

Зоологический журнал, 2022, T. 101, № 7, стр. 736-753
Особенности фауны и населения ногохвосток (Hexapoda, Collembola) Восточного Ямала
А. Б. Бабенко a, *, М. Д. Антипова a, **
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: lsdc@mail.ru
** E-mail: antimar.2410@gmail.com
Поступила в редакцию 10.12.2021
После доработки 20.12.2021
Принята к публикации 16.01.2022
- EDN: ADWRWK
- DOI: 10.31857/S004451342205004X
Аннотация
Обследовано население почвенных ногохвосток в окрестностях пос. Сеяха, расположенного на побережье Обской губы в центральной части Ямала. В 13 растительных ассоциациях, отражающих разнообразие растительности данного тундрового района, обнаружено 104 вида ногохвосток 39 родов из 13 семейств. Проведенный анализ свидетельствует о некоторой обедненности данной конкретной фауны по сравнению со среднесибирскими аналогами. При этом группировки ногохвосток отдельных местообитаний в обследованном районе Ямала не обладают заметными особенностями, в корне отличающими их от аналогичных комплексов в других районах южной части тундровой зоны. По составу и структуре они вполне соответствуют своему зональному положению. Пограничное положение региона между Европой и Азией проявляется лишь в присутствии в фауне небольшого числа западно-палеарктических видов, не играющих, однако, существенной роли в большинстве почвенных группировок.
Полуостров Ямал занимает пограничное положение между Европой и Азией и, несомненно, представляет собой ключевой район, важный для понимания генезиса биоты северной Палеарктики. Вместе с тем данных о фауне беспозвоночных этого региона удивительно мало, причем подавляющее большинство из них посвящено самой южной части полуострова, расположенной в лесотундре. Среди беспозвоночных относительно неплохо изучены только жужелицы (Коробейников, 1980, 1987; Андреева, Еремин, 1991; Рябицев, 1997, 1998; Ковалев и др., 2017), имеется довольно много фаунистических данных о пауках (Esyunin, Efimik, 1996; Tanasevitch et al., 2009; Tanasevitch, Rybalov, 2015). Специальных эколого-фаунистических исследований, посвященных мелким почвенным членистоногим, на территории полуострова практически не проводилось. В частности, имеющиеся в литературе сведения по клещам, одной из основных групп почвенного населения, ограничены самой южной частью полуострова (Гришина, 1985; Давыдова, Никольский, 1986; Давыдова, 1988; Марченко, 2012). Редкое исключение – работа, выполненная на о-ве Белый, северной оконечности полуострова (Макарова и др., 2015). Имеется также ряд указаний на находки в регионе отдельных видов клещей, что связано с картированием их ареалов (Макарова, 2000, 2009, 2012) или со сравнениями при описании новых форм (Линдквист, Макарова, 2011; Lindquist, Makarova, 2012). Еще меньше известно о ногохвостках, второй из основных групп микроартропод в почвах тундровой зоны. Все имеющиеся данные по этой группе в регионе основаны на кратковременных фаунистических сборах одного из авторов данной статьи в двух точках северного Ямала в ходе экспедиции “Экология тундры-94”, а также на случайных материалах из самой южной части полуострова. Эти данные были использованы в обзорных статьях и определителях (Бабенко и др., 1994; Babenko, 1997; Potapov, Babenko, 2000; Babenko, Fjellberg, 2006), но специальных публикаций по фауне ногохвосток полуострова нет. Работ, посвященных анализу структуры их сообществ в пределах данного региона, также нет. В свете всего вышесказанного основная задача настоящего сообщения – первичная инвентаризация фауны и описание особенностей группировок коллембол в одном из районов полуострова – представляется нам вполне оправданной.
МАТЕРИАЛ И МЕТОДИКА
Приведенные ниже данные по ногохвосткам основаны на сборах, которые были проведены авторами в июле 2020 г. в окрестностях пос. Сеяха, восточный Ямал (70.1699° с.ш., 72.5153° в.д.). Район исследований расположен на правом берегу р. Сеяха вблизи ее впадения в Обскую губу и представляет собой всхолмленную равнину, где относительно низкие гряды холмов с максимальными высотами в 20–30 м чередуются с многочисленными озерными и болотными котловинами (рис. 1а). Средняя температура июля по данным местной метеостанции составляет 8.4°С (диапазон колебаний за последние 70 лет от 4° до 15°С) (www.pogodaiklimat.ru). Почвы района, как правило, легкие (песчаные и супесчаные), что в целом характерно для Ямала как аккумулятивной морской равнины.
Рис. 1.
Обследованные растительные ассоциации: а – полуостров Ямал в районе исследований; б – зональная ассоциация: кустарничково-мохово-лишайниковая полигональная тундра; в – кустарничковая ассоциация.

Учеты ногохвосток были проведены в основных растительных ассоциациях, более или менее полно отражающих разнообразие растительности данного района. Зональным типом растительности мы считали занимающие выположенные вершины холмов полигональные тундры (рис. 1б) с кустарничково-лишайниково-моховым покровом на полигонах и моховой дерниной с осоками и единичными кустиками низких ивок в трещинах. На наиболее сухих и теплых перегибах рельефа развиты ивково-дриадовые ассоциации с большой долей оголенных грунтов (рис. 1в), а в обширных озерных котловинах широко представлены разнообразные болотные комплексы (рис. 2а и 2б). Относительно высокие ивняки приурочены лишь к средним частям склонов (рис. 3б). Встречаются в обследованном районе и нивальные ассоциации на месте позднего схода снега (рис. 3в), а также небольшие разнотравные луговины, связанные с несколько повышенной термообеспеченностью южных и юго-восточных склонов и/или с зоогенностью (рис. 3а). Помимо этого, были обследованы маршевые комплексы, сформированные на заливаемых пляжах в эстуарии р. Сеяха (рис. 2в).
Рис. 2.
Обследованные растительные ассоциации: а – озерная котловина с осоково-ивовым болотом, б – морошка на сфагновых буграх, в – береговые марши.

Рис. 3.
Обследованные растительные ассоциации: а – разнотравный лужок с полынью, б – ивняк хвощевый, в – снежник в распадке.

Во всех перечисленных местообитаниях были проведены качественные учеты ногохвосток с помощью почвенных ловушек (всего 1540 ловушко/сут). В тех же биотопах были отобраны почвенные образцы (5 × 5 × 5 см) для последующей экстракции микроартропод в лабораторных условиях. Повторность при этих количественных учетах варьировала от 5 до 9 образцов на биотоп. Экстракцию микроартропод из почвенных монолитов проводили в Москве без дополнительного подогрева в течение 7–8 сут. В целом было обработано 92 почвенных образца, из которых извлечено и определено более 48 тыс. экз. коллембол. Кроме того, для более полного выявления богатства конкретной фауны проводили ручной сбор ногохвосток с помощью эксгаустера и сифтованием.
В пяти типичных местообитаниях района измеряли температуры поверхностного слоя почв с помощью дата-логгеров. Несмотря на весьма незначительную продолжительность периода измерений (09–19 июля), результаты явно свидетельствуют о достаточно заметных различиях в термообеспеченности разных биотопов (табл. 1).
Таблица 1.
Температуры поверхностного слоя почвы разных местообитаний в период исследований (09–19 июля 2020)
| Показатель | Дриадник | Луг | Снежник | Болото | Зональная тундра |
|---|---|---|---|---|---|
| Среднесуточная температура | 9.3 | 10.7 | 6.8 | 8.8 | 8.5 |
| Медиана | 9.0 | 9.8 | 6.5 | 8.3 | 7.5 |
| Максимум | 14.0 | 21.0 | 16.0 | 16.0 | 19.0 |
| Минимум | 5.5 | 5.0 | 3.5 | 3.5 | 3.5 |
| Сумма | 1120.3 | 1291.5 | 837.1 | 1056.5 | 1028.7 |
| Общее число измерений | 120 | 120 | 120 | 120 | 120 |
Статистическая обработка материалов и визуализация полученных результатов осуществлялась с помощью программы PAST 3.12 (Hammer et al., 2001). Для проверки нулевой гипотезы о равномерном распределении отдельных видов ногохвосток в пределах обследованных растительных ассоциаций и для выделения видов, наиболее “значимых” для разграничения группировок, использован непараметрический критерий χ2.
РЕЗУЛЬТАТЫ
Фауна. Обнаружено 104 вида из 39 родов 13 семейств (Приложение 1 ). Из них 13 видов ногохвосток были собраны только вручную (ловушки, сифтер и/или эксгаустер). По своей структуре эта конкретная фауна (определение см. Чернов, 1978) достаточно обычна для северных безлесных ландшафтов. В ней безраздельно господствует семейство Isotomidae, представленное 39 видами, что составляет почти 37% общего видового богатства. Такой “изотомоидный” тип фаун (Капрусь, 2010) свойствен практически всем бореальным районам Палеарктики. Второе и третье места делят семейства Hypogastruridae (17 видов, 16%) и Neanuridae (16 видов, 15%). Замыкают пятерку наиболее крупных семейств в обследованной конкретной фауне Onychiuridae (12 видов, 11%) и Sminthurididae (9 видов, 8.5%). Достаточно традиционен для тундровых районов и набор родов, наиболее богатых видами: Folsomia (14 видов), Desoria (9 видов), Hypogastrura и Anurida (по 7 видов), а также Protaphorura и Sminthurides (по 6 видов). Немногим больше 50% родов конкретной фауны одновидовые, но общее соотношение вид/род довольно велико и равно 2.7.
Ареалогический состав фауны ногохвосток окрестностей пос. Сеяха весьма пестрый (рис. 4). Помимо широко распространенных форм, т.е. космополитов и трансголарктов, фауна включает транспалеарктов, множество восточно-палеарктических форм и несколько западно-палеарктических видов, для которых в данном регионе (или вблизи него) проходит восточная граница известного ареала. Особенно интересна именно эта последняя западно-палеарктическая группа, включающая такие виды как Protaphorura stogovi Pomorski 1993, Ceratophysella scotica (Carpenter et Evans 1899), Anurida balatovae Rusek 1970, Anurida ellipsoides Stach 1949, Desoria tolya Fjellberg 2007, Isotoma anglicana Lubbock 1862 и Entomobrya corticalis (Nicolet 1842). Их общее число, конечно, невелико, но само их присутствие в фауне региона достаточно показательно. Состав фауны обследованной территории гетерогенен и в зонально-широтном плане. При этом криофильные формы составляют около 40% видового списка. Среди них наиболее широко представлены гемиаркты, а число эварктов и гипоарктов невелико.
Рис. 4.
Ареалогический состав фауны ногохвосток окрестностей пос. Сеяха: а – широко распространенные формы (космополиты и трансголаркты), б – транспалеаркты, в – восточно-палеарктические формы, г – западно-палеарктические формы, д – виды с неясным распространением.

Особого упоминания заслуживает только несколько выявленных видов. В первую очередь, это A. balatovae, обитатель разного типа болот в районе исследования. Ранее вид был известен только для центральной Европы: отмечался на юго-востоке Чехии (Rusek, 1970, 1977, 1995) и в Польше (Pomorski, 1994; Skarżyński, Pomorski, 1995), причем также в переувлажненных местообитаниях.
Примечательна для столь северного региона находка Vertagopus asiaticus Potapov et al. 2016. Вид широко распространен в азиатской части России (от Урала до Курильских о-вов), встречается в северном и северо-западном Китае, но в Арктике до настоящего времени не отмечался (Potapov et al., 2016, стр. 119). Его ближайшее к пос. Сеяха местонахождение находится примерно на 1500 км южнее на Среднем Урале (Свердловская обл.). В обследованном районе V. asiaticus в массе населяет гниющий плавник в устье р. Сеяха, но, в отличие от совместно встречающегося V. pseudocinereus Fjellberg 1975, не был отмечен вне прибрежной полосы. Недавнее обнаружение единичных экземпляров V. asiaticus в ближайшем Подмосковье (Potapov, Janion-Scheepers, 2019; Vasenkova, Kuznetsova, 2022) может служить весомым аргументом в пользу высокого инвазионного потенциала этого вида. Вероятно, и в нашем случае можно полагать, что его присутствие в центральной части восточного Ямала объясняется пассивным переносом из более южных таежных районов вдоль побережья Обской губы.
Следует также отметить присутствие в составе фауны Hypogastrura lapponica (Axelson 1902). В отличие от ранее упомянутых видов, единичные находки этой северо-бореальной формы в южной половине тундровой зоны известны (см., например, Бабенко, 2003). В обследованном районе этот вид, обычно считающийся облигатным кортициколом, приурочен к зоогенным луговинам в пределах зональных сообществ (рис. 5). Интересна находка, вероятно, нового вида рода Micranurida (приведен в списке как Micranurida sp. aff. balta Fjellberg 1998). Как и M. balta, он имеет удлиненную латеральную сенсиллу S8 на четвертом членике усиков, но характеризуется наличием хет p2 на всех сегментах от Th.2 до Abd.4 и полным набором хет на тибиотарзусах. Для второго близкого вида рода, M. rostrata Babenko 2007, известного из азиатской части России, также характерна существенная редукция числа тибиотарзальных хет. К сожалению, в наших материалах имеется лишь один экземпляр этого вида. Обнаружение в регионе всех остальных перечисленных в Приложении видов вполне закономерно и не вызывает особых вопросов.

Следует, очевидно, упомянуть и не слишком понятное отсутствие (или редкость) в анализируемой фауне ряда таксонов ногохвосток, обычных для аналогичных тундровых ландшафтов. В первую очередь, это касается видов рода Entomobrya. В имеющемся материале был обнаружен только один неполовозрелый экземпляр. Причем он, как ни странно, принадлежал не к E. nivalis (Linnaeus 1758), обычному для европейских тундр (Babenko et al., 2017), и не к E. bermani Tshelnokov 1977, который встречается восточнее, например на Таймыре (Челноков, 1977), а к бореальному виду E. corticalis, считающемуся типичным кортициколом (Jordana, 2012). Вполне показателен и весьма ограниченный набор видов слитнобрюхих коллембол (Symphypleona). Этот отряд, кроме обычных для тундр гигрофильных Sminthurididae, представлен в наших сборах лишь тремя видами разных семейств: Arrhopalitidae, Katannidae и Bourletiellidae.
Особенности населения. Обследованные ценотические комплексы ногохвосток четко делятся на две группы, сходство между которыми даже на фаунистическом уровне минимально (рис. 6). Первый кластер объединяет основные тундровые местообитания от наиболее влажных и холодных до самых теплых, несмотря на все их различия. Второй – формируют приморские марши, развитые в эстуарии р. Сеяхи (рис. 2в). Фауны этого типа сообществ на низких гипсометрических уровнях обычно сильно обеднены, хотя и включают достаточно богатый набор видов семейства Sminthurididae. Специализированные литоральные формы в наших материалах из приморских стаций отсутствуют, за исключением нескольких особей Folsomia sexoculata (Tullberg 1871), что очевидно связано с сильным опреснением Обской губы в районе исследования. Ценотические комплексы более высоких уровней богаче, как в фаунистическом плане, так и численно (табл. 2).
Рис. 6.
Сходство ценотических фаун коллембол восточного Ямала. Обозначения биотопов см. табл. 2 и 3.

Таблица 2.
Видовой состав и число экземпляров ногохвосток, собранных на маршах в эстуарии р. Сеяха
| Вид, показатель | MI | MII | MIII |
|---|---|---|---|
| Sminthurides malmgreni | 11 | 3 | 8 |
| Folsomia palaearctica | 2 | 5 | |
| Folsomia manolachei | 1 | – | – |
| Folsomia sexoculata | 1 | 2 | 7 |
| Isotomurus stuxbergi | – | 10 | 1 |
| Hymenaphorura sibirica | – | 4 | – |
| Ceratophysella longispina | + | 1 | 4 |
| Hypogastrura assimilis | – | 1 | – |
| Protaphorura subarctica | – | 1 | – |
| Desoria tshernovi | – | – | 427 |
| Desoria violacea | + | – | 393 |
| Parisotoma reducta | – | – | 291 |
| Folsomia quadrioculata | – | – | 280 |
| Desoria fennica | + | – | 170 |
| Friesea mirabilis | + | – | 165 |
| Desoria breviseta | + | – | 65 |
| Folsomia binoculata | – | – | 43 |
| Oligaphorura groenlandica | – | – | 43 |
| Anurida papillosa | – | – | 15 |
| Isotomurus chaos | + | – | 15 |
| Hypogastrura tullbergi | – | + | 13 |
| Hymenaphorura anatolii | – | – | 13 |
| Anurida palustris | – | – | 7 |
| Sphaeridia leutrensis | – | – | 6 |
| Brachystomella parvula | – | + | 4 |
| Sphaeridia pumilis | – | + | 4 |
| Isotoma cf. viridis | + | + | 3 |
| Desoria propinqua | – | – | 3 |
| Ceratophysella czelnokovi | – | – | 1 |
| Ceratophysella scotica | – | – | 1 |
| Pachyotoma miserabilis | + | – | – |
| Friesea quinquespinosa | + | – | – |
| Hypogastrura fjellbergi | + | – | – |
| Hypogastrura viatica | – | + | – |
| Isotoma anglicana | – | + | – |
| Morulina gigantea | – | + | – |
| Sminthurides aquaticus | – | + | – |
| Sminthurides parvulus | – | + | – |
| Sminthurides schoetti | – | + | – |
| Sminthurides cf. signatus | – | + | – |
| Stenacidia violacea | – | + | – |
| Число образцов | 11 | 8 | 8 |
| Обилие, экз./дм2 | 5.5 | 13.5 | 991.0 |
| Число видов* | 4(15) | 8(19) | 25(25) |
Эти две группы ценотических комплексов объединяют приморские разнотравные луга, развитые в виде прерывающейся узкой полосы выше приморских маршей на валу многолетних штормовых выбросов. При кластерном анализе фауны этих лугов образуют единую группу с фаунами наиболее теплых тундровых местообитаний. Однако именно здесь, а также на верхних маршах было зафиксировано около десятка видов (Micranurida sp. aff. balta, Anurida papillosa (Axelson 1902), A. ellipsoides, Hypogastrura viatica (Tullberg 1872), Folsomia ciliata Babenko et Bulavintsev 1993, Desoria fennica (Reuter 1895), D. breviseta Potapov 2017, Isotoma anglicana), не обнаруженных во внутренних местообитаниях данного района. Подавляющее большинство этих видов нельзя назвать уникальными для фаун центрально-палеарктического сектора Арктики, и их полное отсутствие в наших сборах из основных тундровых местообитаний района вполне может быть случайным, но их низкое обилие вне береговых сообществ, по-видимому, бесспорно.
Общая плотность населения ногохвосток в большинстве растительных ассоциаций была достаточно высокой и варьировала от 400 до почти 8000 экз./дм2, в среднем составляя около 2600 экз./дм2, а медианное значение – чуть больше 2000 экз./дм2. Минимальная величина была зарегистрирована в нивальных группировках, максимальная – в зоогенных, а также на приморских луговинах. Средний уровень ценотического или α-разнообразия по данным количественных учетов составлял 29.3 ± 5.6 (здесь и далее Х ± SD), медианное значение 29, а с учетом результатов сифтования, ловушечных и ручных сборов – 33.3 ± 6.2, медиана 32. Дифференцирующее или β-разнообразие, рассчитанное по формуле Уиттекера (β = S/α – 1), составило при этом величину в 2.1 ± 0.5. Уровень доминирования в обследованных группировках ногохвосток, оцененный по величине индекса разнообразия Бергера-Паркера (Мэгарран, 1992), колебался от 1.5 до 6.0, в среднем составляя 4.0 ± 1.4 (медиана 3.9).
Состав массовых видов и их относительное обилие в основных биотопах представлены в табл. 3. Обследованные группировки ногохвосток, как правило, включают от 1 до 3 (эу)доминантов (виды, чье обилие составляет более 12.9% общей численности), богатый набор (2–8) субдоминантов (3.9–12.4%) и множество более редких видов. Из 33 видов, приведенных в табл. 3, 20 были отмечены по крайней мере в половине обследованных местообитаний, два вида (F. quadrioculata и P. pjasinae) обладали 100%-ной встречаемостью и еще четыре (I. cf. viridis, F. manolachei, H. anatolii и F. borealis) не были отмечены только в одном или в двух биотопах. В ловушечных сборах практически по всему ландшафтному профилю безраздельно господствовал один и тот же вид, M. gigantea, который явно избегал лишь заливаемые прибрежные стации. Доля видов с 50%-ной встречаемостью во всех 13 основных биотопах составляла почти 30%. Все это явно свидетельствует о весьма высоком сходстве населения ногохвосток в самых разных местообитаниях обследованного района. Тем не менее результаты тестирования нулевой гипотезы о равномерном распределении видов в обследованном ландшафте показали ее несостоятельность; распределение обилия конкретных видов (с заметной численностью) в разных биотопах отличалось от случайного с высокой долей достоверности. В табл. 4 приведены виды, наиболее “значимые” для разграничения основных обследованных группировок. Показательно, что подавляющее большинство из этих видов, дифференцирующих конкретные группировки, являются массовыми не только в данной группировке, но и присутствуют в достаточно широком спектре разных сообществ. Иными словами, топические группировки ногохвосток в бóльшей степени различаются обилием видов, обычных для района, чем наличием в составе каких-либо специфических форм.
Таблица 3.
Относительное обилие (%) массовых видов ногохвосток в фоновых растительных ассоциациях восточного Ямала
| Вид, показатель | A | B | C | D | E | F | G | H | I | MM | J | K |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Folsomia quadrioculata | 30.6 | 27.0 | + | + | 8.2 | 5.6 | + | + | 6.6 | 16.4 | + | + |
| Protaphorura pjasinae | + | 7.9 | + | 5.0 | 5.2 | + | + | 3.4 | 6.8 | + | 7.3 | 5.5 |
| Isotoma cf. viridis | + | + | + | + | 4.5 | + | – | + | + | + | 4.2 | + |
| Folsomia manolachei | – | + | 18.0 | 21.8 | 11.1 | 40.0 | 19.5 | 33.9 | + | + | 18.1 | 5.9 |
| Hymenaphorura anatolii | – | + | 4.9 | + | 23.0 | + | 19.0 | – | + | 9.6 | + | 7.2 |
| Folsomia borealis | – | + | + | 13.1 | 9.7 | + | – | + | + | 4.4 | + | + |
| Hymenaphorura sibirica | + | + | 6.8 | – | 7.6 | 4.5 | – | + | 25.9 | – | 12.8 | + |
| Folsomia palaearctica | – | + | + | – | – | + | + | + | 21.8 | 12.3 | 12.8 | 64.8 |
| Parisotoma reducta | 6.5 | 25.0 | + | 7.0 | 7.1 | 12.3 | – | – | + | 9.1 | + | – |
| Protaphorura jacutica | – | – | + | + | + | + | 5.1 | + | + | + | – | + |
| Protaphorura subarctica | – | – | + | – | + | 4.6 | – | + | + | 14.9 | + | 3.9 |
| Desoria propinqua | – | 4.9 | – | + | + | + | – | – | + | + | + | + |
| Parisotoma notabilis | + | – | – | + | – | + | – | + | 13.1 | + | + | + |
| Desoria neglecta | – | + | 7.7 | 4.2 | + | + | + | – | – | – | – | + |
| Pseudisotoma sensibilis | – | – | + | 4.2 | – | + | + | + | – | – | + | + |
| Supraphorura furcifera | – | – | 4.1 | 3.9 | + | + | + | – | + | – | – | + |
| Folsomia longidens | – | – | + | 13.3 | 6.8 | + | + | – | – | 3.9 | – | + |
| Desoria tshernovi | 30.6 | 15.7 | + | + | – | + | – | – | + | – | – | – |
| Ceratophysella denticulata | 6.1 | – | – | + | – | – | – | + | + | + | + | – |
| Tetracanthella wahlgreni | – | – | 10.2 | + | – | – | 35.3 | 33.7 | – | + | – | + |
| Brachystomella parvula | + | 4.1 | + | + | – | – | + | – | – | – | – | – |
| Willemia anophthalma | – | – | + | + | – | – | 5.8 | – | + | – | – | + |
| Folsomia binoculata | – | 4.4 | – | – | – | + | + | – | – | + | – | + |
| Folsomia bisetosa | + | – | – | – | – | – | – | + | 6.0 | 11.9 | 6.6 | – |
| Folsomia vasilyevi | – | – | – | + | + | – | – | + | – | – | 4.3 | – |
| Parisotoma ekmani | – | – | 4.9 | + | – | 10.7 | – | – | – | + | – | – |
| Protaphorura tundricola | – | – | – | – | – | + | – | 6.7 | – | – | 16.8 | – |
| Pachyotoma miserabilis | – | – | 15.2 | 10.1 | + | – | – | – | – | – | – | – |
| Oligaphorura groenlandica | – | 4.8 | + | – | – | – | – | – | – | – | – | – |
| Protaphorura stogovi | – | – | – | – | – | – | – | 6.8 | – | – | + | – |
| Protaphorura taimyrica | – | – | – | – | – | – | – | – | – | 4.1 | – | – |
| Oligaphorura ursi | 14.5 | – | – | – | – | – | – | – | – | – | – | – |
| Folsomia rossica | – | – | – | – | – | – | – | – | 7.8 | – | – | – |
| Число проб | 5 | 5 | 9 | 5 | 5 | 9 | 9 | 9 | 5 | 4 | 5 | 5 |
| Общее обилие, экз./дм2 | 408.0 | 2304.8 | 1381.8 | 2131.2 | 2040.8 | 1408.4 | 1133.3 | 1757.3 | 2315.2 | 7733.0 | 2384.8 | 6816.8 |
| Число видов* | 20(29) | 22 | 36(37) | 36(42) | 28 | 31(31) | 28(32) | 24(32) | 27(29) | 37(43) | 30(36) | 33(39) |
Примечания. Нивальные биотопы: A– снежник в распадке: B – нивальный разнотравный луг на северном склоне. Болотные биотопы: C – осоково-ивовое болото с пятнами сфагнумов, D – морошка на сфагновых буграх, E – ерник с морошкой, F – ивняк хвощевый. Зональная ассоциация: G – кустарничково-мохово-лишайниковая полигональная тундра. Наиболее “теплые” биотопы: H – дриадник, I – разнотравный луг на юго-восточном склоне, MM – береговые разнотравные луговины. Зоогенные ассоциации: J – разнотравный лужок с полынью, K – лемминговины в зональной тундре. + – вид присутствует в группировке, но не достигает уровня субдоминирования (3.9%); серым цветом выделены доминанты; * по данным эклекторной выгонки; в скобках – с учетом ручного и ловушечного сборов.
Таблица 4.
Дифференцирующие виды ногохвосток основных растительных сообществ района исследований
| Дриадник | Луг | Снежник | |||
|---|---|---|---|---|---|
| T. wahlgreni | *** (+) | F. palaearctica | *** (+) | D. tshernovi | *** (+) |
| P. stogovi | *** (+) | H. sibirica | *** (+) | O. ursi | *** (+) |
| P. tundricola | *** (+) | P. notabilis | *** (+) | F. quadrioculata | *** (+) |
| F. quinquespinosa | *** (+) | F. rossica | *** (+) | C. denticulata | *** (+) |
| F. manolachei | *** (+) | F. manolachei | *** (–) | S. armatus | *** (+) |
| Зональная тундра | Ивняк | Болото | |||
| H. anatolii | *** (+) | P. ekmani | *** (+) | P. miserabilis | *** (+) |
| T. wahlgreni | *** (+) | P. reducta | *** (+) | D. neglecta | *** (+) |
| W. anophthalma | *** (+) | F. manolachei | *** (+) | O. groenlandica | *** (+) |
| H. sibirica | *** (–) | P. subarctica | *** (+) | I. gorodkovi | *** (+) |
| P. jacutica | *** (+) | H. spei | *** (+) | F. palaearctica | *** (–) |
Примечания. Для каждого местообитания виды расположены в порядке уменьшения их вклада в общую величину критерия χ2. (+) – обилие вида выше, чем предполагалось исходя из нулевой гипотезы об отсутствии различий в относительной численности видов в разных типах местообитаний; (–) – вид более редок в данном типе местообитаний, чем ожидалось. Подчеркнуты названия видов, массовых в данном местообитании; уровень достоверности по критерию χ2: *** p > 0.999; при расчетах использованы средние значения обилия в биотопе в пересчете на одну пробу.
Не менее показательны и результаты многомерного шкалирования, проведенного нами на основе рангового распределения Спирмана (рис. 7): достаточно изолированное положение занимают группировки и наиболее “теплых” местообитаний (дриадник и разнотравный луг), и наиболее “холодных” стаций (снежник). Напротив, точки, соответствующие группировкам из срединной части диапазона обследованных сообществ, включая зональные, образуют единое облако и их диапазоны широко перекрываются.
Рис. 7.
Сходство основных ценотических группировок коллембол восточного Ямала (nonmetric MDS, Rho- index, программа PAST 3.12). Обозначения биотопов см. табл. 3.

ОБСУЖДЕНИЕ
Наиболее характерной чертой арктической биоты по праву считается ее резкая широтная дифференциация (Чернов, 1978, 1980, 1985), и сравнение региональных особенностей арктических фаун (и животного населения) требует достаточно жесткой привязки к той или иной части тундровой зоны. К сожалению, вопрос о подзональном положении района наших исследований на Ямале не имеет пока однозначного ответа. Поселок Сеяха, в окрестностях которого проводилась работа, расположен чуть севернее 70-й широты. На Таймыре примерно по этой широте проходит южная граница тундровой зоны, что связано с более высокими летними температурами среднесибирского сектора Арктики из-за повышенной континентальности климата. Среднеиюльские температуры в пос. Сеяха (~8.5°C) сходны с наблюдаемыми в среднем течении р. Пясины, в частности у устья р. Тареи (см. Чернов, 1989), т.е. в районе, который расположен значительно севернее (~73° с.ш.) и по праву считается олицетворением подзоны типичных тундр Таймыра. Ботаники, работающие на Ямале, выделяют, как правило, две тундровые подзоны: арктических и субарктических (гипоарктических) тундр с делением последних на две части – полосу северных (типичных) и южных (кустарниковых) субарктических тундр (Добринский, 1995; Магомедова и др., 2006; Ребристая, 2013). Несмотря на все терминологические расхождения, эта классификация по смыслу и используемым критериям не сильно отличается от принятого в нашей лаборатории подразделения тундровой зоны на южную, типичную и арктическую подзоны (Чернов, 1978, 1980; Чернов, Матвеева, 1979; Матвеева, 1998). Сложнее обстоит дело с подзональными границами. Так, согласно работе Магомедовой и др. (2006, стр. 150, рис. 3.1 ), пос. Сеяха находится практически на границе между южными и северными субарктическими тундрами (т.е. на южной границе типичных тундр в принятой нами системе), а по мнению Ребристой (2013, стр. 10, рис. 1) – почти в центре северной гипоарктической тундры (= подзона типичных тундр). На циркумполярной карте растительности (CAVM Team, 2003) данный район относится к субзоне D, что соответствует южной полосе типичных тундр в трактовке Матвеевой (1998). Именно такая трактовка принята в нашей работе.
Общее видовое богатство фауны ногохвосток в районе пос. Сеяха несколько уступает фаунам Таймыра, сходным по зональной приуроченности. Например, в среднем течении р. Пясины (Тарея, подзона типичных тундр) при аналогичной интенсивности одноразовых учетов было зарегистрировано 117 видов группы (Бабенко, 2013), а в южных тундрах на той же широте, на которой расположен пос. Сеяха (верховья Нижней Агапы, окрестности оз. Ладаннах), – более 120 (Бабенко, 2005). Общее число видов, зарегистрированных в окрестностях пос. Сеяха, составило всего 104 вида, что ближе по богатству к известным конкретным фаунам Европейского севера: 104 вида – Кольский п-ов, пос. Д. Зеленцы; 103 вида – о-в Долгий, Печорское море (Бабенко, 2012). Кроме того, необходимо учитывать, что почти десять видов конкретной фауны окрестностей пос. Сеяха были обнаружены лишь в приморских стациях. Большинство из этих обитателей прибрежных биотопов – не облигатные “приморские” формы, они вполне могли бы встретиться во внутренних районах, но сам факт их “отсутствия” (или редкости) здесь достаточно показателен. Несколько обеднен состав фауны Ямала и на родовом уровне: зарегистрировано только 39 родов ногохвосток в сравнении с 54 родами, обнаруженными в тундрах Кольского п-ова, 44 родами на о-ве Долгом (Бабенко, 2012), 46 и 43 родами в тундрах Таймыра (в типичной и южной подзонах, соответственно). Несмотря на всю возможную случайность подобных оценок из-за большой доли одновидовых родов в тундровых фаунах, они, похоже, отражают реальность.
Относительную бедность флоры Ямала обычно связывают с ее молодостью и миграционным происхождением (Ребристая, 2013). Наиболее важной эпохой в формировании современного облика полуострова стал верхний голоцен, поскольку лишь тогда произошло его окончательное поднятие (Добринский, 1995). Во время климатического оптимума голоцена (от 9 до 5 тыс. лет назад) на южном Ямале существовали таежные елово-березовые леса, сменившиеся елово-березовой лесотундрой, а затем тундрой (2–3 тыс. лет назад). Возраст растительности современного тундрового типа подчас оценивается величиной всего в 8–9 столетий (Панова, 1990). Очевидно, не менее бурной была история фаун, в том числе фауны ногохвосток. Тем не менее однозначно связать относительно низкое богатство последней лишь с ее молодостью и миграционной природой, наверное, было бы неправильно, поскольку в целом фауна ногохвосток района совсем не ущербна и по структуре вполне соответствует аналогичным фаунам других арктических регионов Палеарктики со сходной зональной приуроченностью. Можно предположить, что определенную роль при этом могут играть чисто экологические факторы, в частности некоторая монотонность растительного покрова и гидротермические условия обычных для Ямала грунтов с легким механическим составом.
Ямал – пограничный регион, в котором должно проявляться взаимное влияние европейских и сибирских арктических биот. Так, например, по мнению Юрцева (Юрцев и др., 1978; Yurtsev, 1994), Ямал – часть Европейско-Западносибирской провинции Арктической флористической области, и по своему составу его флора ближе к флоре востока Большеземельской тундры, чем к таковой Таймыра (Магомедова и др., 2006). Напротив, для животных показано, что в восточно-европейских тундрах “сибирский вклад”, т.е. доля видов с преимущественно сибирским ареалом, может быть весьма значительным (Seebohm, 1880; Штегман, 1938; Еськов, 1988; Городков, 1992; Марусик, 2007) и может даже существенно превышать долю чисто европейских видов в целом ряде ключевых таксонов (Макарова и др., 2019), включая ногохвосток (Babenko et al., 2017). По нашим данным фауна ногохвосток восточного Ямала почти чисто “сибирская”, европейские элементы в ней присутствуют, но их общее число совсем невелико (рис. 4). Тем не менее само наличие подобных видов достаточно показательно.
Несмотря на относительно низкое видовое богатство конкретной фауны, общий уровень ценотического разнообразия ногохвосток в районе исследований существенно не отличался от известного для ряда иных тундровых районов Палеарктики со сходной зональной приуроченностью (рис. 8). Такие обобщенные данные, естественно, очень сильно зависят от числа и характера местообитаний, обследованных в том или ином районе. Так, например, низкий уровень варьирования α-разнообразия на Кольском п-ове, показанный на рис. 8, очевидно представляет собой прямой результат обследования в этом районе всего четырех разных местообитаний. Однако Ямал явно не выпадает из общего диапазона варьирования уровня локального разнообразия и при более строгом подходе (рис. 9), т.е. при учете числа зарегистрированных видов в пересчете на одну пробу в ограниченном наборе однотипных растительных ассоциаций разных районов. Иными словами, уровень локального разнообразия ногохвосток в фоновых сообществах южной половины тундровой зоны – величина довольно стабильная. И группировки ногохвосток Ямала – не исключение из этого правила.
Рис. 8.
Ценотическое богатство группировок ногохвосток в ряде тундровых районов Палеарктики: 1 – Кольский п-ов, 2 – Югорский п-ов (Амдерма), 3 – п-ов Ямал (Сеяха), 4 – п-ов Таймыр (Тарея), 5 – Чукотка (Певек). На этом и рис. 9–11 показаны медианные значения, 25 и 75% квартили (boxes) и полный диапазон варьирования (“усы”).
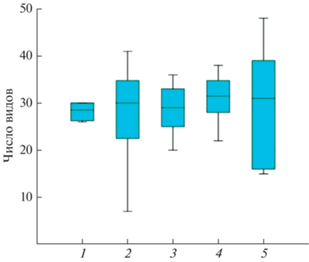
Рис. 9.
Число видов в пробе в четырех аналогичных растительных группировках (а – зональные, б – осоковые болота, в – ивняки и ерники, г – кустарничковые ассоциации) в разных районах Палеарктики. 1 – Кольский п-ов, 2 – Югорский п-ов (Амдерма), 3 – п-ов Ямал (Сеяха), 4 – п-ов Таймыр (Тарея), 5 – Чукотка (Певек).
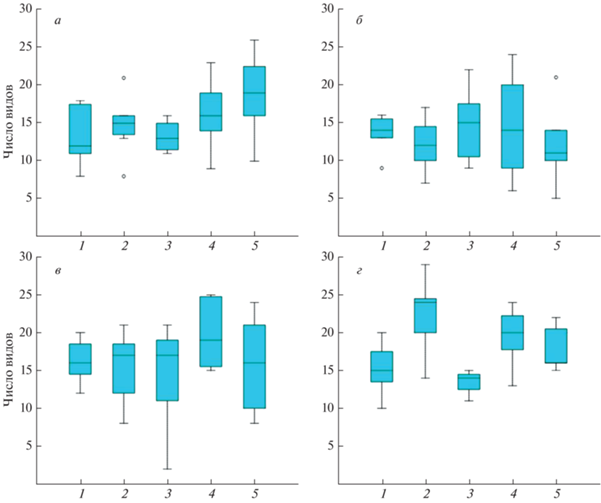
Для большинства обследованных растительных ассоциаций окрестностей пос. Сеяха было характерно достаточно высокое обилие ногохвосток, чьи медианные значения заметно превышали обычные для Таймыра (рис. 10а). Особенно хорошо это заметно если не учитывать выпадающие значения (рис. 10б), которые, как правило, характерны лишь для зоогенных стаций. Тем не менее при сравнении ограниченного числа аналогичных группировок разных районов такой закономерности не наблюдается (рис. 11), и данные для Ямала не выпадают из общего ряда. Обилие – один из наиболее лабильных показателей структуры почвенных группировок (Кузнецова, 2005), поэтому судить, насколько повышенный уровень численности действительно типичен для данного района Ямала, довольно сложно и, вероятно, преждевременно.
Рис. 10.
Плотность населения коллембол на Ямале в сравнении с рядом таймырских районов, сходной зональной приуроченности: а – полный диапазон варьирования, б – без учета выпадающих значений. 1–3 – п-ов Таймыр (Ладаннах, Тарея, Рагозинка), 4 – п-ов Ямал (Сеяха).

Рис. 11.
Плотность населения ногохвосток в четырех аналогичных растительных группировках (зональные, осоковые болота, ивняки и кустарничковые ассоциации) в разных районах Палеарктики: 1 – Кольский п-ов, 2 – Югорский п-ов (Амдерма), 3 – п-ов Ямал (Сеяха), 4 – п-ов Таймыр (Тарея), 5 – Чукотка (Певек).

В завершение хотелось бы подчеркнуть, что, несмотря на некоторую обедненность на видовом уровне по сравнению с более восточными районами Палеарктики, группировки ногохвосток обследованного района Ямала, похоже, не обладают специфическими особенностями, в корне отличающими их от аналогичных комплексов тундровой зоны Евразии. По составу и структуре обследованные группировки вполне соответствуют своему зональному положению в пределах южной полосы типичных тундр. Пограничное положение региона между Европой и Азией проявляется в присутствии в фауне небольшого числа западно-палеарктических видов, не играющих, однако, существенной роли в подавляющем большинстве почвенных группировок.
Список литературы
Андреева Т.Р., Еремин П.К., 1991. Эколого-фаунистический обзор жужелиц (Coleoptera, Carabidae) Южного Ямала // Экологические группировки жужелиц (Coleoptera, Carabidae) в естественных и антропогенных ландшафтах Урала. Свердловск: УрО АН СССР. С. 3–17.
Бабенко А.Б., 2003. Ландшафтная хорология коллембол Таймыра. 1. Биотопическое распределение видов // Зоологический журнал. Т. 82. № 8. С. 937–952.
Бабенко А.Б., 2005. Коллемболы Арктики: структура фауны и особенности хорологии. Автореф. дис. … докт. биол. наук. Москва. 48 с.
Бабенко А.Б., 2012. Ногохвостки (Hexapoda, Collembola) тундровых ландшафтов Кольского полуострова // Зоологический журнал. Т. 91. № 4. С. 411–427.
Бабенко А.Б., 2013. “Коллемболы Западного Таймыра”: сорок лет спустя // Зоологический журнал. Т. 92. № 4. С. 428–444. https://doi.org/10.7868/S0044513413040041
Бабенко А.Б., Потапов М.Б., Стебаева С.К., Чернова Н.М., 1994. Определитель коллембол фауны России и сопредельных стран. Семейство Hypogastruridae. Москва: Наука. 336 с.
Городков К.Г., 1992. Типы ареалов двукрылых (Diptera) Сибири // Э.П. Нарчук (ред.). Систематика, зоогеография и кариология двукрылых насекомых (Insecta: Diptera). Ленинград: ЗИН АН СССР. С. 45–56.
Гришина Л.Г., 1985. Панцирные клещи севера Сибири // Членистоногие Сибири и Дальнего Востока. Новосибирск: Наука. С. 14–24.
Давыдова М.С., 1988. Новые виды гамазовых клещей, связанных со шмелями // Таксономия животных Сибири. Новосибирск: Наука. С. 18–29.
Давыдова М.С., Никольский В.В., 1986. Гамазовые клещи Западной Сибири. Новосибирск: Наука. 125 с.
Добринский Л.Н. (ред.), 1995. Природа Ямала. Екатеринбург: УИФ Наука. 435 с.
Еськов К.Ю., 1988. Пауки (Aranei) Средней Сибири // Материалы по фауне Средней Сибири и прилежащих районов Монголии. Москва: ИЭМЭЖ АН СССР. С. 101–155.
Капрусь И.Я., 2010. Таксономiчна структура i типологiя регiональних фаун ногохвiсток (Collembola) Евразiї // Науковi записки Державного природознавчого музею (Львiв). № 26. С. 39–50.
Коробейников Ю.И., 1980. Материалы по фауне жужелиц Южного Ямала // Фауна и экология насекомых Приобского Севера. Свердловск: УНЦ АН СССР. С. 38–44.
Коробейников Ю.И., 1987. Эколого-фаунистическая характеристика жужелиц южного Ямала // Фауна и экология насекомых Урала. Свердловск: УНЦ АН СССР. С. 33–44.
Ковалев О.Д., Зубрий Н.А., Филиппов Б.Ю., 2017. Экологические факторы и структура населения жужелиц типичных тундр полуострова Ямал // Циркумполярные исследования. Материалы Молодежной науч. школы в рамках междунар. конф. “Живая природа Арктики: сохранение биоразнообразия, оценка состояния экосистем” (3 ноября 2017). Архангельск: ФГБУН ФИЦКИА РАН. С. 22–23.
Кузнецова Н.А., 2005. Организация сообществ почвообитающих коллембол. Москва: Прометей. 244 с.
Линдквист Э.Э., Макарова О.Л., 2011. Два новых циркумполярных вида клещей рода Arctoseius Thor (Parasitiformes, Mesostigmata, Ascidae) // Зоологический журнал. Т. 90. № 8. С. 923–941.
Магомедова М.А., Морозова Л.М., Эктова С.Н., Ребристая О.В., Чернядьева И.В., Потемкин А.Д., Князев М.С., 2006. Полуостров Ямал: растительный покров. Тюмень: Сити-пресс. 360 с.
Макарова О.Л., 2000. К изучению клещей рода Arctoseius (Parasitiformes, Ascidae) Крайнего Севера. 3. Ареалы и экологические предпочтения видов // Зоологический журнал. Т. 79. № 9. С. 1045–1052.
Макарова О.Л., 2009. Мезостигматические клещи (Parasitiformes, Mesostigmata) // Компоненты экосистем и биоразнообразие карстовых территорий Европейского Севера России (на примере заповедника “Пинежский”) (ред. Пучнина Л.В. и др.). Архангельск: Солти. С. 228–239.
Макарова О.Л., 2012. Гамазовые клещи (Parasitiformes, Mesostigmata) Европейской Арктики и их ареалы // Зоологический журнал. Т. 91. № 8. С. 907–927.
Макарова О.Л., Ермилов С.Г., Юртаев А.А., Мансуров Р.И., 2015. Первые сведения о почвенных клещах (Acari) арктического острова Белый (Северный Ямал, Карское море) // Зоологический журнал. Т. 94. № 8. С. 899–904. https://doi.org/10.7868/S0044513415080127
Макарова О.Л., Ануфриев В.В., Бабенко А.Б., Бизин М.С., Глазов П.М., Колесникова А.А., Марусик Ю.М., Татаринов А.Г., 2019. Фауна Восточно-Европейских тундр: вклад “сибирских” видов // Вестник Северо-Восточного научного центра ДВО РАН. № 1. С. 59–71.
Марусик Ю.М., 2007. Пауки (Arachnida: Aranei) азиатской части России: таксономия, фауна, зоогеография. Автореф. дис. … докт. биол. наук. Санкт-Петербург. 36 с.
Марченко И.И., 2012. Почвенные гамазовые клещи (Acari, Mesostigmata) севера Сибири // Евразиатский энтомологический журнал. Т. 11. № 6. С. 517–528.
Матвеева Н.В., 1998. Зональность в растительном покрове Арктики. Санкт-Петербург: БИН РАН. 220 с.
Мэгарран Э., 1992. Экологическое разнообразие и его измерение. Москва: Мир. 184 с.
Панова Н.К., 1990. Новые данные к палеоэкологии и истории растительности Ямала в голоцене // Четвертичный период: методы исследования, стратиграфия и экология. VII Всесоюз. совещание, Таллинн. Т. 1. С. 45–46.
Ребристая О.В., 2013. Флора полуострова Ямал. Современное состояние и история формирования. Санкт-Петербург: изд. СПбГЭТУ “ЛЭТИ”. 312 с.
Рябицев А.В., 1997. Фауна жуков Северного Ямала // Успехи энтомологии на Урале. Екатеринбург. С. 85–88.
Рябицев А.В., 1998. Население и экология жужелиц на севере Ямала. Автореферат диссертации на соискание степени кандидата биологических наук. Лабытнанги, Салехард. 16 с.
Челноков В.Г., 1977. Новые виды ногохвосток рода Entomobrya (Collembola, Entomobryidae) азиатской части СССР // Зоологический журнал. Т. 56. № 7. С. 1030–1037.
Чернов Ю.И., 1978. Структура животного населения Субарктики. Москва: Наука. 167 с.
Чернов Ю.И., 1980. Жизнь тундры. Москва: Мысль. 236 с.
Чернов Ю.И., 1985. Среда и сообщества тундровой зоны // Соколов В.Е. (ред.) Сообщества Крайнего Севера и человек. Москва: Наука. С. 8–22.
Чернов Ю.И., 1989. Тепловые условия и биота Арктики // Экология. № 2. С. 49–57.
Чернов Ю.И., Матвеева Н.В., 1979. Закономерности зонального распределения сообществ на Таймыре // Арктические тундры и полярные пустыни Таймыра. Ленинград: Наука. С. 166–200.
Штегман Б.К., 1938. Основы орнитогеографического деления Палеарктики // Фауна СССР. Птицы. Москва–Ленинград: АН СССР. Т. 1. 157 с.
Юрцев Б.А., Толмачев А.И., Ребристая О.В., 1978. Флористическое ограничение и разделение Арктики // Арктическая флористическая область. Ленинград: Наука. С. 9–104.
Babenko A.B., 1997. The taxonomy and distribution of the genus Anurida (Collembola: Neanuridae) in the northern Palaearctic // European Journal of Entomology. V. 94. P. 511–536.
Babenko A., Fjellberg A., 2006. Collembola septentrionale. A catalogue of springtails of the Arctic regions. Moscow: KMK Scientific Press Ltd. 190 p.
Babenko A.B., Potapov M.B., Taskaeva A.A., 2017. The collembolan fauna of the East European tundra // Russian Entomological Journal. V. 26. № 1. P. 1–30.
CAVM Team, 2003. Circumpolar Arctic Vegetation Map. Scale 1 : 7 500 000. Conservation of Arctic Flora and Fauna (CAFF) Map No. 1. U.S. Fish and Wildlife Service, Anchorage, Alaska.
Esyunin S.L., Efimik V.E., 1996. Catalogue of the spiders (Arachnida, Aranei) of the Urals. Moscow: KMK Scientific Press Ltd. 229 p.
Hammer Ø., Harper D.A.T., Ryan P.D., 2001. PAST – Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. V. 4. № 1. 9 p.
Jordana R., 2012. Synopses on Palaearctic Collembola. V. 7/1. Capbryinae & Entomobryini // Soil Organisms. V. 84. № 1. P. 1–390.
Lindquist L.L., Makarova O.L., 2012. Review of the mite subfamily Arctoseiinae Evans with a key to its genera and description of a new genus and species from Siberia (Parasitiformes, Mesostigmata, Ascidae) // ZooKeys. V. 233. P. 1–20.
Pomorski R.J., 1994. Nowe dane o epilitoralnych gatunkach z rodzaju Anurida Laboulbene, 1865 w Polsce // Przegląd Zoologiczny. V. 38. № 3–4. P. 273–277.
Potapov M.B., Babenko A.B., 2000. Species of the genus Folsomia (Collembola: Isotomidae) of northern Asia // European Journal of Entomology. V. 97. P. 51–74.
Potapov M., Gulgenova A., Babykina M., 2016. Isotomidae (Collembola) of Buryat Republic. III. The genera Vertagopus and Agrenia, with a note on 'Claw index' // Zootaxa. V. 4088. № 1. P. 112–128.
Potapov M., Janion-Scheepers Ch., 2019. Longitudinal invasions of Collembola within the Palearctic: new data on non-indigenous species // Abstracts of the 10th International Seminar on Apterygota. Paris, France. P. 46.
Rusek J., 1970. Zwei neue Collembolen-Arten von den Überschwemmungswiesen in Südmähren // Véstnik Československé Společnosti Zoologické. V. 34. № 4. P. 309–316.
Rusek J., 1977. Collembola // Check list Enumeratio Insectorum Bohemoslovakiae (Dlabola J., Ed.). Acta Faunistica Entomologica Musei Nationalis Pragae. V. 15. Suppl. 4. P. 11–17.
Rusek J., 1995. Collembola // Terrestrial Invertebrates of the Pálava Biosphere Reserve of UNESCO (Rozkosny R. & Vanhara J., Eds). Folia facultatis scientiarum naturalium Universitatis Masarykianae Brunensis. Biologia. V. 92. P. 103–109.
Seebohm H., 1880. Siberia in Europe: a visit to the valley of the Petchora, in North-East Russia, with description of the natural history, migration of birds, etc. London, John Murray, 311 p.
Skarżyński D., Pomorski R.J., 1995. Anurida uniformis Gisin, 1953 nowy dla Polski gatunek skoczogonki (Collembola, Neanuridae) // Przegląd Zoologiczny. V. 39. № 1–2. P. 105–110.
Tanasevitch A.V., Koponen S., Kaarlejarvi E., 2009. Small collection of spiders (Aranei) from the Polar Urals and Yamal Peninsula, Russia // Arthropoda Selecta. V. 17. № 3–4. P. 185–190.
Tanasevitch A.V., Rybalov L.B., 2015. Fauna and distribution of spiders (Aranei) of the arctic tundra in northern Yamal Peninsula, Russia // Arthropoda Selecta. V. 24. № 2. P. 215–230. https://doi.org/10.15298/arthsel.24.2.08
Vasenkova N.V., Kuznetsova N.A., 2022. A multi-scale approach to assessing the structure of Collembola diversity in the boreo-nemoral forests of the Russian Plain // Nature Conservation Research (in press).
Yurtsev B.A., 1994. Floristic division of the Arctic // The Journal of Vegetation Science. V. 5. № 6. P. 765–776.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал