

Зоологический журнал, 2022, T. 101, № 9, стр. 1032-1038
Изменчивость синтаксиса песни у сибирской теньковки (Phylloscopus tristis, Phylloscopidae, Aves)
Я. В. Домбровская a, *, А. С. Опаев a
a Институт проблем экологии и эволюции имени А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: dombrovskayaya@mail.ru
Поступила в редакцию 01.07.2021
После доработки 24.09.2021
Принята к публикации 28.09.2021
- EDN: VTLWGI
- DOI: 10.31857/S0044513422070042
Аннотация
Синтаксис песен/пения подробно описан для многих видов птиц. В большинстве работ исследователей интересовали видовые особенности. Индивидуальная либо ситуативная изменчивости синтаксиса изучены недостаточно – а немногочисленные работы посвящены видам с относительно сложной песней. Неясно, есть ли изменчивость синтаксиса у видов с простой песней и если есть, в чем она состоит. Чтобы выяснить это, мы изучили пение сибирской теньковки (Phylloscopus (collybita) tristis) – вида с простой песней. Всего проанализировали записи 13 самцов. Структуру пения каждого самца мы характеризовали пятью параметрами (медианная длительность песни, медианная длительность пауз, медианное количество элементов в песне, медианное количество типов элементов в песне, размер репертуара элементов). Для изучения организации пения анализировали матрицы переходов между элементами разных типов. По матрицам рассчитывали индекс линейности, индекс постоянства и относительную энтропию первого порядка. В репертуаре каждого самца – 5–8 типов элементов. Разные особи могут использовать репертуар по-разному. Одни исполняют песни с определенной, повторяемой последовательностью элементов. У других конкретные последовательности элементов варьируют от песни к песне. Таким образом, индивидуальная изменчивость в организации пения выражена больше, чем в его структуре. Коэффициенты вариации для параметров, описывающих структуру песен, примерно в 2 раза меньше, чем для параметров организации пения. Открытым остается вопрос о причинах выявленной изменчивости. Мы пока не знаем, является ли она индивидуальной (различия между самцами стабильны), ситуативной либо просто случайной.
Пение воробьиных птиц многофункционально и используется для привлечения брачного партнера, а также регулирования территориальных отношений (Catchpole, Slater, 2008). Пение состоит из стереотипных акустических единиц (например, типов песен или типов звуков), которые обычно чередуются по определенным правилам (Kershenbaum et al., 2014). Поэтому говорят, что пение имеет синтаксис.
Синтаксис песен/пения подробно описан для многих видов птиц (Иваницкий, 2015; Иваницкий, Марова, 2021). Исследователей в большинстве работ интересовали видовые особенности синтаксиса. Выяснилось, в частности, что Марковские цепи низких порядков адекватно описывают организацию пения многих из них. Речь идет о влиянии данного типа песни/звука на следующую за ним песню/звук (Марковская цепь первого порядка), а также на несколько последующих (Марковские цепи более высоких порядков). Оказалось, что одни виды птиц при пении жестко придерживаются определенных правил: например, каждый тип песни четко определяет следующий за ним тип (линейный синтаксис). У других видов выражена свобода комбинирования исходных элементов (комбинаторный синтаксис). При этом индивидуальная либо ситуативная изменчивость таких правил изучена недостаточно (Опаев, 2021). Нам известно немного работ, где такую изменчивость анализировали. У сероголовой очковой пеночки (Phylloscopus tephrocephalus) каждый самец имеет репертуар из 18–48 стереотипных типов песен. Одни особи чередуют их по принципу линейного синтаксиса: самец циклически повторяет все типы песен из своего репертуара в определенной последовательности, а затем начинает сначала. Синтаксис пения других особей комбинаторный: можно выделить несколько групп типов песен, внутри которых возможны изменения порядка исполнения этих песен (Opaev, 2016). У пеночки Клаудии (P. claudiae) выявлена ситуативная изменчивость в организации пения. До проигрывания видовой песни птицы чередовали разные типы песен более свободно, чем во время проигрывания (Opaev et al., 2019). Таким образом, в ответ на трансляцию синтаксис пения становился более линейным.
У многих других видов отсутствуют стереотипные типы песен. Структура их песен может варьировать от случая к случаю. При этом, как и в случае с пением, свобода комбинирования исходных элементов (звуков) в составе песни может различаться у разных видов. Немногие авторы изучали изменчивость такой комбинаторики внутри одного вида. У мексиканской чечевицы (Haemorhous mexicanus) обнаружена ситуативная изменчивость синтаксиса песни. В присутствии конспецифика птицы комбинировали звуки в песне более свободно, чем при спонтанном пении (Ciaburri, Williams, 2019). Мухоловка-белошейка (Ficedula albicollis) также не имеет стереотипных песен. С возрастом упорядоченность пения возрастает – самцы начинают исполнять преимущественно те или иные “любимые” сочетания элементов (Zsebők et al., 2021). В результате их песни в целом более стереотипны, по сравнению с песнями молодых мухоловок. Приведенные материалы позволяют предположить, что внутривидовая изменчивость синтаксиса может играть роль в коммуникации.
Описанные выше 4 вида птиц имеют довольно сложное пение – их репертуары включают десятки типов песен или звуков. Неясно, есть ли изменчивость синтаксиса у видов с простой песней и если есть, в чем она состоит. Чтобы выяснить это, мы изучили изменчивость синтаксиса в песнях сибирской теньковки (Phylloscopus (collybita) tristis) – вида с простой песней.
Пеночки-теньковки (P. collybita s. l.) – комплекс нескольких близких форм со сложной таксономической структурой (del Hoyo, Collar, 2016; Helbig et al., 1996). Вокализация большинства из них изучена в плане уточнения таксономического статуса, а также в рамках исследования гибридизации разных форм (Марова и др., 2018; Marova et al., 2017; Shipilina et al., 2017). В итоге основные особенности структуры пения известны для большинства форм. Например, пение европейской теньковки (P. (c.) collybita) отличается от пения сибирской теньковки тем, что последняя имеет более торопливую песню (выше темп пения, т.е. количество звуков в единицу времени), кроме того, максимальная частота её песен ниже, а частотный диапазон ýже. Помимо этого, звуки сибирской теньковки более разнообразны за счет того, что у европейской теньковки в репертуаре нет звуков с восходящей частотной модуляцией (Шипилина, 2014). Организация пения большинства форм теньковок, за исключением европейской теньковки, изучена недостаточно. У каждого самца европейской теньковки в репертуаре от 9 до 24 типов элементов (звуков). При пении они группируются в 2–8 мотивов (фраз). Была зафиксирована изменчивость пения во времени: спустя дни и тем более годы используемые данной птицей элементы и мотивы могут меняться (Průchová et al., 2017).
МАТЕРИАЛ И МЕТОДЫ
Для исследования мы использовали записи пения 13 самцов сибирской теньковки. 5 из них сделаны В.К. Рябицевым в окрестностях г. Каменск-Уральский (Свердловская обл.) с 8 мая по 8 июня 2020 г., остальные – С.Г. Мещерягиной в г. Асбест (Свердловская обл.) с 5 по 7 мая 2020 г. В записях каждого самца было от 7 до 61 песен (в среднем 29). Записи длились от 51 до 425 с (в среднем 201 с). Суммарная длительность всех изученных записей составила около 75 мин.
Обработка записей велась в программе Syrinx. Мы измерили длительность песен и пауз между ними, а также посчитали количество элементов и их типов в каждой песне. Для каждого самца мы составили каталог репертуара типов элементов. В целом выделение типов элементов у сибирской теньковки достаточно простое. Однако иногда бывают сложности. Например, некоторые элементы птицы могут регулярно не допевать, и такие “недопетые” элементы можно принять за другой тип.
Всем типам элементов мы присвоили номера и представили все песни в виде последовательности цифр. По ним в программе Past3 строили матрицы наблюдаемых переходов и переходных вероятностей. Используя эти данные, мы рассчитывали индексы линейности и постоянства, а также относительную энтропию для каждого самца. Индекс линейности SLIN = размер репертуара/ количество типов переходов между элементами разных типов. Индекс постоянства SCONS = Σ наиболее частых переходов/Σ всех переходов между элементами (Scharff, Nottebohm, 1991). SLIN описывает количество наблюдаемых переходов между элементами разных типов. В линейной последовательности после каждого типа элемента будет всегда следовать другой строго определенный тип (А → В → С → D…). В этом случае SLIN = 1. Если типов переходов больше, то SLIN сокращается, стремясь к нулю. SCONS также изменяется в пределах 0 < SCONS ≤ 1. Этот индекс описывает не то, как именно чередуются звуки, а насколько часто выполняется самый распространенный паттерн. Также мы рассчитали относительную энтропию первого порядка RE1 по матрицам переходных вероятностей: RE1 = E1/E0 (Briefer et al., 2010). Для расчета E1 (энтропия первого порядка) и E0 (энтропия нулевого порядка) использовали формулу Шеннона:
Исходя из этого, E0 описывает гипотетическую последовательность, где все переходы равновероятны, и зависит только от размера репертуара. E1 описывает наблюдаемые переходы. А отношение этих двух параметров (RE1) можно использовать как показатель “свободы выбора” самцом каждого следующего элемента. Например, в случае, если RE1 = 0.70, можно считать, что самец имеет 70% “свободы” выбора (Briefer et al., 2010). Или, по-другому, – последовательность элементов на 70% случайна.
Таким образом, пение каждого самца мы характеризовали восемью параметрами (медианная длительность песни, медианная длительность пауз, медианное количество элементов в песне, медианное количество типов элементов в песне, размер репертуара элементов, индекс линейности, индекс постоянства, относительная энтропия).
Из восьми изученных переменных шесть были распределены нормально (тест Шапиро–Уилка, p > 0.05). Исключения – медианное количество типов элементов в песне (p = 0.02) и индекс линейности (p = 0.01). Поэтому, учитывая также небольшой размер выборки, мы использовали непараметрические методы статистики. Статистическую обработку мы проводили в программе R 3.5.2. (R Core Team, 2016). Для описания взаимосвязей между различными параметрами использовали коэффициент корреляции Спирмена.
Для характеристики изменчивости данного параметра в изученной выборке из 13 самцов использовали коэффициент вариации (выраженное в процентах отношение стандартного отклонения к среднему).
Для визуализации наблюдаемых переходов между разными элементами в песне использовали пакет ‘markovchain’ в среде R (Spedicato et al., 2017).
РЕЗУЛЬТАТЫ
Основные особенности пения сибирской теньковки
В пении сибирской теньковки можно выделить отдельные песни, разделенные паузами. Медианная длительность песни у разных самцов (n = = 13) варьирует от 3.01 до 5.21 с, а паузы между песнями – от 4.07 до 11.43 с. Песня состоит из 15–28 звуков, или элементов (приведен разброс значений медиан разных птиц). Количество типов элементов в одной песне варьирует от 4 до 8. В репертуаре каждого самца имеется 5–8 типов элементов (медиана = 6). Примеры песен трех разных самцов представлены на рис. 1.
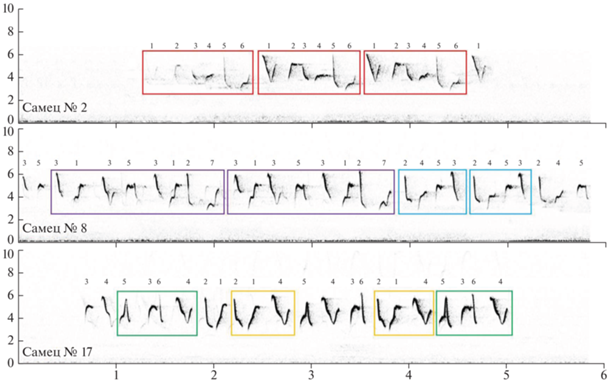
Синтаксис песни: основные особенности и индивидуальная изменчивость
Чаще всего в песне теньковки последовательные элементы разные. Часто в песне выделяются стереотипные сочетания из двух элементов – слоги (например, слог 2 → 1 у самца 17: рис. 1). Нередко слоги и отдельные элементы можно сгруппировать в мотивы (фразы). Под мотивами мы подразумеваем повторяющиеся в составе песни последовательности из нескольких (до восьми) преимущественно разных элементов. В песне можно выделить один или несколько мотивов. При этом большинство элементов относятся к тому или иному мотиву (например, песни самцов № 2 и 8 на рис. 1). Однако бывает, что элементы чередуются более свободно, и тогда ряд элементов сложно отнести к тому или другому мотиву (как у самца № 17 на рис. 1). Особо отметим, что одни и те же элементы могут исполняться в составе разных мотивов (песня самца № 8 на рис. 1). Таким образом, разные самцы могут объединять элементы в песню с разной степенью свободы.
Для характеристики степени свободы группировки разных элементов в составе песни мы использовали ряд индексов. Они заметно различались у разных самцов (табл. 1). При этом индивидуальная изменчивость по индексам выражена существенно больше, чем по структурным характеристикам песни (длительность, количество элементов и их типов, размер репертуара). Это видно по коэффициентам вариации, приведенным в табл. 1.
Таблица 1.
Описательные статистики изученных акустических параметров
| Параметр | Медиана | Минимальное значение | Максимальное значение | Коэффициент вариации, % |
|---|---|---|---|---|
| Медианная длительность, с | ||||
| песни | 3.68 | 3.01 | 5.21 | 19.45 |
| паузы | 7.23 | 4.07 | 11.43 | 30.48 |
| Медианное количество в песне | ||||
| элементов | 20 | 15 | 28 | 19.05 |
| типов элементов | 6 | 4 | 8 | 17.11 |
| Размер репертуара элементов | 6 | 5 | 8 | 13.62 |
| Индекс | ||||
| линейности | 0.37 | 0.19 | 0.86 | 39.34 |
| постоянства | 0.70 | 0.48 | 1 | 16.47 |
| Энтропия | 0.27 | 0.03 | 0.59 | 44.63 |
Различия в упорядоченности пения у четырех разных самцов можно видеть на рис. 2. У самца № 2 энтропия минимальна в нашей выборке. У этой птицы всего один мотив из 6 типов элементов (рис. 1): 1 → 2 → 3 → 4 → 5 → 6. Данная последовательность элементов встречается чаще всего, что видно по частоте переходов на рис. 2. Самое высокое значение энтропии – у самца № 8 (0.59), и соответствующую частоту переходов в его песнях тоже можно наблюдать на рис. 2. У этого самца велико разнообразие мотивов. Он может некоторое время исполнять песню, состоящую из одного и того же мотива, а потом переключиться на другие мотивы. Также на рис. 2 представлены самцы со средними значениями энтропии (самцы № 10 и 12).
Рис. 2.
Наблюдаемые переходы между разными типами элементов (обозначены цифрами в кружках) у четырех самцов сибирской теньковки. Стрелки – переходы, цифры в основании каждой стрелки – частота данного перехода.

Отметим, что индекс постоянства имеет меньший размах изменчивости, чем индекс линейности и относительная энтропия (табл. 1). Кроме того, индекс постоянства довольно высок и обычно выше индекса линейности (табл. 1). Это связано с тем, что у большинства самцов есть предпочитаемые короткие последовательности из двух элементов – слоги. В качестве иллюстрации на рис. 1 (самцы № 2, 10 и 12) красными рамками показаны наиболее частые переходы: в большинстве случаев они как раз соответствуют переходам между элементами в составе слога. Для отдельных самцов, впрочем, слоги менее характерны, и индекс постоянства у таких птиц низок (например, самец № 8 на рис. 1).
Индивидуальная изменчивость индекса линейности и энтропии связана с тем, каким образом разные самцы комбинируют слоги и отдельные элементы при пении – предсказуемо (как самец № 2 на рис. 1) или с существенной степенью свободы (как самец № 8).
Влияние упорядоченности пения на структуру отдельных песен
Результаты корреляционного анализа представлены в табл. 2. Из нее видно, что упорядоченность пения не находит отражение в структуре отдельных песен изученного вида.
Таблица 2.
Коэффициенты корреляции Спирмена между параметрами организации и структуры пения
| Параметр | Индекс линейности | Индекс постоянства | Относительная энтропия |
|---|---|---|---|
| Медианная длительность, с | |||
| песни | –0.29, 0.34 | –0.32, 0.29 | 0.41, 0.17 |
| паузы | 0.38, 0.21 | 0.49, 0.09 | –0.51, 0.08 |
| Медианное количество в песне | |||
| элементов | –0.30, 0.33 | –0.22, 0.47 | 0.23, 0.45 |
| типов элементов | 0.23, 0.46 | –0.02, 0.95 | –0.50, 0.08 |
| Размер репертуара элементов | –0.13, 0.67 | –0.29, 0.34 | –0.03, 0.93 |
ОБСУЖДЕНИЕ
В данной работе мы выяснили, что разные самцы сибирской теньковки могут по-разному использовать свой репертуар. Одни исполняют песни с определенной, повторяемой последовательностью элементов (линейный синтаксис). Другие пользуются репертуаром более свободно. Для пения таких самцов характерны более высокие значения относительной энтропии и низкие значения – индекса линейности. Индекс постоянства же учитывает предпочитаемые слоги в песнях, и его значение всегда выше значения индекса линейности. У самцов с линейным синтаксисом песен относительная энтропия низкая, а индексы линейности и постоянства – высокие.
Важно отметить, что индивидуальная изменчивость в организации пения заметно больше, чем в структуре пения. Так, коэффициенты вариации параметров организации – энтропия и индекс линейности – составили 45% и 39% соответственно. А коэффициенты для параметров, описывающих структуру песни (длительность песни, количество элементов и типов элементов в ней), примерно в 2 раза меньше (17–19%) (табл. 1). Кроме того, параметры организации песни не связаны с параметрами структуры песни. Т.е. организация песни заметно варьирует между самцами. А структура песен остается более или менее постоянной.
Роль структуры песни в коммуникации известна для ряда видов (Опаев, 2021). Обычно в территориальном контексте или при взаимодействии с самкой сложность песни увеличивается. Например, увеличивается длительность песни (как у большой синицы (Parus major): McGregor, Horn, 1992) или количество звуков и типов звуков (элементов) в ней (как у полевого жаворонка (Alauda arvensis): Geberzahn, Aubin, 2014). Индивидуальные различия в структуре песни самцов вне зависимости от контекста также могут быть важны. Например, сложные песни могут вызывать более активный ответ самок (Catchpole et al., 1984; Collins, 1999). В нашем случае различия в структуре песен у разных самцов сибирской теньковки минимальны. Поэтому мы сомневаемся, что они играют роль в коммуникации.
Синтаксис песни варьирует существенно сильнее. Однако неизвестно, насколько предпочитаемый самцом синтаксис песен постоянен. Возможно, это индивидуальная характеристика каждого самца, и со временем организация пения не меняется. Но есть вероятность, что в пении любых самцов имеется определенная изменчивость, и жесткий линейный синтаксис на имеющейся у нас записи через день/месяц/год окажется уже совсем не таким. Упорядоченность пения некоторых самцов менялась на протяжении одной записи. Они пели песни то с одним типом мотива (упорядоченное пение), то с разными (менее упорядоченное пение). Может быть, у других особей такие изменения просто растянуты во времени.
Есть данные по изменчивости песен с течением времени у европейской теньковки. Авторы отмечают, что песни у птиц индивидуальны, и отдельных самцов в конкретный момент можно легко различить по пению. Однако их песня меняется спустя годы, месяцы и даже дни. Т.е. повторная идентификация самцов по одному лишь пению невозможна. Также у европейских теньковок организация песен могла меняться на протяжении одной записи (Průchová et al., 2017) – похожий паттерн мы выявили и у сибирской теньковки.
Также мы не можем исключать ситуативную изменчивость организации песен у сибирской теньковки. К примеру, более упорядоченное пение характерно для одних ситуаций, а менее упорядоченное – для других (как это показано для мексиканской чечевицы – см. выше).
Подводя итог, скажем, что синтаксис песен сибирской теньковки широко варьирует. Однако остается открытым вопрос о причинах такой изменчивости. Мы пока не знаем, является ли она индивидуальной (различия между самцами стабильны), ситуативной либо просто случайной.
Список литературы
Иваницкий В.В., 2015. Песня птиц как микрокосм современной науки: между биоакустикой и лингвистикой // XIV орнитологическая конференция Северной Евразии. II. Доклады. Алматы. С. 418–435.
Иваницкий В.В., Марова И.М., 2021. Синтаксическая организация песни птиц // Зоологический журнал. Т. 100. № 10. С. 1145–1158.
Марова И.М., Ильина И.Ю., Квартальнов П.В., Грабовский В.И., Иваницкий В.В., 2018. От Босфора до Копетдага: биоакустическая дифференциация теньковок в Турции, на Кавказе и в западном Туркменистане // Орнитология: история, традиции, проблемы и перспективы. Материалы Всероссийской конференции, посвященной 120-летию со дня рождения профессора Г.П. Дементьева. М.: Товарищество научных изданий КМК. С. 232–236.
Опаев А.С., 2021. Коммуникативное значение сложного пения воробьиных птиц // Поволжский экологический журнал. № 2.
Шипилина Д.А., 2014. Взаимоотношения восточно-европейской и сибирской теньковок (Phylloscopus collybita abietinus, Ph. (c.) tristis) в зоне симпатрии: морфологические, биоакустические и генетические аспекты. Дис. … канд. биол. наук. М.: Московский государственный университет им. М.В. Ломоносова. 27 с.
Briefer E., Osiejuk T., Rybak F., Aubin T., 2010. Are bird song complexity and song sharing shaped by habitat structure? An information theory and statistical approach // Journal of Theoretical Biology. V. 262. P. 151–164.
Catchpole C.K., Dittami J., Leisler B., 1984. Differential responses to male song repertoires in female songbirds implanted with oestradiol // Nature. V. 312. P. 563–564.
Catchpole C.K., Slater P.J.B., 2008. Bird song: biological themes and variations // 2nd ed. Cambridge: Cambridge University Press. 335 p.
Ciaburri I., Williams H., 2019. Context-dependent variation of house finch song syntax // Animal Behaviour. V. 147. P. 33–42.
Collins S.A., 1999. Is female preference for male repertoires due to sensory bias? // Proceeding of the Royal Society B. V. 266. P. 2309–2314.
del Hoyo J., Collar N.J., 2016. HBW and Bird Life International Illustrated Checklist of the Birds of the World. V. 2. Passerines. Barcelona: Lynx Edicions. 1013 p.
Geberzahn N., Aubin T., 2014. How a songbird with a continuous singing style modulates its song when territorially challenged // Behavioral Ecology and Sociobiology. V. 68. P. 1–12.
Helbig A.J., Martens J., Seibold I., Henning F., Schottler B., Wink M., 1996. Phylogeny and species limits in the palaearctic chiffchaff Phylloscopus collybita complex: Mitochondrial genetic differentiation and bioacoustic evidence // Ibis. V. 138. № 4. P. 650–666.
Kershenbaum A., Blumstein D.T., Roch M.A., Akcay C., Backus G. et al., 2014. Acoustic sequences in non-human animals: a tutorial review and prospectus // Biological Reviews of Cambridge Philosophical Society. V. 91. P. 13–52.
Marova I., Shipilina D., Fedorov V., Alekseev V., Ivanitskii V., 2017. Interaction between common and Siberian chiffchaff in a contact zone // Ornis Fennica. V. 94. № 2. P. 66–81.
McGregor P.K., Horn A.D., 1992. Strophe length and response to playback in great tit // Animal Behaviour. V. 43. P. 667–676.
Opaev A., 2016. Relationships between repertoire size and organization of song bouts in the Grey-crowned Warbler (Seicercus tephrocephalus) // Journal of Ornithology. V. 157. № 4. P. 949–960.
Opaev A., Kolesnikova Yu., Liu M., Kang Z., 2019. Singing of Claudia’s Leaf-warbler (Phylloscopus claudiae) in aggressive contexts: role of song rate, song type diversity and song type transition pattern // Journal of Ornithology. V. 160. P. 297–304.
Průchová A., Jaška P., Linhart P., 2017. Cues to individual identity in songs of songbirds: testing general song characteristics in Chiffchaffs Phylloscopus collybita // Journal of Ornithology. V. 158. № 4. P. 911–924.
R Core Team, 2016. R: A language and environment for statistical computing. Vienna. https://www.R-project.org
Scharff C., Nottebohm F., 1991. A comparative study of the behavioral deficits following lesions of various parts of the zebra finch song system: implications for vocal learning // Journal of Neuroscience. V. 11. № 9. P. 2896–2913.
Shipilina D., Serbyn M., Ivanitskii V., Marova I., Backström N., 2017. Patterns of genetic, phenotypic, and acoustic variation across a chiffchaff (Phylloscopus collybita abietinus/tristis) hybrid zone // Ecology and Evolution. V. 7. № 7. P. 2169–2180.
Spedicato G.A., Kang T.S., Yalamanchi S.B., Thoralf M., Yadav D. et al., 2017. Easy handling discrete time Markov chains. https://cran.r-project.org/web/packages/markovchain
Zsebők S., Herczeg G., Laczi M., Nagy G., Vaskuti E. et al., 2021. Sequential organization of birdsong: relationships with individual quality and fitness // Behavioral Ecology. V. 32. P. 82–93.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал