

Зоологический журнал, 2023, T. 102, № 3, стр. 266-283
Переописание Canthocamptus latus Borutzky 1947 (Copepoda, Harpacticoida, Canthocamptidae) из озера Байкал
Т. М. Алексеева a, *, Т. Д. Евстигнеева a, Н. Г. Шевелева a, О. А. Тимошкин a
a Лимнологический институт СО РАН
664033 Иркутск, ул. Улан-Баторская, 3, Россия
* E-mail: atm171@mail.ru
Поступила в редакцию 16.09.2022
После доработки 08.10.2022
Принята к публикации 10.10.2022
- EDN: HQKNEQ
- DOI: 10.31857/S0044513423020034
Аннотация
На основе нового материала приведено иллюстрированное переописание самки и самца Canthocamptus latus Borutzky 1947 – массового эндемичного вида гарпактикоид из литоральной зоны озера Байкал, играющего важную роль в пищевых цепях мелководья. В сравнении с другими эндемичными байкальскими видами рода Canthocamptus Westwood 1836, данный представитель хорошо отличается от остальных видов строением каудальных ветвей и плавательных ног; морфологически вид близок только к Canthocamptus bulbifer Borutzky 1947. Приведены сведения о неотипе (типовые серии были утеряны). Впервые с использованием сканирующего электронного микроскопа была подробно изучена морфология данного вида; приводится описание ротовых конечностей и антеннул, значительно дополнено описание плавательных ног, показано строение генитального поля; представлены данные по морфологической изменчивости.
Байкальские гарпактикоиды – одна из важнейших групп мейозообентоса, отличающаяся необычным таксономическим разнообразием, глубиной эндемизма, а также важной ролью в функционировании бентосных сообществ. В последней таксономической сводке этой группы для Байкала указано 69 видов и еще 9 видов, определенных только до рода (Евстигнеева, Окунева, 2001). Более 80% видов являются эндемиками озера. Помимо них, в коллекции авторов, хранящейся в Лимнологическом институте СО РАН, находилось к тому времени более 30 видов неясного таксономического положения (Евстигнеева, Окунева, 2001). Следовательно, в результате дальнейших исследований с применением современных методов исследования (в частности, СЭМ) видовое богатство байкальских гарпактикоид может значительно возрасти. Сходное мнение недавно высказано Фефиловой с соавторами (2018): имеются предпосылки к описанию большего числа новых таксонов, особенно при изучении глубоководной фауны.
Численность гарпактикоид в прибрежной зоне может достигать нескольких сотен тысяч на метр квадратный, что подчеркивает их исключительную важность при изучении трофических взаимоотношений и круговорота органического вещества в Байкале (Евстигнеева, Окунева, 2001). Гарпактикоиды широко освоили не только биотопы прибрежной зоны (Окунева, 1976; Евстигнеева, 2007); они были найдены также в самых глубоких районах озера (Тахтеев и др., 1993). Несмотря на это, Harpacticoida – одна из наименее изученных групп байкальских ракообразных. Прежде всего это касается сведений об их морфологии на современном светооптическом, а также на электронно-микроскопическом уровнях. Подавляющее большинство видов семейства Canthocamptidae, куда входят почти все байкальские гарпактикоиды, нуждается в современной ревизии. Предыдущими исследователями проведена огромная работа по первоописанию новых таксонов, исследованию их экологии (Боруцкий, 1931, 1947, 1952; Окунева, 1983, 1989). К сожалению, в процессе этих исследований типовой материал авторами, как правило, не выделялся (либо в большинстве своем был утерян), что придает таксономическим исследованиям байкальских гарпактикоид особую важность и сложность. Аспекты экологии и биологии, связанные с байкальскими гарпактикоидами, также изучены мало. Это касается их трофических взаимоотношений, биотопической приуроченности, вертикального и горизонтального распределения в озере. Практически не изучены особенности эндемического видообразования байкальских гарпактикоид, современные схемы филогенетических отношений этой удивительной группы отсутствуют.
Известно, что примерно с 2010 г. мелководная зона Байкала – основной район обитания гарпактикоид – претерпевает сильнейший экологический кризис (Кравцова и др., 2012; Timoshkin et al., 2016). Каким образом негативные экологические процессы отразились на разнообразии гарпактикоид и их роли в донных биоценозах? Ответ на этот важный вопрос также пока отсутствует. Следовательно, актуальность междисциплинарных исследований Harpacticoida озера Байкал не вызывает сомнений. Вначале необходимо проведение таксономической ревизии подавляющего большинства известных и еще не известных науке видов на современном уровне.
В качестве первого объекта в данном направлении исследований нами выбран род Canthocamptus Westwood 1836. До настоящего момента в оз. Байкал он был представлен пятью эндемичными видами, два из которых ранее относились к подроду Baikalocamptus Borutzky 1931 (Боруцкий, 1931). После ревизии рода Canthocamptus Новиковым (Novikov, Sharafutdinova, 2022) данный подрод сведен в синонимы, а вид Canthocamptus gibba Okuneva 1983 оказался младшим синонимом Attheyella (Neomrazekiella) nordenskioldii Lilljeborg 1902, т.к. описания и рисунки видов идентичны. Оставшиеся представители рода значительно различаются между собой, кроме двух морфологически близких видов Canthocamptus latus и Canthocamptus bulbifer Borutzky 1947. Также были высказаны предположения, что C. latus является переходной формой от подрода Baikalocamptus к остальным байкальским представителям рода Canthocamptus (Боруцкий, 1952).
Нами выяснено, что типовой материал, по которому Е.В. Боруцким было выполнено первоописание вида C. latus, утерян.
Цель данной статьи – привести подробное, иллюстрированное описание одного из наиболее многочисленных, широко распространенных видов мелководной зоны озера – Canthocamptus latus.
Материал был собран в Южной котловине оз. Байкал на отрезке Утулик–Мурино в районе БЦБК (Байкальский целлюлозно-бумажный комбинат) 6 июня 2018 г., на каменистом грунте, с глубины 6.5 м. Отбор производили при помощи аквалангистов: камни поднимали со дна и помещали в непроницаемые мешки. На палубе корабля с камней счищали обрастания и промывали отфильтрованной байкальской водой через сачок из мельничного газа с ячеей 60 мкм. Материал фиксировали 40%-ным раствором формалина и затем промывали в лаборатории водопроводной водой. Пробу разбирали под бинокуляром МБС–10. Гарпактикоид препарировали, части монтировали в жидкости Фора–Берлезе. Исследование препаратов проводилось при помощи микроскопов Olympus CX21 FS 1 и Биомед–6. Идентификацию на уровне вида производили при помощи табличных ключей (Боруцкий, 1952; Окунева, 1989). Все рисунки были сделаны с помощью микроскопа Nikon Optiphot–2. Измерения производили по постоянным препаратам и микрофотографиям в программе “Levenhuk Lite”.
Для сканирующей электронной микроскопии образцы подготавливали следующим образом: особей обезвоживали в 96%-ном этаноле в течение суток, затем держали в гексаметилдисилазане в течение 5 мин; далее рачков рассекали в спирте и части переносили на покрытый клеем столик; производили напыление золотом. Фотографии выполнены на сканирующем электронном микроскопе (СЭМ) FEI Company Quanta 200.
Тотальные препараты в жидкости Фора–Берлезе (аббревиатура ПЖФБ – от “препараты в жидкости Фора–Берлезе”) с препарированными особями гарпактикоид имеют нумерацию следующего вида: № H1/1–01–060618, где Н1 – номер особи на предметном стекле, 1 – номер препарата (предметного стекла) из пробы, 01 – номер обработанной пробы, 060618 – дата отбора пробы.
Описание внешнего вида дано по фиксированным особям. Описательная терминология (нумерация каудальных щетинок, обозначение плавательных ножек) по: Huys, Boxshall, 1991.
Весь типовой материал хранится в лаборатории биологии водных беспозвоночных Лимнологического института СО РАН, Иркутск.
Canthocamptus latus Borutzky 1947 (рис. 1–13)
Рис. 1.
Canthocamptus latus, самка неотип: A – внешний вид, дорсально; B – внешний вид, латерально. Масштаб 400 мкм.
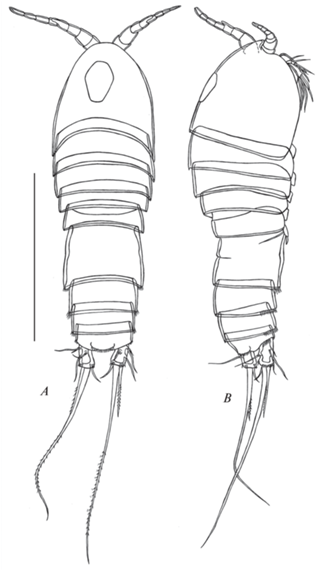
Рис. 2.
Canthocamptus latus, самка неотип: A – абдомен, дорсально; B – абдомен, вентрально; C – часть абдомена, латерально: I–VII – фуркальные щетинки. Масштаб 200 мкм.
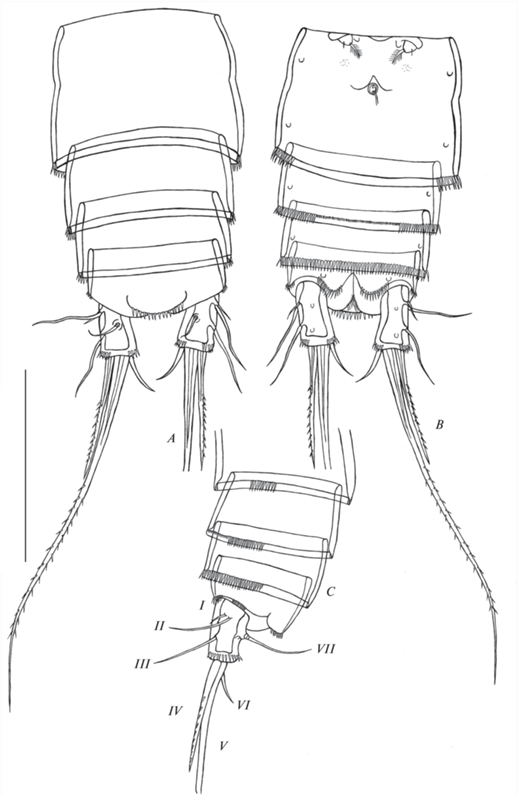
Рис. 3.
Canthocamptus latus, самка: A – внешний вид, дорсально; B – фурка, дорсально; C – фурка, вентрально: стрелкой показана гладкость внутреннего края; D – абдомен, дорсально; E – P6, стрелкой показаны ситовидные поры. Масштаб, мкм: A – 500; B–C – 50; D – 100; E – 20.

Рис. 4.
Canthocamptus latus, самка неотип: A – антеннула и рострум; B – ламбрум, внешняя сторона; C – ламбрум, внутренняя сторона; D – генитальное поле. Масштаб 200 мкм.
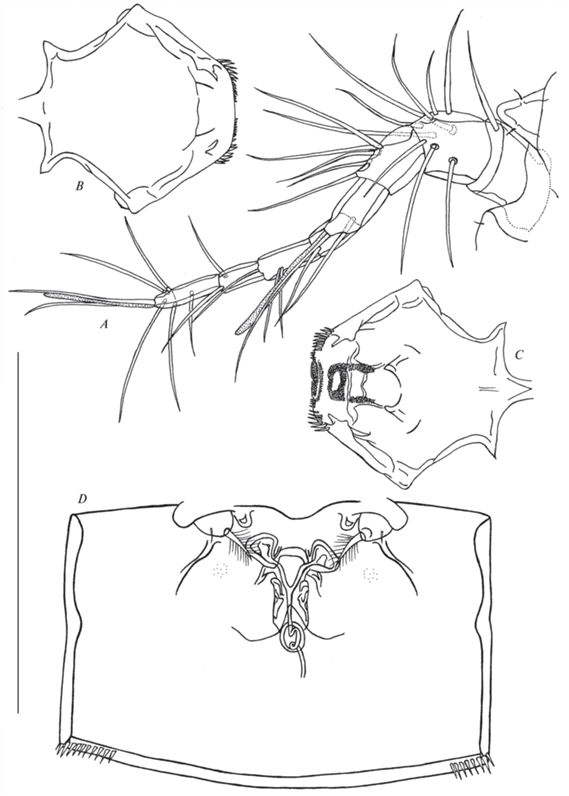
Рис. 5.
Canthocamptus latus, самка: А – антенна; B – антенна, эндоподит; C – мандибулы; D – максиллулы; E – максиллы; F – максиллипеды. Масштаб 200 мкм.
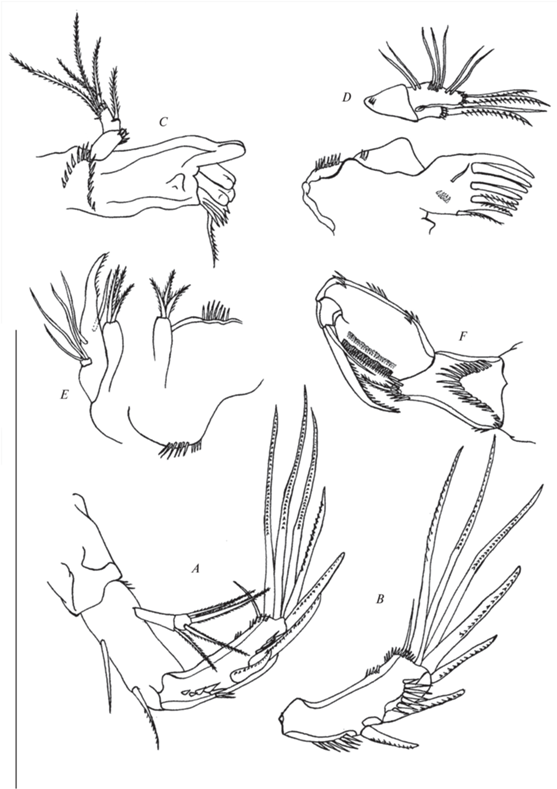
Рис. 6.
Canthocamptus latus, самка: A – ротовые конечности; B – пальпа мандибулы; C – мандибула; D – максиллипеды. Масштаб, мкм: A, D – 50; B – 30; C – 40.

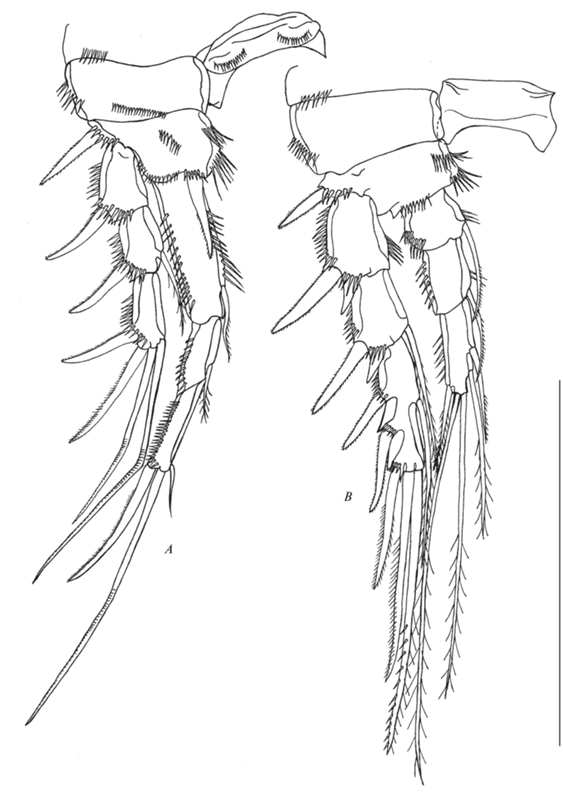
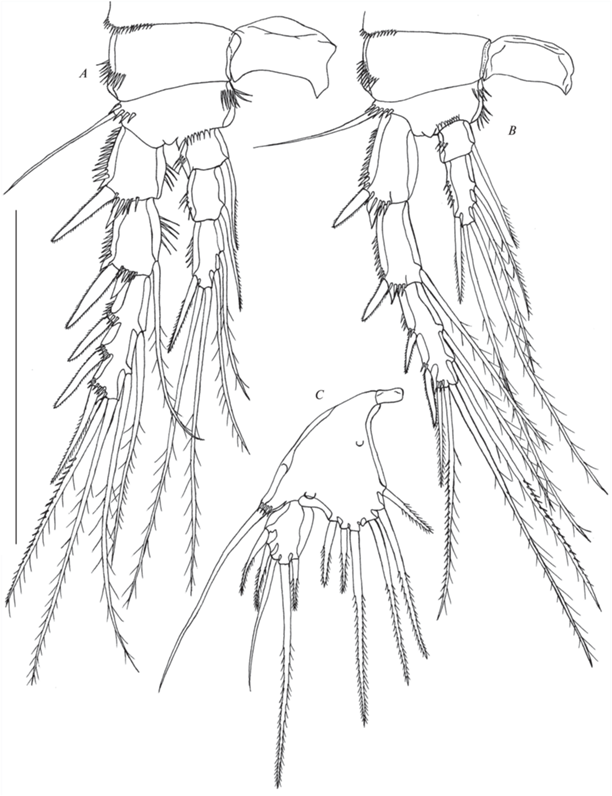
Рис. 9.
Canthocamptus latus, самец: A – внешний вид, дорсально; B – внешний вид, латерально. Масштаб 400 мкм.
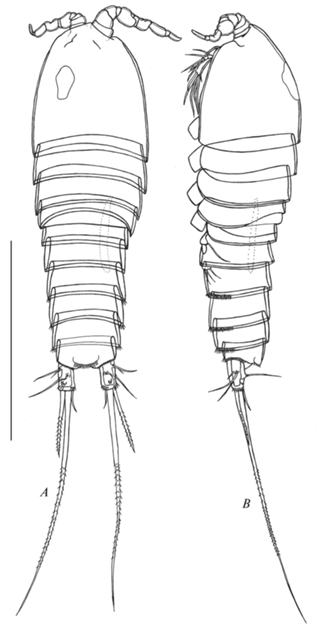
Рис. 10.
Canthocamptus latus, самец: A – абдомен, дорсально; B – абдомен, вентрально; C – часть абдомена, латерально: I–VII – фуркальные щетинки. Масштаб 200 мкм.
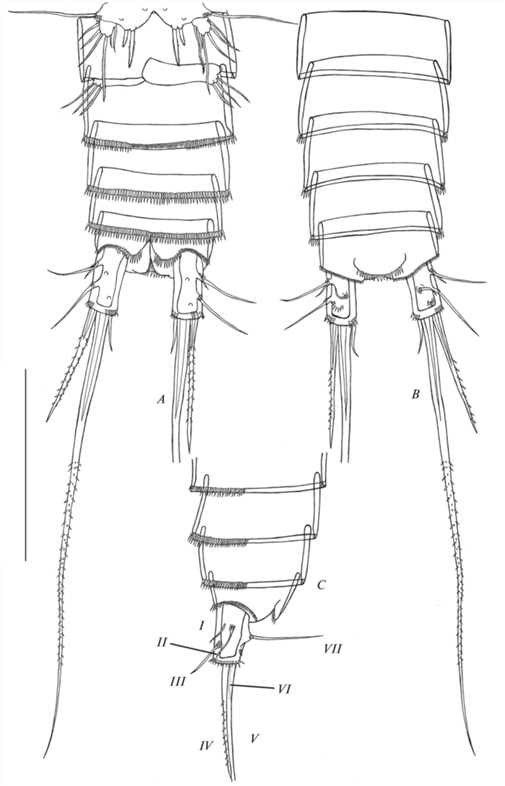
Рис. 11.
Canthocamptus latus, самец: A – внешний вид, дорсально; B – P5: стрелкой показано отсутствие оперения шипов; C – эндоподит P3: стрелкой показана зазубренность апофизы второго сегмента и щетинки на третьем сегменте; D – эндоподит P4: стрелкой показан обычный шип. Масштаб, мкм: A – 400; B–D – 50.

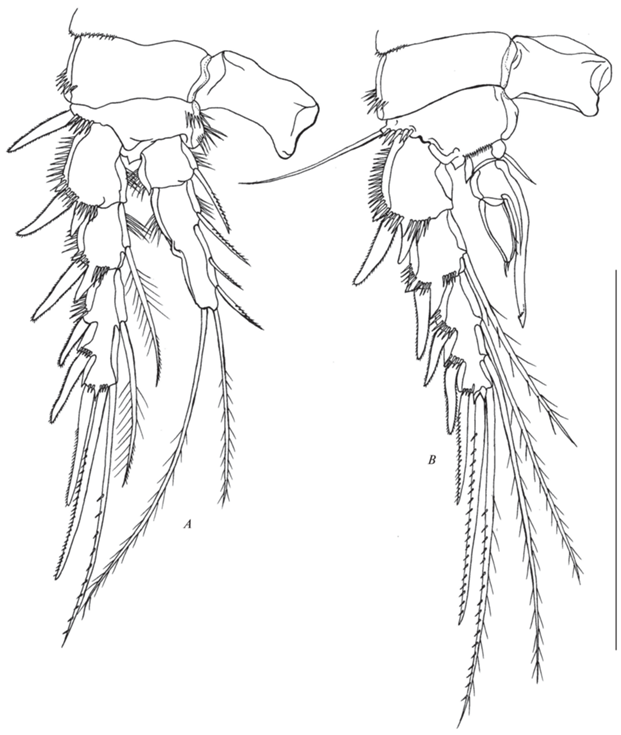
Рис. 13.
Canthocamptus latus, самец: A – антеннула и рострум; B – 3 и 4 сегменты антеннулы; C – P4; D – P5; E – P6. Масштаб 200 мкм.

Материал исследования. Неотип № 1 самка: ПЖФБ № H1/1–01–060618; из озера Байкал (Южная котловина), на отрезке Утулик-Мурино в районе БЦБК (51°31′22.4″ N, 104°11′13.4″ E), глубина 6.5 м, грунт крупные валуны, камни размером 2–5 см, расположенные на песке.
Дополнительный материал: 5 ♀♀: № 1 ♀: ПЖФБ № H2/1–01–060618; № 2, 3: ♀♀ ПЖФБ № H1–2/2–01–060618; № 4, 5 ♀♀: ПЖФБ № H1–2/3–01–060618, все особи собраны там же, где и неотип. 6 ♂♂: № 1, 2 ♂♂: ПЖФБ № H1–2/4–01–060618; № 3, 4 ♂♂: ПЖФБ № H1–2/5–01–060618; № 5, 6: ♂♂ № ПЖФБ H1–2/6–01–060618, собраны там же, где и неотип.
Описание. Самка (рис. 1–8).
Тело (рис. 1A–1B, 3A). Длина от кончика рострума до дистального края каудальных ветвей составляет 804–960 мкм (среднее 845 мкм, n = 6). Цвет фиолетовый. Нукальный орган расположен по центру цефалосомы и имеет форму вытянутого шестиугольника. Интегумент с мелкими волосками по всей поверхности (рис. 3A, 3D). Задний край сомитов тела с простой гиалиновой оборкой.
Рострум (рис. 4A) размером 22–26 мкм (среднее 25 мкм, n = 6), треугольный с одной парой сенсилл.
Генитальный двойной сомит (рис. 4D) шире своей длины, с шипиками на задних боковых углах. Генитальное поле расположено по центру. Копулятивная пора небольшая, ведет к короткому семенному протоку; семенные сосуды маленькие. Рудиментарная Р6 в виде одной перистой щетинки. Ниже каждой рудиментарной ножки есть ситовидные поры (рис. 3E).
Свободные абдоминальные сомиты (рис. 2A–2C, 3D) с мелкими шипиками на вентральной стороне, которые уменьшаются в размере к центру сегментов. Терминальный сегмент с шипиками на вентральной стороне и по бокам, анальная пластинка полукруглая с 15–19 зубчиками (среднее 16, n = 6). Анальное отверстие расположено терминально между каудальными ветвями; анальная бахрома глубоко надрезана, образует лопасти из щетинок.
Каудальные ветви (рис. 2A–2C, 3B–3C) в 2.3 раза больше своей ширины, с семью щетинками. Медиальный край более или менее ровный, латеральный край ровный. Дистальный край ветви окружен не сплошным рядом шипиков, который прерывается на дорсальной, вентральной и внутренней сторонах. Щетинки I–III голые; IV щетинка почти параллельна соседней, вооружена шипиками; V щетинка имеет небольшой изгиб, оперенная; VI щетинка короткая и голая; VII голая, на двойном цоколе.
Антеннулы (рис. 4A) длинные, 8-сегментные. Первый сегмент самый широкий, с одной щетинкой. Второй сегмент широкий, с девятью щетинками, одна из них с цоколем. Третий сегмент с пятью щетинками. Четвертый сегмент с одной свободной щетинкой и сросшимися в основании щетинкой и эстетаском. Пятый сегмент с одной щетинкой. Шестой сегмент с тремя щетинками. Седьмой сегмент с двумя щетинками, одна из них с цоколем. Восьмой сегмент с пятью щетинками с цоколем и акротеком (две щетинки + эстетаск). Формула вооружения: 1 (1), 2 (9), 3 (5), 4 (1 + (1 + э)), 5 (1), 6 (3), 7 (2), 8 (5 + акротек). Щетинки, имеющие цоколи в основании, присутствуют на сегментах 2 (1), 7 (1) и 8 (5).
Антенны (рис. 5A–5B, 6A). Кокса с рядом шипиков. Аллобазис голый; абексоподальные щетинки односторонне перистые. Экзоподит двухсегментный с четырьмя щетинками (с одной мелко оперенной на первом сегменте и тремя мелко оперенными на втором). Свободный эндоподальный сегмент в средней части имеет два мощных шипа и два ряда шипиков (один переходит на фронтальную сторону), расположенных каудально, и две группы мощных шипиков, расположенных фронтально; в апикальной части два шипа, три коленчатые (геникулирующие) щетинки и одна маленькая щетинка.
Ламбрум (рис. 4B–4C) ромбовидный, на дистальных углах шипики, на внутренней стороне на дистальном конце зубчики, расположенные как на рисунке.
Мандибулы (рис. 5C, 6A–6C). Кокса крепкая, гнатобаза с хорошо развитыми округлыми раздвоенными зубами, pars incisiva на фронтальной стороне, на проксимальном конце коксы полукруглый ряд шипиков, на дистальном углу одна щетинка. Пальпа двухсегментная; базис несет ряд шипиков на фронтальной стороне; эндоподит с одной щетинкой на внутренней стороне, четырьмя апикальными щетинками и рядом шипиков на фронтальной стороне.
Максиллулы (рис. 5D, 6A). Прекокса с рядом коротких шипиков на дистальном крае. Прекоксальный артрит с семью шипами (два из которых оперены) и одной оперенной щетинкой на дистальном крае; на фронтальной поверхности располагается голая короткая щетинка, на каудальной – короткий ряд шипиков. Кокса с рядом шипиков на фронтальной стороне; коксальный эндит с одной оперенной и одной голой щетинкой, и группой шипиков на дистальном его конце. Аллобазис с двумя оперенными щетинками на дистальном крае и группами щетинок: две с внешнего края фронтальной стороны, три срединные, две внутренние; также имеются две группы шипиков – у трех срединных щетинок и на дистальном крае.
Максиллы (рис. 5E, 6A). Синкокса с двумя эндитами, которые несут по три оперенные щетинки; на внутреннем и внешнем краях ряды шипиков. Аллобазис с эндитом в виде мощного когтя с шипиками имеет одну щетинку на фронтальной стороне и одну на каудальной. Эндоподит несет три щетинки.
Максиллипеды (рис. 5F, 6A, 6D). Синкокса с оперенной щетинкой на дистальном выступе, и рядами шипиков. Базис немного больше синкоксы, с рядами шипиков на внутреннем и внешнем краях. Эндоподит несет мощный коготь с голой щетинкой у основания.
Р1 (рис. 7A). Интеркоксальный склерит почти в форме полумесяца с двумя рядами шипиков на дистальном крае. Прекокса с рядом шипиков на внешнем крае. Кокса c одним рядом шипиков на наружном крае и одним рядом шипиков на передней стороне. Базис с наружной и внутренней мелкоперистыми щетинками (каждая имеет на базисе ряд шипиков, “покрывающих” основание щетинки), двумя рядами тонких длинных шипиков на внутренней стороне, рядом шипиков на передней поверхности и рядом шипиков на дистальном крае. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль дистального и внешнего краев; проксимальный сегмент с одним внешним шипом; медиальный сегмент с одним внешним шипом и одной внутренней оперенной щетинкой; дистальный сегмент с одним внешним шипом, одним апикальным шипом и двумя апикальными щетинками. Эндоподит трехсегментный, длиннее экзоподита; проксимальный сегмент почти достигает конца медиального сегмента экзоподита, с рядом шипиков на внешнем и внутреннем крае и внутренней щетинкой; медиальный сегмент с рядом шипиков на внешнем крае и одной внутренней щетинкой, тесно соединен с последующим сегментом; дистальный сегмент с рядом шипиков на внешнем крае, частично заходящим на дистальный край, и тремя элементами (одна внутренняя короткая щетинка, одна длинная апикальная щетинка и один апикальный длинный шип).
Р2 (рис. 7B). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом шипиков на внешнем крае. Кокса почти прямоугольная, с одним рядом шипиков на наружном крае. Базис с наружным мелкоперистым шипом и рядом шипиков на наружном крае у основания шипа, у основания эндоподита и двумя рядами тонких длинных шипиков на внутреннем крае. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль внешнего края; проксимальный сегмент с одним внешним шипом, шиповидным выростом на дистально-латеральном углу и коротким рядом тонких длинных шипиков на внутреннем крае; медиальный сегмент с одним внешним шипом, шиповидным выростом на дистально-латеральном углу, оперенной внутренней щетинкой и коротким рядом шипиков на внутреннем крае; дистальный сегмент с шестью элементами (три внешних шипа, две апикальные щетинки и одна внутренняя щетинка). Эндоподит трехсегментный, достигает середины третьего сегмента экзоподита; проксимальный сегмент с рядом шипиков на внешнем крае, коротким рядом шипиков на внутреннем крае и одной мелкооперенной щетинкой там же; медиальный сегмент с рядом шипиков на внешнем крае, коротким рядом шипиков на внутреннем крае и одной мелкооперенной щетинкой там же; дистальный сегмент с пятью элементами (одна внешняя апикальная короткая щетинка, две апикальные и две внутренние щетинки).
Р3 (рис. 8A). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом шипиков на внешнем крае. Кокса прямоугольная, с одним рядом шипиков на наружном крае. Базис с наружной голой щетинкой, рядами шипиков по внешнему и внутреннему краю и у основания эндоподита. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль внешнего края; проксимальный сегмент с одним внешним шипом, шиповидным выростом и коротким рядом шипиков на внутреннем крае; медиальный сегмент с одним внешним шипом, шиповидным выростом, коротким рядом тонких длинных шипиков на внутреннем крае и внутренней оперенной щетинкой; дистальный сегмент с семью элементами (три внешних шипа, две апикальные и две внутренние щетинки). Эндоподит трехсегментный; проксимальный и медиальный сегмент с коротким рядом шипиков на внешнем крае и одной внутренней щетинкой; дистальный сегмент с пятью элементами (один внешний шип, две апикальные и две внутренние щетинки).
Р4 (рис. 8B). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом шипиков на внешнем крае. Кокса почти прямоугольная, с одним рядом шипиков на внешнем крае. Базис с наружной голой щетинкой, одним рядом шипиков на внешнем крае, двумя рядами шипиков на внутреннем крае и рядом шипиков у основания эндоподита. Экзоподит трехсегментный; проксимальный и медиальный сегмент с рядом шипиков вдоль внешнего края, одним внешним шипом и шиповидным выростом на дистальном углу; медиальный сегмент с внутренней оперенной щетинкой; дистальный сегмент с семью элементами (три внешних шипа, две апикальные щетинки и две внутренние щетинки). Эндоподит двухсегментный; проксимальный сегмент с коротким рядом шипиков на внешнем крае и внутренней щетинкой; дистальный сегмент с коротким рядом шипиков на внешнем крае и пятью элементами (один внешний шип, две апикальные и две внутренние щетинки).
Формула вооружения плавательных ног представлена в табл. 1.
Таблица 1.
Формула вооружения плавательных ног (по: Sewell, 1949)
| Ноги | Кокса | Базис | Экзоподит | Эндоподит |
|---|---|---|---|---|
| P1 | 0–0 | I–I | I–0; I–1; II,2,0 | 0–1; 0–1; 1,1,1 |
| P2 | 0–0 | 0–I | I–0; I–1; III,2,1 | 0–1; 0–1; 1,2,2 |
| P3 | 0–0 | 0–1 | I–0; I–1; III,2,2 | 0–1; 0–1; I,2,2 |
| P4 | 0–0 | 0–1 | I–0; I–1; III,2,2 | 0–1; I,2,2 |
Р5 (рис. 8C). Интеркоксальный склерит очень маленький прямоугольный. Базиэндоподит и экзоподит отчетливо разные. Базиэндоподит вооружен шестью мелкооперенными щетинками, из которых первая, пятая и шестая от внутреннего края – самые короткие. Экзоподит немного длиннее внутренней лопасти базиэндоподита и несет 5 щетинок; вторая от внутреннего края – самая длинная, третья лишена оперения.
Самец (рис. 9–13).
Тело (рис. 9A–9B, 11A). Длина от конца рострума до дистального конца каудальных ветвей 752–925 мкм (среднее 847 мкм, n = 6). Форма тела, интегумент как у самки.
Рострум от 25 до 28 мкм (среднее 26 мкм, n = 6), треугольный с парой сенсилл.
Абдоминальные сегменты (рис. 8A–8C) несут на вентральной стороне более мощные ряды шипиков, чем у самки.
Каудальные ветви (рис. 10A–10C) такой же длины и ширины, как у самки. На дорсальной стороне ветвей есть группа шипиков между VII дорсальной щетинкой и дистальным концом. Дистальный край ветви окружен не сплошным рядом шипиков, который прерывается на дорсальной, вентральной и внутренней сторонах. Щетинки I–III голые, в основании имеют 2–4 шипика; IV–VII щетинки как у самки.
Антеннулы (рис. 13A–13B) 10-сегментные, гаплоцерные с геникуляцией между седьмым и восьмым сегментами. Первый сегмент с одной щетинкой и рядом шипиков у основания. Второй сегмент с девятью щетинками, одна из них с цоколем. Третий сегмент с семью щетинками, одна из них с цоколем. Четвертый сегмент – небольшой, с двумя щетинками. Пятый сегмент немного вздутый, с шестью щетинками (из них одна с цоколем и две оперенные) и сросшимися в основании щетинкой и эстетаском. Шестой сегмент короткий с тремя щетинками, одна из них оперенная. Седьмой сегмент с одной щетинкой, зубчатым выростом и углублением в центре, в котором расположены три ряда мелких шипов. Восьмой сегмент с углублением в центре и зубчатым отростком. Девятый сегмент короткий с одной щетинкой. Десятый сегмент с семью щетинками, из них шесть с цоколем, и акротеком (две щетинки + эстетаск). Формула вооружения: 1(1), 2(9), 3(7), 4(2), 5(4 + 2 плюмозы + (1 + э)), 6(2 + 1 плюмоза), 7(1 + зубчатый отросток), 8(зубчатый отросток), 9(1), 10(7 + акротек). Щетинки с цоколями присутствуют на сегментах 2(1), 3(1), 5(1) и 10(6).
Антенны, ламбрум, мандибулы, максиллулы, максиллы, максиллипеды и Р1 как у самки.
Р2 (рис. 12A). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом шипиков на дистальном крае. Кокса почти прямоугольная, с рядом шипиков на внешнем крае. Базис с наружным мелкоперистым шипом, одним рядом шипиков на внешнем крае и двумя рядами шипиков на внутреннем крае. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль внешнего края; проксимальный и медиальный сегменты с наружным шипом и шиповидным выростом и коротким рядом шипиков на внутреннем крае; медиальный сегмент с внутренней оперенной щетинкой; дистальный сегмент с шестью элементами (3 внешних шипа, две апикальные и одна внутренняя щетинки). Эндоподит двухсегментный с неясно отделенным третьим сегментом, оба сегмента с короткими рядами шипиков на внутреннем и внешнем краях; проксимальный сегмент короткий с внутренней щетинкой; медиальный сегмент с круглым хитиновым утолщением, двумя апикальными и тремя внутренними щетинками.
Р3 (рис. 11С, 12B). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом коротких шипиков на дистальном крае. Кокса почти прямоугольная, с рядом шипиков на внешнем крае. Базис с наружной голой щетинкой, одним рядом шипиков на внешнем крае и одним рядом шипиков на передней стороне у основания эндоподита. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль внешнего края; проксимальный и медиальный сегменты с наружным шипом и шиповидным выростом и коротким рядом шипиков на внутреннем крае; медиальный сегмент с внутренней оперенной щетинкой; дистальный сегмент с семью элементами (3 внешних шипа, две апикальные и две внутренние щетинки). Эндоподит трехсегментный; проксимальный сегмент короткий, с внутренней щетинкой; медиальный сегмент несет апофизу (с зазубренной апикальной частью как на рис. 11C) во внутреннем дистальном углу, которая примерно в 3 раза длиннее базальной части сегмента; дистальный сегмент округлый, с двумя короткими щетинками на дистальном конце и двумя щетинками на внутреннем крае (рис. 11C).
Р4 (рис. 11D, 13C). Интеркоксальный склерит прямоугольный голый. Прекокса с рядом коротких шипиков на дистальном крае. Кокса почти прямоугольная, с рядом шипиков на внешнем крае. Базис с наружной голой щетинкой, одним рядом шипиков на внешнем крае, двумя рядами шипиков на внутреннем крае и одним рядом шипиков на фронтальной стороне у основания эндоподита. Экзоподит трехсегментный, каждый сегмент с рядом шипиков вдоль внешнего края; проксимальный и медиальный сегменты с наружным шипом и шиповидным выростом; медиальный сегмент с внутренней оперенной щетинкой; дистальный сегмент с семью элементами (3 внешних шипа, две апикальные и две внутренние щетинки). Эндоподит двухсегментный почти как у самки; проксимальный сегмент короткий, с внутренней оперенной щетинкой; дистальный сегмент с одним внешним шипом (рис. 11D), двумя голыми апикальными щетинками, двумя внутренними короткими редуцированными щетинками (у самки длинные) и коротким рядом тонких шипиков на внешнем крае.
Р5 (рис. 10A, 11B, 13D) медиально срощены, базиэндоподит и экзоподит разделены. Базиэндоподит с наружной щетинкой и коротким рядом наружных шипиков у ее основания. Эндоподальная лопасть меньше, чем у самки, с двумя оперенными шипами разной длины. Экзоподит с парой шипиков на наружном крае и шестью щетинками, из которых самая короткая вторая от внутреннего края.
Р6 (рис. 10A, 13E) срощены с сомитом, каждая из ножек имеет по три щетинки; внутренняя щетинка оперенная, срединная и внешняя – голые. Пластинка покрывает функционирующий гонопор.
Изменчивость. На P5 самки вторая щетинка от наружного края может быть равна первой по длине либо может быть несколько короче нее. Е.В. Боруцкий указывал наличие мелкого шипика на экзоподите P5; все проанализированные нами особи имели развитую щетинку. Первоописание приводит данные, что экзоподит P5 самца вооружен как у самки (т.е. несет 5 щетинок и мелкий шипик) – наши особи имеют 6 щетинок и ряд мелких шипиков на внешнем крае. Также у самцов на P5 наблюдается различие в длине первой внутренней щетинки на экзоподите: у особи № 1 ♂ на одном из сегментов эта щетинка в два раза длиннее и оперена (рис. 10A). Шипы на базиэндоподите P5 самца могут быть голыми (рис. 11B). Покрывающая пластинка на P6 расположена либо справа, либо слева в зависимости от того, какой семенник функционирует.
Дифференциальный диагноз. По морфологическим признакам данный вид сходен с C. bulbifer, главным образом отличается строением фуркальных ветвей, а именно прямой IV щетинкой и ровными основаниями IV и V щетинок. Каудальные ветви менее расширены у дистального края. Такое небольшое количество различий между C. latus и проанализированными нами самками C. bulbifer, а также отсутствие описания самца этого вида, несмотря на его массовость, оставляет вопрос о систематическом положении C. bulbifer открытым. Для выяснения реального таксономического статуса C. bulbifer необходимо использовать методы молекулярной биологии, т.к. этот таксон, вероятно, имеет статус ниже видового.
Географическое распространение. Вид встречается по всему Байкалу, на глубине 1–20 м на песчаных и каменистых грунтах (Окунева, 1989).
Сокращения, используемые в тексте и обозначениях на рисунках согласно Huys, Boxshall, 1991: P1–P6 – плавательные ножки 1–6, э – эстетаск.
Canthocamptus latus – один из наиболее распространенных видов байкальских эндемичных гарпактикоид, чья морфология из-за широкого ареала претерпевает некоторую изменчивость. В статье выполнено подробное описание морфологии самца и самки этого вида с привлечением СЭМ, впервые дано описание ротовых конечностей, расширено описание плавательных ног, антеннул; представлены данные по морфологической изменчивости. Данный вид принадлежит к литофильному комплексу гарпактикоид и массово обитает на глубинах 1–20 м; редко встречается и на бо́льших глубинах – вплоть до 260 м (Окунева, 1989; Евстигнеева, 2007).
Список литературы
Боруцкий Е.В., 1931. Пресноводная фауна. Пресноводные и солоноватоводные Harpacticoida СССР. Выпуск 3. Л.: Печатня. 246 с.
Боруцкий Е.В., 1947. Материалы по фауне Copepoda, Harpacticoida Байкала // Доклады Академии Наук СССР. Т. 58. № 8. С. 1825–1829.
Боруцкий Е.В., 1952. Harpacticoida пресных вод. Фауна СССР. Ракообразные. Т. 3. № 4. М., Л.: Изд-во Академии наук СССР. 426 с.
Евстигнеева Т.Д, Окунева Г.Л., 2001. Гарпактициды (Harpacticoida) // Анн. список фауны озера Байкал и его водосборного бассейна. Новосибирск: Наука, Сибирское отделение. Т. I. Водоемы и водотоки Восточной Сибири и Северной Монголии. Кн. 1. С. 451–467.
Евстигнеева Т.Д., 2007. Экологическая характеристика гарпактицид в литорали озера Байкал // Актуальные вопросы изучения микро-, мейозообентоса и фауны зарослей пресноводных водоемов. Борок, 2–7 октября 2007 г. Нижний Новгород: Вектор ТиС. С. 163–166.
Кравцова Л.С., Ижболдина Л.А., Ханаев И.В., Помазкина Г.В., Домышева В.М. и др., 2012. Нарушение вертикальной зональности зеленых водорослей в прибрежной части залива Лиственничный озера Байкал // Доклады Академии наук. Т. 447. № 2. С. 227–229.
Окунева Г.Л., 1976. Гарпактициды Южного Байкала // Новые материалы по фауне и флоре Байкала. Отв. ред. О.М. Кожова. Иркутский государственный университет. Иркутск: Изд-во Иркутского университета. С. 94–116.
Окунева Г.Л., 1983. Новые виды Copepoda, Harpacticoida в фауне озера Байкал // Зоологический журнал. Т. 62. № 9. С. 1343–1352.
Окунева Г.Л., 1989. Гарпактициды озера Байкал. Иркутск: Изд-во Иркутского университета. 152 с.
Тахтеев В.В., Снимщикова Л.Н., Окунева Г.Л., Тимошкин О.А., Оболкина Л.А., Таничев А.И., 1993. Характеристика донного населения глубинной зоны Байкала // Экология. № 6. С. 60–68.
Фефилова Е.Б., Евстигнеева Т.Д., Майор Т.Ю., 2018. Перспективы исследования гарпактицид (Crustacea: Copepoda: Harpacticoida) Байкала // Пресноводные экосистемы – современные вызовы. Иркутск, 10–14 сентября 2018 г. Иркутск: ООО “Мегапринт”. С. 143.
Huys R., Boxshall G.A., 1991. Copepod Evolution. London: The Ray Society Publ. 468 p.
Novikov A.A., Sharafutdinova D., 2022. Revision of the genus Canthocamptus (Copepoda: Harpacticoida) with a description of a new species from the Lena River Delta (North-eastern Siberia) // European Journal of Taxonomy. V. 826. P. 33–63.
Sewell R.B.S., 1949. The littoral and semi-parasitic Cyclopoida, the Monstrilloida and Notodelphyoida // Scientific Reports of the John Murray Expedition. Publ. British Museum (Natural History). V. 9. № 2. P. 17–199.
Timoshkin O.A., Samsonov D.P., Yamamuro M., Moore M.V., Belykh O.I. et al., 2016. Rapid ecological change in the coastal zone of Lake Baikal (East Siberia): Is the site of the world’s greatest freshwater biodiversity in danger? // Journal of Great Lakes Research. V. 42. P. 487–497.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал