

Зоологический журнал, 2023, T. 102, № 3, стр. 284-290
Ассоциативное обучение и память наездников Trichogramma telengai (Hymenoptera, Trichogrammatidae)
М. А. Федорова a, *, С. Э. Фарисенков a, А. В. Тимохов a, А. А. Полилов a
a Московский государственный университет имени М.В. Ломоносова,
биологический факультет, кафедра энтомологии
119234 Москва, Россия
* E-mail: mariafedorova1997@gmail.com
Поступила в редакцию 13.09.2022
После доработки 10.01.2023
Принята к публикации 14.01.2023
- EDN: BWOYYL
- DOI: 10.31857/S0044513423030054
Аннотация
Вследствие миниатюризации перед насекомыми встает задача сохранить жизненно необходимые функции организма при значительном уменьшении размеров тела. На примере паразитоидов Trichogramma telengai исследовали влияние миниатюризации на функционирование нервной системы насекомых. Несмотря на уменьшение числа нейронов и их объема, у T. telengai было выявлено наличие ассоциативного обучения и сохранение следов памяти вплоть до 6 часов после обучения. Обучение и тестирование памяти микронасекомых происходило на термоарене. Установка работала по принципу водного лабиринта Морриса, что давало возможность сравнивать скорости обучения животных разных таксономических групп. Полученные данные расширяют представление о влиянии миниатюризации на когнитивные способности животных и позволяют определить, какие структурные факторы ограничивают минимальные размеры полноценно функционирующего мозга.
Центральная нервная система и органы чувств миниатюрных перепончатокрылых подробно исследуются в последние годы (Макарова, Полилов, 2013; Makarova et al., 2015, 2021; Fischer et al., 2018, 2019). ЦНС миниатюрных наездников имеет ряд особенностей, характерных для микронасекомых, в частности уменьшенные число нейронов, их объем, а также компактизация нервных ганглиев (Макарова, Полилов, 2013). Характерная для микронасекомых аллометрия отдельных зон мозга подвергается регуляции со стороны естественного отбора. Он помогает предотвратить фатальные изменения в функционировании нервной системы, которые привели бы к снижению когнитивных способностей (Bolstad et al., 2015). План строения головных ганглиев Trichogramma (Hymenoptera, Trichogrammatidae) соответствует общему плану для насекомых (Макарова, Полилов, 2013; Makarova et al., 2021). Мозг T. telengai состоит из примерно 17 000 нейронов со средним диаметром 2.26 ± 0.08 мм (Makarova et al., 2021). Уменьшение их размеров происходит за счет сокращения объема цитоплазмы, в результате которого ядро занимает 50–60% клетки (Макарова, Полилов, 2013). Объем мозга T. evanescens составляет 0.46 нл, и он занимает 7.3% от объема тела (Макарова, Полилов, 2013).
Предшествующие исследования показали способность трихограммы к запоминанию запахов (Huigens et al., 2009; Kruidhof et al., 2012; Farahani et al., 2014) и визуальных стимулов (Keasar et al., 2000). Исследования памяти наездников рода Trichogramma показывают возможности формирования кратковременной (van der Woude et al., 2018), устойчивой к анестезии и долговременной памяти, длительностью вплоть до 24 ч (Huigens et al., 2009; Kruidhof et al., 2012) после обучения на ольфакторных стимулах. При этом уменьшение размеров тела не влияло на способность к сохранению следов памяти – крупные и мелкие особи T. evanescens показывали одинаковые результаты в экспериментах с ольфакторными и визуальными стимулами (van der Woude et al., 2018).
Trichogramma telengai (Sorokina 1987) (Hymenoptera, Trichogrammatidae) – распространенный яйцевой паразит бабочек (Сорокина, 1987). Это вид с телитокическим партеногенезом, самцы встречаются крайне редко (Сорокина, 1987). В дикой природе T. telengai приурочен к лиственным лесам и садам (Теленга, 1959), трофически связан с листовертками Tortricicdae (Сорокина, 1987; Теленга, 1959). В лабораториях разводится, в основном, на яйцах Sitotroga cerealella (Lepidoptera, Gelechiidae).
Данная статья продолжает серию работ авторов по изучению ассоциативного обучения микронасекомых (Федорова и др., 2022). Ранее была показана способность трипсов Thrips tabaci Lindeman 1889 (Thysanoptera, Thripidae) к ассоциативному обучению, а также к сохранению следов памяти до часа после обучения. Термическая арена, использованная в исследовании, является универсальной для любых видов миниатюрных членистоногих и позволяет сравнивать их скорости обучения и длительность сохранения следов памяти.
МАТЕРИАЛ И МЕТОДЫ
Объект исследования – имаго Trichogramma telengai Sorokina 1987 (Hymenoptera, Trichogrammatidae) (длина тела около 460 мкм). Трихограмма, задействованная в эксперименте, культивируется в Лаборатории экспериментальной энтомологии С.Я. Резника, Зоологический институт РАН, Санкт-Петербург, Россия, на яйцах зерновой моли Sitotroga cerealella. Выведение трихограммы из яиц хозяина происходило при температуре 25 ± 1°C и 12-часовом световом дне. В экспериментах использовали имаго не старше трех дней. Эксперименты проводили в августе и сентябре 2020 г. Между тестами на наличие следов памяти насекомых помещали в отдельные пробирки с ниткой, смоченной в медовом растворе.
Экспериментальная установка, на которой проводили исследования ассоциативного обучения и памяти, была подробно описана нами в предыдущей работе, посвященной когнитивным способностям трипсов (Федорова и др., 2022) (рис. 1). Установка представляет собой термическую арену со светодиодным экраном на периферии. Диаметр арены составляет 28 мм, что является достаточно большой площадью для микронасекомого и делает случайный поиск неэффективной стратегией. Арена была нагрета до температуры, вызывающей реакцию избегания, – 37 ± 0.5°C. В каждый момент времени во время обучения на арене присутствовал участок с комфортной температурой, которая соответствовала температуре содержания насекомых и составляла 25 ± 1°C. Комфортный участок мог находиться в одном из четырех возможных положений, и его положение меняли после каждой попытки обучения. Рисунок установленного вокруг арены экрана составлял целевой паттерн – вертикальная черная полоса, обрамленная двумя вертикальными белыми полосами, и горизонтальные полосы, обеспечивающие равномерное освещение. Положение целевого паттерна в экспериментах с тестовой группой совпадало с комфортным участком, за счет чего насекомые обучались находить его, ориентируясь по рисунку на экране.
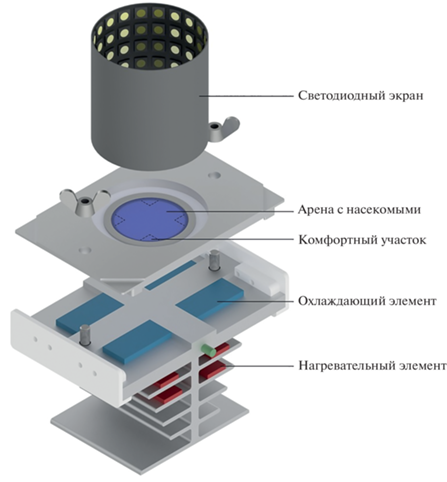
Протокол эксперимента подробно был описан в предыдущей работе (Федорова и др., 2022). Каждую особь помещали на арену, где она проходила тестирование изначальных предпочтений (ТП), за которым следовали 10 циклов обучения. Далее проводили тестирование на наличие следов памяти: сразу после обучения (Т0), а также через 2 ч (Т2) и через 6 ч (Т6) после обучения.
Эксперимент был проведен на 180 особях (по 90 в тестовой и контрольной группах), из которых в итоговом анализе использованы 80 (по 40 в тестовой и контрольной группах). Особи, которые не могли найти холодный участок в течение 5 мин или не смогли выполнить 10 циклов поиска, были выбракованы.
Регистрация и анализ данных
Передвижение насекомых было зафиксировано на цифровую камеру Moticam 3. С помощью программы Tracker 5.0.5 (https://physlets.org/tracker) были получены координаты траекторий движения. По координатам были рассчитаны длина треков при поиске комфортного участка, а также время, проведенное в каждом из четырех секторов.
Поведение насекомых во время сеансов обучения и во время тестов анализировали раздельно. В первом случае анализировали длину пути, пройденную от одного комфортного участка до другого. Во втором – распределение времени, проведенного насекомым в каждом из секторов, доля времени в целевом секторе (четверть арены, соответствующая целевому паттерну экрана) и индекс обучения (отношение разности между временем в целевом секторе и временем в противоположном секторе к общему времени в этих двух секторах) в тестах ТП – Т6. Целевой сектор в каждом тесте является соседним от того, в котором насекомое находится изначально. Индекс обучения показывает, правильно ли насекомым было выбрано направление поиска, поскольку целевой и противоположный от него сектора находятся на одинаковом расстоянии от изначального, но в противоположных направлениях.
Статистический анализ был выполнен в STATISTICA 12 (t-test и ANOVA). При проведении множественных попарных сравнений применялась поправка Бонферрони.
РЕЗУЛЬТАТЫ
Тесты памяти
Достоверные различия наблюдаются при сравнении доли времени, проведенного насекомыми из тестовой группы в целевом секторе в ТП и Т0 (t-test, df = 63, F = 1.317, t = –4.392, p < 0.001), в ТП и Т2 (t-test, df = 59, F = 1.380, t = –3.595, p < 0.001) и ТП и Т6 (t-test, df = 68, F = 2.754, t = –2.049, p = 0.044). (рис. 2)
Рис. 2.
Доля (%) времени, проведенного T. telengai в целевом секторе, в каждом из четырех тестов (M ± SE). t-test: * 0.01 ≤ p < 0.05, ** p < 0.01.

В тестовой группе наблюдаются достоверные различия при сравнении доли времени пребывания в целевом секторе с аналогичным показателем в остальных секторах в ТП и Т0 (ANOVA, df = 60, F = 5, p < 0.001), ТП и Т2 (ANOVA, df = 56, F = 4, p < 0.001) и ТП и Т6 (ANOVA, df = 65, F = 1, p = 0.027) (рис. 2). Доля времени нахождения в целевом секторе достоверно различалось у тестовой и контрольной групп во всех тестах памяти – Т0 (t-test, df = 71, F = 2.801, t = 5.433, p < 0.001), Т2 (t-test, df = 60, F = 1.032, t = 3.206, p = 0.002) и Т6 (t-test, df = 73, F = 1.950, t = 2.189, p = 0.032) (рис. 3). По индексу обучения достоверные различия в тестовой группе наблюдаются при сравнении ТП и Т0 (t-test, df = 63, F = 2.177, t = –3.767, p < 0.001), ТП и Т2 (t-test, df = 59, F = 1.401, t = –2.255, p = 0.028) и ТП и Т6 (t-test, df = 68, F = 1.406, t = –1.998, p = 0.049) (рис. 3).
Рис. 3.
Индекс обучения (отношение разности между временем в целевом секторе и временем в противоположном секторе к общему времени в этих двух секторах) T. telengai в каждом из четырех тестов (M ± SE). t-test: * 0.01 ≤ p < 0.05, ** p < 0.01.

Индекс обучения при сравнении тестовой и контрольной групп достоверно различался во всех тестах памяти – Т0 (t-test, df = 71, F = 1.507, t = 4.875, p < 0.001), Т2 (t-test, df = 60, F = 1.670, t = 2.403, p = = 0.019) и Т6 (t-test, df = 73, F = 1.255, t = 2.647, p = = 0.010) (рис. 3).
Динамика обучения
Для исследования динамики обучения сравнивали длину пути до комфортного участка в тестовой и контрольной группах в каждой из десяти попыток. Начиная с 5-й попытки длина пути особей из тестовой группы становится достоверно короче (df = 76, t = 2.107, F = 4.982, p = 0.038), эта тенденция сохраняется и в последующих попытках (6: df = 70, t = 3.661, F = 4.952, p < 0,001; 7: df = 70, t = 2.199, F = 1.544, p = 0.031; 8: df = 70, t = 2.700, F = 4.281, p = 0.009; 9: df = 67, t = 2.098, F = 1.950, p = 0.03; 10: df = 70, t = 2.443, F = 2.364, p = 0.017) (рис. 4). Средний путь особей из тестовой группы в попытках 5–10 достоверно отличается от среднего пути тех же особей в попытках 1–2 (df = 282, t = 3.636, F = 2.261, p < 0.001) и от среднего пути особей из контрольной группы в попытках 5–10 (df = 433, t = –6.227, F = 3.204, p < 0.001) (рис. 4).

ОБСУЖДЕНИЕ
В процессе миниатюризации насекомые не лишаются способностей к ассоциативному обучению и памяти. Напротив, перепончатокрылые паразитоиды, значительная часть которых является представителями миниатюрного размерного класса, соперничают с социальными насекомыми за статус насекомых с самым сложным поведением (Farris, Schulmeister, 2011). Способности к ассоциативному обучению паразитоидов подробно исследуются в связи с их широким использованием в качестве модельных объектов нейробиологии, а также из-за их частого применения в качестве биологических агентов контроля численности вредителей сельского хозяйства. В экспериментах с ольфакторнымы стимулами паразитоиды Leptopilina boulardi (Barbotin et al. 1979) (Hymenoptera, Figitidae) показывают кратковременную память, до 2 ч после одной попытки обучения, и долговременную, от 24 ч после многократных попыток (Kaiser et al., 2003). После 1 ч обучения паразитоиды Nasonia vitripennis (Walker 1836) (Hymenoptera, Pteromalidae) были способны сохранять память до 96 ч (Schurmann et al., 2009), а Lariophagus distinguendus (Förster 1841) (Hymenoptera, Pteromalidae) – до 144 ч (Müller et al., 2006).
На разных видах паразитоидов можно наблюдать, как различия в способностях к обучению и памяти эволюционировали в соответствии с экологией и образом жизни насекомого (Hoedjes et al., 2011).
Сравнительные исследования когнитивных способностей насекомых разных видов были ограничены из-за сложности подбора универсального стимула, который имел бы одинаковую силу для разных объектов (Kruidhof et al., 2012). Термоарена на основе водного лабиринта Морриса позволяет сравнивать скорости обучения животных из разных групп. Возникновение ассоциативной связи между стимулами у T. telengai наблюдается после пяти повторностей обучения, что сравнимо с результатами обучения у других насекомых и млекопитающих (табл. 1). Сходные скорости обучения в водном лабиринте Морриса и его аналогах у животных со столь разными объемами мозга можно объяснить тем, что более крупный мозг, обеспечивая бóльшую чувствительность и детализацию, не обязательно имеет преимущества при решении конкретных задач (Chittka, Niven, 2009).
Таблица 1.
Число попыток, после которых формируется достоверно выявляемое обучение у разных животных в водном лабиринте Морриса и его аналогах
| Объект | Число попыток, необходимое для обучения |
Экспериментальная установка |
Ссылка |
|---|---|---|---|
| Rattus norvegicus (Rodentia, Muridae) |
3 | Водный лабиринт Морриса | Муровец, Александров, 2020 |
| Drosophila melanogaster (Diptera, Drosophilidae) |
4 | Термоарена | Ofstad et al., 2011 |
| Gryllus bimaculatus (Orthoptera, Gryllidae) |
4 | Термоарена | Wessnitzer et al., 2008 |
| Trichogramma telengai (Hymenoptera, Trichogrammatidae) |
5 | Термоарена для микронасекомых | Данная статья |
| Thrips tabaci (Thysanoptera, Thripidae) |
8 | Термоарена для микронасекомых | Федорова и др., 2022 |
У насекомых могут формироваться разные типы памяти, различающиеся по длительности и устойчивости. Кратковременная память, или чувствительная к анестезии, консолидируется в две формы памяти – устойчивую к анестезии и долговременную. Две эти формы формируются независимо (Tully et al., 1994), однако при усилении одной из них другая оказывается менее производительной (Lagasse et al., 2012). У паразитоидов рода Cotesia (Hymenoptera, Braconidae) консолидация памяти происходила через 1–2 ч после одной попытки обучения, причем тип консолидированной памяти зависел от условного стимула (van den Berg et al., 2011; Kruidhof et al., 2012). Эксперименты с T. evanescens (Huigens et al., 2009) показывают, что через час после одной попытки обучения память еще не успевает консолидироваться и остается кратковременной, а через 24 ч наблюдается уже долговременная память (Huigens et al., 2009). Мы наблюдаем сохранение следов памяти у T. telengai сразу после эксперимента, через 2 и через 6 ч после него, поэтому можем предположить наличие как кратковременной памяти, так и ее консолидированных форм.
Список литературы
Макарова А.А., Полилов А.А., 2013. Особенности строения и ультраструктуры головного мозга насекомых, связанные с миниатюризацией. 2. Мельчайшие перепончатокрылые (Hymenoptera, Mymaridae, Trichogrammatidae) // Зоологический журнал. Т. 92. № 6. С. 695–706.
Муровец В.О., Александров А.А., 2020. Особенности влияния мемантина на обучение крыс в водном тесте Морриса // Журнал высшей нервной деятельности. Т. 70. № 1. С. 50–61.
Сорокина А.П., 1987. Биологическое и морфологическое обоснование видовой самостятельности Trichogramma telengai sp. n. (Hymenoptera: Trichogrammatidae) // Энтомологическое обозрение. Т. 1. С. 32–46.
Теленга Н.А., 1959. Таксономическая и экологическая характеристика видов рода Trichogramma (Hymenoptera: Trichogrammatidae) // Научные труды украинского института защиты растений. Т. 8. С. 124–130.
Федорова М.А., Фарисенков C.Э., Тимохов А.В., Полилов А.А., 2022. Ассоциативное обучение и память трипсов // Зоологический журнал. Т. 101. № 8. С. 1–12.
Berg van den M., Duivenvoorde L., Wang G., Tribuhl S., Bukovinszky T., Vet L.E.M., Dicke M., Smid H.M., 2011. Natural variation in learning and memory dynamics studied by artificial selection on learning rate in parasitic wasps. // Animal Behaviour. Vol. 81. № 1. P. 325–333.
Bolstad G.H., Cassara J.A., Márquez E., Hansen T.F., van der Linde K., Houle D., Pélabon C., 2015. Complex constraints on allometry revealed by artificial selection on the wing of Drosophila melanogaster // Proceedings of the National Academy of Sciences of the United States of America. Vol. 112. № 43. P. 13284–13289.
Chittka L., Niven J., 2009. Are bigger brains better? // Current Biology. Vol. 19. № 21. P. 995–1008.
Farahani H.K., Ashouri A., Goldansaz S.H., Shapiro M.S., Golshani A., Abrun P., 2014. Associative learning and memory duration of Trichogramma brassicae // Progress in Biological Sciences. Vol. 4. № 1. P. 87–96.
Farris S.M., Schulmeister S., 2011. Parasitoidism, not sociality, is associated with the evolution of elaborate mushroom bodies in the brains of hymenopteran insects // Proceedings of the Royal Society B: Biological Sciences. Vol. 278. № 1707. P. 940–951.
Fischer S., Lu Z., Meinertzhagen I.A., 2018. From two to three dimensions: the importance of the third dimension for evaluating the limits to neuronal miniaturization in insects // Journal of Comparative Neurology. Vol. 526. P. 653–662.
Fischer S., Lu Z., Meinertzhagen I.A., 2019. Three-dimensional ultrastructural organization of the ommatidium of the minute parasitoid wasp Trichogramma evanescens // Arthropod Structure and Development. Vol. 48. P. 35–48.
Hoedjes K.M., Kruidhof H.M., Huigens M.E., Dicke M., Vet L.E.M., Smid H.M., 2011. Natural variation in learning rate and memory dynamics in parasitoid wasps: opportunities for converging ecology and neuroscience // Proceedings of the Royal Society B: Biological Sciences. Vol. 278. P. 889–897.
Huigens M.E., Pashalidou F.G., Qian M., Bukovinszky T., Smid H.M., van Loon J.J.A., Dicke M., Fatouros N.E., 2009. Hitch-hiking parasitic wasp learns to exploit butterfly antiaphrodisiac // PNAS. Vol. 106. № 3. P. 820–825.
Kaiser L., Pérez-Maluf R., Sandoz J.C., Pham-Delècue M.H., 2003. Dynamics of odour learning in Leptopilina boulardi, a hymenopterous parasitoid // Animal Behaviour. Vol. 66. № 6. P. 1077–1084.
Keasar T., Ney-Nifle M., Mangel M., 2000. Evidence for learning of visual host-associated cues in the parasitoid wasp Trichogramma thalense // Israel Journal of Zoology. Vol. 46. № 3. P. 243–247.
Kruidhof H.M., Pashalidou F.G., Fatouros N.E., Figueroa I.A., Vet L.E.M., Smid H.M., Huigens M.E., 2012. Reward value determines memory consolidation in parasitic wasps // PLoS ONE. Vol. 7. № 8. P. 1–10.
Lagasse F., Moreno C., Preat T., Mery F., 2012. Functional and evolutionary trade-offs co-occur between two consolidated memory phases in Drosophila melanogaster // Proceedings of the Royal Society B: Biological Sciences. Vol. 279. № 1744. P. 4015–4023.
Makarova A.A., Veko E.N., Polilov A.A., 2021. Metamorphosis of the central nervous system of Trichogramma telengai (Hymenoptera: Trichogrammatidae) // Arthropod Structure and Development. Vol. 60. № 101005.
Makarova A.A., Polilov A., Fischer S., 2015. Comparative morphological analysis of compound eye miniaturization in minute hymenoptera // Arthropod Structure and Development. Vol. 44. № 1. P. 21–32.
Müller C., Collatz J., Wieland R., Steidle J.L.M., 2006. Associative learning and memory duration in the parasitic wasp Lariophagus distinguendus // Animal Biology. Vol. 56. № 2. P. 221–232.
Ofstad T.A., Zuker C.S., Reiser M.B., 2011. Visual place learning in Drosophila melanogaster // Nature. Vol. 474. № 7350. P. 204–207.
Schurmann D., Collatz J., Hagenbucher S., Ruther J., Steidle J.L.M., 2009. Olfactory host finding, intermediate memory and its potential ecological adaptation in Nasonia vitripennis // Naturwissenschaften. Vol. 96. № 3. P. 383–391.
Tully T., Preat T., Boynton S.C., del Vecchio M., 1994. Genetic dissection of consolidated memory in Drosophila // Cell. Vol. 79. № 1. P. 35–47.
Wessnitzer J., Mangan M., Webb B., 2008. Place memory in crickets // Proceedings of the Royal Society B: Biological Sciences. Vol. 275. № 1637. P. 915–921.
van der Woude E., Huigens M.E., Smid H.M., 2018. Differential effects of brain size on memory performance in parasitic wasps // Animal Behaviour. Vol. 141. P. 57–66.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал