (у I, J), и центр активности участка М3 обусловили значительное (64.7–65.5%) перекрытия участка обитания М3 заходами самцов, на 53.5% центра активности.
На других стационарных площадях доля перекрытия для участков самцов со второго года жизни составляла от 10% до более 40%, менялась в связи с конкретной социальной ситуацией (по данным 1975–1983 гг.; Зайцев, 1991). Соответственно, мы не можем использовать четкий критерий перекрытия для того, чтобы отнести вид к территориальному (т.е. при обособлении территории – охраняемого индивидуального пространства) или не территориальному виду без учета действий и их результатов, направленных на охрану и обособление территории. Подобные критерии, имеющие явно условный характер, предлагаются при моделировании участков обитания одиночно живущих хищных зверей (например, больше или меньше 10%; Sandell, 1989). В нашем случае при проявлении охраны участка обитания (патрулирование, сопровождение следом) лишь на части (46.5%) общего центра активности не отметили заходов соседей.
С увеличением числа заходов, длительности пребывания соседа в зоне наложения участков обитания М3, соответственно, смещал свою активность ближе к местам заходов той или иной кабарги. Возрастал и показатель синхронизации посещений с нарушителем. У зон контактов с соседями и в них формировались своеобразные функциональные элементы структуры – модули, в основном приуроченные к локальным ядрам. В отличие от ядер активности, которые можно выделить дистанционным способом по частотам регистрации, мы определяли модули по свойствам поведения и траекторий движения, что требует непосредственных наблюдений за деятельностью животного. Характерный признак модуля – концентрация (группы) лежек вместе с замкнутыми к ним траекториями “лепестковой” формы, охватывающими небольшие площади (удаление переходов от лежек, по средним на $\overline {{{R}_{{{\text{mod}}}}}} $ 143–178 м).
По результатам восстановления плотности распределения линейных и угловых параметров траекторий, мы установили уникальные характерные портреты движения особи внутри модулей и между ними (при h × 1; рис. 6). В совокупности переходы в модулях, на которых преобладали паттерны сбора пищи и отдыха, отличались от таковых между модулями меньшими длинами векторов Va и большими углами отклонения $\varphi _{i}^{j}$. В модулях наибольшие значения плотности имели разграничение на две области. Можно утверждать, что нередко при также активном сборе пищи на переходах между модулями структура поведения особи в модулях и между ними различалась в связи с разной целью движения. При уникальности каждой траектории, обобщенная характеристика параметров движения, полученная методом восстановления распределения с использованием ядерной оценки плотности, имеет перспективу в создании имитационных агентных моделей (Белотелов и др., 2018) движения животных.
Находясь в одном или двух-трех соседних модулях от нескольких часов до суток и дольше, самец поддерживал свою жизнедеятельность и эффективно охранял территорию. Эти функции обеспечивались сочетанием интенсивного сбора пищи, комфортного поведения с комплексным патрулированием. Модульная структура в данном варианте использования участка обитания согласована с защитой от претендентов, прежде всего, общего центра активности, часть которого нередко посещали самцы-соседи. “Дежурство” в модулях обеспечивало эффект близкого присутствия самца, ограничивало возможность длительного присутствия соседа и проникновение в ядро активности. Подобная форма охраны совмещалась с распределением большого числа (54.1%) специфических меток (GC) в зоне перекрытия участков и в общем центре активности.
С проявлением социальной активности связаны также протяженные обходы периферии с преобладанием паттерна комплексного патрулирования, в том числе и вслед за посещением части участка соседями. Протяженные обходы с активным патрулированием, достаточно обычным в других условиях (Зайцев, 1991), появлялись эпизодически.
ЗАКЛЮЧЕНИЕ
Комплексная методика позволила выяснить связи параметров и структуры участка обитания с поведенческой активностью особи в условиях влияния разных факторов среды и социального окружения. В результате выделен своеобразный вариант использования участка обитания, в котором кабарга поддерживала длительное время его внешние размеры и форму, но значительно меняла свою пространственную активность, преимущественно, в общем центре активности. В условиях почти повсеместной обеспеченности пищей, такое перераспределение согласовано с расположением участков самок и с проникновением самцов-соседей на участок обитания резидента.
Переходы особи формировали своеобразный каркас в форме широких “полос” обходов периферии и центральной системы, объединяющей локальные ядра активности. В благоприятных для кабарги условиях в хвойном лесу данная структура имела согласование с формами, линиями перегиба рельефа и границами растительных сообществ. В результате, наиболее частые контакты между особями с эпизодической их концентрацией приурочены к определенным местам, включенным в систему переходов по участку обитания.
Между самцами преобладали дистанционные контакты с отсутствием явной агрессии при индивидуальных отношениях между соседями. Широкие заходы самцов к центру активности участка обитания самца-резидента сопровождались формированием последним модулей – функциональных элементов структуры, обеспечивающих жизнедеятельность особи вместе с охраной в форме “дежурства”, создающей заслоны от проникновения самцов-соседей.
БЛАГОДАРНОСТИ
Авторы выражают благодарность всем сотрудникам и администрации Сихотэ-Алинского заповедника, Г.И. Банникову, Д.А. Беляеву, сопровождавших нас на маршрутах, И.В. Середкину (Тихоокеанский институт географии ДВО РАН), Дж. Г. Микеллу (WCS) за содействие и обеспечение полевых работ.
Список литературы
Белотелов Н.В., Коноваленко И.А., Назарова В.М., Зайцев В.А., 2018. Некоторые особенности групповой динамики в агентной модели “ресурс-потребитель” // Компьютерные исследования и моделирование. Т. 10. № 6. С. 833–850.
Воронов А.Г., 1973. Геоботаника. Ред. Н.А. Соколова. М.: Высшая школа. 384 с.
Доманов Т.А., 2013. Экология кабарги Moschus moschiferus (Linnaeus, 1758) хребта Тукурингра. Автореф. дис. … канд. биол. наук. Иркутск. 23 с.
Зайцев В.А., 1991. Кабарга Сихотэ-Алиня. Экология и поведение. Ред. Баскин Л.М., Филонов К.П. М.: Наука. 216 с.
Зайцев В.А., 1994. Пространственная структура популяции лося центральной части Европейской России // Бюллетень Mосковского общества испытателей природы. Отд. биологии. Т. 99. № 3. С. 3‒14.
Зайцев В.А., 2002. Векторные системы и ритмы в перемещениях и ориентации лосей (Alces alces L.) и других зверей (Mammallia) // Журнал общей биологии. Т. 63. № 4. С. 335–350.
Зайцев В.А., 2006. Кабарга: экология, динамика численности, перспективы сохранения. Ред. Травина И.В., Зименко А.В. М.: Центр охраны дикой природы. 120 с.
Зайцев В.А., 2014. Применение метода тропления для исследования перемещений копытных и развития концепции участка обитания // Ареалы, миграции и другие перемещения диких животных. Владивосток: ООО “Рея”. С. 103–115.
Зайцев В.А., 2019. Влияние “нового хищника” – хохлатого орла (Nisaëtus nipalensis) – на выбор мест отдыха кабаргой (Moschus moschiferus) // Зоологический журнал. Т. 98. № 4. С. 1–15.
Зайцев В.А., Зайцева В.К., 1980. Методы изучения экологии и поведения кабарги в Сихотэ-Алине // Бюллетень Московского общества испытателей природы. Отдел. биологии. Т. 85. Вып. 4. С. 3‒10.
Зайцев В.А. Середкин И.В., Максимова Д.А., Пименова Е.А., Милаковский Б., Слат Д. К., Микелл Д.Г., 2013. Методы учета плотности населения и исследования распределения кабарги (Moschus moschiferus) в местообитаниях Сихотэ-Алиня / Научно-методические основы составления государственного кадастра животного мира республики Казахстан и сопредельных стран. Алматы: РПГ Институт зоологии. С. 73–80.
Максимова Д.А., Середкин И.В., Зайцев В.А., Микелл Д.Г., 2014. Использование участка обитания кабаргой // Поведение и поведенческая экология млекопитающих. М.: Товарищество научных изданий КМК. С. 72.
Максимова Д.А., Середкин И.В., Зайцев В.А., Микелл Д.Г., 2014a. Участки обитания и суточные перемещения кабарги на Сихотэ-Алине // Ареалы, миграции и другие перемещения диких животных. Владивосток: ООО “Рея”. С. 175–180.
Максимова Д.А., Середкин И.В., Зайцев В.А., Микелл Д.Г., 2015. Участок обитания молодого самца кабарги в Сихотэ-Алинском заповеднике // Материалы XV совещания географов Сибири и Дальнего Востока. Иркутск: изд-во Института географии им. В.Б. Сочавы. СО РАН. 424–426 с.
Приходько В.И., 2003. Кабарга. Происхождение, систематика, экология, поведение и коммуникация. М: ГЕОС. 443 с.
Серёдкин И.В., Зайцев В.А., Петруненко Ю.К., Максимова Д.А., Микелл Д.Г., 2017. Кабарга в питании тигра и медведей на Сихотэ-Алине // Экология. № 4. С. 299–303,
Устинов С.К., 1967. Биология кабарги в связи с ее промысловым использованием. Автореф. дис. … канд. биол. наук. М.: Московская ветеринарная академия. 17 с.
Холодова М.В., Приходько В.И., 1984. Потребление и перевариваемость кормов у кабарги (Moschus moschiferus L.) // Зоологический журнал. Т. 63. № 6. С. 923–928.
Bekoff, M., Mech L.D., 1984. Simulation analyses of space use home range estimates, variability, and sample size // Behavior Research Methods, Instruments and Computers. V. 16. Is. 1. P. 32–37.
Burt W.H., 1943. Territoriality and home range concepts as applied to mammals // J. of Mammalogy. V. 24. № 3. P. 346–352.
Dmitriev E., Kozoderov V., Donskoy S., Melnik P., Sokolov A., 2019. Remote Sensing Methods for the Retrieval of Inventory and Bioproductivity Parameters of Forests Using High Resolution Satellite Images // E3S Web of Conferences, 75. № 01003. (https://doi.org/).https://doi.org/10.1051/e3sconf/20197501003
Green V.J.B., 1985. Aspect of the ecology of the Himalayan musk deer. Ph.D. Dissertation. Cambridge: University of Cambridge. 280 p.
Harris R.B., Cai G., 1993. Autumn home range of musk deer in Baizha Forest, Tibetan Platean // J. Bom. Nat. Hist. Soc. V. 90. № 3. P. 430–436.
Hayne D.W., 1949. Calculation of size of home range // J. of Mammalogy. V. 30. P. 1–18.
Kattel B., 1990. Ecology and conservation of the Himalayan musk deer in Nepal // Intern. Congress of Ecology Wildlife Conservation Symp. Yokohama. P. 26.
Kaufmann J.H., 1983. On the definition and functions of dominance and territoriality // Biol. Rev., Cambridge Phil. Soc. V. 58. № 1. P. 1–20.
Kie J.G., Matthiopoulos J., Fieberg J., Powell R.A., Cagnacci F., Mitchell M.S., Gaillard J.M., Moorcroft P.R., 2010. The home-range concept: are traditional estimators still relevant with modern telemetry technology? // Philosophical Transaction. The Royal Society. B. № 365. P. 2221–2231.
Lychkov I.I., Alfimtsev A.N., Sakulin S.A., 2018. Tracking of Moving Objects with Regeneration of Object Feature Points // Global Smart Industry Conference 2018. № 8 570 061. https://doi.org/10.1109/GloSIC/20188570061)
Maltsev G.N., Yakimov V.L., Soloviev S.V., Lebedeva N.V., 2018. Primary processing of telemetric information using dynamic models of parameter change and partial nonlinear filtration // Informatsionno-Upravliaiushchie Sistemy. № 5. P. 22–34.
Moorcroft P.R., Barnett A., 2008. Mechanistic home range models and resource selection analysis: a reconciliation and unification // Ecology. V. 89. № 4. P. 1112–1119.
Parzen E., Rosenblatt M., 1956. Remarks on Some Nonparametric Estimates of a Density Function // Annual Mathematical Statistic. V. 27. № 3. P. 832–837.
Powell R.A., 2000. Animal home ranges and territories and home range estimators // Research Techniques in Animal Ecology (Pearl M.C., ed.). N.-Y.: Columbia Univer. Press. P. 66–110.
Powell R.A., Mitchell M.S., 2012. What is a home range? // Journal of Mammalogy. V. 93. P. 948–958.
Sandell M., 1989. The mating tactics and spacing patterns of solitary carnivores // Carnivore behavior, ecology and evolution. (J.L. Gatterman, ed.). V. 1. New York: Cornel univ. P. 164–182.
Scott D.W., 2010. Scott′s rule // Journal of WIREs Computational Statistics archive. V. 2. Is. 4. P. 497–502.
Shen K., Neusypin K.A., Liu R., Proletarsky A.V., Guo R., 2017. Technology of error compensation in navigation systems based on nonlinear Kalman filter // Guofang Keji Daxue Xuebao. J. of National University of Defense Technology. V. 39. № 2. P. 84–90.
Worton B.J., 1989. Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies // Ecology. V. 70. № 1. P. 164–168.
Zaitsev V.A., Seryodkin I.V., Maksimova D.A., Soutyrina S.V., 2015. Study of the musk deer population structure in Sikhote-Alin reserve // Achievements of Life Sciences. V. 9. Is. 2. P. 83–86.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал