

Зоологический журнал, 2022, T. 101, № 1, стр. 23-36
Анализ распространения ящериц рода Lacerta на северо-востоке Кавказа (Дагестан, Россия) с использованием ГИС-технологий и методов моделирования пространственного распределения
М. А. Доронина a, *, Л. Ф. Мазанаева b, **, И. В. Доронин a, *
a Зоологический институт РАН
199034 Санкт-Петербург, Россия
b Дагестанский государственный университет
367025 Махачкала, Республика Дагестан, Россия
* E-mail: igor.doronin@zin.ru
** E-mail: mazanaev@mail.ru
* E-mail: igor.doronin@zin.ru
Поступила в редакцию 17.11.2020
После доработки 17.02.2021
Принята к публикации 24.02.2021
- EDN: AAVWHL
- DOI: 10.31857/S0044513421110052
Аннотация
Приведен анализ пространственного распространения зеленых ящериц рода Lacerta в пределах Дагестана. Уточнены и детализированы ареалы L. agilis, L. media и L. strigata. Создан набор векторных (точек находок) и растровых синтетических карт, отражающих пространственное распределение видов. Проведено биоклиматическое моделирование пригодности местообитаний. Минимальное перекрывание экологических (эколого-климатических) ниш отмечено для L. media со всеми изученными видами, максимально у L. agilis и L. strigata.
Одним из центров таксономического разнообразия зеленых ящериц рода Lacerta Linnaeus 1758, включающего по современным представлениям 10 видов (Kornilios et al., 2019), выступает Кавказ. Здесь обитает три вида рода – прыткая, Lacerta agilis Linnaeus 1758, представленная шестью подвидами11, номинативный подвид средней ящерицы, L. m. media Lantz et Cyrén 1920, и полосатая ящерица, L. strigata Eichwald 1831, рассматриваемая как монотипический вид (Uetz et al., 2020; www.lacerta.de). Если принять восточную границу Кавказа по схеме “Фонда сотрудничества для сохранения экосистем, находящихся в критическом состоянии” (CEPF) (https://wwf.panda.org/discover/knowledge_hub/where_we_work/black_sea_basin/ caucasus/projects/english/), то к этому списку следует добавить и южный подвид зеленой ящерицы, L. viridis meridionalis Cyrén 1933, чей ареал охватывает, в том числе, Понтийские горы. Ранее популяции зеленой ящерицы севера Анатолии выделяли в самостоятельные узкоареальные подвиды L. v. paphlagonica Schmidtler 1986 и L. v. infrapunctata Schmidtler 1986, сведенные в младшие синонимы L. v. meridionalis (Marzahn et al., 2016).
Отметим, что на протяжении всей истории герпетологических исследований Кавказ относили к наиболее уникальным с точки зрения биоразнообразия территориям Северной Евразии. Согласно результатам изучения уровня эндемизма рептилий Западной Палеарктики (Ficetola et al., 2018) Кавказ входит в число наиболее значимых регионов. В пределах его российской части “горячими точками” разнообразия герпетофауны можно признать территорию Дагестана и Краснодарского края. Здесь обитают все три обозначенных выше вида зеленых ящериц.
История изучения лацертид в пределах Дагестана, занимающего северо-восточную часть Кавказского региона, насчитывает почти 200 лет и ведет свое начало с экспедиции Эйхвальда, наблюдавшего в 1825 г. в районе Тарки “Lacerta deserti, viridis, velox, muralis (agilis L.)” (S. 97, Eichwald, 1834) – речь вероятнее всего идет об Eremias arguta deserti (Gmelin 1789), E. v. velox (Pallas 1771), L. a. boemica и L. strigata (Доронин и др., 2017). Красовский (1932) впервые обнаружил на территории республики среднюю ящерицу. Шибанов (1935) указал для окрестностей г. Кизляр прыткую ящерицу со следующим примечанием: “Единственный экземпляр коллекции я определяю как subsp. exigua Eichw, хотя по отдельным признакам он приближается к “Lacerta boemica Suchow”, оценивать таксономическое значение которой я в настоящее время воздерживаюсь” (С. 66). В 1940 г. герпетологические исследования в Южном Дагестане проводил Банников (1954). По данным этого автора, прыткая, которую он определил как “Lacerta agilis exigna” (lap.) (С. 75, 79), и полосатая ящерицы симпатричны на исследованной территории и, вероятно, “конкурируют, так как в местах, где много прыткой ящерицы, – мало полосатой, и наоборот. Однако каких-либо разграничений в биотопах подметить не удалось” (С. 79).
Сухов (1948) впервые опубликовал карты ареалов зеленых ящериц фауны СССР, приведя для Дагестана все три вида. В дальнейшем новые данные о распространении зеленых ящериц в регионе были получены Хонякиной (1970, 1972), как и Шибанов указавшей для изученных особей прыткой ящерицы (выборка из окрестностей Махачкалы) морфологические признаки, общие для L. a. boemica и L. a. exigua (С. 89, Хонякина 1970). К настоящему времени мы не располагаем достоверными сведениями об обитании второго подвида в регионе, хотя на ряде карт его ареал занимает север республики (Щербак, 1966; Bischoff, 1988; Калябина-Хауф, Ананьева, 2004 и др.), и он был включен в список герпетофауны Дагестана при зоогеографическом анализе (Мазанаева, Туниев, 2011). Есть вероятность, что L. a. exigua может быть обнаружена в долине р. Кума, где прыткая ящерица известна на сопредельной территории Черноземельского р-на Калмыкии (Киреев, 1981).
Ройтберг с соавторами (Roitberg et al., 2000) подытожил предшествующие исследования хорологии и опубликовал наиболее полный на сегодняшний день кадастр находок ящериц Дагестана. Мазанаева и Орлова (2014), делая краткий обзор таксономического и экологического разнообразия заурофауны, отнесли L. agilis и L. strigata к широко распространенным в регионе видам. По их данным полосатая ящерица освоила субаридные и мезофитные биотопы низменности и предгорий, проникая местами во внутригорные районы и высокогорье. Прыткая ящерица на низменности занимает мезофитные биотопы лугово-степных и лесостепных ландшафтов, но в горах придерживается сухих биотопов. L. media в высокогорье населяет сухие горные степи и зону нагорных ксерофитов, во внутригорных районах – послелесные луга, лесостепь и горные леса.
Сведения о ландшафтно-зональном распределении зеленых ящериц во Внутригорном Дагестане содержатся в работе Мазанаевой и Аскендерова (2014): по их данным прыткая ящерица распространена в сланцевом районе, где населяет степные и лесостепные участки, а в субальпийском поясе – склоны южной экспозиции до 2300 м над ур. м.; полосатая ящерица населяет здесь степной и лесостепной пояса до 1200–1300 м, тогда как в известняковом – те же участки, что и прыткая ящерица; средняя ящерица в сланцевом районе отмечена в степном и лесостепном поясе ущелья р. Курах (до 1400 м), а в известняковом – в тех же поясах, а также в нагорных ксерофитах ущелий до 1500 м. Результаты изучения ареала L. media на территории Дагестана были опубликованы Мазанаевой и Аскендеровым (2016): ими выявлены четыре изолированные популяции вида в семиаридных котловинах крупных горных рек – Самур, Курах, Андийское Койсу и Аварское Койсу.
Перечисленные выше публикации основаны на эмпирическом методе изучения географического распространения видов. В настоящее время ареалогические (хорологические) исследования в зоологии вобрали в себя значительный арсенал ГИС-программ (см. Лисовский и др., 2020). К сожалению, среди отечественных публикаций герпетогеографических (терминология по: Боркин, Литвинчук (2013)) работ с применением ГИС пока явно недостаточно. Вслед за орнитологами (Коблик и др., 2011) мы можем сказать, что “Прикладная ареалогия – наше слабое звено” (Ананьева и др., 2019). В то же время в некоторых зарубежных публикациях зачастую даются искаженные и даже ошибочные представления о распространении зеленых ящериц на Кавказе (к примеру – Honegger, 1981; Troidl, Troidl, 2000, 2001; Ahmadzadeh et al., 2013 и др.). Вероятно, одна из причин этого кроется в ошибках при определении их видовой и подвидовой принадлежности. Последнее мы выявили при работе с коллекциями Британского музея естественной истории (www. nhm.ac.uk/our-science/collections.html), Дрезденского зоологического музея, Зенкенбергского музея естественной истории (www.sesam.senckenberg.de/) и др., в базе Global Biodiversity Information Facility (GBIF) (www.gbif.org).
Цель данной публикации – дать анализ распространения зеленых ящериц на территории Дагестана с применением геоинформационных систем и создать набор моделей потенциального пространственного распределения видов с использованием методов моделирования экологических ниш.
Актуальность исследования подчеркивает как природоохранный (L. media включена в Красную книгу РФ22 (Даревский, 2001) и Дагестана (Мазанаева, Орлова, 2009)), так и эпидемиологический (зеленые ящерицы, в том числе и на Кавказе, играют важную роль в поддержании природных очагов трансмиссивных заболеваний человека и домашних животных (Балашов, 2009; Кидов и др., 2016)) аспекты. Полученные данные могут быть использованы и для уточнения путей расселения ящериц: через территорию республики в силу физико-географических условий (меньшее развитие горного оледенения, более низкие по сравнению с Западным и Центральным Кавказом отметки высот, особенности расположения высотных растительных поясов) проходили пути фаунистического обмена между Северным Кавказом и Закавказьем (Мазанаева, Туниев, 2011), при этом явно в обоих направлениях. Кроме того, Дагестан выделяется среди других регионов Кавказа и Европы (если проводить ее южную границу по Главному Кавказскому хребту) разнообразием представленных здесь типов климата по Кеппену-Гейгеру (Peel et al., 2007), что делает эту территорию крайне удобной площадкой для биоклиматического моделирования.
МАТЕРИАЛ И МЕТОДИКА
Материалами для исследования послужили коллекции Зоологического института РАН (Санкт-Петербург, ZISP), Зоологического музея Московского государственного университета (Москва, ZMMU), Харьковского национального университета им. В.И. Каразина (Харьков, MNKhU), Национального музея естественной истории (Вашингтон, NMNH), собственные полевые исследования 1970–2020 гг., а также литературные данные. Полевые сборы второго автора охватили все административные районы Дагестана, что, в совокупности с литературными и коллекционными сведениями, позволяет говорить о достаточном объеме данных для достоверных построений.
Определение географических координат (десятичных, до четвертого знака после точки) и высоты точек регистрации ящериц в полевых условиях проводили с помощью GPS навигатора (Garmin). При работе с музейными коллекциями и литературными источниками были использованы интерактивные карты (www.wikimapia.org и www. maps.google.ru), а их локализацию определяли исходя из известных биотопических предпочтений ящериц. За одну точку был принят локалитет, удаленный от близлежащего на расстоянии не менее 1 км, что обусловлено как спецификой работы с ГИС, так и данными об индивидуальных перемещениях зеленых ящериц (Тертышников и др., 1976). Расчеты по описательной статистике выполнены с применением программы Statistica v. 7.0.
Распределение точек регистрации по территории региона показано на картах, созданных с помощью программы Quantum GIS (QGIS) v. 3.10.2 (www.qgis.org/ru/site/). На них продемонстрирован точечный (дискретный) метод построения карт – первый этап исследования географического распространения видов. На втором этапе в QGIS был реализован сеточный (растровый) метод, или метод формальных квадратов (терминология по: Воронов и др., 2002) – выделение области с заданными размерами (квадраты со стороной, соответствующей 15 км местности, т.е. площадью 225 км2), в пределах которой известна хотя бы одна точка; нами также построены так называемые тепловые карты (heatmap) с цветовой градацией количества находок каждого вида и количества видов в стандартном квадрате. При ранжировании территории мы использовали следующее цветовое обозначение: белый – отсутствие находок в пределах условного квадрата, синий – 1 точка, зеленый – 1–5 точек, красный – 5–10 точек. На тепловой карте распределения числа видов ящериц белый цвет квадрата обозначает отсутствие в нем находок, синий – присутствие 1 вида, зеленый – 2 видов, красный – 3 видов.
Отметим, что в наше время сеточный метод был применен для создания последней версии атласа амфибий и рептилий Европы (Sillero et al., 2014; https://www.seh-herpetology.org/distribution-atlas), карт ареалов зеленых ящериц в Красных книгах Краснодарского края (Островских, 2017; Туниев, Островских, 2017; Туниев Б.С., Туниев С.Б., 2017; Туниев С.Б., Туниев Б.С., 2017) и Южной Осетии (Туниев, Лотиев, 2017, 2017а). Составителями второй сводки (Замотайлов и др., 2017) было отмечено, что ареалогическая информация об особо охраняемых таксонах, публикуемая в Красной книге, должна способствовать наиболее достоверной оценке природоохранной значимости различных территорий региона. В этом плане особенно удобен сеточный метод, включающий цветовое обозначение критических мест обитания, т.к. оценки значимости часто искажены именно точечными картами ареалов слабо изученных видов, зачастую не отражающими реального экологического ареала (охватывающего заселенные места обитания) таксона в границах его географического регионального ареала. В третьем издании (Бутаева и др., 2017) такой подход был объяснен природоохранными соображениями – нежеланием обнародовать точные места нахождений таксонов, подверженных коммерческому вылову, но при этом очерки для всех редких и исчезающих представителей герпетофауны содержат точную локализацию, что купирует данный посыл.
На третьем этапе нами было проведено эколого-климатическое ГИС-моделирование ареалов ящериц с использованием 19 параметров среды (предикторов Bio 1 – Bio 19, см. табл. 2), отражающих данные по температуре и осадкам территории (Busby, 1991), а также по высотам, которые позволяют провести интерполяцию наблюдаемых данных с 1950 по 2000 гг. (Current) (Hijmans et al., 2005). Эти данные были получены из базы Worldclim (http://www.worldclime.org) в разрешении 30 угловых секунд или ~ 1 км на пиксель в районе экватора.
На начальном этапе производилась оценка значимости всех биоклиматических переменных. В дальнейшем для построения модели были исключены коррелирующие друг с другом предикторы. Для этого мы рассчитали коэффициент корреляции Пирсона для всех пар предикторов с помощью ENMTools v. 1.4.3 (Warren et al., 2010). Мы исключили переменную из коррелированной пары с |r| > 0.7, которую сочли менее биологически важной на основании известных предпочтений для изученных видов (Щербак и др., 1976; Ahmadzadeh et al., 2013; Kafash et al., 2019; данные авторов). Полученный набор данных содержал 6 биоклиматических предикторов: Bio 2 (cреднесуточная амплитуда, среднее для месяца, max–min), Bio 6 (минимальная температура наиболее холодного месяца, ×10, °С), Bio 7 (годовой размах температур, ×10, °С), Bio 8 (средняя температура наиболее влажного сезона, ×10, °С), Bio 10 (средняя температура наиболее жаркого сезона. ×10, °С), Bio 14 (осадки наиболее сухого месяца, мм), Bio 15 (сезонность осадков, CV) и Bio 19 (осадки наиболее холодного сезона, мм).
С помощью программы ENMTools были удалены локалитеты, близко расположенные друг к другу и находящиеся в одном и том же квадрате (1 × 1 км). При моделировании применили программу MaxEnt (Maximum Entropy Species Distribution Modelling) v. 3.4.1 (www.cs.princeton.edu/~schapire/maxent/), в которой экстраполяция пространственного распределения биологических видов проводится методом максимальной энтропии (Elith et al., 2011). С ее помощью созданы карты наиболее вероятных областей распространения (пригодности местообитаний) ящериц и определен вклад (в %) каждого фактора в построение модели.
Чтобы правильно параметризовать модели, мы оценили выполнение различных комбинаций из десяти вариантов настройки MaxEnt (от 0.5 до 5.0 с шагом 0.5; см. Vences et al., 2017). Самый подходящий вариант модели был параметризован с коэффициентом регуляризации 1.0 (30 повторов). Нами использованы настройки по умолчанию, т.е. задействованы все классы пространственных объектов, максимальное количество итераций (500) и максимальное количество фоновых точек (10 000) (Phillips, Dudík, 2008). Для тестирования полученных моделей использовали 25% точек. Мы применили метод “складного ножа” (jackknife analysis) для оценки относительного вклада переменных в модель MaxEnt.
Достоверность модели оценивали с использованием встроенной функции построения кривой AUC (area under receiver operating characteristic (ROC) curve), отражающей чувствительность вида к параметрам и его специфичность. Значение индекса AUC выше 0.75 и близкое к единице означает высокую степень достоверности (Elith, 2002). Визуализацию карт MaxEnt провели с помощью программы DIVA-GIS v. 7.5.0 (www.diva-gis.org) (Scheldeman, van Zonneveld, 2010).
Для оценки перекрывания экологических (эколого-климатических) ниш при помощи ENMTools был вычислен индекс Шенера (Schoener’s D) (D) и стандартизированное расстояние Хеллингера (Hellinger’ s I) (I) значение которых статистически измеряется от нуля – в случае полного отсутствия перекрывания ниш, до единицы – в случае их идентичности; по этой причине этот показатель получил название теста идентичности, I-test. Его рекомендуется проводить со всеми предикторами в 100-кратной повторности (100 реплик) (Warren et al., 2010). Высокая эффективность использования индекса D ранее была показана на примере партеногенетических и двуполых видов ящериц рода Darevskia 1999 (Petrosyan et al., 2019, 2020).
РЕЗУЛЬТАТЫ
По состоянию на 01.09.2020 кадастр находок зеленых ящериц на территории Дагестана включает 361 точку регистрации (L. agilis – 78, L. media – 30, L. strigata – 253) (приложение 1 – http://sev-in.ru/ru/zoologiceskii-zurnal). По источнику информации (табл. 1) более половины (188 точек, или 52.1%) получены только по результатам наблюдений авторов и ранее не были опубликованы. Показательно, что данные по находкам средней ящерицы были обнародованы на 100%, что связано с повышенным вниманием специалистов к этому виду, занесенному в региональную и федеральную Красные книги.
Таблица 1.
Распределение точек регистрации ящериц рода Lacerta на территории Дагестана по источнику информации
| Вид | Источник информации | |||||||
|---|---|---|---|---|---|---|---|---|
| Общее колич. точек | Наблюдения авторов, % от общего колич. | Коллекционные данные, % от общего колич. | Литературные данные, % от общего колич. | 3 + 4 | 3 + 5 | 4 + 5 | 3 + 4 + 5 | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| L. agilis | 78 | 34 (43.6) | 4 (5.1) | 7 (9) | 1 (1.3) | 25 (32.1) | 4 (5.1) | 3 (3.8) |
| L. media | 30 | 0 | 0 | 0 | 4 (13.3) | 21 (70) | 0 | 5 (16.7) |
| L. strigata | 253 | 154 (60.9) | 22 (8.7) | 18 (7.1) | 3 (1.2) | 35 (13.8) | 7 (2.8) | 14 (5.5) |
| Итого | 361 | 188 (52.1) | 26 (7.2) | 25 (6.9) | 8 (2.2) | 81 (22.4) | 11 (3.1) | 22 (6.1) |
Таблица 2.
Высотное распределение точек регистрации ящериц рода Lacerta на территории Дагестана
| Вид | n | Диапазон распространения, м над ур. м. | Среднее значение (μ) | Стандартное отклонение (σ) | Коэффициент вариации (CV, %) |
|---|---|---|---|---|---|
| L. agilis | 78 | –30–2300 | 914.61 | 737.09 | 80.59 |
| L. media | 30 | 560–1710 | 1278.66 | 253.91 | 19.85 |
| L. strigata | 253 | –30–1780 | 281.60 | 356.70 | 126.66 |
| Общее | 361 | –30–2300 | 501.23 | 575.18 | 114.75 |
Диапазон населяемых видами высот варьирует от –30 м – популяции прыткой и полосатой ящериц на побережье Каспийского моря (минимальные показатели для этих видов в целом по всему ареалу) до 2300 м над ур. м. – популяция прыткой ящерицы в окрестностях с. Шари Агульского р-на (ZISP № 22 021; Калябина-Хауф, Ананьева, 2004; данные Л.Ф. Мазанаевой, 2005, 2007). Высотное распределение находок представлено в табл. 2, из которой следует, что в условиях северо-востока Кавказа по средним показателям L. strigata (диапазон –30–1780 м, μ – 281.60) может быть ассоциирована с зоной равнин (по: Гурлев, 1972), L. agilis (–30–2300 м, μ – 914.61) – предгорий, L. media (560–1710 м, μ –1278.66) – гор.
На рис. 1 для L. agilis отмечен максимальный охват высот – это единственный вид, присутствующий во всех высотных диапазонах (на гистограмме был выбран промежуток между делениями = 150 м). Наибольшее количество находок зеленых ящериц (143 точки, или 39.7%), сделано на высотах от –30 до 120 м, что демонстрирует их высотный преферендум в условиях Дагестана. Аналогичные цифры характерны для прыткой и полосатой ящериц, при этом для второго вида на этих отметках известно 127 точек (50.4%). Большинство локалитетов средней ящерицы – 9 (30%) известно на высотах 1170–1320 м.
Рис. 1.
Высотное распределение точек регистрации (n = 361) ящериц рода Lacerta на территории Дагестана.

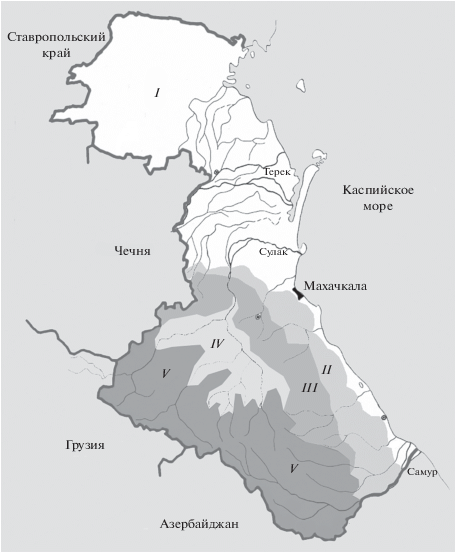
При сопоставлении точечных карт (рис. 2) с картой ландшафтных зон Дагестана (рис. 3) мы видим, что прыткая и полосатая ящерицы обнаружены во всех пяти зонах: низменности, предгорий, передовых хребтов, внутригорного и высокогорного Дагестана. Наибольшее число находок L. agilis приходится на третью из них, а L. strigata – на первую и вторую; L. media обнаружена в четвертой и пятой с количественным преобладанием в высокогорной зоне. Это соответствует анализу высотного распределения точек регистрации видов.
Рис. 2.
Карты точек регистрации ящериц рода Lacerta на территории Дагестана: A – L. agilis, B – L. media, C – L. strigata.
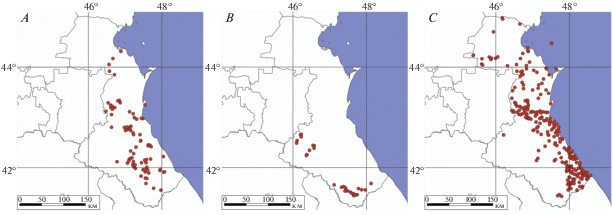
Рис. 3.
Карта ландшафтных зон Дагестана: I – низменности, II – предгорья, III – передовые хребты, IV– внутригорья, V – высокогорья (по: Федина, 1963; из: Мазанаева, Туниев, 2011, с изменениями).
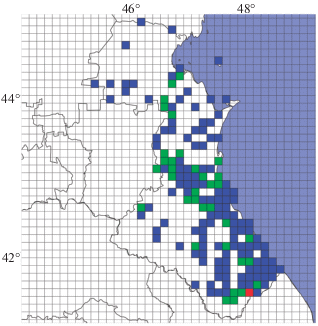
На сеточных картах (рис. 4), где территория региона покрыта 421 полным квадратом33, 150 (35.6%) включают хотя бы одну находку представителей Lacerta. Наибольшее распространение в регионе получила полосатая ящерица – она зарегистрирована в 117 (27.8%) квадратах и выступает фоновым видом; прыткая известна в 51 (12.1%), а средняя только в 17 (4%) (табл. 3). Для L. agilis характерно отсутствие областей с максимальным числом находок: наибольшее – 4 локалитета, отмечено в верховьях рек Казикумухское Койсу и Хунних; для L. media единственный красный растр выявлен в среднем течение р. Самур, L. strigata – в верховьях р. Сулак в районе Чиркейского водохранилища и в прибрежной зоне Каспия в низовье р. Уллучай.
Рис. 4.
Карты распределения числа точек регистрации ящериц рода Lacerta при сеточном (растровом) методе разделения территории Дагестана: A – L. agilis; B – L. media; C – L. strigata. Цветная заливка обозначает количество точек регистрации в стандартном квадрате 15 × 15 км: белая – 0, синяя – 1, зеленая – 1–5, красная – 5–10.

Таблица 3.
Распределение числа находок ящериц рода Lacerta при сеточном (растровом) методе разделения территории Дагестана
| Вид | Число квадратов (%) с находками ящериц | |||
|---|---|---|---|---|
| белых (0 точек) | синих (1 точка) | зеленых (1–5 точек) | красных (5–10 точек) | |
| L. agilis | 370 (87.9) | 35 (8.3) | 16 (3.8) | 0 |
| L. media | 404 (96) | 9 (2.1) | 7 (1.7) | 1 (0.2) |
| L. strigata | 304 (72.2) | 52 (12.3) | 63 (15) | 2 (0.5) |
На полученной карте видового обилия (рис. 5) максимальный показатель отмечен только для одного (0.2%) квадрата, расположенного в долине р. Курах – небольшой аридной котловине с растительностью средиземноморского типа; с присутствием одного вида – 116 (27.6%), двух видов – 33 (7.8%). “Пустые” квадраты с нулевыми показателями сосредоточены, главным образом, на севере региона в Терско-Кумской низменности и на юго-западе в районе высокогорных водораздельных хребтов и плато. Вероятнее всего это отражает реальное отсутствие здесь популяций ящериц в силу непригодности территории: низменность характеризуется недостаточным увлажнением (самая засушливая часть республики) и очень жарким летом (самые высокие среднегодовые температуры), а высокогорья – резко континентальным климатом с холодной зимой и прохладным летом (самые низкие среднегодовые температуры).
Рис. 5.
Карта распределения числа видов ящериц рода Lacerta при сеточном (растровом) методе разделения территории Дагестана. Цветная заливка обозначает количество видов, отмеченных в стандартном квадрате 15 × 15 км: белая – 0 видов; синяя – 1, зеленая – 2, красная – 3.
При моделировании распространения видов с помощью программы MaxEnt были получены хорошие результаты производительности модели потенциального распределения (индекс AUC = = 0.98–0.99). По этим данным карты надежно характеризуют особенности распространения изученных видов и позволяют нам в совокупности с новым материалом дать свои замечания и дополнения.
Наиболее значимый вклад в построение моделей (≥10%) современных ареалов всех трех видов внесли минимальная температура наиболее холодного месяца (Bio 6), количество осадков наиболее сухого месяца (Bio 14) и количество осадков наиболее холодного сезона (Bio 19) (табл. 4). Суммарный вклад этих факторов в формирование модели достигает 76.2%. Для L. agilis и L. media в число доминирующих предикторов попал годовой размах температур (Bio 7); только для L. media – сезонность осадков (Bio 15). Таким образом, при наличии значительного числа общих показателей, каждый вид в условиях северо-востока Кавказа продемонстрировал индивидуальный набор доминирующих при построении модели параметров. Обращает на себя внимание отсутствие влияния высоты местности на построение карт.
Таблица 4.
Значимость задействованных биоклиматических параметров (%) в построении моделей распространения (пригодности местообитаний) ящериц рода Lacerta на территории Дагестана
| Параметр | Аббревиатура | Вид | |||||
|---|---|---|---|---|---|---|---|
| L. agilis | L. media | L. strigata | |||||
| Вклад | Пермутация | Вклад | Пермутация | Вклад | Пермутация | ||
| Среднесуточная амплитуда (среднее для месяца, max–min) | BIO 2 | 5.6 | 26.2 | 5.1 | 43.8 | 8.2 | 33 |
| Минимальная температура наиболее холодного месяца (×10, °С) | BIO 6 | 11.1 | 0.5 | 11.9 | 2.4 | 39.3 | 41.1 |
| Годовой размах температур (×10, °С) | BIO 7 | 32.1 | 33.5 | 29.4 | 37.5 | 8.7 | 0.1 |
| Средняя температура наиболее влажного сезона (×10, °С) | BIO 8 | 1.9 | 0 | 6.1 | 0.2 | 0.8 | 0.1 |
| Средняя температура наиболее жаркого сезона (×10, °С) | BIO 10 | 0.1 | 0 | 2.9 | 0.1 | 1.7 | 0 |
| Осадки наиболее сухого месяца, мм | BIO 14 | 11.8 | 0.1 | 11.8 | 0 | 16.9 | 5 |
| Сезонность осадков (CV) | BIO 15 | 1.7 | 0.6 | 19.6 | 2.8 | 4.4 | 0.7 |
| Осадки наиболее холодного сезона, мм | BIO 19 | 35.7 | 39.1 | 13.2 | 13.2 | 20 | 20 |
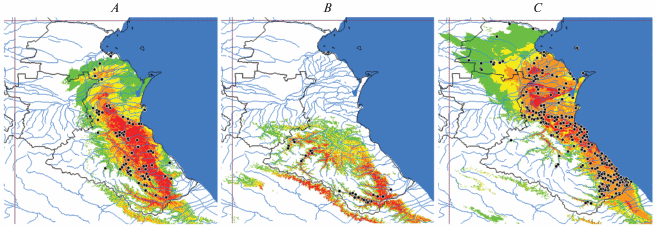
На полученных ГИС-картах (рис. 6) наибольшая распространенность, т.е. доля территории, на которой мы ожидаем наличие вида, отмечена для полосатой ящерицы. Прыткая незначительно уступает ей в этом показателе, при этом число ее фактических находок меньше в 3.2 раза. Выявлены различия в расположении территорий, максимально пригодных для них, которые можно обозначить как ядра ареалов: для L. strigata – это низменности и предгорья, L. agilis – предгорья и передовые хребты.
Рис. 6.
Карты прогнозируемых областей географического распространения ящериц (SDM-карты) рода Lacerta на территории Дагестана, построенные с помощью программы MaxEnt v. 3.4.1: A – L. agilis; B – L. media; C – L. strigata. Цветная заливка обозначает области вероятного распространения (пригодности местообитаний) с высокой (100–80%; красный цвет), средней (80–60%; оранжевый цвет), низкой (60–40%; желтый цвет) и крайне низкой (40–20%; зеленый цвет) вероятностью встречи; черные точки – места находок.
Перекрывание экологических (эколого-климатических) ниш в нашем исследовании минимально для средней ящерицы со всеми изученными видами (табл. 5), с которыми по данным полевых исследований она симпатрична в единичных локалитетах. Максимальное перекрывание отмечено для совместно обитающих на значительной площади региона (Roytberg, 1994) прыткой и полосатой ящериц (0.52/0.78).
ОБСУЖДЕНИЕ
Несмотря на значительные отметки высот, ни один из трех изученных видов не достигает в Дагестане предела своего распространения, известного на Кавказе: L. agilis ≈ 2730 м в районе Ардаганского перевала в Турции (Туниев и др., 2014), L. media ≈ 2700 м в окрестностях горы Аг-юрт в Нахичеванской АР Азербайджана (Кузьмин, 1981), L. strigata ≈ 2100 м на горе Ара (= Аралер, Араилер) в Армении (ZISP № 20 216).
В отличие от Закавказья (Рудик, 1986), на Северном Кавказе совместное обитание всех трех видов зеленых ящериц ранее было неизвестно. В ходе полевых исследований в долине р. Курах 28.07.2003 нами выявлена зона их симпатрии: полосатая ящерица (2–3 экз./200 м учетного маршрута) встречена между с. Кутул и с. Куказ на участке, где шибляк образует сомкнутые заросли по северным склонам ближе к речной долине. Средняя ящерица (1–4 экз./500 м учетного маршрута) была обнаружена между с. Куказ и с. Курах на северных склонах с изреженными шибляком и трагакантником, а также на участке ущелья между с. Курах и с. Хута с горно-степной растительностью; на склоне с горно-луговой растительностью этой части ущелья отмечена прыткая ящерица (1 экз./500 м). Учитывая высокую научную ценность обозначенной территории, мы рекомендуем расширить границы Государственного природного заказника “Касумкентский” в юго-западном направлении, что будет способствовать сохранению заурофауны региона.
Согласно ГИС-картам перспективной территорией для поисков аналогичных территорий может быть признано среднее и верхнее течение рек Аварское Койсу (c притоками Каракойсу и Казикумухское Койсу) и Андийское Койсу. Так, в окрестностях с. Ботлих 03.07.2002 были обнаружены популяции L. media и L. strigata, а высокое ценотическое разнообразие этого района (широколиственные леса, луговые степи, субальпийские луга) (Абдулаев и др., 2011) позволяет предполагать обитание здесь и L. agilis. Показательно, что при герпетогеографическом районировании республики данная территория была отнесена к Внутригорному округу известнякового Дагестана – наиболее оригинальному выделу в пределах всего Большого Кавказа, представляющему значительную научную и природоохранную ценность (Мазанаева, Туниев, 2011).
Говоря о прогностической роли ГИС-карт, отметим, что на первом этапе моделирования мы не располагали сведениями о находках прыткой ящерицы в долине р. Курах, но эта территория вошла в область с максимальными показателями пригодности для вида.
Неразрешенным вопросом остается обитание прыткой ящерицы в долине р. Самур: для низовий этой реки вид был приведен в публикации Хонякиной (1970), Щербака с соавторами (1976) и Ройтберга с соавторами (Roitberg et al., 2000), при этом во второй работе локалитет дан со ссылкой на данные Хонякиной, а в третьей – на статью Банникова (1954), в которой, в действительности, нет указаний на точные места наблюдений за ящерицами. Сборы вида с этой территории отсутствуют во всех известных нам коллекциях. На полученной ГИС-карте дельта Самура не вошла в число благоприятных территорий, но высокая пригодность выявлена в среднем и верхнем течении этой реки, в долинах рек Ахтычай и Усухчай, а также на сопредельной территории Азербайджана от верхнего и среднего течения р. Кусарчай до р. Вельвеличай. Эта территория уже была включена в ареал прыткой ящерицы (Щербак и др., 1976), но без достоверного подтверждения этого: к сожалению, кадастр в монографии “Прыткая ящерица” был составлен с многочисленными ошибками. Новые находки L. agilis ожидаемы и в низовьях р. Терек, в долине рек Аварское Койсу и Андийское Койсу.
В коллекции ZMMU (№ 2014) хранится сбор L. agilis, сделанный Б.А. Красавцевым в с. Тушиловка Кизлярского р-на 25.06.1932. В настоящее время вид здесь не обнаружен (полевые исследования 24.05.2006 и 18.06.2010), а на ГИС-карте (SDM) эта точка, выступающая самой северной в ареале L. a. boemica, выпала из пригодной области. Можно предположить вымирание этой популяции, находящейся в зоне пессимума на границе ареала.
С высокой вероятностью обнаружение полосатой ящерицы возможно в долине рек Аварское Койсу и Андийское Койсу, что было отмечено и для прыткой ящерицы; в этом случае ГИС-карта указывает на вероятные пути проникновения этих видов во внутригорную и высокогорную зоны. Кроме того, необходим поиск ящериц в долине р. Самур выше с. Ахты. Подробный анализ модели распространения L. media был дан нами ранее (Доронин и др., 2018).
Интенсивное развитие ГИС и их внедрение в зоологию вновь поднимает один из ключевых вопросов – что определяет границу видового ареала? Если следовать подходу современных исследователей экологических ниш (см: Hargreaves et al., 2014; Lee-Yaw et al., 2016), выбор должен быть сделан между двумя основными ответами – ограничением расселения из-за изоляционных барьеров или же ухудшением условий обитания, включая климатические. В нашем случае, исходя из полученных данных, в частности – отсутствия зависимости построения модели от высот, предпочтение должно быть отдано второму варианту.
Нельзя не учитывать и временно́й (исторический) фактор. Наиболее показателен пример L. media: на ее филогенетическом древе популяции с территории Дагестана входят, согласно нашим неопубликованным данным, в кладу Армянское нагорье – Большой Кавказ, появление которой относят к завершающему этапу филогенеза вида и датируют около 1 млн лет назад (нижний плейстоцен), а дивергенцию внутри нее – менее 200 тыс. лет назад (средний плейстоцен) (Ahmadzadeh et al., 2013a; Sagonas et al., 2014). Возможно, данный вид, проникнув из Закавказья на территорию Дагестана, не успел расселиться на Северном Кавказе, хотя благоприятные для него климатические условия выявлены на сопредельной территории Чечни и в районе Кавказских Минеральных вод (Доронин и др., 2018). С другой стороны, существующие ранее северокавказские популяции могли исчезнуть из анклавов во время последнего ледникового (гляциального) максимума около 22 тыс. лет назад, после чего L. media не смогла восстановить прежний ареал.
В дальнейшем необходимо сопоставить MaxEnt-модели ареалов ящериц, построенных только на основе точек с территории Дагестана, с моделями, охватывающими Кавказ, или всю область их распространения. Это позволит более детально осветить региональные особенности изученных нами популяций, дать оценку чувствительности ГИС к изменению анализируемого объема первичных данных – локалитетов, как это уже было сделано при моделировании ареала болотной черепахи (Emys orbicularis (Linnaeus 1758)) (Дуйсебаева и др., 2019). Здесь мы можем ожидать различий в показателях перекрывания ниш зеленых ящериц в различных частях ареалов. При этом, если они обитают в различающихся климатических условиях, то очевидно, что показатель сходства экологических ниш популяций одного вида может иметь низкое значение, но это еще не свидетельствует об их эколого-климатической дивергенции, т.к. они могут обитать в разных условиях за счет экологической пластичности (см. Zink, 2015).
Список литературы
Абдулаев К.А., Атаев З.В., Братков В.В., 2011 Современные ландшафты Горного Дагестана. Махачкала: ДГПУ. 116 с.
Ананьева Н.Б., Доронин И.В., Мазанаева Л.Ф., 2019. VII съезд Герпетологического общества им. А.М. Никольского при РАН // Известия высших учебных заведений. Поволжский регион. Естественные науки. № 1 (25). С. 5–11.
Балашов Ю.С., 2009. Паразитизм клещей и насекомых на наземных позвоночных. СПб.: Наука. 357 с.
Банников А.Г., 1954. Материалы по биологии земноводных и пресмыкающихся Южного Дагестана // Ученые записки Московского городского педагогического института имени В.П. Потемкина. Т. 28. С. 75–88.
Боркин Л.Я., Литвинчук С.Н., 2013. Амфибии Палеарктики: таксономический состав // Труды Зоологического института РАН. Т. 317. № 4. С. 494–541.
Бутаева Ф.Г., Комаров Ю.Е., Лотиев К.Ю., 2017. Введение. Отв. ред.: Кабулов А.З. Красная книга Республики Южная Осетия. Нальчик: Издательство М. и В. Котляровых (ООО “Полиграфсервис и Т”). С. 8–11.
Воронов А.Г., Дроздов Н.Н., Криволуцкий Д.А., Мяло Е.Г., 2002. Биогеография с основами экологии. М.: Издательство Московского университета; Издательство “Высшая школа”. 392 с.
Гурлев И.А., 1972. Природные зоны Дагестана. Махачкала: Дагучпедгиз. 210 с.
Даревский И.С., 2001. Средняя ящерица – Lacerta media Lantz et Cyren, 1920. В.И. Данилов-Данильян и др. (Гл. редкол.). Красная книга Российской Федерации (животные). М.: АСТ-Астрель. С. 333–334.
Доронин И.В., Ананьева Н.Б., Барабанов А.В., Мильто К.Д., Хайрутдинов И.З., 2017. К вопросу о типовых экземплярах амфибий и рептилий из коллекции Эдуарда Эйхвальда // Материалы юбилейной отчетной научной сессии, посвященной 185-летию Зоологического Института РАН. Санкт-Петербург, Россия 13–16 ноября 2017. СПб.: ЗИН РАН. С. 77–80.
Доронин И.В., Мазанаева Л.Ф., Доронина М.А., 2018. Использование ГИС-моделирования для анализа распространения средней ящерицы, Lacerta media Lantz et Cyrén, 1920, на территории Дагестана (Россия) // Труды Зоологического института РАН. Т. 322. № 4. С. 463–480.
Дуйсебаева Т.Н., Доронин И.В., Малахов Д.В., Кукушкин О.В., Бакиев А.Г., 2019. ГИС-анализ распространения и условий обитания Emys orbicularis оrbicularis (Testudines, Emydidae): методические аспекты // Известия высших учебных заведений. Поволжский регион. Естественные науки. № 1 (25). С. 28–40.
Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. и др., 2017. Введение. Отв. ред.: Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. Красная книга Краснодарского края. Животные. Адм. Краснодар. края. 3-е изд. Краснодар. С. 9–18.
Калябина-Хауф С.А., Ананьева Н.Б., 2004. Филогеография и внутривидовая структура широкоареального вида ящериц Lacerta agilis L., 1758 (Lacertida, Sauria, Reptilia) (опыт использования митохондриального гена цитохрома b). СПб. 108 с. (Труды Зоологического института РАН, т. 302).
Кидов А.А., Коврина Е.Г., Тимошина А.Л., Матушкина К.А., 2016. Паразито-хозяинные отношения иксодового клеща (Ixodes ricinus L.) и терской ящерицы (Lacerta boemica S.) в Северной Осетии // Известия Тимирязевской сельскохозяйственной академии. Вып. 5. С. 81–90.
Киреев В.А., 1981. Земноводные и пресмыкающиеся Калмыкии. Дис. ... канд. биол. наук. Киев. 235 с.
Коблик Е.А., Лаппо Е.Г., Редькин Я.А., Томкович П.С., Калякин М.В., 2011. Прикладная ареалогия – наше слабое звено // Зоологический журнал. Т. 90. № 7. С. 835–845.
Красовский Д.Б., 1932. Материалы к познанию фауны наземных позвоночных Рутульского кантона Дагестанской АССР // Известия 2-го Северо-Кавказского педагогического института. Т. 9. С. 185–218.
Кузьмин С.Л., 1981. К экологии ящериц Нахичеванской АССР // Пятая Всесоюзная герпетологическая конференция. Вопросы герпетологии. Авторефераты докладов, Ашхабад, 22–24 сентября 1981 г. Л.: Наука, Ленинградское отделение. С. 78–79.
Лисовский А.А., Дудов С.В., Оболенская Е.В., 2020. Преимущества и ограничения методов экологического моделирования ареалов. 1. Общие подходы // Журнал общей биологии. Т. 81. № 2. С. 123–134.
Мазанаева Л.Ф., Аскендеров А.Д., 2014. Ландшафтно-зональное распределение амфибий и рептилий во внутригорном Дагестане // Вестник Дагестанского научного центра. № 54. С. 53–58.
Мазанаева Л.Ф., Аскендеров А.Д., 2016. К распространению и экологии средней ящерицы Lacerta media Lantz et Cyren, 1920 (Reptilia: Sauria: Lacertidae) в Дагестане // Вестник Санкт-Петербургского университета. Серия 3. Биология. Вып. 3. С. 93–99.
Мазанаева Л.Ф., Орлова В.Ф., 2009. Средняя ящерица Lacerta media Lantz et Cyrén, 1920. Отв. Ред. и сост.: Абдурахманов Г.М. Красная книга Республики Дагестан. Махачкала. С. 382–383.
Мазанаева Л.Ф., Орлова В.Ф., 2014. Таксономическое и экологическое разнообразие ящериц Дагестана // Горные экосистемы и их компоненты: Материалы V Всерос. конференции с международным участием, посвященной 25-летию научной школы чл.-корр. РАН А.К. Темботова и 20-летию Института экологии горных территорий им. А.К. Темботова КБНЦ РАН. Нальчик. С. 74–75.
Мазанаева Л.Ф., Туниев Б.С., 2011. Зоогеографический анализ герпетофауны Дагестана // Современная герпетология. Т. 11. Вып. 1/2. С. 55–76.
Островских С.В., 2017. Ящерица полосатая Lacerta strigata Eichwald, 1831 Отв. ред.: Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. Красная книга Краснодарского края. Животные. Адм. Краснодар. края. 3-е изд. Краснодар. С. 493–494.
Рудик А.М., 1986. О находке полосатой ящерицы (Lacerta strigata Eichwald) на Черноморском побережье Кавказа // Герпетологические исследования на Кавказе. Л.: ЗИН АН СССР. С. 187–188. (Труды ЗИН АН СССР, т. 158).
Сухов Г.Ф., 1948. Обзор ящериц подрода Lacerta (Sauria), встречающихся в СССР // Сборник работ по систематике, зоогеографии и экологии. М. Л.: Из-во АН СССР. С. 101–117. (Труды ЗИН АН СССР. Т. 7. Вып. 3).
Тертышников М.Ф., Щепотьев Н.В., Рашкевич Н.А., Стрельцов А.Б., Булахов В.Л., Константинова Н.Ф., Жаркова В.К., 1976. Глава IX. Перемещение и миграции. Прыткая ящерицы. Монографическое описание вида. М.: Наука. С. 246–251.
Туниев Б.С., Лотиев К.Ю., 2017. Ящерица прыткая Lacerta agilis Linnaeus, 1758. Отв. ред.: Кабулов А.З. Красная книга Республики Южная Осетия. Нальчик: Издательство М. и В. Котляровых (ООО “Полиграфсервис и Т”). С. 225–226.
Туниев Б.С., Лотиев К.Ю., 2017а. Ящерица средняя Lacerta media Lantz et Cyren, 1920. Отв. ред.: Кабулов А.З. Красная книга Республики Южная Осетия. Нальчик: Издательство М. и В. Котляровых (ООО “Полиграфсервис и Т”). С. 226–227.
Туниев Б.С., Островских С.В., 2017. Ящерица средняя Lacerta media Lantz et Cyren, 1920. Отв. ред.: Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. Красная книга Краснодарского края. Животные. Адм. Краснодар. края. 3-е изд. Краснодар. С. 492–493.
Туниев Б.С., Туниев С.Б., 2017. Ящерица прыткая грузинская Lacerta agilis grusinica Peters, 1960. Отв. ред.: Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. Красная книга Краснодарского края. Животные. Адм. Краснодар. края. 3-е изд. Краснодар. С. 490–491.
Туниев Б.С., Туниев С.Б., Авджи А., Ильгаз Ч., 2014. Герпетологические исследования в восточной и северо-восточной Турции // Современная герпетология. Т. 14. Вып. 1/2. С. 44–53.
Туниев С.Б., Туниев Б.С., 2017. Ящерица прыткая мзымтинская Lacerta agilis mzymtensis (Tuniyev S. et Tuniyev B., 2008). Отв. ред.: Замотайлов А.С., Лохман Ю.В., Вольфов Б.И. Красная книга Краснодарского края. Животные. Адм. Краснодар. края. 3-е изд. Краснодар. С. 491–492.
Федина А.Е., 1963. Основные закономерности ландшафтной дифференциации горного Дагестана и их влияние на хозяйственное использование территории // Вопросы ландшафтоведения. Алма-Ата: Изд-во АН КазССР. С. 5–96.
Хонякина З.П., 1970. Материалы по биологии прыткой ящерицы Дагестана // Исследования по зоологии и паразитологии в Дагестане. Сборник научных сообщений кафедры зоологии Дагестанского государственного университета. Махачкала. С. 88–93.
Хонякина З.П., 1972. Полосатая и средняя ящерицы Дагестана // Исследования по зоологии и паразитологии в Дагестане. Сборник научных сообщений кафедры зоологии Дагестанского государственного университета. Махачкала. С. 150–159.
Шибанов Н.В., 1935. Материалы к фауне рептилий Дагестана // Сборник трудов Государственного Зоологического музея (при МГУ). Т. II. С. 63–68.
Щербак Н.Н., 1966. Земноводные и пресмыкающиеся Крыма (Herpetologia Taurica). Киев: Наукова думка. 239 с.
Щербак Н.Н., Осташко Н.Г., Даревский И.С., Баранов А.С., Андрушко А.М., Ведмедеря В.И., Гаранин В.И., Ищенко В.Г., Лукина Г.П., Окулова Н.М., Рашкевич Н.А., Тертышников М.Ф., Топорков Л.Я., Хонякина З.П., Швецов Ю.Г., Щербань М.И., 1976. Глава I. Ареал // Прыткая ящерица. Монографическое описание вида. Отв. ред.: А.В. Яблоков. М.: Наука. С. 9–52.
Ahmadzadeh F., Flecks M., Carretero M.A., Böhme W., Ilgaz C., Engler J.O., Harris D.J., Üzüm N., Rödder D., 2013. Rapid lizard radiation lacking niche conservatism: ecological diversification within a complex landscape. Journal of Biogeography. V. 40. № 9. P. 1807–1818.
Ahmadzadeh F., Flecks M., Rödder D., Böhme W., Ilgaz Ç., Harris D.J., Engler J.O., Üzüm N., Carretero M.A., 2013a. Multiple dispersal out of Anatolia: Biogeography and evolution of oriental green lizards // Biological Journal of the Linnean Society. V. 110. № 2. P. 398–408.
Bischoff W., 1988. Zur Verbreitung und Systematik der Zauneidechse, Lacerta agilis Linnaeus, 1758 // Mertensiella. № 1. P. 11–30.
Busby J.R., 1991. BIOCLIM – a bioclimate analysis and prediction system // Plant Protection Quarterly. V. 6. P. 8–9.
Claudia A., Franke F., Bleidorn C., Bernhard D., Schlegel M., 2014. Phylogenetic analysis of the Lacerta agilis subspecies complex // Systematics and Biodiversity. V. 12. № 1. P. 43–54.
Eichwald E., 1834. Reise auf dem Caspischen Meere und in den Caukasus. Unternommen in den Jahren 1825–1826. Erster band. Stuttgart: Verlag der J.G. Gotta’schen Buchhandlung. 472 s.
Elith J., 2002. Quantitative methods for modeling species habitat: comparative performance and an application to Australian plants // Quantitative methods for conservation biology. N.Y.: Springer. P. 39–58.
Elith J., Phillips S.J., Hastie T., Dudík M., Chee Y.E., Yates C.J., 2011. A statistical explanation of MaxEnt for ecologists // Divers. Distrib. V. 17. № 1. P. 43–57.
Ficetola G.F., Falaschi M., Bonardi A., Padoa-Schioppa E., Sindaco R. 2018. Biogeographical structure and endemism pattern in reptiles of the Western Palearctic // Progress in Physical Geography. V. 42. № 2. P. 220–236.
Hargreaves A.L., Samis K.E., Eckert C.G., 2014. Are species’ range limits simply niche limits writ large? A review of transplant experiments beyond the range // The American Naturalist. V. 183. № 2. P. 157–173.
Hijmans R.J., Cameron S.E., Parra J.L., Jones P.G., Jarvis A., 2005. Very high resolution interpolated climate surfaces for global land areas // International journal of Climatology. V. 25 P. 1965–1978.
Honegger R.E., 1981. Threatened amphibians and reptiles in Europe. Supplementary volume of Hanbuch der Reptilien und Amphibien Europas. Wiesbaden: Akademische Verlagsgesellschaft. 158 p.
Kafash A., Ashrafi S., Ohler A., Schmidt B.R., 2019. Environmental predictors for the distribution of the Caspian green lizard, Lacerta strigata Eichwald, 1831, along elevational gradients of the Elburz Mountains in northern Iran // Turkish Journal of Zoology. V. 43. P. 106–113.
Kornilios P., Thanou E., Lymberakis P., Ilgaz Ç., Kumlutaş Y., Leaché A., 2019. A phylogenomic resolution for the taxonomy of Aegean green lizards // Zoologica Scripta. V. 49. № 1. P. 14–27.
Lee-Yaw J.A., Kharouba H.M., Bontrager M., Mahony C., Csergo A.M., Noreen A.M.E., Li Q., Schuster R., Angert A.L., 2016. A synthesis of transplant experiments and ecological niche models suggests that range limits are often niche limits // Ecology Letters. V. 19. P. 710–722.
Marzahn E., Mayer W., Joger U., Ilgaz Ç., Jablonski D., Kindler C., Kumlutaş Y., Nistri A., Schneeweiss N., Vamberger M., Žagar A., Fritz U., 2016. Phylogeography of the Lacerta viridis complex: mitochondrial and nuclear markers provide taxonomic insights // Journal of Zoological Systematics and Evolutionary Research. V. 54. № 2. P. 85–105.
Peel M.C., Finlayson B.L., McMahon T.A., 2007. Updated world map of the Köppen-Geiger climate classification // Hydrology and Earth System Sciences. V. 11. P. 1633–1644.
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Nazarenko E., Omelchenko A., Danielyan F., Arakelyan M., 2019. Analysis of geographical distribution of the parthenogenetic rock lizard Darevskia armeniaca and its parental species (D. mixta, D. valentini) based on ecological modeling // Salamandra. V. 55. № 3. P. 173–190.
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Omelchenko A., Varshavskiy A., Danielyan F., Arakelyan M., 2020. Species Distribution Models and Niche Partitioning among Unisexual Darevskia dahli and Its Parental Bisexual (D. portschinskii, D. mixta) Rock Lizards in the Caucasus // Mathematics. V. 8. № 1329. P. 1–21.
Phillips S.J., Dudík M., 2008. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation // Ecography. V. 31. № 2. P. 161–175.
Roitberg E.S., Mazanaeva L.F., Ilyina E.V., Orlova V.F., 2000. Die Echsen Dagestans (Nordkaukasus, Russland): Artenliste und aktuelle Verbreitungsdaten (Reptilia: Sauria: Gekkonidae, Agamidae, Anguidae, Scincidae et Lacertidae) // Faunistische Abhandlungen, Staatliches Museum für Tierkunde Dresden. Bd. 22. Hf. 8. S. 97–118.
Sagonas K., Poulakakis N., Lymberakis P., Parmakelis A., Pafilis P., Valakos E.D., 2014. Molecular systematics and historical Biogeography of the green lizards (Lacerta) in Greece: Insights from mitochondrial and nuclear DNA // Molecular Phylogenetics and Evolution. V. 76. P. 144–154.
Scheldeman X., van Zonneveld M., 2010. Training manual on spatrial analysis of plant diversity and distribution. Rome: Bioversity International. 179 p.
Sillero N., Campos J., Bonardi A., Corti C., Creemers R., Crochet P.-A., Crnobrnja Isailović J., Denoël M., Ficetola G.F., Gonçalves J., Kuzmin S., Lymberakis P., de Pous P., Rodríguez A., Sindaco R., Speybroeck J., Toxopeus B., Vieites D.R., Vences M., 2014. Updated distribution and Biogeography of amphibians and reptiles of Europe // Amphibia-Reptilia. V. 35. P. 1–31.
Troidl A., Troidl S., 2000. Lacertidae. Eidechsen Porträt, Ausgabe Nr. 1. Text & Bild, Fürth. 10 p.
Troidl A., Troidl S., 2001. Lacertidae. Eidechsen Porträt, Ausgabe Nr. 2. Text & Bild, Fürth. 10 p.
Uetz P., Cherikh S., Shea G., Ineich I., Campbell P.D., Doronin I.V., Rosado J., Wynn A., Tighe K.A., Mcdiarmid R., Lee J.L., Köhler G., Ellis R., Doughty P., Raxworthy C.J., Scheinberg L., Resetar A., Sabaj M., Schneider G., Franzen M., Glaw F., Böhme W., Schweiger S., Gemel R., Couper P., Amey A., Dondorp E., Ofer G., Meiri S., Wallach V., 2019. A global catalog of primary reptile type specimens // Zootaxa. V. 4695. № 5. P. 438–450.
Vences M., Brown J.L., Lathrop A., Rosa G.M., Cameron A., Crottini A., Dolch R., Edmonds D., Freeman K.L.M., Glaw F., Grismer L.L., Litvinchuk S., Milne M.G., Moore M., Solofo J.F., Noël J., Nguyen T.Q., Ohler A., Randrianantoandro C., Raselimanana A.P., van Leeuwen P., Wogan G.O.U., Ziegler T., Andreone F., Murphy R.W., 2017. Tracing a toad invasion: lack of mitochondrial DNA variation, haplotype origins, and potential distribution of introduced Duttaphrynus melanostictus in Madagascar // Amphibia-Reptilia. Vol. 38. № 2. P. 197–207.
Warren D.L., Glor R.E., Turelli M., 2010. ENMTools: a toolbox for comparative studies of environmental niche models // Ecography. V. 33. P. 607–611.
Zink R.M., 2015. Genetics, morphology, and ecological niche modeling do not support the subspecies status of the endangered Southwestern Willow Flycatcher (Empidonax traillii extimus) // The Condor. V. 117. № 1. P. 76–86.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал