

Зоологический журнал, 2023, T. 102, № 2, стр. 237-240
Средообразующая роль туруханской пищухи (Ochotona turuchanensis, Lagomorpha, Ochotonidae): влияние на произрастание крапивы двудомной (Urtica dioica) на каменистых осыпях
С. В. Попов a, *, Н. Г. Борисова a, Д. Г. Чимитов a, С. Ю. Ленхобоева a, А. И. Старков a
a ФГБУН Институт общей и экспериментальной биологии СО РАН
670047 Улан-Удэ, ул. Сахьяновой, 6, Россия
* E-mail: zoosvp79@gmail.com
Поступила в редакцию 08.10.2022
После доработки 11.11.2022
Принята к публикации 22.11.2022
- EDN: HPBSDU
- DOI: 10.31857/S0044513423020125
Исходным пунктом настоящего исследования послужило эмпирическое наблюдение, что на каменистых осыпях, заселенных туруханскими пищухами, присутствует, иногда в изобилии, крапива двудомная (Urtica dioica L. (1753)). В то время как в окружающих эти осыпи лесных и степных ассоциациях крапива не обнаруживается. Это наблюдение вызвало вопрос о наличии и природе возможных связей распределения в пространстве пищух и крапивы.
В описаниях экологии крапивы двудомной (Olsen, 1921; Ivins, 1952; Шалпыков и др., 2021) подчеркивается два важных момента: 1) крапива – типичное рудеральное растение, произрастающее в биоценозах с нарушенным растительным покровом; 2) крапива – нитрофильное растение, которое может хорошо произрастать лишь на почвах, содержащих значительное количество азотосодержащих веществ. Эти требования хорошо сочетаются в типичных местообитаниях крапивы – антропогенных фитоценозах, в огородах, на свалках, на выпасах. Однако такие естественные местообитания, как каменистые осыпи, будучи нарушенными, в то же время лишены очевидных источников азотистых соединений (Aho et al., 1998). Каковы источники азота, необходимого для произрастания крапивы?
Пищухи имеют такую особенность питания, как облигатная аутокопрофагия и связанный с нею механизм симбионтной азотфиксации (Формозов и др., 2012). При этом они выделяют во внешнюю среду с фекалиями и мочой значительные объемы азотосодержащих веществ, поскольку именно в их выделениях, в первую очередь в первичном помете – цекотрофах, происходит концентрация азота. Кроме того, часть обильных запасов растительности, которые эти зверьки создают на зиму, не используется, перегнивает и также становится дополнительным источником азота. В отличие от других, обитающих на осыпях, мелких фитофагов (полевок, бурундуков), пищухи оставляют фекалии и мочевые пятна в нескольких точках индивидуального участка, которые часто совпадают с местами хранения запасов. Такие точки могут сохраняться в течение жизни нескольких поколений зверьков и обеспечивать локальную концентрацию азота (Aho et al., 1998).
Роль “норных” видов пищух, в числе других млекопитающих-норников, как “экосистемных инженеров” (ecosystem engineer), т.е. животных, имеющих возможность прямо или косвенно регулировать доступность ресурсов для других видов (Jones et al., 1994), давно привлекала внимание исследователей и хорошо изучена (Кучерук, 1963; Исаков, Панфилов, 1970; Реймес, 1970; Злотин и др., 1975; Wesche et al., 2007; Davidson et al., 2012; Valkó et al., 2021 и др.). Роль пищух-обитателей каменистых осыпей (к которым относится Ochotona turuchanensis) в модификации условий существования окружающих их биоценозов очевидно иная, т.к. эти пищухи не роют нор, а используют в качестве убежищ полости между камнями. Эта роль рассматривается лишь в немногих работах. Травина с соавторами (Травина и др., 1998, 2000) изучали роль пищух в ходе микросукцессий травяного покрова в Туве. Согласно результатам исследований, на ранних стадиях зарастания каменистых россыпей пионерные растения, в первую очередь крапива двудомная, оказываются тесно связанными с местами, где благодаря деятельности алтайских (O. alpina) и в меньшей степени северных (O. hyperborea) пищух появляется и скапливается перегной, состоящий из остатков прошлогодних запасов и помета зверьков. При этом крапива вообще не встречалась вне поселений пищух. Авторы приходят к закономерному выводу, что “Неиспользуемые скопления следов жизнедеятельности петрофильных пищух обеспечивают основу субстрата для сосудистых растений, которые первыми появляются между камнями” (Травина и др., 2000). Кроме того, в своей работе Aho с соавторами (1998) отмечают, что американские пищухи способны изменять количество и доступность питательных веществ для растений: было установлено, что в почве под неиспользованными стожками выше содержание азота, выше скорость его минерализации, а в растениях, произрастающих на стожках, содержание азота выше, чем у растений, произрастающих вне стожков.
В связи с этим мы попытались обнаружить связь между заселенностью осыпей туруханскими пищухами и произрастанием на этих же осыпях двудомной крапивы и, таким образом, выявить возможную роль этого вида пищух как “экосистемного инженера”.
В августе 2022 г. в окрестностях деревень Малое Голоустное и Нижний Кочергат Иркутского р-на Иркутской обл. нами обследованы 30 осыпей. Все осыпи находились на ранней или средней стадии зарастания (были лишены сплошного почвенного и растительного покрова), а также различались по размеру, заселенности пищухами и присутствию крапивы. Каждую осыпь обходили по периметру и дважды пересекали, фиксируя наличие крапивы двудомной и следы жизнедеятельности пищух – запасы, кучки помета и мочевые пятна.
Обилие крапивы и следов жизнедеятельности пищух оценивали в баллах, пользуясь следующими критериями (табл. 1).
Таблица 1.
Критерии для экспертной оценки обилия следов жизнедеятельности пищух и обилия произрастания крапивы двудомной на каменистых осыпях в Иркутской области
| Балл | Следы жизнедеятельности пищух | Крапива двудомная |
|---|---|---|
| 1 | Отсутствуют | Отсутствует |
| 2 | Единичные (1–2 на среднюю осыпь) кучки помета, других следов пребывания нет | Единичные растения по нижнему срезу осыпи |
| 3 | Единичные (1–2 на среднюю осыпь) кучки помета + + единичные запасы | Группы растений по краям осыпи |
| 4 | Многочисленные кучки помета, единичные латрины, единичные запасы | Густые заросли по краям осыпи |
| 5 | Многочисленные следы пребывания (помет, мочевые пятна, запасы) | Заросли по краям и на каменистых склонах осыпи |
Помимо результатов обследования указанных 30 осыпей, в обсуждении приведены результаты разборов запасов пищух на осыпях около пос. Нижний Кочергат (Lenkhoboeva et al., 2021).
Статистическую обработку результатов проводили с использованием пакета Statistica 13.0, избрав для оценки уровня корреляции ранговый критерий Kendall Tau Correlations, поскольку распределение наших данных отличалось от нормального.
По результатам обследования на 7 осыпях (23.3%) крапива не была обнаружена, на 2 осыпях (6.7%) она присутствовала единично или отдельными группами (2–3 балла), а на 21 осыпи встречалась обильно (4–5 баллов).
Следы пребывания пищух отсутствовали на 4 осыпях (13.3%), присутствовали в небольших количествах (2–3 балла) на 4 осыпях (13.3%) и были многочисленны (4–5 баллов) на 22 осыпях (73.4%).
Надо отметить, что в лесу, окружающем осыпи, крапива двудомная нами не обнаружена, а по обочинам проходящих поблизости от осыпей дорог найдены редкие единичные растения.
Между показателями обилия крапивы двудомной и следов жизнедеятельности туруханских пищух обнаружена высоко достоверная корреляция (рис. 1).
Рис. 1.
Обилие крапивы на осыпях и обилие следов пребывания пищух. По горизонтальной оси – баллы оценок обилия, по вертикальной оси – число случаев, линия – линейный тренд коэффициента корреляции.
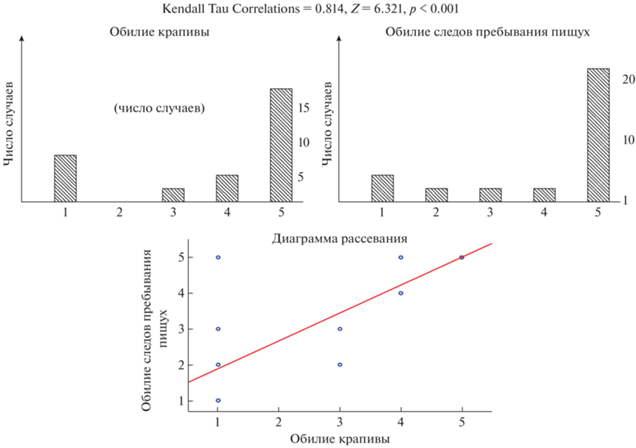
Представляется важным, что на тех четырех осыпях, где следы пребывания пищух полностью отсутствуют (№№ 2, 3, 6, 30), нет и крапивы, в то время как на осыпях № 1 и № 4 отсутствие крапивы сочетается с наличием фекалий и запасов пищух, а на осыпи № 22, при полном отсутствии крапивы, следы пребывания пищух встречаются в изобилии.
При анализе содержания стожков было показано, что во многих случаях пищухи избирательно запасают крапиву, доля которой в стожках значительно больше, чем доля крапивы в растительном покрове осыпи и ее ближайших окрестностей. Доля крапивы в стожках туруханской пищухи более 8%, при этом в окружении крапива встречалась в малых количествах, что отражается в высоком индексе избирательности +0.89 (индексы, говорящие о предпочтении от 0.4 до 1) (Lenkhoboeva et al., 2021).
Само по себе наличие корреляции между обилием крапивы и концентрацией следов жизнедеятельности пищух может свидетельствовать либо о том, что пищухи создают условия, необходимые для произрастания крапивы, либо о том, что обилие крапивы делает осыпь пригодной и благоприятной для заселения пищухами, либо о том, что как пищухи, так и крапива связаны в своем распространении с действием одних и тех же неустановленных факторов. Однако наличие пищух на осыпях, где полностью отсутствует крапива, в сочетании с отсутствием крапивы на осыпях, где нет следов жизнедеятельности пищух, дает нам основания считать первый вариант более предпочтительным по сравнению со вторым и третьим. Действительно, наши результаты позволяют утверждать, что на Приморском хребте наличие на осыпях следов жизнедеятельности туруханских пищух делает такие осыпи пригодными для заселения их крапивой. Эти результаты и сложившиеся на их основе представления о механизмах влияния жизнедеятельности туруханских пищух на растительность осыпей полностью подтверждают результаты, полученные ранее Травиной с соавторами (1998, 2000) при исследовании в сходных условиях двух видов пищух (алтайской и северной), наиболее близких таксономически и по образу жизни к туруханской пищухе.
Места, пригодные для произрастания рудеральных нитрофилов, подобных крапиве двудомной, определяются, видимо, не только активностью пищух, но и воздействием водных (дождями и талыми водами) потоков, которые, сбегая по склонам, обеспечивают перераспределение и накопление образовавшегося гумуса. С этим, как мы считаем, связана концентрация зарослей крапивы по нижним краям осыпей. Для появления достаточного количества плодородной почвы, очевидно, необходимо накопление следов жизнедеятельности нескольких поколений пищух. Этим объясняется отсутствие крапивы на осыпях, по-видимому, недавно заселенных пищухами.
Наши результаты, таким образом, подтверждают роль туруханских пищух как “экосистемных инженеров”. Эти животные изменяют биоту голых каменистых осыпей и влияют на состав растительных сообществ, заселяющих эти осыпи. Наличие аналогичных данных по алтайской и северной пищухам позволяют говорить о том, что эта роль свойственна и другим петрофильным видам семейства Ochotonidae.
В заключение рассмотрим отношение пищух к крапиве. Не будучи критически зависимыми от этого растения, пищухи, тем не менее, активно используют листья крапивы в пищу и избирательно ее запасают. Это позволяет считать, что, создавая своей жизнедеятельностью условия для вселения на осыпь крапивы, пищухи повышают пригодность местообитаний для самих себя, а значит выступают не только как “экосистемные инженеры”, но и как “конструкторы экологических ниш” (Laland et al., 2016).
Список литературы
Злотин Р.И., Исаков Ю.А., Ходашова К.С., 1975. Цели и задачи совещания по изучению роли животных в функционировании экосистем // Роль животных в функционировании экосистем. М.: Наука. С. 3–5.
Исаков Ю.А., Панфилов Д.В., 1970. Основные аспекты средообразующей деятельности животных // Средообразующая деятельность животных. Материалы к совещанию 17–18 декабря 1970 г. М.: Изд-во МГУ. С. 3–9.
Кучерук В.В., 1963. Воздействие травоядных млекопитающих на продуктивность травостоя степи и их значение в образовании органической части степных почв // Биология, биогеоценология и систематика млекопитающих СССР. М.: Изд-во АН СССР. С. 157–193.
Реймерс Н.Ф., 1970. О некоторых особенностях средообразующей роли позвоночных животных // Средообразующая деятельность животных. Материалы к совещанию 17–18 декабря 1970 г. М.: Изд-во МГУ. С. 14–15.
Травина И.В., Дервиз Д.Г., Дмитриев П.П., 1998. Роль пищух (Ochotona, Mammalia) в зарастании каменистых россыпей и каменников в Туве // Доклады Академии Наук. Т. 363. № 2. С. 282–286.
Травина И.В., Дервиз Д.Г., Дмитриев П.П., 2000. Взаимосвязи пищух (Ochotona, Mammalia) Тувы с растительностью // Экология. № 1. С. 41–47.
Формозов Н.А., Кизилова А.К., Пантелеева А.Н., Наумова Е.И., 2012. Азотфиксация – возможная физиологическая основа копрофагии у пищух (Ochotona, Lagomorpha, Mammalia) // Доклады Академии Наук. Т. 443. № 5. С. 646–649.
Шалпыков К.Т., Рогова Н.А., Долотбаков А.К., Кайыркулова А.К., Жакыбалиев Б.Э., 2021. Биологические ресурсы некоторых лекарственных растений в восточном Прииссыккулье Киргизии // Научное обозрение. Биологические науки. № 2. С. 41–47.
Aho K., Huntly N., Moen J., Oksanen T., 1998. Pikas (Ochotona princeps: Lagomorpha) as allogenic engineers in an alpine ecosystem // Oecologia. 114. P. 405–409.
Davidson A.D., Detling J.K., Brown J.H., 2012. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands // Front. Ecol. Environ. 10 (9). P. 477–486.
Ivins J.D., 1952. Concerning the Ecology of Urtica dioica L. // Journal of Ecology. V. 40. № 2. P. 380–382.
Jones C.G., Lawton J.H., Shachak M., 1994. Organisms as Ecosystem Engineers // In: Ecosystem Management. NY: Springer New York.
Laland K., Matthews B., Feldman M., 2016. An introduction to niche construction theory // Evol. Ecol. 30. P. 191–202.
Lenkhoboeva S.Y., Chepinoga V.V., Borisova N.G., Chimitov D.G., Belova V.A., Skornyakova A.M., Nikulin A.A., Nikulina N.A., Ilchenko O.G., 2021. The composition of haypiles of Turuchan pika (Preliminary analysis) // IOP Conf. Ser.: Earth Environ. Sci. 908 012019.
Olsen C., 1921. The ecology of Urtica dioica // Ecol. Vol. 9. № 1. P. 1–18.
Valkó O., Tölgyesi C., Kelemen A., Bátori Z., Gallé R., Rádai Z., Bragina T.M., Bragin Y.A., Deák B., 2021. Steppe Marmot (Marmota bobak) as ecosystem engineer in arid steppes // Journal of Arid Environments. 184. 104244
Wesche K., Nadrowski K., Retzer V., 2007. Habitat engineering under dry conditions: the impact of pikas (Ochotona pallasi) on vegetation and site conditions in southern Mongolian steppes // J. Veg. Sci. 18 (5). P. 665–674.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал