

Зоологический журнал, 2022, T. 101, № 5, стр. 525-536
Сезонная динамика повреждения насекомыми-филлофагами листьев дуба черешчатого (Quercus robur) на северо-востоке его ареала
И. В. Ермолаев a, *, А. А. Васильев b
a Ботанический сад УрО РАН
620130 Екатеринбург, Россия
b Удмуртский государственный университет
426034 Ижевск, Россия
* E-mail: ermolaev-i@yandex.ru
Поступила в редакцию 24.04.2021
После доработки 07.06.2021
Принята к публикации 07.08.2021
- EDN: CFVQSL
- DOI: 10.31857/S0044513422050051
Аннотация
Исследована сезонная динамика повреждения насекомыми-филлофагами дуба черешчатого (Quercus robur L.) на северо-востоке его ареала. Работу провели в течение 2013−2016 гг. близ биостанции Удмуртского государственного университета (56°82′ с.ш., 53°90′ в.д.) в пойменной дубраве р. Сива (Воткинский район, Удмуртская Республика). Комплекс насекомых-филлофагов дуба включает 71 вид насекомых из 23 семейств четырех отрядов: Lepidoptera (59.2%), Coleoptera (25.3%), Hymenoptera (12.7%) и Diptera (2.8%). Видовое разнообразие филлофагов трех отрядов (Coleoptera, Hymenoptera и Diptera) максимально в весенний и раннелетний периоды. В период с 10 июня по 30 августа количество видов Lepidoptera варьировало в диапазоне 21–25. Степень трофической специализации выявленного комплекса филлофагов дуба тесно связана с их экологической принадлежностью. Большинство видов минеров (71.4%) и все насекомые, образующие галлы, являются монофагами. Среди насекомых, формирующих объедание, скелетирование и трубки, доминируют полифаги: 92.4, 85.7 и 75%. Основное количество повреждений листьям дуба филлофаги наносят в весенний период – 77.1 ± 2.2%, в раннелетний, летний и осенний периоды – 12.2 ± 1.6, 3.3 ± 1.0 и 7.4 ± 2.1% (во всех случаях n = 8) соответственно.
Изучение системы “Quercus robur–насекомые-филлофаги” представляет значительный теоретический и практический интерес. Такие работы позволяют изучить процессы, протекающие при реализации вспышек массового размножения филлофагов (Feeny, 1970; Воронцов, 1978; Рубцов, Рубцова, 1984; Иерусалимов, 1986, 1994, 2004; Уткина, Рубцов, 1989, 1994; Уткина, 1991; Рубцов, Уткина, 2008; Visakorpi et al., 2020), оценить их экологические последствия (Воронцов и др., 1967; Лямцев, 1995; Рубцов, Уткина, 1995; Иерусалимов, 2004), выявить трансформацию динамики численности насекомых в связи с изменением климата (Рубцов, Уткина, 2010, 2016), исследовать географическую изменчивость взаимодействий в системе “растение–фитофаг–энтомофаг” (Valdés-Correcher et al., 2020).
Исследование особенностей повреждения листьев древесных растений комплексом насекомых филлогагов – простой и информативный метод оценки консортивных отношений экологических групп насекомых с растением. Согласно данным фундаментального исследования Герасимова (1952, стр. 156−158), филлофаги могут образовывать следующие типы повреждений: объедание, скелетирование, минирование, галлы и сигары. Последний тип повреждения часто называют трубками (Бей-Биенко, 1980). Перечисленные типы можно отнести к качественной характеристике повреждения. Количественной характеристикой повреждения является изъятие листовой поверхности, т.е. величины отчуждаемой насекомыми площади листьев (Богачева, 1990). Этот показатель адекватно характеризует нагрузку филлофага на растение (потери фотосинтезирующей поверхности) и тесно коррелирует с видовым разнообразием (числом видов, входящих в консорцию) и численностью этих видов (Богачева, 1990, стр. 12). Изъятие листовой поверхности можно оценить в случае объедания, скелетирования и минирования.
Цель представленной работы – исследовать сезонную динамику повреждения дуба Q. robur насекомыми-филлофагами на северо-востоке его ареала.
МАТЕРИАЛ И МЕТОДИКА
Особенности сезонной динамики поврежденности и изъятия листовой пластинки листьев дуба черешчатого (Quercus robur) комплексом насекомых-филлофагов исследовали в течение вегетационных сезонов 2013–2016 гг. близ биостанции Удмуртского университета (56°82′ с.ш., 53°90′ в.д.) в пойменной дубраве р. Сива (Воткинский р-н, Удмуртской Республики). Для этого были заложены пробные площади 1 (в 2013 г.) и 2 (в 2014 г.) около 0.5 га каждая. Выбор места пробной площади осуществляли исходя из разнообразия видового состава древостоя. Все пойменные дубравы долины р. Сива представлены липовым дубняком. Однако на пробной площади 1, помимо традиционных ассоциаций дуба черешчатого (Q. robur), липы мелколистной (Tilia cordata Mill.) и осины (Populus tremula L.), около 20% деревьев было представлено вязом гладким (Ulmus laevis Pall.). Карта расположения и описание пробных площадей были представлены нами ранее (Ермолаев и др., 2021). На каждой площади методом случайной выборки было выбрано 30 модельных деревьев дуба.
Исследование листьев дуба проводили 15 июня, 15 июля, 15 августа и 15 сентября на постоянной, выделенной маркерной лентой ветви первого порядка нижнего яруса восточной экспозиции каждого модельного дерева. Повреждения, наносимые в первый период, называли весенними, во второй, третий и четвертые периоды – раннелетними, летними и осенними, соответственно. Число листьев на модельной ветви составляло не менее 100 шт. Типы повреждения листьев даны в соответствии с классификацией Герасимова (1952): объедание, скелетирование, минирование, галлы. Помимо этого, выделяли трубки – тип повреждения, при котором филлофаг сворачивает повреждаемый лист (обычно в рулон).
Сбор филлофагов дуба проводили в течение вегетации. Имаго и личинок свободноживущих филлофагов, питающихся листьями дуба, собирали, обтряхивая ветви (не модельных деревьев) в энтомологический сачок. Личинок выкармливали листьями дуба в полевой лаборатории до стадии куколок и выводили имаго. Минеров собирали на стадии куколки. Всех насекомых на стадии имаго монтировали по общепринятой методике для последующего изучения. В течение сезона собирали листья с минами и галлами. Материал гербаризировали и снабжали этикеткой. Часть редких чешуекрылых-филлофагов, специализированных на Q. robur, была выявлена с помощью световой ловушки (на основе лампы ДРЛ-250) на биостанции “Сива”. Подтверждение правильности определения собранного материала проведено экспертами.
При анализе широты потенциальных трофических связей филлофагов подразделяли выделяли на монофагов, специализирующихся на представителях одного рода, олигофагов, – развивающихся на растениях одного семейства, и полифагов, – живущих на разных семействах растений. Для проведения анализа использовали справочники: Вредители леса (1955); Насекомые и клещи – вредители сельскохозяйственных культур. Т. II (1974), Т. III. Ч. 1 (1994) и Ч. 2 (1999), а также монографию Матова и Кононенко (2012).
Рассчитывали следующий показатель: поврежденность листьев = (число листьев с определенным типом повреждения) × (общее число листьев на ветви)–1 × 100, %.
Во всех случаях рассчитывали среднее арифметическое значение и его ошибку. Статистическую обработку материала проводили стандартными методами (Ивантер, Коросов, 2011).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Комплекс насекомых-филлофагов Q. robur долины р. Сива включает 71 вид. Это представители 23 семейств четырех отрядов: Lepidoptera (59.2%), Coleoptera (25.3%), Hymenoptera (12.7%) и Diptera (2.8%) (табл. 1). При этом 58 видов наносят повреждения на стадии личинки и 13 − при дополнительном питании на стадии имаго. Из них 42 вида (59.1%) объедают листья; 8, 7 и 1 вид (11.3, 9.9 и 1.4% соответственно) минируют, образуют галлы и скелетируют. 13 видов (18.3%) дают смешанные типы повреждений. Например, скелетирование у личинок младшего и объедание у личинок старшего возраста характерно для ряда видов листоедов (Luperus flavipes и Altica quercetorum), коконопрядов (Malacosoma neustria), хохлаток (Ptilodon capucina и Phalera bucephala), волнянок (Calliteara pudibunda, Euproctis similis) и совок (Meganola strigula, Moma alpium). У листоверток (Tortrix viridana, Acleris quercinana и Ancylis mitterbacheriana) и трубковертов (Byctiscus populi) личинки сначала скелетируют, затем объедают лист внутри трубок.
Таблица 1.
Тип питания и сезонная динамика встречаемости насекомых-филлофагов дуба черешчатого (Q. robur) долины р. Сива
| ТП | Вид | Месяц/декада | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | VI | VII | VIII | IX | |||||||||||
| 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | ||
| Scarabaeidae | |||||||||||||||
| о | Melolontha hippocastani Fabricius 1801 | + | + | + | + | ||||||||||
| Buprestidae | |||||||||||||||
| о | Agrilus biguttatus (Fabricius 1777) | + | + | + | + | ||||||||||
| о | A. angustulus angustulus (Illiger 1803) | + | + | + | + | + | |||||||||
| Elateridae | |||||||||||||||
| о | Selatosomus cruciatus (Linnaeus 1758) | + | + | + | + | + | + | ||||||||
| Cerambycidae | |||||||||||||||
| о | Acanthoderes clavipes (Schrank 1781) | + | + | + | + | + | + | + | |||||||
| о | Saperda scalaris hieroglyphica (Pallas 1773) | + | + | + | + | + | + | + | |||||||
| Chrysomelidae | |||||||||||||||
| о | Cryptocephalus flavipes Fabricius 1781 | + | + | + | + | + | |||||||||
| с, о | Luperus flavipes (Linnaeus 1758) | + | + | + | + | + | + | + | |||||||
| с, о | Altica quercetorum Foudras 1860 | + | + | + | + | + | + | + | + | + | + | + | + | ||
| Attelabidae | |||||||||||||||
| т, с, о | Byctiscus populi (Linnaeus 1758) | + | + | + | + | + | + | + | + | + | + | + | |||
| Curculionidae | |||||||||||||||
| о | Curculio glandium Marsham 1802 | + | + | + | + | + | + | + | + | ||||||
| о | C. venosus (Gravenhorst 1807) | + | + | + | + | + | + | + | + | ||||||
| о | C.villosus Fabricius 1781 | ? | |||||||||||||
| о | Archarius pyrrhoceras (Marsham 1802) | ? | |||||||||||||
| м | Orchestessignifer (Creutzer 1799) | + | + | + | |||||||||||
| о | Phyllobius pyri (Linnaeus 1758) | + | + | + | + | + | |||||||||
| о | Polydrusus flavipes (Degeer 1775) | + | + | + | + | + | + | ||||||||
| о | P. undatus (Fabricius 1781) | + | + | + | + | + | + | ||||||||
| Tenthredinidae | |||||||||||||||
| о | Allantus togatus (Panzer 1801) | + | + | + | + | ||||||||||
| с | Caliroa cinxia (Klug 1816) | + | + | + | + | + | |||||||||
| м | Profenusa pygmaea (Klug 1816) | + | + | + | + | + | |||||||||
| о | Mesoneura opaca (Fabricius 1775) | + | + | + | + | + | |||||||||
| Cynipidae | |||||||||||||||
| г | Neuroterus quercusbaccarum (Linnaeus 1758) | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| г | Cynips quercusfolii Linnaeus 1758 | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| г | Andricus curvator Hartig 1840 | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| г | A. foecundatrix (Hartig 1840) | + | + | + | + | + | + | + | + | + | |||||
| г | A. quadrilineatus Hartig 1840 | + | + | + | + | + | + | ||||||||
| Nepticulidae | |||||||||||||||
| м | Stigmella sp. 1 | + | + | + | + | + | + | + | |||||||
| м | Stigmella sp. 2 | + | + | + | + | + | + | + | |||||||
| Tischeriidae | |||||||||||||||
| м | Tischeria ekebladella (Bjerkander 1795) | + | + | + | + | ||||||||||
| Gracillariidae | |||||||||||||||
| м | Acrocercops brongniardella (Fabricius 1798) | + | + | + | + | ||||||||||
| м | Phyllonorycter roboris (Zeller 1839) | + | + | + | + | ||||||||||
| Coleophoridae | |||||||||||||||
| м | Coleophora sp. | + | + | + | |||||||||||
| Limacodidae | |||||||||||||||
| о | Apoda limacodes (Hufnagel 1766) | + | + | + | + | + | + | + | |||||||
| Tortricidae | |||||||||||||||
| т, с, о | Tortrix viridana Linnaeus 1758 | + | + | + | + | ||||||||||
| т, с, о | Acleris quercinana (Zeller 1849) | + | + | + | + | + | |||||||||
| т, с, о | Ancylis mitterbacheriana ([Denis & Schiffermüller] 1775) | + | + | + | + | + | + | + | + | ||||||
| Geometridae | |||||||||||||||
| о | Ennomos autumnaria (Werneburg 1859) | + | + | + | + | + | + | ||||||||
| о | E. erosaria ([Denis & Schiffermüller] 1775) | + | + | + | + | + | + | ||||||||
| о | Selenia lunularia (Hübner [1788]) | + | + | + | + | + | + | + | + | ||||||
| о | Plagodis dolabraria (Linnaeus 1767) | + | + | + | + | + | + | ||||||||
| о | P. pulveraria (Linnaeus 1758) | + | + | + | + | + | + | ||||||||
| о | Hypomecispunctinalis (Scopoli 1763) | + | + | + | + | + | + | + | + | + | |||||
| о | H. roboraria ([Denis & Schiffermüller] 1775) | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| о | Parectropis similaria (Hufnagel 1767) | + | + | + | + | + | + | + | + | + | + | ||||
| о | Cyclophora porata (Linnaeus 1767) | + | + | + | + | + | + | + | + | + | |||||
| о | C. punctaria (Linnaeus 1758) | + | + | + | + | + | + | + | + | + | + | ||||
| о | C. quercimontaria (Bastelberger 1897) | + | + | + | + | + | + | + | + | ||||||
| Lasiocampidae | |||||||||||||||
| с, о | Malacosoma neustria (Linnaeus 1758) | + | + | + | + | + | + | ||||||||
| о | Gastropacha quercifolia (Linnaeus 1758) | + | + | + | + | + | + | + | + | + | |||||
| Shingidae | |||||||||||||||
| о | Mimas tiliae (Linnaeus 1758) | + | + | + | + | + | + | ||||||||
| Notodontidae | |||||||||||||||
| о | Drymonia dodonaea ([Denis & Schiffermüller] 1775) | + | + | + | + | + | |||||||||
| c, о | Ptilodon capucina (Linnaeus 1758) | + | + | + | + | + | + | + | + | + | |||||
| с, о | Phalera bucephala (Linnaeus 1758) | + | + | + | + | + | |||||||||
| Lymantriidae | |||||||||||||||
| с, о | Calliteara pudibunda (Linnaeus 1758) | + | + | + | + | + | + | + | + | ||||||
| с, о | Euproctis similis (Fuessly 1775) | + | + | + | + | + | + | + | + | ||||||
| о | Lymantria dispar (Linnaeus 1758) | + | + | + | + | + | |||||||||
| Noctuidae | |||||||||||||||
| с, о | Meganola strigula ([Denis & Schiffermüller] 1775) | + | + | + | + | + | |||||||||
| о | Pseudoips prasinana (Linnaeus 1758) | + | + | + | + | + | + | ||||||||
| о | Trisateles emortualis ([Denis & Schiffermüller] 1775) | + | + | + | + | + | + | + | |||||||
| о | Catephia alchymista ([Denis & Schiffermüller] 1775) | + | + | + | + | + | |||||||||
| о | Catocala promissa ([Denis & Schiffermüller] 1775) | + | + | + | + | ||||||||||
| о | C. sponsa (Linnaeus 1767) | + | + | + | + | ||||||||||
| о | Colocasia coryli (Linnaeus 1758) | + | + | + | + | + | + | + | |||||||
| с, о | Moma alpium (Osbeck 1778) | + | + | + | + | + | + | + | |||||||
| о | Acronicta aceris (Linnaeus 1758) | + | + | + | + | + | + | + | + | ||||||
| о | A. auricoma ([Denis & Schiffermüller] 1775) | + | + | + | + | + | + | + | + | + | |||||
| о | Conistra vaccinii (Linnaeus 1761) | + | + | + | + | + | |||||||||
| Lycaenidae | |||||||||||||||
| о | Nordmannia w-album (Knoch 1782) | + | + | + | + | + | |||||||||
| Cecidomyiidae | |||||||||||||||
| г | Macrodiplosis dryobiae (Low 1877) | + | + | + | + | + | |||||||||
| г | M. volvens Kieffer 1895 | + | + | + | + | + | |||||||||
В таксономическом отношении выявленный состав филлофагов характеризуется относительной бедностью. Это связано с низкой теплообеспеченностью места проведения исследования (Ермолаев и др., 2021). Для сравнения в европейской части бывшего СССР было известно 530 видов членистоногих, питающихся листьями Q. robur (Довнар-Запольский, 1954). Из них в дубравах Северного Кавказа, Молдавии, Николаевской обл. Украины, Воронежской и Волгоградской областей РФ – 250 (Дейко, Арефьев, 2004), 243 (Плугарь, 1963), 108 (Вайнштейн, 1950), 184 (Моравская, 1954, 1957, 1975) и 140 (Моравская, 1952) видов-филлофагов, соответственно.
Степень трофической специализации выявленного комплекса филлофагов дуба тесно связана с их экологической принадлежностью. К высокоспециализированным можно отнести представителей эндофагов. Среди минеров большинство видов (71.4%), среди галлообразователей – все являются монофагами. Два вида минеров (Profenusa pygmaea и Tischeria ekebladella) способны питаться и на других представителях Fagaceae, например, на каштане, и поэтому относятся к олигофагам. Среди насекомых, формирующих объедание, скелетирование и трубки, доминируют полифаги: 92.4, 85.7 и 75%. Так, среди насекомых, для которых характерно объедание, выявлено только два вида моно- и два вида олигофагов. Это Agrilus biguttatus, Archarius pyrrhoceras и Agrilus angustulus angustulus, Catocala promissa соответственно. При этом ксилофаги рода Agrilus используют листья дуба в качестве источника дополнительного питания. Среди насекомых, для которых характерно скелетирование, выявлено два монофага: Caliroa cinxia и Acleris quercinana. Последний вид также образует трубки.
Наше исследование позволило уточнить трофическую специализацию пяденицы Cyclophora quercimontaria. Несмотря на сложившееся мнение об узкой трофической специализации этого вида на представителях рода Quercus (Hausmann, 2004), авторы собирали гусениц этого вида и выводили имаго при выполнении аналогичной работы на липе мелколистной (Tilia cordata Mill.) в г. Ижевске (Ермолаев, Сидорова, 2011). В связи с этим вид был отнесен нами к полифагам.
Сезонная динамика видового разнообразия филлофагов Q. robur из четырех отрядов насекомых представлена на рис. 1. Максимальное число видов отмечено в первой половине периода вегетации. При этом в начале сезона общее число видов растет до 20 июня (49 видов), затем с той же скоростью снижается до конца июля, стабилизируется в августе (27−29 видов) и снова сокращается в сентябре. Видовое разнообразие филлофагов трех отрядов (Coleoptera, Hymenoptera и Diptera) было максимальным в весенний и раннелетний периоды. В отличие от них после весеннего роста общее число видов Lepidoptera варьировало в диапазоне 21−25 в течение большей части сезона (с 10 июня по 30 августа). Очевидно, что в разных частях ареала Q. robur сезонная динамика общего числа видов филлофагов дуба из одного отряда может быть разной. Так, в Великобритании максимальное число видов Lepidoptera на дубе наблюдали в мае–первой половине июня (около 110), затем число видов падало до середины августа (около 60) и снова росло в сентябре (чуть более 80) (Feeny, 1970).

Динамика таксономической структуры филлофагов в течение вегетации, вероятно, тесно связана с изменением качества корма. Общеизвестно, что пищевое качество листа дуба Q. rodur значительно меняется в течение вегетации. Согласно классической работе Фини (Feeny, 1970), содержание воды в листьях дерева максимально в мае и в дальнейшем снижается в течение сезона (наиболее интенсивно с конца мая до середины июня). Общая концентрация соединений азота снижается в течение сезона (Salminen et al., 2004). Концентрация белка снижается до второй декады мая, и в дальнейшем стабилизируется. Количество таннинов нарастает в течение вегетации (наиболее интенсивно в августе–сентябре) (Feeny, 1970). При этом молодые листья дуба более богаты гидролизуемыми таннинами и флавоноидными гликозидами, старые – проантоцианидинами (Salminen et al., 2004). Содержание хлорофилла a и b в листьях Q. rodur растет с июня по август, затем падает (Gond et al., 1999). Тот же тренд имеет общая концентрация соединений углерода (Salminen et al., 2004). Анализ географической изменчивости в системе “Q. rodur–фитофаг– хищник”, проведенный в 2018−2019 гг. группой исследователей в разных странах Европы, показал, что представленность ряда высокоспециализированных экологических групп филлофагов может зависеть от концентрации в листьях определенных соединений (Valdés-Correcher et al., 2020). Так, в консорции Q. robur доля минеров растет с увеличением в тканях листа концентрации гидролизуемых таннинов, а доля галлообразователей – растет с повышением концентрации растворимого сахара и уменьшением концентрации лигнина (Valdés-Correcher et al., 2020).
Общая поврежденность листьев дуба – суммарный показатель (рис. 2), включающий в себя объедание (рис. 3), скелетирование (рис. 4), минирование (рис. 5), образованием галлов и трубок (рис. 6 и 7), а также смешанные на одном листе повреждения (рис. 8). В районе исследований доминировали первых два типа повреждения. Общая поврежденность листьев дуба на двух пробных площадях росла в течение периода вегетации (рис. 2). Это происходило преимущественно за счет увеличения доли скелетированных (рис. 4) и минированных (рис. 5) листьев. К концу сезона этот показатель был в диапазоне от 69.6 ± 1.2 (2016 г., пробная площадь № 2) до 94.6 ± 1.0% (2014 г., пробная площадь № 2). Если осенний показатель общей поврежденности на конкретной пробной площади принять за 100%, то основное количество повреждений листьям дуба филлофаги наносят в весенний период – 77.1 ± 2.2%, в раннелетний, летний и осенний периоды – 12.0 ± 1.5, 3.8 ± 1.1 и 7.1 ± 2.2% (во всех случаях n = 8), соответственно. Таким образом, основа сезонной картины повреждения зависит от активности весеннего комплекса филлофагов. Схожий вывод был получен при исследовании динамики поврежденности листьев березы, осины и ивы в Челябинской обл. (Рафес, Соколов, 1976), березы и рябины в лесах Нижнего Приангарья (Петренко, Дрянных, 1978), липы мелколистной в г. Ижевске (Ермолаев, Сидорова, 2011). По всей вероятности, синхронизация стадий питания и роста основного количества филлофагов с весенним изменением биохимического состава листьев кормового растения является общей адаптацией представителей с гетеродинамным развитием.
Рис. 2.
Динамика общей поврежденности (%) листьев дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.). На рис. 2−8 по вертикальной оси указана доля (%) поврежденных листьев.

Рис. 3.
Динамика объедания (%) листьев дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).
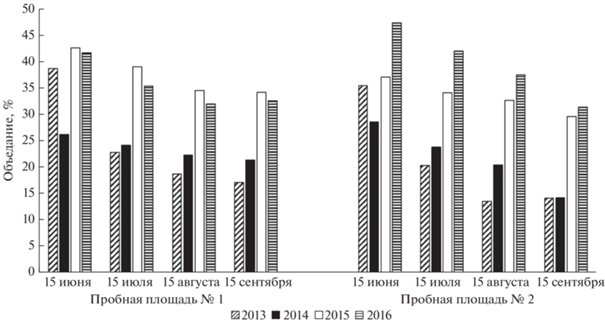
Рис. 4.
Динамика скелетирования (%) листьев дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).

Рис. 5.
Динамика минирования (%) листьев дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).

Рис. 6.
Динамика образования галлов (%) на листьях дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).

Рис. 7.
Динамика образования трубок (%) на листьях дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).

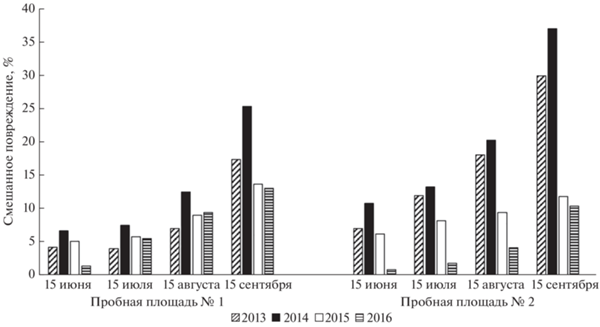
Рис. 8.
Динамика смешанного повреждения (%) на листьях дуба черешчатого на двух пробных площадях в долине р. Сива (2013−2016 гг.).
Объедание листьев – доминирующий тип повреждений, вызываемых филлофагами дуба. В долине р. Сива объедание отмечено для 37 видов представителей Lepidoptera, 17 видов – Coleoptera и 2 видов Hymenoptera (табл. 1). Листья с таким типом повреждения наблюдали преимущественно в весенний период (рис. 3). В дальнейшем доля такого повреждения снижалась в течение периода вегетации. Это происходило на фоне увеличения размера листьев на модельной ветви. У Q. robur наряду с весенним хорошо представлено коррелятивное побегообразование (“ивановы побеги”) (Грудзинская, 1960). Если весеннее побегообразование у дуба происходит в период от набухания почек до первой половины июня, то коррелятивное имеет место в июне (Добрынин, Комиссарова, 2012). Дополнительное летнее побегообразование обусловлено существованием корреляции между ростом и образованием листьев при определенном соотношении корневой и листовой поверхности (Грудзинская, 1960).
Скелетирование листьев дуба как тип повреждения отмечалось так же часто. В течение периода вегетации число листьев, несущих этот тип повреждения, росло не равномерно (рис. 4). Весной листья скелетировали гусеницы Malacosoma neustria, Euproctis similis, Meganola strigula, а также имаго жуков Luperus flavipes. Основное повреждение листья получали в раннелетний период. При этом доминировали личинки листоеда Altica quercetorum. Кроме того, в этот период были активны личинки пилильщика Caliroa cinxia, а также гусеницы Ptilodon capucina, Calliteara pudibunda, Moma alpium.
Минирование листьев было обычно. Количество листьев, несущих мины, обычно росло в течение сезона (рис. 5). В конце вегетации 2013−2014 гг. доля таких листьев не превышала 5%. В 2016 г. показатель достигал значений 8.3% (пробная площадь № 1, июль), в 2015 г. − 13.5% (пробная площадь № 1, август). Повреждения наносили представители восьми видов шести семейств: весной − Orchestes signifer (Curculionidae), Coleophora sp. (Coleophoridae), Acrocercops brongniardella (Gracillariidae), в раннелетний период − Profenusa pygmaea (Tenthredinidae), Stigmella sp. 1, Stigmella sp. 2 (Nepticulidae), Tischeria ekebladella (Tischeriidae), Phyllonorycter roboris (Gracillariidae), осенью − Stigmella sp. 1, Stigmella sp. 2. Наиболее обычными были мины Ph. roboris и Stigmella sp.1.
Образование галлов на листьях дуба встречали относительно редко. За четыре года наблюдений число листьев, несущих галлы, составляло от менее 0.5 (2014 г.) до 5% (пробная площадь № 1, июль, август 2016 г.) (рис. 6). Максимальное количество листьев, несущих галлы, наблюдали в раннелетний период. В дальнейшем увеличение числа повреждений этого типа не наблюдали. Повреждение было связано с активностью семи видов насекомых: Neuroterus quercusbaccarum, Cynips quercusfolii, Andricus curvator, A. foecundatrix, A. quadrilineatus (Cynipidae), Macrodiplosis dryobiae и M. volvens (Cecidomyiidae) (табл. 1). К началу проведения июльских учетов в 2013, 2015 и 2016 гг. наблюдали 2–3-кратный рост повреждения по сравнению с предыдущими показателями (рис. 6). Это происходило в результате появления галлов, образуемых личинками агамного поколения C. quercusfolii в середине и во второй половине июня. Вид доминировал на обеих пробных площадях.
Образование трубок отмечали также редко. Если в 2013−2014 гг. на обеих пробных площадях это повреждение не превышало 0.5%, то в 2015− 2016 гг. – 2.5% листьев (рис. 7). Максимальное количество листьев, несущих трубки, наблюдали в раннелетний период. В дальнейшем роста этого типа повреждений не наблюдали. Повреждение было вызвано четырьмя видами: тополевым трубковертом (Byctiscus populi) и тремя листовертками: Tortrix viridana, Acleris quercinana и Ancylis mitterbacheriana. Гусеницы первых двух видов листоверток проходят развитие в мае – июне, личинки B. populi в июне – июле, гусеницы A. mitterbacheriana − с конца июня по август (табл. 1). Последний вид обычен.
Количество листьев, несущих смешанные повреждения (сочетание на одном листе двух и более типов повреждения), увеличивается в течение периода вегетации (рис. 8). Если к концу вегетации 2013−2014 гг. таких листьев было более 30%, то в 2015−2016 гг. – чуть более 10%.
Разница между показателями поврежденности на двух пробных площадях пойменных дубрав была незначительной (рис. 2−8). Это связано со сходством структуры древостоев (Ермолаев и др., 2021) на обеих площадях, а также сходством встречающихся в этих площадях комплексов филлофагов.
Список литературы
Бей-Биенко Г.Я., 1980. Общая энтомология. М.: Высшая школа. 416 с.
Богачева И.А., 1990. Взаимоотношения насекомых-фитофагов и растений в экосистемах Субарктики. Свердловск: УрО РАН СССР. 137 с.
Вайнштейн Б.А., 1950. Листогрызущие вредители дуба и их сезонная динамика // Зоологический журнал. Т. 29. Вып. 2. С. 107−112.
Воронцов А.И., Иерусалимов Е.Н., Мозолевская Е.Г., 1967. Роль листогрызущих насекомых в лесном биогеоценозе // Журнал общей биологии. Т. 28. № 2. С. 172−187.
Воронцов А.И., 1978. Патология леса. М.: Лесная промышленность. 272 с.
Вредители леса, 1955. М.−Л.: Издательство АН СССР. Т. I, Т. II. 1097 с.
Герасимов А.М., 1952. Фауна СССР. Насекомые чешуекрылые. Гусеницы. М. − Л.: Издательство АН СССР. Т. 1. Вып. 2. Ч. 1. 338 с.
Грудзинская И.А., 1960. Летнее побегообразование у древесных растений и его классификация // Ботанический журнал. Т. 45. № 7. С. 969−978.
Дейко В.Г., Арефьев Ю.Ф., 2004. О надзоре и учете главнейших филлофагов дуба в Краснодарском крае // Лесное образование и лесная наука в XXI веке. Материалы региональной науч.-практ. юбилейной конф., посвященной 85-летию высшего лесного образования в г. Воронеже и ЦЧР (г. Воронеж, 12–13 февраля 2004 г.). Воронеж: Воронежская государственная лесотехнологическая академия. С. 87−89.
Добрынин А.П., Комиссарова М.Г., 2012. Самые северные дубравы России. Вологда: Полиграф-Книга. 188 с.
Довнар-Запольский Д.П., 1954. Очерк энтомофауны черешчатого дуба (Quercus robur) в европейской части СССР // Зоологический журнал. Т. 33. Вып. 4. С. 794−806.
Ермолаев И.В., Сидорова О.В., 2011. Сезонная динамика повреждения липы мелколистной комплексом членистоногих-филлофагов // Зоологический журнал. Т. 90. № 5. С. 552−558.
Ермолаев И.В., Пономарев В.И., Васильев А.А., Кумаева М.С., 2021. Насекомые-фитофаги дуба черешчатого на северо-востоке его ареала // Зоологический журнал. Т. 100. № 6. С. 640−651.
Ивантер Э.В., Коросов А.В., 2011. Введение в количественную биологию. Петрозаводск: Изд-во ПетрГУ. 302 с.
Иерусалимов Е.Н., 1986. Последствия повреждения дуба комплексом листогрызущих насекомых // Растительноядные животные в биогеоценозах суши. Материалы Всесоюз. совещания. Валдай, 3–6 июня 1984 г. М.: Наука. С. 76−84.
Иерусалимов Е.Н., 1994. Особенности дефолиации дуба в очагах массового размножения непарного шелкопряда и зимней пяденицы // Лесоведение. № 3. С. 10−21.
Иерусалимов Е.Н., 2004. Зоогенная дефолиация и лесное сообщество. М.: Товарищество научных изданий КМК. 263 с.
Лямцев Н.И., 1995. Влияние листогрызущих насекомых на прирост дуба в порослевых дубравах // Лесоведение. № 6. С. 23−33.
Матов А.Ю., Кононенко В.С., 2012. Трофические связи гусениц совкообразных чешуекрылых фауны России (Lepidoptera, Noctuoidea: Nolidae, Erebidae, Euteliidae, Noctuidae). Владивосток: Дальнаука. 346 с.
Моравская А.С., 1952. Насекомые-вредители дуба в условиях степного лесоразведения засушливого Юго-Востока. Автореф. дис. … канд. биол. наук. Москва: МГУ. 15 с.
Моравская А.С., 1954. Вредители листвы основных древесных пород Теллермановского леса (предварительное сообщение) // Сообщения института леса АН СССР. Вып. 3. С. 30−38.
Моравская А.С., 1957. Повреждаемость насекомыми рано- и позднораспускающихся форм дуба и ильмовых пород // Сообщения института леса АН СССР. Вып. 8. С. 44−63.
Моравская А.С., 1975. Насекомые-фитофаги дубравных лесов // Дубравы лесостепи в биогеоценологическом освещении. М.: Наука. С. 199−210.
Насекомые и клещи – вредители сельскохозяйственных культур, 1974. Жесткокрылые. Т. II. Л.: Наука. 335 с.
Насекомые и клещи – вредители сельскохозяйственных культур, 1994. Чешуекрылые. Т. III. Часть 1. СПб.: Наука. 316 с.
Насекомые и клещи – вредители сельскохозяйственных культур, 1999. Чешуекрылые. Т. III. Часть 2. СПб.: Наука. 410 с.
Петренко Е.С., Дрянных Н.М., 1978. Биоценотические закономерности использования кормовой базы насекомыми-филлофагами в лесах нижнего Приангарья // Экология питания лесных животных. Новосибирск: Наука. С. 76−88.
Плугарь С.Г., 1963. Некоторые результаты изучения энтомофауны дуба в лесах Молдавии // Вредная энтомофауна Молдавии и меры борьбы с ней. Кишинев: Издательство “Картя Молдовеняскэ”. С. 7−38.
Рафес П.М., Соколов В.К., 1976. О взаимодействии фоновых вредителей листвы с кормовым деревом // Доклады Академии наук СССР. Т. 228. № 1. С. 246−247.
Рубцов В.В., Рубцова Н.Н., 1984. Анализ взаимодействия листогрызущих насекомых с дубом. М.: Наука. 183 с.
Рубцов В.В., Уткина И.А., 1995. Влияние насекомых-филлофагов на прирост древесины дуба черешчатого // Лесоведение. № 2. С. 22−30.
Рубцов В.В., Уткина И.А., 2008. Адаптационные реакции дуба на дефолиацию. М.: Институт лесоведения РАН. 302 с.
Рубцов В.В., Уткина И.А., 2010. Филлофаги лесных экосистем в условиях изменяющегося климата // Вестник Марийского государственного технического университета. Серия Лес. Экология. Природопользование. № 3. С. 3−15.
Рубцов В.В., Уткина И.А., 2016. Дуб черешчатый и насекомые-филлофаги как объект изучения биоценотических взаимоотношений в лесной экосистеме Теллермановской дубравы // Лесотехнический журнал. № 4. С. 79−89.
Уткина И.А., Рубцов В.В., 1989. Особенности восстановления листвы дуба черешчатого после дефолиации крон листогрызущими насекомыми // Состояние дубрав лесостепи. М.: Наука. С. 116−137.
Уткина И.А., Рубцов В.В., 1994. Рефолиация дуба черешчатого после повреждения насекомыми // Лесоведение. № 3. С. 23−31.
Уткина И.А., 1991. Восстановление листвы дуба черешчатого после дефолиации крон листогрызущими насекомыми. Автореф. дис. … канд. биол. наук. Москва: лаборатория лесоведения АН СССР. 24 с.
Feeny P., 1970. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter moth caterpillars // Ecology. V. 51 (4). P. 565−581.
Gond V., de Pury D.G.G., Veroustraete F., Geulemans R., 1999. Seasonal variation in leaf area index, leaf chlorophyll, and water content; scaling-up to estimate fAPAR and carbon balance in a multilayer, multispecies temperate forest // Tree Physiology. V. 19. P. 673−679.
Hausmann A., 2004. The Geometrid Moths of Europe. V. 2. Sterrhinae. Apollo Books, Stenstrup. 600 p.
Salminen J.-P., Roslin T., Karonen M., Sinkkonen J., Pihlaja K., Pulkkinen P., 2004. Seasonal variation in the content of hydrolysable tannins, flavonoid glycosides, and proanthocyanidins in oak leaves // Journal of Chemical Ecology. V. 30 (9). P. 1693−1711.
Valdés-Correcher E., Moreira X., Augusto L., Barbaro L., Bouget C., Bouriaud O., Branco M., Centenaro G., Csóka G., Damestoy T., Dobrosavljevic J., Duduman L., Dulaurent A.-M., Eötvös C., Faticov M., Ferrante M., Fūrjes-Miko A., Galman A., Gossner M.M., Hampe A., Harvey D., Howe A.G., Kadiri Y., Kaennel-Dobbertin M., Koricheva J., Kozel A., Kozlov M.V., Lövei G.L., Lupastean D., Milanovic S., Mrazova A., Opgennoorth L., Pitkänen J.-M., Popova A., Popovic M., Prinzing A., Queloz V., Roslin T., Sallé A., Sam K., Scherer-Lorenzen M., Schuldt A., Selikhovkin A., Suominen L., Tack A.J.M., Tahadlova M., Thomas R., Castagneyrol B., 2020. Search for top-down and bottom-up drivers of latitudinal trends in insect herbivory in oak tree in Europe // Global Ecology and Biogeography. 00. P. 1–15. https://doi.org/10.1111/geb. 13244
Visakorpi K., Riutta T., Malhi Y., Salminen J.-P., Salinas N., Gripenberg S., 2020. Changes in oak (Quercus robur) photosynthesis after winter moth (Operophtera brumata) herbivory are not explained by changes in chemical or structural leaf traits // PLoS ONE. V. 15 (1). P. 1−20. [Электронный ресурс]. Дата обновления: 11.03.2020. Режим доступа: https://doi.org/10.1371/journal.pone.0228157
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал