

Зоологический журнал, 2022, T. 101, № 5, стр. 483-491
Сравнительная морфология биссусной бороздки ноги у трех видов митилид (Bivalvia, Mytilidae) из Японского моря
Е. Е. Вехова *
Национальный научный центр морской биологии имени А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: evechova@gmail.com
Поступила в редакцию 18.03.2021
После доработки 03.11.2021
Принята к публикации 03.11.2021
- EDN: YFGFRY
- DOI: 10.31857/S0044513422050063
Аннотация
У массовых видов митилид Mytilus coruscus, Crenomytilus grayanus, Modiolus modiolus из Японского моря изучены особенности морфологического строения биссусной бороздки ноги. Сравнительный анализ показал, что эти виды различаются длиной, шириной, глубиной биссусной бороздки и особенностями ее строения, а также размерами и формой дистальной ямки, где происходит образование биссусной нити и прикрепительного диска соответственно. Особенности морфологического строения биссуса у Myt. coruscus, C. grayanus, Mod. modiolus объясняются различиями в строении секреторного органа.
Двустворчатые моллюски семейства Mytilidae часто являются доминирующим компонентом бентосных сообществ прибрежных вод Японского моря. Кроме этого, митилиды играют важную роль в структуре и функционировании морских экосистем (Скарлато и др., 1967; Скарлато, 1981). По мнению этого автора (Скарлато, 1981), митилиды обитают в верхней сублиторали Японского моря на глубине 1–60 м, образуя основные промысловые скопления не глубже 20 м.
Для выживания в прибойной зоне верхней сублиторали такие виды митилид как мидия блестящая (Mytilus coruscus Gould 1861), мидия Грея (Crenomytilus grayanus (Dunker 1853)) и модиолус (Modiolus modiolus (Linnaeus 1758)) на протяжении всей жизни прикрепляются к субстрату биссусом, возникшим у них в результате неотении (Yonge, 1962; Lutaenko, Noseworthy, 2012). Для мест обитания этих видов митилид характерна высокая волновая активность, соответствующая 3-й степени прибойности побережья (Лукин, Фадеев, 1982). Их биссусные нити прикрепляются к субстрату по направлению к макушке, заднему концу раковины и в боковых направлениях. Это позволяет смягчить фронтальное, отраженное и боковые воздействия волн на раковину этих митилид (Denny, 1987; Denny et al., 1998). На протяжении всей жизни эти двустворчатые моллюски могут отбросить биссус и, переместившись с помощью ноги в пригодное местообитание, вновь образовать новый биссус с помощью желез ноги (Waite, 1983, 1997; Vekhova, 2019, 2021; Вехова, 2007).
В настоящее время морфология биссусной бороздки ноги митилид изучена методами сканирующей электронной микроскопии лишь у немногих представителей этого семейства. Среди них такие представители рода Mytilus, как Myt. edulis и Myt. trossulus (Pujol, 1967; Allen et al., 1976; Price, 1983; Vekhova, 2021; Вехова, 2021; Бергер и др., 1985). Строение биссусной бороздки ноги у Myt. coruscus, C. grayanus, Mod. modiolus до сих пор не изучено. По данным автора (Vekhova, 2006), при повторном прикреплении к антропогенным субстратам (бетону, дереву и железу) каждый из этих видов имел свои особенности поведения. C. grayanus предпочитал прикрепляться к твердому субстрату (бетону), а Mod. modiolus прикреплялся ко всем субстратам, кроме дерева. Сведения о колонизации антропогенных субстратов Myt. coruscus в научной литературе в настоящее время отсутствуют. Для Myt. coruscus, C. grayanus и Mod. modiolus установлено, что они различаются размерами и морфологией биссусных нитей (Vekhova, 2019; Вехова, 2007), т.е. имеют видоспецифические адаптации к прикрепленному образу жизни. По-видимому, на строение биссуса влияют особенности строения секреторного органа, что определяет прочность прикрепления к субстрату и выживаемость видов в верхней сублиторали.
Цель работы – провести сравнительное исследование особенностей морфологического строения биссусной бороздки ноги у Mytilus coruscus, Crenomytilus grayanus, Modiolus modiolus из залива Восток Японского моря.
МАТЕРИАЛ И МЕТОДИКА
Для изучения особенностей морфологии биссусной бороздки ноги использовали моллюсков, собранных с крупных валунов и скал в 2019–2021 гг. с помощью водолазной службы Национального научного центра морской биологии имени А.В. Жирмунского ДВО РАН на глубине 1.5‒3 м в заливе Восток Японского моря (42°53′03.35″ с.ш., 132°43′41.72″ в.д.). В лабораторных условиях у 10 экз. каждого вида с помощью штангенциркуля с точностью до ± 0.1 мм измеряли длину раковины (мм). По наружным кольцам роста на поверхности раковины определяли возраст двустворчатых моллюсков (Золотарев, 1989). Далее перед анатомированием каждую исследуемую особь двустворчатого моллюска расслабляли с помощью инъекции 1 М раствора хлорида калия. Используя метод световой микроскопии (СМ), у 30 особей каждого вида размером 40.0 ± 4.0 мм удаляли биссус с биссусными нитями, при этом у каждой особи отрезали ногу и изучали ее параметры под бинокуляром при окулярах ×8 и тубусе ×0.6, оценивали длину ноги (мм) и ее ширину (мм).
У 5 экз. каждого вида митилид изучали особенности морфологического строения биссусной бороздки ноги методом сканирующей электронной микроскопии (СЭМ). Для этого использовали одновозрастных моллюсков размером 40.0 ± 4.0 мм. Для СЭМ образцы ног митилид фиксировали в 2.5% растворе глютаральдегида, приготовленном на 0.1 М какодилатном буфере (pH 7.4), в течение 2–4 ч при температуре 4°С. После этого образцы промывали в 0.1 М какодилатном буфере в течение 15–20 мин. Далее образцы обезвоживали в спиртах возрастающей концентрации, постепенно доводя образцы до чистого ацетона (Миронов и др., 1994). После этого образцы окончательно высушивали в диоксиде углерода по методике сушки в критической точке, используя прибор critical point dryer 030 фирмы “BAL-TEC”, помещали на поверхность алюминиевых столиков и напыляли хромом, используя вакуумный прибор для покрытия тонких мембран Q 150T ES. Далее особенности морфологии образцов биссусной бороздки ноги митилид исследовали на сканирующем электронном микроскопе Carl Zeiss, Sigma 300 VP.
Все полученные изображения редактировали с использованием графической программы Adobe Photoshop CS6. С помощью программы Smartiff измеряли длину и ширину биссусной бороздки ноги, глубину биссусной бороздки, а также оценивали размер дистальной ямки на дистальном конце биссусной бороздки ноги, измеряли длину ресничек и сосочков на дне и стенке биссусной бороздки и дистальной ямки. Измеряли ширину складок на стенке и дне биссусной бороздки. Статистическую обработку данных выполняли на персональном компьютере по стандартным алгоритмам, реализованным в пакете прикладных программ Microsoft Excel.
РЕЗУЛЬТАТЫ
Морфология ноги
Секреторную функцию формирования биссусных нитей и биссуса в целом выполняет нога, которая по внешнему виду напоминает язык. Как правило, в ноге очень много мышц, поэтому нога очень подвижна и может увеличиваться в длину, превышающую ее оригинальный размер. Кроме этого, мышцы ноги поддерживают железы, которые локализованы в ноге. С помощью ноги эти виды могут передвигаться по поверхности субстрата, покидая неблагоприятные местообитания.
Так, у вскрытой мидии Myt. coruscus размером 40 мм длина ноги 7.65 ± 0.40 мм, ширина ноги 5.0 ± 0.3 мм. Нога ‒ сильно пигментированный с вентральной поверхности темно-бордового цвета орган, способный при секреции биссусных нитей вытягиваться, увеличиваясь в длину в 2 раза. С дорсальной поверхности нога желтого цвета.
Результаты показали, что у C. grayanus размером 40 мм длина ноги 5.2 ± 0.4 мм, ширина ноги 3.3 ± 0.5 мм. Нога окрашена в оранжевый цвет с двух сторон. Как и в случае с Myt. coruscus, мидия С. grayanus способна при формировании биссусных нитей удлинять ногу в 2‒3 раза по сравнению с ее оригинальной длиной.
У модиолуса Mod. modiolus размером 40 мм длина ноги 4.2 ± 0.2 мм, ширина ноги 3.3 ± 0.2 мм. Нога полностью окрашена в желтый цвет. В процессе формирования биссусных нитей нога может удлиняться в 4 раза.
Строение биссусной бороздки ноги
На вентральной поверхности ноги Myt. coruscus, C. grayanus и Mod. modiolus расположена биссусная бороздка, где образуется биссус. Согласно результатам СЭМ, у этих видов бороздка располагается вдоль центральной оси ноги и ее длина приближается к длине ноги (рис. 1А, 1Б; 2А, 2Б; 3А, 3Б).
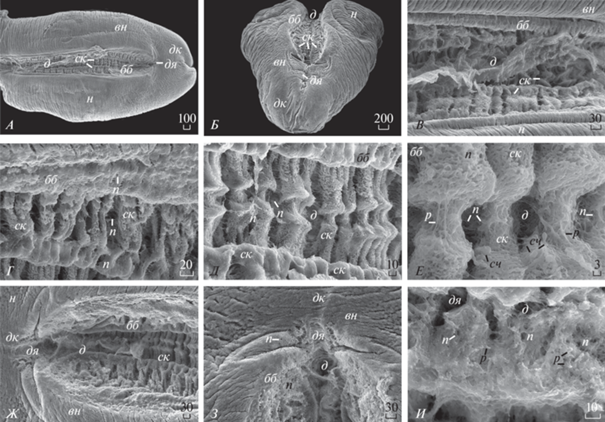
Рис. 1.
Морфологическое строение (мкм) биссусной бороздки ноги у мидии блестящей (Mytilus coruscus) (СЭМ): А – вентральная поверхность ноги и биссусной бороздки, вид сверху; Б – дистальный конец ноги, фронтально; В – биссусная бороздка, вид сверху; Г – боковая стенка биссусной бороздки, увеличение; Д – дно биссусной бороздки, вид сверху; Е ‒ дно и боковая поверхность стенки биссусной бороздки, увеличение; Ж – дистальная ямка на вентральной поверхности ноги с участком биссусной бороздки; З – дистальная ямка на дистальном конце ноги, увеличение; И – дно дистальной ямки ноги, увеличение.

По данным СЭМ, длина ноги у мидии Myt. coruscus 3870‒4450 мкм. Длина биссусной бороздки ноги 3850‒4250 мкм (рис. 1А). В отличие от двух других видов, у Myt. coruscus с двух сторон вдоль биссусной бороздки ноги тянутся продольные смыкающие складки (рис. 1А–1В). Ширина ноги в центральной части 2367‒2680 мкм (рис. 1Б). Ширина биссусной бороздки в этой части ноги 750–875 мкм (рис. 1В). Глубина биссусной бороздки 233–275 мкм (рис. 1Б, 1Г). Складки стенок биссусной бороздки ноги имеют форму, напоминающую квадраты и прямоугольники разного размера (рис. 1А, 1В, 1Г). Поверхность складок стенок и дна биссусной бороздки покрыта многочисленными порами (рис. 1Г, 1Д). Ширина складок стенки биссусной бороздки варьирует и составляет 90.0–187.5 мкм (рис. 1Г). У Myt. coruscus стенки биссусной бороздки располагаются относительно параллельно друг другу (рис. 1А‒1В, 1Д, 1Ж). Ширина дна биссусной бороздки 120‒135 мкм (рис. 1Д). На дне биссусной бороздки ноги образуются складки в виде поперечных прямоугольников (рис. 1Д). Ширина складок дна биссусной бороздки ноги 40.0–67.0 мкм. Поверхность дна биссусной бороздки ноги покрыта многочисленными порами и ресничками (длиной 3.0–3.75 мкм) и сосочками (длиной 2.0‒2.25 мкм) (рис. 1Д, 1Е). На дистальном конце ноги располагается чашевидная, хорошо заметная дистальная ямка (ее поперечная ширина 250‒270 мкм, продольная ширина ямки 128‒138 мкм) (рис. 1Ж, 1З). Поверхность дистальной ямки покрыта многочисленными, хорошо заметными порами (рис. 1И). Длина многочисленных ресничек на дне дистальной ямки 2.0–3.3 мкм (рис. 1И). Длина сосочков на дне дистальной ямки 0.7–1.0 мкм (рис. 1И). В этом месте нога более узкая. Биссусная бороздка ноги также сужается на дистальном конце ноги, ее ширина здесь 86‒112 мкм (рис. 1Ж).
Результаты СЭМ показали, что длина ноги у мидии C. grayanus 3700‒4500 мкм, ширина ноги в ее центральной части 2280‒2500 мкм (рис. 2А, 2Б). Длина биссусной бороздки 2050‒2365 мкм (рис. 2А). Глубина биссусной бороздки 262‒ 343 мкм (рис. 2Б). Ширина биссусной бороздки в центральной части ноги 300‒320 мкм (рис. 2В). Стенки и особенно дно биссусной бороздки ноги имеют длинные и округлые складки, напоминающие по внешнему виду колонны, которые сужаются или расширяются вдоль своей длины (рис. 2Д). Ширина складок стенки биссусной бороздки 10.0–30.0 мкм (рис. 2Г). Поверхность стенок и дна биссусной бороздки покрыта многочисленными порами (рис. 2Г‒2Е). У C. grayanus стенки биссусной бороздки располагаются параллельно друг другу (рис. 2А‒2В, 2Д). Ширина дна биссусной бороздки 125‒200 мкм (рис. 2Д). Ширина складок дна биссусной бороздки 19.5–36.0 мкм (рис. 2Д, 2Е). Дно биссусной бороздки покрыто ресничками (длиной 10.5–13.5 мкм) и сосочками (длиной 3.75‒4.5 мкм) (рис. 2Е). На дистальном конце нога сужается, также сужается биссусная бороздка, ширина которой в этом месте 115‒240 мкм (рис. 2Ж). На дистальном конце ноги располагается дистальная ямка, покрытая многочисленными порами (рис. 2А, 2З). По внешнему виду дистальная ямка напоминает полумесяц (рис. 2Ж, 2З). Поперечная длина дистальной ямки 300‒400 мкм (рис. 2Ж, 2З). Продольная длина дистальной ямки 30.0‒50.0 мкм (рис. 2Ж, 2З). Длина ресничек на дне дистальной ямки 7.0–8.0 мкм (рис. 2И).
Рис. 2.
Морфологическое строение (мкм) биссусной бороздки ноги у мидии Грея (Сrenomytilus grayanus) (СЭМ): А – вентральная поверхность ноги и биссусной бороздки, вид сверху; Б – дистальный конец ноги, фронтально; В – биссусная бороздка, вид сверху; Г – боковая стенка биссусной бороздки, увеличение; Д – дно биссусной бороздки, вид сверху; Е – дно и боковая поверхность стенки биссусной бороздки, увеличение; Ж – дистальная ямка на вентральной поверхности ноги с участком биссусной бороздки; З – дистальная ямка на дистальном конце ноги, увеличение; И – дно дистальной ямки ноги, увеличение.
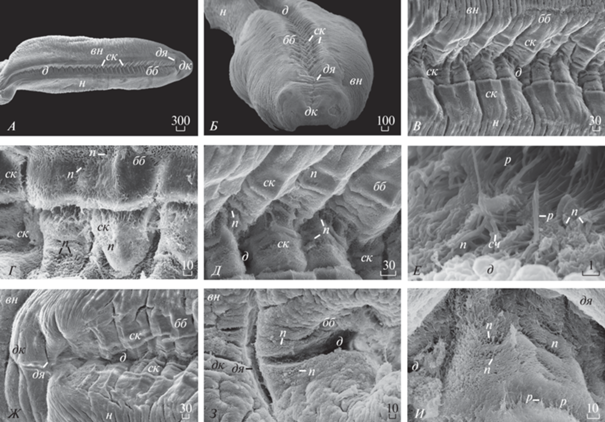
Результаты СЭМ показали, что длина ноги у модиолуса Mod. modiolus 4457‒5225 мкм, ширина ноги 1114.3‒1500 мкм (рис. 3А, 3Б). Длина биссусной бороздки, которая проходит вдоль центральной оси ноги, 3857‒4125 мкм (рис. 3А). Ширина биссусной бороздки может варьировать вдоль ноги моллюска и составляет в ее центральной части 300‒375 мкм (рис. 3В). Глубина биссусной бороздки варьирует вдоль ее длины и составляет 115–180 мкм (рис. 3Б, 3В). Стенки биссусной бороздки образуют складки под углом около 90° и более (рис. 3Д). Ширина складок 75–100 мкм. Поверхность складчатых стенок и дна биссусной бороздки покрыты хорошо заметными, многочисленными порами, ресничками и сосочками (рис. 3Д, 3Е). У модиолуса стенки биссусной бороздки соединяются под углом, образуя узкий, зигзагообразный желоб на дне бороздки (рис. 3А‒3В, 3Д). Ширина дна биссусной бороздки 45‒65 мкм (рис. 3Д). Ширина складок дна биссусной бороздки 66–126 мкм (рис. 3Д). Длина ресничек на дне бороздки 2.6–3.2 мкм. Длина сосочков на дне биссусной бороздки 1.0‒1.35 мкм (рис. 3Е). К дистальному концу нога модиолуса сужается и ее ширина на самом конце 429‒ 600 мкм (рис. 3А). Здесь рядом располагается длинная, щелевидная дистальная ямка, покрытая многочисленными порами (рис. 3А, 3Ж, 3З). Поперечная длина дистальной ямки 165‒183 мкм, продольная длина дистальной ямки 8.5‒10.0 мкм (рис. 3Ж, 3З). Дно дистальной ямки покрыто многочисленными порами и ресничками длиной 5.0–7.5 мкм.
Рис. 3.
Морфологическое строение (мкм) биссусной бороздки ноги у модиолуса (Modiolus modiolus) (СЭМ): А – вентральная поверхность ноги и биссусной бороздки, вид сверху; Б – дистальный конец ноги, фронтально; В – биссусная бороздка, вид сверху; Г – боковая стенка биссусной бороздки, увеличение; Д – дно биссусной бороздки, вид сверху; Е – дно и боковая поверхность стенки биссусной бороздки, увеличение; Ж – дистальная ямка на вентральной поверхности ноги с участком биссусной бороздки; З – дистальная ямка на дистальном конце ноги, увеличение; И – дно дистальной ямки ноги, увеличение.
ОБСУЖДЕНИЕ
Наши результаты СЭМ и сравнительный анализ литературных данных показали, что нога митилид Myt. coruscus, C. grayanus и Mod. modiolus, как и у Myt. trossulus (Vekhova, 2021; Вехова, 2021), преобразована в биссусообразующий орган, имеющий биссусную бороздку, где образуется биссус (Carter et al., 2012). У исследованных видов митилид бороздка располагается на вентральной стороне ноги вдоль центральной оси и ее длина приближается к длине ноги. Наряду со сходством эти виды характеризуются также некоторыми различиями особенностей морфологии биссусной бороздки, обусловленными обитанием этих видов митилид в разных биотопах (Vekhova, 2013; Скарлато и др., 1967; Скарлато, 1981).
У одноразмерных вскрытых моллюсков размером 40 мм самую длинную и широкую ногу имеет мидия Myt. coruscus (длина ноги 7.65 ± 0.4, ширина 5.0 ± 0.3 мм). У трех других видов различия в длине и ширине ноги носят случайный характер. У C. grayanus длина ноги составляет 5.2 ± 0.4, ширина ноги 3.3 ± 0.5 мм, у Mod. modiolus и у Myt. trossulus размер ноги сходный (длина 4.2 ± 0.2, ширина 3.3 ± 0.2 мм).
Мы сравнили результаты СЭМ строения биссусной бороздки ноги у исследуемых видов митилид из Японского моря. По нашим предыдущим данным (Vekhova, 2021; Вехова, 2021), Myt. trossulus, Myt. coruscus, C. grayanus и Mod. modiolus имеют особенности длины, ширины и глубины биссусной бороздки (рис. 1А–1Г; 2А–2Г; 3А–3Г). Длина биссусной бороздки у C. grayanus 2050‒2365 мкм, что значительно меньше, чем у других видов митилид. У Myt. coruscus длина биссусной бороздки 3850‒4250 мкм, у Mod. modiolus 3857‒4125, у Myt. trossulus – 3600 мкм. Наиболее широкая биссусная бороздка у Myt. coruscus, ширина которой 750–875 мкм. У C. grayanus этот параметр 240‒ 320 мкм, у остальных двух видов – Mod. modiolus 300‒375 мкм, Myt. trossulus 286–300 мкм. Самую глубокую биссусную бороздку имеет C. grayanus (300‒343 мкм). У Myt. coruscus глубина бороздки составляет 233–275 мкм. Отметим, что других два вида не различаются глубиной биссусной бороздки: у Mod. modiolus 115–180 мкм, а у Myt. trossulus 70.0–250 мкм. При этом в отличие от трех других видов только Myt. coruscus имеет биссусную бороздку в виде смыкающихся складок, проходящих по двум сторонам от биссусной бороздки (рис. 1А–1В). Все это объясняет различия в размере биссуса и биссусных нитей у этих видов и других видов митилид (Brown, 1952; Tamarin, Keller, 1972; Bairati, Vitellaro-Zuccarello, 1974; Tamarin, 1975; Allen et al., 1976; Vekhova, 2019, 2021; Бергер и др., 1985; Вехова, 2007, 2021).
Сравнительный анализ наших результатов СЭМ и литературных данных (Vekhova, 2021; Вехова, 2021) свидетельствует о различиях в особенностях строения и размерах стенок, а также дна биссусной бороздки ноги у Myt. coruscus, C. grayanus, Mod. modiolus и Myt. trossulus (рис. 1Г, 1Д; 2Г, 2Д; 3Г, 3Д). У Myt. coruscus стенки и дно биссусной бороздки образуют складки в форме прямоугольников разной ширины (у стенок 90.0–187.5, у дна 40.0–67.0 мкм), которые расположены параллельно друг другу. У Myt. coruscus ширина дна биссусной бороздки 120‒135 мкм. Дно выстилают реснички размером 3.0–3.75 и сосочки длиной 2.0‒2.25 мкм (рис. 1Г–1Е). У C. grayanus складки стенок (шириной 10.0–30.0 мкм) расположены параллельно друг другу и складки дна (шириной 19.5–36.0 мкм) имеют форму колон, которые сужаются или расширяются вдоль их длины. У C. grayanus ширина дна биссусной бороздки составляет 125‒200 мкм; дно выстлано ресничками размером 10.5–13.5 мкм и сосочками размером 3.75‒4.5 мкм (рис. 2Г–2Е). В отличие от Myt. coruscus и C. grayanus, у Mod. modiolus стенки биссусной бороздки образуют складки под углом 90°, которые сужаются ко дну и образуют зигзагообразный желоб на дне (рис. 3В–3Д). Ширина складок стенок может варьировать и составляет в среднем 75–100 мкм. У Mod. modiolus ширина дна биссусной бороздки 45‒65 мкм. Дно биссусной бороздки у Mod. modiolus покрыто ресничками длиной 2.6–3.2 мкм и сосочками длиной 1.0‒ 1.35 мкм (рис. 3Е). По данным автора (Vekhova, 2021; Вехова, 2021), у Myt. trossulus стенки биссусной бороздки образуют округлые складки размером 100–200 мкм, которые сужаются ко дну (их ширина 50.0–67.0 мкм), образуя на дне зигзагообразный желоб, чем напоминают биссусную бороздку модиолуса Mod. modiolus. По данным автора (Vekhova, 2021; Вехова, 2021), у Myt. trossulus ширина дна биссусной бороздки 65 мкм. При этом стенки и дно биссусной бороздки покрыты ресничками длиной 1.5–2.0 мкм и сосочками длиной 3.4 мкм.
Литературные данные (Tamarin, Keller, 1972; Eckroat, Steel, 1993; Vekhova, 2021; Вехова, 2021) и анализ полученных результатов показали (рис. 1А, 1Б; 2А, 2Б; 3А, 3Б), что на дистальном конце нога у Myt. coruscus, C. grayanus, Mod. modiolus и Myt. trossulus, как и у других видов митилид, сужается. Также сужается покрытая многочисленными порами биссусная бороздка, образуя на конце дистальную ямку (рис. 1Ж, 1З; 2Ж, 2З; 3Ж, 3З). Все эти виды различаются между собой строением и размерами дистальной ямки. У Myt. coruscus дистальная ямка чашевидная (ее поперечная ширина 250‒270, продольная ширина 128‒138 мкм). У Myt. coruscus на дне дистальной ямки длина ресничек 2.0–3.3 мкм, длина сосочков 0.7–1.0 мкм (рис. 1Ж‒1И). У C. grayanus дистальная ямка напоминает полумесяц (ее поперечная ширина 300‒400, продольная ширина 30.0‒50.0 мкм). У C. grayanus длина ресничек на дне дистальной ямки 7.0–8.0 мкм, сосочки не обнаружены (рис. 2Ж‒2И). У Mod. modiolus щелевидная дистальная ямка (поперечная ширина дистальной ямки 165‒183, продольная ширина 8.5‒10.0 мкм). У Mod. modiolus длина ресничек составляет 5.0–7.5 мкм, сосочки не обнаружены (рис. 3Ж‒3И). По данным автора (Vekhova, 2021; Вехова, 2021), у Myt. trossulus дистальная ямка узкая щелевидная (поперечная ширина 180, продольная ширина 5.0 мкм). У этого вида дно биссусной бороздки покрыто ресничками длиной 1.0 мкм и сосочками длиной 0.7–1.0 мкм.
Согласно литературным данным, при сходстве внешнего строения биссуса и биссусных нитей у Myt. coruscus, C. grayanus, Mod. modiolus и Myt. trossulus размеры прикрепительных дисков и биссусных нитей и степень их армированности существенно различаются (Vekhova, 2019, 2021; Вехова, 2007, 2021). Показано, что формирование биссуса тесно связано со сходной для митилид природой механических процессов, протекающих в биссусной бороздке и дистальной ямке ноги в момент формирования биссусной нити и ее прикрепления к поверхности субстрата (Pujol, 1967; Price, 1983; Waite, 1983). В значительной мере различия в строении биссусной нити вызваны морфологическими особенностями строения биссусной бороздки ноги, ее размером и топографией поверхности у этих митилид (рис. 1А–1И; 2А–2И; 3А–3И). По-видимому, не последнюю роль здесь играют формы складок стенок и дна биссусной бороздки (Carter et al., 2012). Многочисленные поры, которые пронизывают стенки и дно биссусной бороздки ноги, а также дистальную ямку, – это протоки желез, которые также играют важную роль в процессе образования биссуса. Биссусообразование – биохимический процесс, при этом секреты пяти желез ноги вытекают через протоки желез в биссусную бороздку. Поверхность биссусной бороздки ноги покрыта сосочками и многочисленными ресничками, с помощью которых при формировании биссусных нитей секреты всех желез смешиваются в вентральной бороздке ноги у этих видов митилид (рис. 1Г–1Е; 2Г–2Е; 3Г–3Е). Важную роль при формировании биссуса имеют форма и строение дистальной ямки, где образуется прикрепительный диск биссусной нити (рис. 1Ж–1И; 2Ж–2И; 3Ж–3И), которые существенно различаются у разных видов митилид (Pujol, 1967; Price, 1983; Eckroat, Steel, 1993; Vekhova, 2021; Вехова, 2021). Известно, что в биохимическом процессе формирования биссусной нити важную роль играет количество секрета, вырабатываемого каждой из пяти желез ноги. На этот процесс также оказывает влияние скорость его смешивания с помощью многочисленных ресничек и сосочков, выстилающих стенки и дно биссусной бороздки ноги (рис. 1Д, 1Е, 1З, 1И; 2Д, 2Е, 2З, 2И; 3Д, 3Е, 3З, 3И). Образование нового биссуса происходит после поступления продуктов секреции пяти желез в биссусную бороздку и дистальную ямку ноги (Allen et al., 1976; Бергер и др., 1985), посредством сокращения мощной мускулатуры ноги и в результате дальнейшего задубливания фенолами (Allen et al., 1976; Price, 1983; Vekhova, 2021; Вехова, 2021).
Таким образом, биссус митилид имеет неклеточное белковое строение (Waite, 1983, 1997). Это продукт синтеза биохимических веществ (включая коллаген), секретирующихся в пяти железах ноги. В момент образования биссусной нити большое количество продуктов секреции желез попадает через многочисленные протоки в биссусную бороздку и там смешивается ресничками и сосочками, затем происходит фенольное задубливание белкового материала, в т.ч. автозадубливание биссуса (Pujol, 1967; Price, 1983). Предположительно, морфологическое разнообразие в строении биссуса у Myt. coruscus, C. grayanus, Mod. modiolus и Myt. trossulus вызвано структурными особенностями секреторного органа, поскольку образование биссусной нити в биссусной бороздке ноги происходит по принципу отливки в фильерах (Vekhova, 2019, 2021; Вехова, 2007, 2021).
Буквенные обозначения на рис. 1, 2 и 3: бб – биссусная бороздка ноги, вн – вентральная поверхность ноги, д ‒ дно биссусной бороздки, дк ‒ дистальный конец, дя – дистальная ямка, н – нога моллюска, п ‒ протоки желез, р ‒ реснички биссусной бороздки, ск – складки биссусной бороздки, см ‒ смыкающие складки биссусной бороздки ноги, сч ‒ сосочек биссусной бороздки ноги.
Список литературы
Бергер В.Я., Летунов В.Н., Вшевцов Г.В., Саранчова О.Л., 1985. Морфофункциональные и экологические аспекты биссусообразования у мидии (Mytilus edulis L.) // Экология обрастания в Белом море. Л.: ЗИН АН СССР. С. 67‒75.
Вехова Е.Е., 2007. Сравнительная морфология биссусных нитей трех представителей семейства Mytilidae (Bivalvia) из Японского моря // Зоологический журнал. Т. 86. № 2. С. 154‒162.
Вехова Е.Е., 2021. Биссусный аппарат мидии тихоокеанской (Mytilus trossulus, Bivalvia, Mytilidae) из Японского моря // Зоологический журнал. Т. 100. № 5. С. 483‒492.
Золотарев В.Н., 1989. Склерохронология морских двустворчатых моллюсков. Киев: Наукова думка. 112 с.
Лукин В.И., Фадеев В.И., 1982. Особенности планирования гидробиологических работ на акваториях большой протяженности // Подводные гидробиологические исследования. Владивосток: ДВНЦ АН СССР. С. 13‒20.
Миронов А.А., Комиссарчик Я.Ю., Миронов В.А., 1994. Методы электронной микроскопии в биологии и медицине: методическое руководство. СПб.: Наука. 400 с.
Скарлато О.А., Голиков А.Н., Василенко С.В. и др., 1967. Состав, структура и распределение донных биоценозов в прибрежных водах залива Посьет (Японское море) // Исследования фауны морей. Т. 5. № 13. С. 5‒61.
Скарлато O.A., 1981. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука. 479 с.
Allen J.A., Cook M., Jackson D.J. et al., 1976. Observations on the rate of production and mechanical properties of the byssus threads of Mytilus edulis L. // Journal of Molluscan Studies. V. 42. № 2. P. 279‒289.
Brown C.H., 1952. Some structural proteins of Mytilus edulis // The Quarterly Journal of Microscopical Science. V. 93. P. 487‒502.
Bairati A., Vitellaro-Zuccarello L., 1974. The ultrastructure of the byssal apparatus of Mytilus galloprovincialis. II. Observations by microdissection and scanning electron microscopy // Marine Biology. V. 28. P. 145‒158.
Carter J.G., Harries P.J., Malchus N., Sartori A.F., Anderson L.C., Bieler R., Bogan A.E., Coan E.V., Cope J.C.W., Cragg S.M., Cartἰa-March J.R., Hylleberg J., Kelley P., Kleemann K., Kříž J., McRoberts C., Mikkelsen P.M., Pojeta J., Jr., Tëmkin I., Yancey T., Zieritz A., 2012. Illustrated Glossary of the Bivalvia. Treatise online № 48. V. 1. P. 15.
Denny M.W., 1987. Lift as a mechanism of patch initiation in mussel beds // Journal of Experimental Marine Biology and Ecology. V. 113. P. 231‒245.
Denny M., Gaylord B., Helmuth B., Daniel T., 1998. The menace of momentum: Dynamic forces of flexible organisms // Limnology and Oceanography. V. 43. № 5. P. 955‒968.
Eckroat L.R., Steel L.M., 1993. Comparative morphology of the byssi of Dreissena polymorpha and Mytilus edulis // American Malacological Bulletin. V. 10. P. 103‒108.
Lutaenko K.A., Noseworthy R.G., 2012. Catalogue of the Living Bivalvia of the Continental Coast of the Sea of Japan (East Sea). Vladivostok: Dalnauka. 247 p.
Price H.A., 1983. Structure and formation of the byssus complex in Mytilus (Mollusca, Bivalvia) // Journal of Molluscan Studies. V. 49. № 1. P. 9−17.
Pujol J.P., 1967. Le complex byssogéne des mollusques bivalves: Histochimie comparee des secretions chez Mytilus edulis et Pinna nobilis // Bulletin de la Société Lennéenne de Normandie. V. 10. P. 308–332.
Tamarin A., Keller P.J., 1972. An ultrastructural study of the byssal thread forming system in Mytilus // Journal of Ultrastructure Research. V. 40. P. 401–416.
Tamarin A. 1975. An ultrastructural study of byssus stem formation in Mytilus califomianus // Journal Morphology. V. 145. P. 151–178.
Vekhova E.E., 2006. Reattachment of certain species of mytilid bivalves to various substrates // Russian Journal of Marine Biology. V. 32. № 5. P. 308‒311.
Vekhova E.E., 2013. Growth and shell morphology of three Mytilidae (Bivalvia) species from the Sea of Japan // Biology Bulletin. V. 40. № 9. P. 728–737.
Vekhova E.E., 2019. The adaptive morphology of byssus in Mytilus coruscus, Crenomytilus grayanus, and Modiolus modiolus (Mytilidae, Bivalvia) from the Sea of Japan // Biology Bulletin. V. 46. № 9. P. 1030–1044.
Vekhova E.E., 2021. The byssal apparatus in the Pacific mussel, Mytilus trossulus (Bivalvia, Mytilidae), from the Sea of Japan // Biology Bulletin. V. 48. № 9. P. 1443‒1451.
Waite J., 1983. Adhesion in byssally attached bivalves // Biological Reviews of the Cambridge Philosophical Society. V. 58. № 2. P. 209–231.
Waite J.H., 1997. Marine bioadhesion: unraveling the chemistry // Journal of The Adhesion Society of Japan. V. 33. № 5. P. 186–194.
Yonge C.M., 1962. On the primitive significance of the byssus in the Bivalvia and its effects in evolution // Journal of the Marine Biological Association of the United Kingdom. V. 42. № 1. P. 113–125.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал