

Зоологический журнал, 2021, T. 100, № 6, стр. 652-659
Первые сведения по морфологии личинок рода Strongylophthalmyia Heller 1902 (Diptera, Strongylophthalmyiidae)
Н. П. Кривошеина *
Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Ленинский пр., 33, Россия
* E-mail: dipteranina@rambler.ru
Поступила в редакцию 12.02.2020
После доработки 23.04.2020
Принята к публикации 24.04.2020
Аннотация
Установлено, что личинки рода Strongylophthalmyia развиваются в зоне коры древесных стволов (березы, осины, дуба, ильма, клена и маакии), предпочитают лежащие на земле или надломленные стволы, где обитают в частично переработанном или относительно свежем лубе вблизи скоплений сока. Личинки − сапрофлеофаги, взрослые личинки выступают в роли сапро-некрофагов. Для личинок характерны древовидно разветвленные передние и своеобразные трехлопастные задние дыхальца, расположенные в середине заднего края последнего сегмента. Фарингеальный склерит имеет слабо развитый дорсальный бугорок в основании вентрального отростка. Морфологическое своеобразие личинок подтверждает вывод Хеннига о выделении рода в самостоятельное семейство.
Семейство Strongylophthalmyiidae представлено одним родом Strongylophthalmyia Heller 1902. Род длительное время рассматривался в семействе Psilidae, но Хенниг (Hennig, 1958) признал его обособленность от родов семейства Psilidae и выделил в самостоятельное семейство. Ряд авторов включает его в семейство Tanypezidae (Colless, McAlpine, 1970; Cogan, 1980; Ferrar, 1987). Strongylophthalmyia – новое название для рода Strongylophthalmus Hendel 1902, так как последнее было преоккупировано. Виды рода обитают преимущественно в Ориентальной области, где зарегистрировано около 31 вида (Iwasa, 1998). В Палеарктике известны 8 видов: S. pictipes Frey 1935, S. raricornis Shatalkin 1981, S. stackelbergi Krivosheina 1981, S. ustulata (Zetterstedt 1847), S. japonica Iwasa 1992, S. caliginosa Iwasa 1992, S. paula Shatalkin 1993 и S. crinita Hennig 1940. В Неарктике и Неотропике известно по одному виду (Кривошеина, 1981; Krivosheina, 1984; Шаталкин, 1981, 1993; Shatalkin, 1995).
Сведения по биологии видов практически отсутствуют. Имеется лишь краткое указание на обитание личинок под корой лиственных деревьев (Мамаев и др., 1977). Имаго S. ustulata были собраны на стволе осины в Финляндии (Steyskal, 1977). Личинки этого вида обитают под корой осины на территории России (Московская и Вологодская обл., Краснодарский край, Бурятия и Амурская обл.), а S. pictipes – на территории Горного Алтая и Бурятии. Личинки S. stackelbergi выведены из-под коры березы на о-ве Кунашир и ильма, дуба, клена и маакии амурской в Приморском крае.
Известные личинки Psilidae – типичные фито- и флеофаги – питаются растительными тканями, проделывая ходы в стеблях и корнях однодольных и двудольных растений или питаются тканями коры и древесины деревьев (Ferrar, 1987). Некоторые Chyliza выводятся в Северной Америке из скоплений сока: Ch. annulipes Macquart 1835 из скоплений сока на Pinus virginiana Mill. (Teskey, 1976); пупарии Сh. erudita Melander 1920 найдены в скоплениях сока по краям ран Pinus strobus L. (Wehrle, 1930), а Ch. notata Loew 1869 – из ран на стволах вяза (Pechuman, 1943).
На основании имеющихся в коллекции ИПЭЭ РАН материалов в данной статье приводятся сведения по образу жизни и морфологии личинок 3 видов рода Strongylophthalmyia. Каждой серии собранных личинок, куколок и вышедших из них имаго присвоен сквозной порядковый номер, который приводится в материале.
Род Strongylophthalmyia Heller 1902 (рис. 1, 1−6; 2, 1−10)
Рис. 1.
Личинка Strongylophthalmyia stackelbergi Krivosheina 1981: 1 – ротоглоточный аппарат, 2 – переднее дыхальце, 3 – терминальный конец тела сбоку, 4 – терминальный конец тела снизу, 5 – задние дыхальца, 6 – кутикулярные структуры последнего сегмента тела (f – фарингеальный склерит, g – гипостомальный склерит, h – парастомальный склерит, k – фарингеальная мембрана, l – дорсальный отросток фарингеального склерита, m – вентральный отросток фарингеального склерита, n – ротовые крючки, p – вентральный выступ базального склерита ротовых крючков).
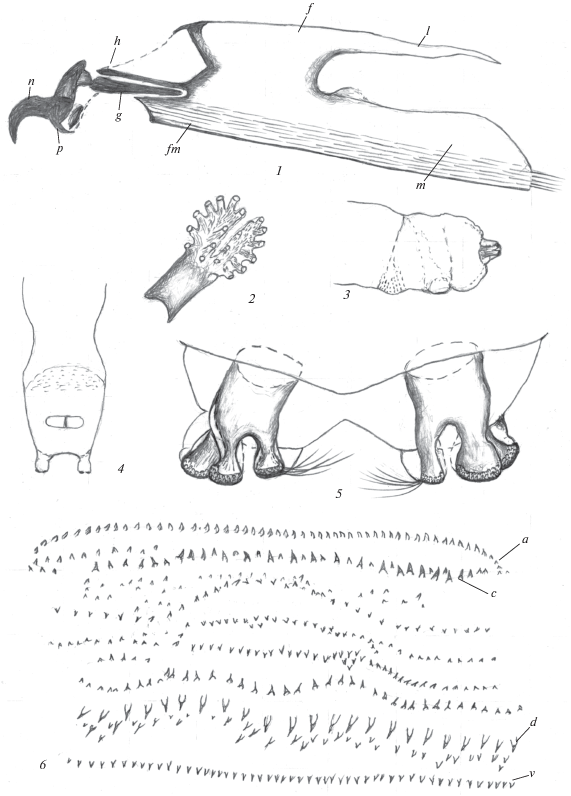
Характеристика личинок. Тело удлиненное, слегка уплощено дорсо-вентрально и несколько сужено в переднем отделе, состоит из 3 грудных и 8 брюшных сегментов, головной сегмент очень маленький. Ротоглоточный аппарат склеротизован, небольшой, полностью втянут в первые грудные сегменты, и ротовые крючки не выступают снаружи. Покровы тела светлые, беловатые или слегка желтоватые, гладкие, без заметных папилл или бугорков. Слегка выступают лишь ползательные валики, расположенные на всех брюшных сегментах. Последний сегмент тела с двумя терминальными короткими конусовидными бугорками. Передние дыхальца, расположенные на боковых сторонах тела на границе I и II грудных сегментов, удлиненные, с двумя разобщенными в вершинной части отделами, несущими пальцевидные выступы с овальными дыхальцевыми отверстиями. Задние дыхальца расположены на коротких конусовидных бугорках, расстояние между бугорками почти в 2 раза превышает их ширину. Атриум дыхалец слегка удлиненный, на вершине с тремя разобщенными выступами, при этом один более узкий, заметно обособлен от двух сближенных более широких выступов. Их вершины со стигмальными пластинками, несущими по одному отверстию, окруженному решетчатыми структурами (рис. 1, 5).
Анальная щель, расположенная в среднем отделе последнего сегмента с вентральной стороны, продольная. Анальная пластинка поперечная, достаточно узкая, ее длина в несколько раз меньше ширины (рис. 1, 4).
Ротоглоточный аппарат типичного для личинок круглошовных двукрылых строения (рис. 1, 1; 2, 1). Ротовые склериты сильно склеротизованы, как и удлиненные гипостомальные и парастомальные склериты. Фарингеальный склерит светлый, практически не склеротизован, кроме переднего краевого отдела. Характерный признак личинок двух исследованных видов – наличие загнутого назад вентрального выступа базального склерита ротовых крючков (мандибул). Фарингеальный склерит с хорошо развитой фарингеальной мембраной, что подтверждает отсутствие хищничества. Образ жизни личинок, их медленные, спокойные перемещения наиболее соответствуют такому типу питания, как сапрофагия, точнее сапрофлеофагия. В то же время для них, как и для большинства ксилобионтов, наиболее характерна не просто сапрофагия, а сапро-некрофагия, способствующая ускорению развития при питании разлагающимися органическими остатками.
Биология. Виды рода приурочены к лесным районам. Личинки обычны во влажном лубе лиственных пород деревьев, окукливаются в его верхних слоях под мертвой коркой (Krivosheina, 1984).
Strongylophthalmyia stackelbergi Krivosheina 1981 (рис. 1, 1–6; 2, 8–10)
Материал. 6 личинок, 2 пупария, 6♂♂, 4♀♀, Россия, Приморский край, Лазовский заповедник, Корпадь, 22.06.1979, 6.07.1979, 20.07.1979, № 143, 167, 342, личинки под корой дуба (Quercus mongolica Fisch.) (сем. Fagaceae), клена (Acer mandshuricum Maxim.) (сем. Aceraceae) и маакии (Maackia amurensis Rupr.et Maxim.) (сем. Leguminosae); Приморский край, Уссурийский заповедник, 17.04.1969, № 27, под корой ильма (Ulmus propinqua Koidz.) (сем. Ulmaceae); Сахалинская обл., Кунашир, Серноводск, 23.09.1972, № 177, 178, под корой березы (Betula mandshurica (Regel) Nakai) (Betulaceae); там же, Менделеево, 22.09.1976, № 94, 25.09.1976, № 105, в лубе березы (Н. Кривошеина).
Описание. Личинка. Тело удлиненное, белое с легким желтоватым оттенком и с четкой сегментацией, слегка расширено в среднем отделе, сужено и закруглено в переднем отделе и не расширено сзади. Все брюшные сегменты с четкими овальными ползательными валиками, но на границе III грудного и I брюшного сегментов значительно меньше остальных. Анальная пластинка расположена в среднем отделе последнего сегмента, поперечная, с параллельными передней и задней сторонами, не сужена и равномерно закруглена на боковых сторонах (рис. 1, 4). Ее длина в 4 раза меньше ширины. Анальная щель продольная.
Ползательные валики широкие с шипиками различных размеров, часть которых образуют поперечные ряды различной длины (рис. 1, 6). Поля ограничены спереди и сзади простым рядом небольших сближенных однородных шипиков (a, b). К ним примыкают 2 ряда более крупных шипиков (c, d), между которыми расположены 4 ряда мелких структур. Крупные шипики сопровождают разрозненные мелкие шипики.
Передние дыхальца удлиненные, лопастевидные, их передняя часть разделена на 2 части, каждая из которых несет по 10 пальцевидных выступов со стигмальными овальными отверстиями на вершине (рис. 1, 2). Задние дыхальца расположены на коротких цилиндрических бугорках в среднем отделе последнего сегмента (вид сбоку, рис. 1, 3). Бугорки удалены друг от друга на расстояние, в 2 раза превышающее их ширину (рис. 1, 4). Дыхальца с удлиненным атриумом, с тремя выступами в верхнем отделе, из них один более узкий, удален на большее расстояние от двух более широких и сближенных (рис. 1, 5). Все они несут на вершине по одному овальному отверстию, окруженному узкой полоской темных решетчатых структур, занимающих всю вершинную часть выступов. Стигмальная пластинка дыхалец с хорошо развитыми периспиракулярными железами, несущими длинные темные волоски.
Ротоглоточный аппарат с достаточно хорошо склеротизованными ротовыми, гипостомальными и парастомальными склеритами и светлым, практически прозрачным фарингеальным склеритом (рис. 1, 1). Фарингеальная мембрана хорошо развита, массивная. Ротовые крючки обычной серповидной формы, акцессорные склериты отсутствуют. Вентральный выступ базального склерита ротовых крючков узкий, удлиненный, направлен назад. Дорсальный отросток фарингеального склерита узкий, короче вентрального. Вентральный отросток широкий массивный, с небольшим слабо очерченным дорсальным бугорком в основании.
Длина тела 5 мм.
Пупарий удлиненно-овальный, слегка сужен на уровне грудных и трех последних брюшных сегментов, светло-коричневый, окрашен равномерно, заметно уплощен дорсо-вентрально. Дорсальная и вентральная поверхности сегментов на большем протяжении гладкие. Боковые стороны брюшных сегментов с узкими складками, наиболее заметными на четырех последних брюшных сегментах.
Передние дыхальца по строению соответствуют аналогичным структурам личинок, двухлопастные, ветвистые, расположены на боковой стороне I грудного сегмента (рис. 2, 8, 9). Последний сегмент тела с конусовидными бугорками, на вершине которых расположены задние дыхальца (рис. 2, 10). Вершина атриума с одним более узким выступом, четко отделенным от более сближенных двух других относительно широких выступов. В целом структура атриума аналогична его структуре у личинок, но бугорки, несущие дыхальца, иной формы. Они конусовидные, в основании удалены друг от друга на расстояние, не превышающее размер дыхалец.
Рис. 2.
Личинка Strongylophthalmyia ustulata (Zetterstedt 1847) (1−7), пупарий Strongylophthalmyia stackelbergi Krivosheina 1981 (8−10): 1 – ротоглоточный аппарат; 2, 3 – задние дыхальца; 4, 5 – терминальный конец тела сбоку и снизу; 6 – кутикулярные структуры последнего сегмента тела; 7 – переднее дыхальце; 8, 9 – передний конец тела снизу и сбоку; 10 – терминальный конец тела снизу: s – переднее дыхальце, t – заднее дыхальце.
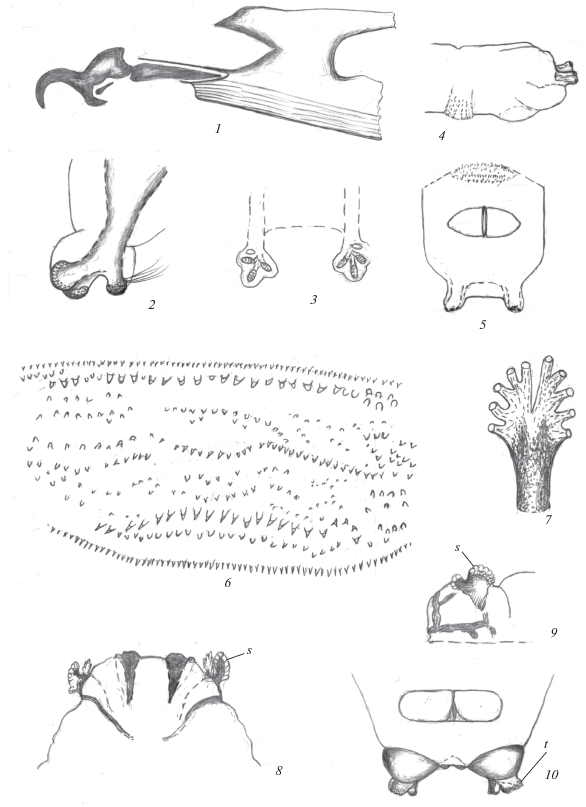
Длина пупария 3.5 мм.
Биология. Личинки предпочитают лежащие на земле или нависающие над землей стволы березы, ильма, ясеня, клена, дуба и маакии амурской. Обитают в частично переработанном, буроватом лубе или в относительно свежем лубе вблизи скоплений сока. Перед окукливанием личинки концентрируются в верхних частях луба. Пупарии встречаются в темном лубе среди древесной трухи непосредственно под коркой. На территории Лазовского заповедника личинки обнаружены в стволах дуба в свалке стволов около арбузной бахчи, заселенных личинками дровосеков Mesosa myops Dalman 1817, и златками (Н. Кривошеина, М. Кривошеина, 2019). Личинки S. stackelbergi сосредоточены преимущественно с нижней стороны стволов, часто непосредственно в колыбельках дровосеков. В таких же свалках около бахчи, в стволах маакии амурской с сохранившимися узкими полосками свежего луба, перемежающегося с участками частично переработанного бурого луба, личинки S. stackelbergi обитают в лубяных волокнах, частично пронизанных мицелием, на границе с заболонью. Совместно с ними встречаются небольшие личинки-сапрофлеофаги Teucholabis esakii (Alexander 1924) и T. yezoensis Alexander 1924 (Limoniidae). На относительно свежих участках луба первоначально поселяются личинки-флеофаги – Lenitovena pteropleuralis (Hendel 1927) (Tephritidae), вблизи которых поселяются личинки жуков Elacatis kraatzii Reitter 1879 (Othniidae). Тип питания личинок жуков не известен, но судя по их постоянным биотопическим связям с личинками пестрокрылок, они способны питаться или непосредственно соком, или пропитанными соком субстратами. На частично отработанных этими видами субстратах им постоянно сопутствуют личинки-сапронекрофаги львинок Maackiana laminiformis Krivosheina 1973 (Stratiomyidae). Вокруг скоплений этих видов постоянно присутствуют хищные личинки жуков Histeridae, двукрылых Lonchaea Fallén 1820 (Lonchaeidae) и Phaonia Robineau-Desvoidy 1830 (Muscidae). Совместно с личинками S. stackelbergi в лубяных волокнах встречаются личинки Milichiella argyrogaster (Perris 1876) (Milichiidae), являющиеся широко известными сапрофагами, питающимися древесными остатками (Ferrar 1987), и личинки-сапроксилофаги Sciaridae. На участках с плесенью под корой клена личинки S. stackelbergi встречались совместно с коллемболами, а хищники были представлены личинками Phaonia sp. (Miscidae).
На о-ве Кунашир имаго выведены в основном из лубяных волокон упавших стволов березы. Кора стволов достаточно прочная, но эпидермис отходит в виде тонкой пленки. Под корой в скоплениях натеков сока встречались личинки рассмотренного вида Strongylophthalmyia, но взрослые личинки, как правило, концентрировались непосредственно под эпидермисом.
Распространение. Вид зарегистрирован в Приморском крае (Лазовский и Уссурийский заповедники) и на о-ве Кунашир (Менделеево, Серноводск).
Strongylophthalmyia ustulata (Zetterstedt 1847) (рис. 2, 1−7)
Материал. 4 личинки, 6 ♂♂, 5 ♀♀. Россия, Московская обл., Красная Пахра, 20.05.1973; Вологодская обл., Никольское, 6.06.1983, № 122, 123; Краснодарский край, ст. Северская, 10.07.1970, № 61, 62; Бурятия, Бабушкин, 5.06.1976, 6.06.1976, № 92, 96, 107, 108 (Н. Кривошеина); Амурская обл, Кундур, 4.05.1975, 14.05.1975, № 95, 99, 165 (А. Зайцев), личинки под корой осины − Populus tremula L. (сем. Salicaceae).
Описание. Личинка. Тело светло-желтое, с четкой сегментацией и поперечными овальными ползательными валиками, несущими поперечные ряды заостренных и овальных шипиков различных размеров. Шипики образуют четкий широкий параллельносторонний поясок (вид сбоку), ограниченный спереди и сзади рядом небольших и однородных, плотно сидящих шипиков. На границе III грудного и I брюшного сегментов валик меньших размеров и с более слабо выраженными шипиками. Сегменты тела гладкие, без развитых бугорков или папилл. Последний сегмент с дорсальной стороны с глубокой продольной узкой срединной складкой, доходящей до основания бугорков (рис. 2, 4). Такие же глубокие складки расположены на наружной стороне сегмента, сбоку от бугорков. Терминальный конец последнего сегмента с двумя короткими светлыми цилиндрическими бугорками, которые окрашены, как и все сегменты тела, и которые несут на вершине задние дыхальца. Терминальные бугорки широко расставлены, удалены друг от друга на расстояние, в 2 раза превышающее ширину бугорка (рис. 2, 5).
Анальная пластинка поперечно-овальная с продольной узкой щелью. Ширина пластинки в 2.5 раза превосходит ее длину (рис. 2, 5).
Трахейная система амфипнейстического типа. Передние дыхальца расположены на боковой стороне грудных сегментов, на границе I и II грудных сегментов. Дыхальца лопастевидные, с двумя разветвленными вершинными долями, несущими по 5 пальцевидных коротких выступов (рис. 2, 7). Атриум удлиненный, цилиндрический, длина его неразветвленного основания равна длине разветвленного вершинного отдела. Задние дыхальца с удлиненным атриумом, вершина которого с тремя короткими выступами, из которых один несколько ýже двух других и отделен от них широкой выемкой (рис. 2, 2, 3). На вершинах всех выступов по одному стигмальному отверстию, окруженному темной полоской из решетчатых структур, занимающей всю вершинную часть выступов (рис. 2, 2). Поверхность стигмальной пластинки с периспиракулярными железами, несущими длинные волоски.
Ротоглоточный аппарат типичного для личинок круглошовных двукрылых строения, с хорошо развитыми серповидными ротовыми крючками, но без акцессорных, дополнительных, склеритов (рис. 2, 1). Вентральные выступы ротовых крючков (мандибул) достаточно массивные, конусовидные, их вершина направлена назад. Гипостомальные и парастомальные склериты удлиненные, последние стержневидные. Фарингеальный склерит массивный, но светлый, слабо склеротизован. Его дорсальные отростки узкие, короче массивных вентральных. Вентральные отростки фарингеального склерита с небольшим дорсальным бугорком в основании. Фагингеальная мембрана хорошо развита, широкая, что подтверждает отсутствие хищничества.
Биология. Личинки зарегистрированы под пологом деревьев только в стволах осин, преимущественно лежащих на земле или опирающихся на ветви и нависающих на небольшой высоте над землей. В Краснодарском крае личинки обитали в лежащих на земле под пологом леса стволах осины, где концентрировались с их северной стороны. Личинки обитали в толстой коре, местами светлой, почти свежей, с ходами короедов; взрослые личинки концентрировались на участках с переработанными волокнами в темной трухе под коркой вблизи трещин. Под корой личинкам сопутствовали личинки-некрофаги Lonchaeidae, личинки-сапронекрофаги Neopachygaster meromelaena Dufour 1841 (Stratiomyidae) и хищные личинки Empididae.
В Вологодской обл. личинки были характерны для смешанных переувлажненных лесов вблизи водоемов. Личинки обитали в лежащих на земле стволах осины преимущественно с темным, значительно переработанным лубом, где концентрировались на теневой стороне стволов в темной трухе непосредственно под коркой. Личинки были рассеяны по стволу, иногда наблюдались скопления нескольких экземпляров. На территории Бурятии (Бабушкин) и Амурской обл. (Кундур) личинки также были обнаружены в лубяных волокнах лежащих на земле стволов и их обрубков, где обитали как в толще луба, так и в его поверхностных слоях непосредственно под коркой. В качестве их спутников среди волокон луба были отмечены личинки-сапронекрофаги Homalocephala albitarsis Zetterstedt 1838 [=H. biumbrata (Wahlberg 1839)] (Otitidae).
Личинки – сапрофлеофаги, питаются бродящим соком и разлагающимся лубом, в конце развития у взрослых личинок преобладает сапронекрофагия, т.е. питание органическими остатками животного происхождения.
Распространение. Вид распространен в Европе, включая европейскую часть России, а также в Бурятии и Амурской обл.
Strongylophthalmyia pictipes Frey 1935
Материал. 2 пупария, 2 ♂♂, 1 ♀. Россия, Горный Алтай, Артыбаш, 21.06.1981, № 139, личинки под корой осины, вылет имаго 30.06.1981; Бурятия, Бабушкин, 5.06.1976, № 94; 6.06.1976, № 107, 110, под корой осины; там же 7.06.1976 № 123; 10.06.1976 № 156, под корой березы − Betula sp. (Н. Кривошеина).
Распространение. Вид описан по материалам из Финляндии, зарегистрирован на территории России в Горном Алтае (Артыбаш) и в Бурятии (Бабушкин).
Биология. Материал по личинкам недостаточен для их морфологического исследования, поэтому приводится только описание мест обитания личинок S. pictipes.
Личинки были обнаружены в лесных насаждениях на открытых местах, вдоль линий электропередач, в лежащих на земле стволах осины (Бурятия, Бабушкин). Личинки встречались среди лубяных волокон, при этом взрослые личинки концентрировались в их верхних слоях непосредственно под коркой. Их обычно сопровождали личинки-сапронекрофаги рода Homalocephala Zetterstedt 1838 (Ulidiinae) и хищные личинки Megamerina dolium (Fabricius 1805) (Megamerinidae).
На Алтае личинки обнаружены в стволе осины, нависающей над землей на высоте 1 м. Ствол заселен личинками дровосека Saperda, а толстые ветки заселены короедами рода Trypophloeus Fairmaire 1869. Кора прочная, луб местами светлый, местами темный, по границе светлого и темного луба обитают личинки-сапронекрофаги Xylomya czekanovskii Pleske 1925 (Xylomyidae). По границе старых ходов личинок усачей на участках с черным лубом обитают личинки-сапронекрофаги Neopachygaster secernibilis Krivosheina 1973 (Stratiomyidae). За ними охотятся по всему стволу личинки Phaonia sp. (Muscidae). Cреди волокон луба обитают личинки-сапрофаги Pseudotephritis corticalis (Loew 1873) (=trypetoptera Hennig 1939) и личинки-сапронекрофаги Lonchaea sp. (Lonchaeidae). Вокруг сучков под коркой энергично перерабатывают лубяные волокна личики Sciaridae. Личинки S. pictipes встречаются среди лубяных волокон в различных участках коры, но предпочитают поверхностные слои волокон непосредственно под коркой, где концентрируются также пупарии.
Личинки двух исследованных видов различаются следующим образом.
– Передние дыхальца несут около 20 пальцевидных выступов. Анальная пластинка узкая, параллельносторонняя, ее ширина в 4 раза превышает длину. Вершина вентрального выступа базального склерита мандибул узкая, заостренная, направлена назад. Конусовидные бугорки пупария, несущие задние дыхальца, разобщены. Ползательные валики вдоль средней линии не менее чем с 8 поперечными рядами шипиков. Личинки обитают под корой дуба, клена, маакии, ильма и березы.......................... ………………..Strongylophthalmyia stackelbergi Kriv.
– Передние дыхальца несут не более 10 пальцевидных выступов. Анальная пластинка несколько расширена, ее ширина в 2.5 раза превосходит длину. Вершина вентрального выступа базального склерита мандибул конусовидная, направлена назад. Ползательные валики вдоль средней линии не более, чем с 6 поперечными рядами шипиков. Личинки обитают под корой осины.…...................................................... ......................Strongylophthalmyia ustulata (Zett.).
ЗАКЛЮЧЕНИЕ
До последнего времени положение семейства в системе Acalyptrata окончательно не установлено. Род Strongylophthalmyia сближался то с Psilidae, то с Tanypezidae. Имеющиеся сведения по изученным личинкам Psilidae (Ferrar, 1987) говорят об их существенных отличиях от личинок Strongylophthalmyia. Так, для личинок Chyliza Fallén 1820 и Psila Meigen 1803 характерны массивные гипостомальные склериты и слабо выраженный вентральный выступ базального склерита мандибул. Отростки фагингеального склерита равной длины или даже дорсальные несколько превосходят вентральные. Не выражен дорсальный бугорок вентрального фарингеального отростка. Склеротизованные бугорки, несущие задние дыхальца, слиты или сближены в основании. Перитрема дыхалец массивная и сильно склеротизована. Передние дыхальца в виде простой вееровидной пластики с 5–15 пальцевидными выступами. Личинки в стеблях и на корнях различных растений, фитофаги, отдельные виды наносят существенный вред корнеплодам.
Кажется вполне закономерным вывод о существенном отличии видов рассматриваемого рода от представителей семейства Psilidae, в состав которого род был включен при описании (Heller, 1902).
Семейство Tanypezidae представлено в основном в Неотропическом регионе, в Палеарктике известен один вид Tanypeza longimana Fallén 1820, который вместе с T. picticornis Knab & Shannon, 1916 был зарегистрирован в Америке севернее Мексики (Foote, 1970).
Опубликованное морфологическое исследование личинки T. longimana (Foote, 1970) говорит о некотором ее сходстве с личинками Strongylophthalmyia. Для личинки характерны удлиненные, двуветвистые с 17 пальцевидными выступами передние дыхальца, хорошо развиты вентральные конусовидные выступы базальных склеритов мандибул, вершины которых направлены вентрально вниз или назад, удлиненные гипостомальные склериты. Но наблюдается совершенно иной тип строения заднего отдела трахейной системы и задних дыхалец, хотя образ жизни личинок обоих родов однотипен. Наблюдаются также различия в строении дорсальной перемычки и дорсальных отростков фарингеального склерита ротоглоточного аппарата. Таким образом, можно отметить несколько большее сходство рода Strongylophthalmyia c Tanypeza Fallén 1820, чем с представителями Psilidae, но в целом для решения таксономических вопросов необходимо дальнейшее более детальное исследование всего комплекса признаков родственных групп Acalyptrata.
Список литературы
Кривошеина Н.П., 1981. Новые палеарктические виды двукрылых рода Strongylophthalmyia Hell. (Diptera, Strongylophthalmyiidae // Энтомологическое обозрение. Т. 60. Вып. 1. С. 183–186.
Кривошеина Н.П., Кривошеина М.Г., 2019. Ксилофильные двукрылые (Insecta, Diptera) Лазовского государственного заповедника (Россия) // Nature Conservation Research. Заповедная наука. Т. 4. № 3. С. 78−92.
Мамаев Б.М., Кривошеина Н.П., Потоцкая В.А., 1977. Определитель личинок хищных насекомых – энтомофагов стволовых вредителей. М.: Наука. 392 с.
Шаталкин А.И., 1981. Claraeola crassula sp. n. (Pipunculidae) и Strongylophthalmyia raricornis sp. n. (Strongylophthalmyiidae) – новые своеобразные двукрылые фауны СССР // Зоологический журнал. Т. 60. № 5. С. 791–793.
Шаталкин А.И., 1993. К систематике мух семейства Strongylophthalmyiidae (Diptera) // Зоологический журнал. Т. 72. № 10. С. 124–131.
Colless D.Y., McAlpine D.K., 1970. Diptera (Flies). Chapter 34. The Insects of Australia. Australia: Melbourne University Press. P. 656–670.
Cogan B.H., 1980. Family Tanypezidae. In: Crosskey R.W. (ed.). Catalogue of the Diptera of the Afrotropical Region. London: British Museum (Natural History). P. 588.
Ferrar P., 1987. A Guide to the breeding habits and immature stages of Diptera Cyclorrhapha. Entomonograph V. 8. Part 1: text. Leiden – Copenhagen: E.J. Brill/ Scandinavian Science Press. 478 p.
Foote B.A., 1970. The larvae of Tanypeza longimana (Dipteras: Tanypezidae) // Annals of the Entomological Society of America. V. 63. P. 235–238.
Frey R., 1935. Neue Diptera brachycera aus Finland und angrenzenden Ländern. III // Notula Entomologica. V. 15. P. 97–101.
Heller K.M., 1902. Strongylophthalmyia nom. nov. für Strongylophthalmus Hendel // Wiener entomologische Zeitung. Bd. 21. P. 226.
Hendel F., 1902. Strongylophthalmus, eine neue Gattung der Psiliden // Wiener entomologische Zeitung. Bd. 21 (5). P. 179–182.
Hennig W., 1958. Die Familien der Diptera Schizophora und ihre phylogenetischen Verwandtschaftsbeziehungen // Beiträge zur Entomologie. Bd. 8 (5/6). P. 506–688.
Iwasa M., 1992. The genus Strongylophthalmyia Heller (Diptera, Strongylophthalmyiidae) from Japan, with descriptions of two new species // Japanische Journal of Entomology. V. 60 (3). P. 660–666.
Iwasa M., 1995. Occurrence of Strongylophthalmyia ustulata Zetterstedt (Diptera, Strongylophthalmyiidae) from Japan and North Korea // Japanische Journal of Entomology. V. 63. P. 57–58.
Iwasa M., 1998. Family Strongylophthalmyiidae. In: Papp L., Darvas B. (eds). Contributions to a Manual of Palaearctic Diptera. V. 3. Higher Brachycera. Budapest: Science Herald. 880 p.
Krivosheina N.P., 1984. Family Strongylophthalmyiidae. In.: Soós A. and Papp L. (eds). Catalogue of Palaearctic Diptera. V. 9. Micropezidae – Agromyzidae. Budapest: Akadémiai Kiadó. P. 27–28.
Pechuman L.L., 1943. Notes on the biology of Chyliza notata (Diptera, Psilidae) // Bulletin of the Brooklyn Entomological Society. V. 38. P. 97.
Shatalkin A.I., 1995. New and little known species of Lauxaniidae and Strongylophthalmyiidae (Diptera) // Russian Entomological Journal. V. 4. № 1–4. P. 145–157.
Steyskal G.C., 1971. Notes on the genus Strongylophthalmyia Heller, with a revised key to the species (Diptera: Strongylophthalmyiidae) // Annals of the Entomological Society of America. V. 64 (1). P. 141–144.
Steyskal G.C., 1977. Family Strongylophthalmyiidae. In: M.D. Delfinado, D.E. Hardy (eds). A catalog of the Diptera of the Oriental Region. V. III. Honolulu: University Press of Hawaii. P. 21–23.
Teskey H.J., 1976. Diptera larvae associated with trees in North America // Memoirs of the Entomological society of Canada. № 100. P. 1–51.
Wehrle L.P., 1930. A note on the food habits of Chilyza erudita Mel. (Diptera) // Journal of Economic Entomology. V. 23. P. 286.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал