

Биология моря, 2020, T. 46, № 1, стр. 24-31
Остеологические особенности Ipnops agassizii Garman, 1899 (Aulopiformes: Ipnopidae) с батиальных и ультраабиссальных глубин Австрало-Новозеландского района с замечаниями о биогеографическом значении данных находок
В. П. Природина 1, *, А. В. Неелов 1
1 Зоологический институт (ЗИН) РАН
199034 Санкт-Петербург, Россия
* E-mail: antarct@zin.ru
Поступила в редакцию 01.11.2018
После доработки 17.05.2019
Принята к публикации 30.05.2019
Аннотация
На основании материала, полученного с батиальных и ультраабиссальных глубин (3020–5410 м), дано остеологическое описание глазного отдела головы и грудного пояса (по ализариновым препаратам) Ipnops agassizii Garman, 1899 (семейство Ipnopidae); приведено детальное описание осевого скелета и проанализированы особенности скелета плавников (по рентгенограммам). Описаны морфологические преобразования в скелете, возникшие как результат глубоководного образа жизни. Показан широкий диапазон функционально-морфологической специализации, которая носит адаптивный характер, направленный на приспособление к жизни на больших глубинах в контакте с дном. Плавниковые лучи непарных плавников и поддерживающие их элементы (соответственно интерневралии и интергемалии) имеют усиленные костные головки. Брюшные плавники находятся на вертикали окончания лучей грудного плавника несколько впереди от уровня середины туловища. Спинной плавник расположен далеко впереди от анального. Хвостовой стебель одинаково уплощен как латерально, так и дорсовентрально.
Рыбы семейства Ipnopidae, как и представители некоторых других семейств, входящих согласно современной классификации в отряд Aulopiformes (Nelson, 2006), относятся к морским донным глубоководным видам, принадлежащим к батиально-абиссальной и ультраабиссальной (хадобентальной) ихтиофауне (Парин, 1988). Семейство включает 5 родов: Bathypterois Günther, 1878 (20 видов); Ipnops Günther, 1878 (3 вида); Bathymicrops Hjort, Koefoed, 1912 (5 видов); Bathytyphlops Norman, 1937 (4 вида) и Discoverichthys Merrett, Nielsen, 1966 (1 вид). Относящиеся к роду Ipnops виды локально распространены в Атлантическом (I. murrayi Günther, 1878), Индийском и Тихом океанах (I. agassizii Garman, 1899 и I. meadi Nielsen, 1966) (Gray, 1956; Nielsen, 1966; Sulak, 1977; Щербачев, 1980; Merrett, Nielsen, 1987; Johnson et al., 1996; McEachran, Fechhelm, 1998; Nelson, 2006; Franco et al., 2009, и др.). В ходе экспедиции НИС “Дмитрий Менделеев” (1975–1976 гг.) особи Ipnops agassizii впервые были обнаружены в приантарктических водах Австрало-Новозеландского района на ультраабиссальных глубинах 5400–5410 м. По итогам экспедиции опубликовано описание выловленных экземпляров I. agassizii и дана характеристика основных видовых признаков этого вида и вида I.meadi Nielsen, 1966 из коллекции Института океанологии РАН; проведено сравнение всех трех видов рода Ipnops, приведены сведения о их распространении (Щербачев, 1980).
Цель настоящей работы – на примере I. agassizii выявить морфологические преобразования в скелете, возникшие как результат глубоководного образа жизни рыб семейства Ipnopidae, уделяя основное внимание строению головной “костной пластинки”, скелета плечевого пояса и грудных плавников.
МАТЕРИАЛ И МЕТОДИКА
В работе использован материал из западной части индо-тихоокеанской провинции (море Фиджи), Юго-Восточной Австралии (Большой Австралийский залив, плато Натуралистов) и желоба Маккуори-Хьорт. Рыбы были выловлены донным тралом “Галатея” в 16-м рейсе НИС “Дмитрий Менделеев” в 1975–1976 гг. с участием одного из авторов настоящей статьи (В.П. Природиной).
Проанализированы следующие материалы, хранящиеся в коллекциях ЗИН РАН: Ipnops agassizii − № 45332, 1 экз. SL 74.6 мм, ст. 1263, южная часть моря Фиджи, 30°36′ ю.ш., 169°22′ в.д., гл. 3305–3470 м, 04.01.1976; колл. А.П. Андрияшев, В.П. Природина. – № 45333, 1 экз. SL 118.0 мм, ст. 1292, южная часть Маккуорийского желоба, 54°59′ ю.ш., 159°21′ в.д., гл. 5400–5410 м, температура 0.9°C, темно-коричневый алевритовый ил c обломками минералов, 25.01.1976; колл. А.П. Андрияшев, В.П. Природина. – № 45334, 2 экз. SL 62.0 и 117.0 мм, ст. 1365, Большой Австралийский залив, 34°26′ ю.ш., 128°12′ в.д., гл. 3870–3880 м, красная глубоководная глина, 26.02.1976; колл. А.П. Андрияшев, В.П. Природина. – № 45335, 5 экз., SL 55.5−132.3 мм, ст. 1254, южная часть моря Фиджи, 30°00′ ю.ш., 169°02′ в.д., гл. 3020–3060 м, 01.01.1976; колл. А.П. Андрияшев, В.П. Природина.
В плавниках подсчитывали число лучей и отмечали, на вертикали какого позвонка начинается первый луч плавника. Остеологию грудного плавника и его пояса, а также головной “костной пластинки” изучали на окрашенных ализариновых препаратах по методике, разработанной в лаборатории ихтиологии ЗИН РАН (Якубовский, 1970; Андрияшев и др., 1977).
В работе приняты терминология и порядок изложения материалов, использованные при описании внешней морфологии видов рода Ipnops (см.: Nielsen, 1966), примененные при изучении рыб других таксономических групп (Сазонов, 1986; Неелов, 1979; Природина, Цветков, 2016) и дополненные авторами данной статьи.
В статье приняты следующие сокращения: SL – стандартная длина рыбы; D – спинной плавник; А – анальный плавник; Р – грудной плавник; V – брюшной плавник; С – хвостовой плавник, хвостовой стебель (расстояние от вертикали основания последнего луча анального плавника до конца чешуйного покрова); aD – антедорсальное расстояние; аА – антеанальное расстояние; rad – радиалии; pu – предуростилярный позвонок; vert – общее число позвонков, включая уростилярный; U – уростиль; inn – интерневралии, inhm – интергемалии. Сокращения, принятые при описании грудного пояса, осевого скелета и хвостового плавника, приведены в подписях к рисункам.
РЕЗУЛЬТАТЫ
Ipnops agassizii Garman, 1899 – ипнопс Агассица
Синонимия. Ipnops agassizii Garman, 1899: 259, Pl. H. (figs. 2–2a) (Галапагосские острова, 2°34′ N, 92°06′ S, гл. 1360 fathoms). – Theisen, 1965: 17 (Ipnops, висцеральный скелет, головная пластина Ipnops murrayi). – Nielsen, 1966: 56–57, fig. 4 (лектотип MCZ 28501, выделен Нильсеном, без описания, рентген, состояние гонад и размножение, Индо-Пацифика, донный образ жизни, глубина 1392–2820 м). – Щербачев, 1980: 47–67 (описание вида, изменчивость, сравнительные данные, распространение). – Okiyama, 1981: 247–253 (личинка SL 13.9 мм в эпипелагиали и молодь 49 мм; ализариновое окрашивание). – Okiyama, 1984: 20 (поимка молоди ранней стадии онтогенеза, онтогенетическая вертикальная миграция). – Okiyama, 1986: 443–445, fig. 1, SL 55 мм (батипелагиаль, первая находка молодого экземпляра в толще воды на глубине 1810–1990 м, метаморфоз). – Sulak, 1986: 260–281 (род Ipnops в семействе Chlorophthalmidae, меристические признаки I. agassizii + I. murrayi).
Рис. 1.
Скелетные элементы Ipnops agassizii (ЗИН № 45333): а – рентгенограмма головы и передней части туловища до начала брюшных плавников; б – левая часть лобной костной пластинки и левая ветвь проходящего в ней надглазничного сенсорного канала (ализариновый препарат). pm1 – передняя пара пор предкрышечно-нижнечелюстного канала; so1, so3 и so6 – передняя, средняя и задняя поры левой ветви надглазничного канала.
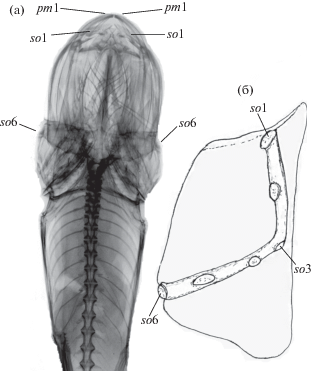
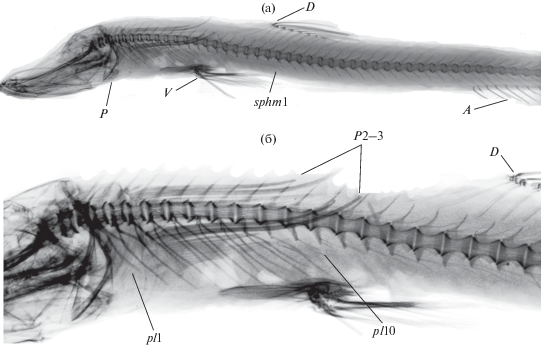
Рис. 2.
Рентгенограмма передней части тела (а) Ipnops agassizii и расположение непарных плавников, плевральных ребер, грудного и брюшного плавников и 1–2-го лучей спинного плавника относительно друг друга (б). D – начало спинного плавника (лучи 1–2); P – грудной плавник; A − анальный плавник; P2–3 – удлиненные второй и третий лучи грудного плавника; V – брюшной плавник; pl1–10 – плевральные ребра; sphm1 – первый гемальный отросток.
Ipnops pristibrachium Fowler, 1943: 53–91 (биология, Филиппинский архипелаг и прилегающие регионы).
Диагноз. Vert 21–23 + 37 = 58–60 (61); лучей: D 9, А 14–16, P 13–15, V 8−7 (редко), C 21–22; жаберных тычинок 22–23, жаберные отверстия очень большие, снизу доходят до симфизиса нижних челюстей. У рыб стандартной длиной (SL) от 40 мм морфологически сформированные глаза отсутствуют, у пелагических личинок имеются.
В предкрышечно-нижнечелюстном канале 7 крупных пор (pm), открывающихся вниз; рm1 парные, так как каждый канал открывается самостоятельной порой, широко расставленные.
Грудных радиалий 4, слабо окостеневшие, радиальные вырезки имеются во второй и третьей радиалиях; верхний и нижний лучи Р редуцированы, основные лучи грудного и брюшного плавников разветвляются только на самом конце. Первый луч D находится на вертикали 18–19-го позвонка; первый луч А расположен на вертикали 37–38-го позвонка. Основание Р на уровне 3-го позвонка; основание V на уровне 12-го позвонка. Гипуральных пластинок в хвостовом отделе 5 (H1–H5); число краевых лучей С 1–2/2.
В процентах стандартной длины SL ширина головной “костной пластинки” составляет 9.3–13.6%; аА – 63.1–68.5%, aD – 34.1–40.3%, aP – 19.9–29.5%, aV – 26.9–31.5%, основание D – 6.4–11.0%, основание A – 16.4–19.6%, длина хвостового стебля – 14.1–18.2 (12.9)%, высота хвостового стебля – 2.8–4.6%.
Головная “костная пластинка” и осевой скелет
Расположенная на верхней части головы рыб “костная пластинка” – это сросшиеся левая и правая лобные кости. На ализариновых препаратах I. agassizii хорошо видны проходящие в ней два надглазничных канала, каждый из которых открывается шестью порами (so1–so6); задняя сторона лобных костей плотно срощена с теменными костями, шов между ними хорошо различим (рис. 1).
Общий вид осевого скелета I. agassizii до начала анального плавника представлен на рентгенограмме (рис. 2). Туловищный отдел позвоночника (рис. 2б) состоит из 21–23 туловищных позвонков; остистый отросток невральной дуги имеется уже на первом позвонке туловищного отдела; зигапофизы разной степени развитости имеются на всех позвонках. Первая interneurale (inn) спинного плавника внедряется между остистыми отростками 17-го и 18-го позвонков; перед первым лучом D может быть одна свободная inn; свободных interhaemalia (inhm) перед первым лучом анального плавника, как правило, нет; первая inhm анального плавника внедряется между гемальными отростками 37-го и 38-го позвонков (рис. 2а). Эпиневральные и эпиплевральные ребра у исследованных экземпляров имеются, начинаются с первого туловищного позвонка и присутствуют вплоть до 19–23-го позвонков. Истинных хорошо развитых плевральных ребер 10–11 пар; 3–8-я пары прикрепляются к телу позвонка над презигапофизом; 9–13-я расположены на парапофизах и постепенно смещаются к их вершине (рис. 2).
Хвостовой отдел позвоночника. Хвостовых позвонков 37 (рис. 2а, 3). Первый гемальный остистый отросток (sphm1) у данного вида отходит от гемальной дуги 22–24-го позвонков. Три последних (преуральных) позвонка (pu2–pu4) по сравнению с предыдущими хвостовыми позвонками несколько более удлиненные (рис. 3). Первый преуральный позвонок (pu2) имеет сложное строение: от него отходит длинный гемальный отросток, к которому причленяются два нижних краевых луча С. От проксимальной части гемального отростка этого позвонка, как у предшествующего и последующего (здесь от parhypurale) позвонков, отходит вниз вперед и назад костное образование, хорошо различимое на рентгенограммах; мы сочли возможным назвать его “каудальный гемальный отросток”. Невральная дуга последнего из этих позвонков у ипнопсов хорошо развита, и ее невральный отросток располагается над преуральным (pu2) и первым уральным (u1) центрами.
Рис. 3.
Рентгенограмма хвостового комплекса Ipnops agassizii: С – основные лучи хвостового плавника; H3–H5 – эпаксиальная гипуральная пластинка; H1–H2 – гипаксиальная гипуральная пластинка; ер – epurale; ph – parhypuralia; pu1 и pu2 − преуральные центры 1 и 2; pu1 + u1 + u2 – уростилярный центр; r – верхние и нижние краевые лучи хвостового плавника; hspu2 – гемальный отросток центра pu2; nspu1 – редуцированный невральный отросток центра pu1; lhaem – каудальная гемальная пластинка; hppz – haemopostparapophysis.
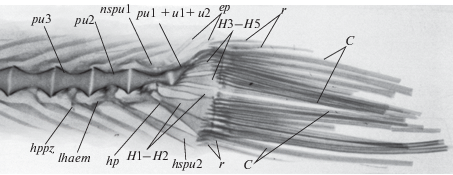
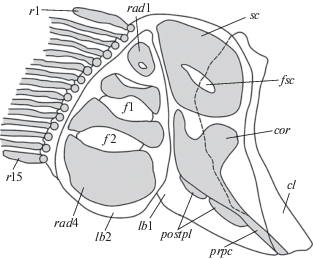
Рис. 4.
Схема строения грудного пояса и грудного плавника Ipnops agassizii: cl – клейтрум; lb1 и lb2 – передняя и задняя базальные хрящевые пластинки; sc – лопатка; fsc – скапулярное отверстие; cor – коракоид; prpc – посткоракоидный отросток; postpl – ребра жесткости; rad1–rad4 – радиалии; f1 и f2 – межрадиальные отверстия; r1 и r15 – редуцированные лучи грудного плавника.
Уростилярный позвонок. Пять гипуральных пластинок (H1–H2 + H3–H5) служат опорой для 17–18 основных лучей хвостового плавника. Как видно на рентгенограмме (рис. 3), к уростилю причленяются верхние эпаксиальные гипуральные пластины H3–H5.
Особенностью урального центра является наличие двух так называемых переходных зон, которые обозначают границу между pu1 и тонким изогнутым уростилем (первая зона), а также в уростилярном позвонке между u1 и u2 (вторая зона). К уральному центру u1 причленяются гипуралии H5, H4 и H3; наибольшая из них – Н4. Над уростилем расположены 2 удлиненные и слабо обызвествленные epuralia; ep2 служит опорой для двух верхних краевых лучей. Гипаксиальные пластинки представлены H1 + H2 + parhypurale. Parhypurale плотно примыкает верхним краем к Н1.
Непарные и парные плавники
Спинной плавник один, с 9 лучами, которые поддерживаются 9–10 interneuralia; его начало находится на вертикали между остистыми отростками 19-го и 20-го туловищных позвонков; перед первым очень коротким лучом плавника обычно имеется свободная, слабо окостеневшая inn; первый и второй лучи D сближены (рис. 2а, 2б).
Анальный плавник. В анальном плавнике 14–16 лучей, которые поддерживаются 13–15 interhaemalia; анальный плавник значительно сдвинут назад, его первый луч находится на вертикали 37–38 позвонков (рис. 2а). Лучи анального плавника значительно расставлены друг от друга и поддерживаются срединной (хрящевой) частью inhm, которая имеет длинную заднюю часть, вплотную подходящую к следующей inhm, что и обусловливает раздвинутость лучей А. Последний луч А двойной, поддерживается единой inhm.
Хвостовой плавник. В хвостовом плавнике 21–22 луча (рис. 3), из которых 17–18 лучей основные, они поддерживаются гипуральными пластинками Н1–Н5, parhypurale и задней частью uroneurale; имеется только один или два верхних краевых луча, которые поддерживаются ep2, и 1–2 нижних краевых луча, их поддерживает гемальный отросток pu2. Между гипуральными пластинками H2 и H3 обычно расположен один луч, который, как правило, не связан ни с одной из пластинок. Формула хвостового плавника: 1–2 + + 8–9 + 1 + 8 + 2 = 21–22. На рентгенограмме (рис. 3) видно, что верхние 2–3 основных луча своими основаниями заходят далеко вперед над основными лучами С. Поддерживаемые ep2, своими основаниями они достигают почти половины уростиля, что отмечено у многих видов рыб с двулопастным хвостовым плавником (Gosline, 1995). Следует отметить, что первый верхний и последний нижний краевые лучи по величине и степени окостенения сходны с основными хвостовыми лучами. Лучи хвостового плавника слабо сегментированы; второй и третий основные лучи сверху и снизу слабо раздвоены.
Плечевой пояс и грудной плавник (рис. 4). В передней хрящевой базальной пластинке (lb1) хорошо различимы лопатка и коракоид, которые занимают практически всю ее площадь, оставляя свободными лишь узкие участки хряща по краю соприкосновения со второй пластинкой lb2. Коракоид полулунной формы, несет две небольшие разноразмерные рукоятки, каждая из которых независимо причленяется к клейтруму. В хрящевой базальной пластинке lb2 расположены четыре удлиненные радиалии (rad1−rad4). Две средние (rad2 и rad3) гантелеобразной формы, так как имеют глубокие верхние и нижние радиальные вырезки; размер радиалий увеличивается от верхней к нижней, самая маленькая верхняя (rad1) и самая крупная нижняя (rad4) имеют квадратно-округлую форму. Rad1 целиком примыкает к заднему краю лопатки; в ее центре имеется маленькое замкнутое отверстие, отсутствующее в трех других радиалиях. Все радиалии расположены близко друг к другу, так как мало раздвинуты хрящом базальной пластинки.
В грудном плавнике 13–15 лучей (рис. 2б). Второй и третий верхние лучи самые длинные, в дистальной части на внешней стороне несут 19–20 (22) бугорковидных мягких сосочков. Как правило, второй луч заканчивается бульбообразным расширением и его конец несколько загибается кверху; остальные лучи в прижатом состоянии направлены прямо назад вдоль тела. На рентгенограмме (рис. 2а) видно, что концы целых лучей плавника заходят за вертикаль 15–18-го туловищных позвонков и немного заходят за вертикаль основания первого луча спинного плавника, что хорошо видно и на теле рыбы.
Брюшные плавники. В брюшных плавниках 8, редко 7 лучей. Правый и левый плавники широко расставленные, расположены практически горизонтально по отношению к телу рыбы; основание их внешних лучей находится на вертикали между 11-м и 12-м туловищными позвонками; наиболее длинные срединные лучи плавника немного заходят за вертикаль третьего–четвертого луча спинного плавника (рис. 2а).
Распространение
Ipnops agassizii – батиально-абиссально-ультраабиссальный вид. Во время экспедиции НИС “Дмитрий Менделеев” (1975–1976 гг.) впервые пойман на ультраабиссальных глубинах 5400–5410 м в приантарктических водах. Ранее представители семейства Ipnopidae в данном регионе известны не были. Во время экспедиции рыбы этого вида были пойманы также на материковом склоне (батиаль) и практически на абиссальных глубинах в юго-западной части моря Фиджи и в Большом Австралийском заливе (плато Натуралистов) на глубине 3020–3880 м. Среди исследованных нами рыб три молодых экземпляра стандартной длиной (SL) 55, 62 и 74.6 мм выловлены донным тралом вместе со взрослыми рыбами на глубинах 3020–3880 м, причем экземпляр SL 55 мм находился в трале с самым крупным экземпляром среди пойманных представителей этого вида ипнопид (SL 132.3 мм).
ОБСУЖДЕНИЕ
Во взрослом состоянии рыбы рода Ipnops обладают уникальной и не встречающейся у других рыб анатомической особенностью – строением глазного отдела головы. Первая характеристика этого рода дана Гюнтером (Günther, 1880), который в диагнозе рода указал на наличие у рыб широкого уплощенного рыла, вся верхняя поверхность которого представляет собой продольно разделенный на две симметричные половины особенный орган зрения (или люминесценции); он же заметил, что этот орган зрения, “по-видимому, потерял свою функцию и принял на себя функцию производства света” (Günther, 1880, с. 585), т.е. свечения. Давая характеристику ипнопид, Парин (1971) указал, что рыбы данного семейства вообще не имеют морфологически сформированных глаз; функция головных органов, расположенных под парными костными пластинками, осталась для автора неясной, хотя позднее он высказал мнение, что “так называемые глазные пластины исполняют роль светочувствительных органов” (Парин, 1988, с. 189).
По сути расположенная на верхней части головы “костная пластинка” – это сросшиеся левая и правая лобные кости, так как в каждой из них проходит надглазничный канал, хорошо и четко видимый на ализариновом препарате; при этом задняя сторона лобных костей плотно срощена с теменными костями. Эта слитая воедино лобно-теменная кость покрывает большие дорсально направленные “глаза” (Theisen, 1965), вернее, как показали гистологические исследования, особую аналогичную сетчатке глаза светочувствительную ткань или видоизмененную сетчатку (Theisen, 1965; Franco et al., 2009). Более подробно “костная пластинка” изучена по ализариновому препарату у I. murrayi (см.: Munk, 1959). Именно этот автор впервые выявил и показал наличие в ней суборбитальных (надглазничных) каналов, а также отметил наличие под боковым краем пластинки другого − инфраорбитального (подглазничного) канала, чем подтвердил предположение, высказанное ранее при характеристике рода Ipnops (Parr, 1928), что пластинка представляет собой не что иное, как лобные кости. Изучив строение “головной пластинки” по ализариновому препарату у I. agassizii, мы обнаружили проходящие в парных “пластинках” надглазничные каналы, каждый из которых открывается шестью порами (рис. 1). Как и у I. murrayi (см.: Theisen, 1965), оба канала начинаются и заканчиваются открытыми крупными порами so1 и so6 соответственно.
Изучение анатомических особенностей головы, осевого скелета, скелета парных и непарных плавников и хвостового комплекса I. agassizii позволило выявить морфологические преобразования этих структур в процессе специализации в связи с жизнью на глубине. Как и у всех видов рода, у I. agassizii имеются светочувствительные органы головы, покрытые прозрачными черепными костями. Особый интерес представляет то, что у личинки ипнопса SL 13.9 мм имеются большие латерально расположенные и еще морфологически не измененные настоящие глаза (Okiyama, 1981, c. 248, рис. 1). Изменение морфологии зрительного органа у донных глубоководных рыб во взрослом состоянии, скорее всего, обусловлено необходимостью лучшего восприятия слабого свечения организмов – объектов их питания, обитающих в придонно-пелагическом биоценозе. Эта уникальная особенность, а также большой рот с острыми тонкими мелкими зубами для удержания крупной добычи и хорошо развитые при ловле мелких объектов жаберные тычинки (рис. 2б) − признаки, присущие глубоководным аулопиформам, ведущим своеобразный образ жизни в полной темноте в условиях дефицита пищи на максимальных глубинах океана. К этому следует добавить и выявленный у данных рыб гермафродитизм (Marshall, Staiger, 1975; Helfmann et al., 1997; Davis, Fielitz, 2010).
Необходимо отметить редукционный характер скелета хвостового отдела плавника, который выражается в наличии лишь двух epuralia, сближенном положении parhypurale и hypurale1 и малом числе краевых лучей в верхней и нижней частях хвостового плавника − соответственно 1−2/2. На первом преуральном позвонке обнаружено отходящее вниз вперед и назад костное образование, хорошо различимое на рентгенограммах и названное нами каудальным гемальным отростком. Этот морфологический элемент пока не выявлен у других рыб (Monod, 1968; Kusaka, 1974). Однако сходный элемент на гемальной дуге последних туловищных позвонков описан у бычков-крючкорогов (Неелов, 1979: рис. 69 , с. 105), а сильно уплощенные широкие гемальные отростки, как и невральные в виде пластин, имеются на последних хвостовых позвонках у скумбриевых рыб и тунцов (Hotta, 1961).
Для чрезвычайно специализированного рода Ipnops, к которому относится исследованный нами вид, характерно более сильное развитие костной составляющей, уменьшение хряща в грудном поясе и усиление окостенения поддерживающих элементов всех лучей непарных плавников при слабом окостенении структурных элементов позвонков (Sulak, 1977; наши данные). На примере рыб отряда Aulopiformes также показано, что лучи грудного плавника хорошо иннервируются в отличие от слабо иннервированных лучей брюшных и хвостового плавников, которые испытывают меньшую силовую нагрузку (Sulak, 1977).
Исследованный авторами материал по Ipnops agassizii добыт с батиальных и ультраабиссальных глубин (“хадаль” по: Андрияшев, 1979, 2003; “абиссо-хадобенталь” по: Парин, 1988). Анализ глубинного распределения I. agassizii выявил значительный диапазон его встречаемости по глубине (Щербачев, 1980). Известно, что в субтропической части западной Пацифики молодь I. agassizii (SL 42–55 мм) ловили в батипелагиали на глубине 1810–1990 м (Okiyama, 1986), а личинка SL 13.3 мм была поймана в эпипелагиали (Okiyama, 1981, 1984). Наиболее молодые экземпляры среди исследованных нами рыб были пойманы в самой нижней батиальной вертикальной зоне – в подножии континентальной ступени (по: Андрияшев, 1979). Таким образом, молодые рыбы могут быть обнаружены как на абиссальных, так и на батиальных глубинах, а личинки разной стадии метаморфоза – в толще воды в эпи-, мезо- и батипелагиали. Поимка молоди ранней стадии развития этих глубоководных донных рыб позволяет говорить об онтогенетической вертикальной миграции рыб рода Ipnops и, возможно, других представителей семейства Ipnopidae (Okiyama, 1984).
Список литературы
Андрияшев А.П. О некоторых вопросах вертикальной зональности морской донной фауны // Биологические ресурсы Мирового океана. М.: Наука. 1979. С. 117–138.
Андрияшев А.П. Липаровые рыбы (Liparidae, Scorpaeniformes) Южного океана и сопредельных вод // Исслед. фауны морей. СПб.: Наука. 2003. Т. 53(61). 478 с.
Андрияшев А.П., Неелов А.В., Природина В.П. К методике изучения морфологии и систематики рыб сем. Liparidae // Зоол. журн. 1977. Т. 56. Вып. 1. С. 141–147.
Неелов А.В. Сейсмосенсорная система и классификация керчаковых рыб. Л.: Наука. 1979. 208 с.
Парин Н.В. Семейство Ипноповые (Ipnopidae) // Жизнь животных. Т. 4. Ч. 1: Рыбы. М.: Просвещение. 1971. С. 223.
Парин Н.В. Рыбы открытого океана. М.: Наука. 1988. 271 с.
Природина В.П., Цветков И.В. Остеологическая характеристика осевого скелета и пояса грудных плавников некоторых видов евразийских пресноводных рыб семейства Cottidae // Тр. Зоол. ин-та РАН. 2016. Т. 320. № 1. С. 71–83.
Сазонов Ю.И. Морфология и классификация рыб семейства Platytroctidae (Salmoniformes, Alepocephaloidei) // Тр. Ин-та океанол. 1986. Т. 121. С. 51–96.
Щербачев Ю.Н. Предварительный обзор индоокеанских видов семейства Chlorophthalmidae (Myctophiformes) // Рыбы открытого океана. М.: Ин-т океанол. АН СССР. 1980. С. 47–67.
Якубовский М. Методы выявления и окраски системы каналов боковой линии и костных образований у рыб in toto // Зоол. журн. 1970. Т. 49. Вып. 9. С. 1398–1402.
Davis M.P., Fielitz Ch. Estimating divergence of lizardfishes and their allies (Euteleostei: Aulopiformes) and the timing of deep-sea adaptations // Mol. Phylogenet. Evol. 2010. V. 57. P. 1194–1208.
Fowler H.W. Descriptions and figures of new fishes obtained in Philippine seas and adjacent waters by the United States Bureau of Fisheries Steamer “Albatross” // Contributions to the Biology of the Philippine Archipelago and Adjacent Regions. U. S. Natl. Mus. Bull. № 100. 1943. V. 14. Part 2. P. 51−91.
Franco M.A.L., Braga A.C., Nunan G.W.A., Costa P.A.S. Fishes of the family Ipnopidae (Teleostei: Aulopiformes) collected on the Brazilian continental slope between 11° and 23° S // J. Fish Biol. 2009. № 75. P. 797–815.
Garman S. Reports on an exploration off the West coasts of Mexico, Central and South America, and off the Galapagos Islands, in Charge of Alexander Agassiz, by the United States Fish Commission Steamer “Albatross,” during 1891, Lieut. Commander Z.I. Tanner, U. S. N., Commanding. V. XXVI: The Fishes // Mem. Mus. Comp. Zool. Harvard College. 1899. V. XXIV. 431 p.
Gosline W.A. Function and structure in the paired fins of scorpaeniform fishes // Environ. Biol. Fishes. 1995. № 40. P. 219–226.
Gray M. The distribution of fishes found below a depth of 2000 meters // Fieldiana: Zool. 1956. V. 36. № 2. P. 75–337.
Günther A.C. An Introduction to the Study of Fishes. Edinburgh: Adam and Charles Black. 1880. P. 582–585.
Helfman G.S., Collette B.B., Facey D.E. The Diversity of Fishes. Maiden, Mass.: Blackwell Science Inc. 1997. P. 1–544.
Hotta H. Comparative Study of the Axial Skeleton of Japanese Teleostei // Tokyo: Nippon Gyogaku Shinkokai. 1961. 155 p.
Johnson G.D., Baldwin C.C., Okiyama M., Tominaga Y. Osteology and relationships of Pseudotrichonotus altivelis (Teleostei: Aulopiformes: Pseudotrichodontidae) // Ichthyol. Res. 1996. V. 43. P. 17–45.
Kusaka T. The Urohyal of Fishes // Tokyo: Univ. of Tokyo Press. 1974. XIV + 320 p.
Marshall N.B., Staiger J.C. Aspects of the structure, relationships, and biology of the deep-sea fish Ipnops murrayi (Family Bathypteroidae) // Bull. Mar. Sci. 1975. V. 25. № 1. P. 101–111.
McEachran J.D., Fechhelm J.D. Fishes of the Gulf of Mexico: Myxiniformes to Gasterosteiformes. Austin, Tex.: Univ. of Texas Press. 1998. V. 1. 1112 p.
Merrett N.R., Nielsen J.G. A new genus and species of the family Ipnopidae (Pisces, Teleostei) from the Eastern North Atlantic, with notes on its ecology // J. Fish Biol. 1987. V. 31. P. 451−464.
Monod T. Le complexe urophore des poissons téléostéens // Mem. Inst. Fondam. Afr. Noire. 1968. № 81. S. I-VI + I-705. 989 Figs. 3 Tabls.
Munk O. The eyes of Ipnops murrayi Günther, 1878 // Galathea Rep. 1959. V. 3. P. 79–87.
Nelson J.S. Fishes of the World. New York: John Wiley & Sons. 2006. 602 p.
Nielsen J.G. Synopsis of the Ipnopidae (Pisces, Iniomi) with description of two new abyssal species // Galathea Rep. 1966. V. 8. P. 49–75.
Okiyama M. A larval Ipnops and its possible metamorphosing process // Jpn. J. Ichthyol. 1981. V. 28. № 3. P. 247–253.
Okiyama M. Travel between surface and abyssal bottom during early life of the ipnopid fish: a risky migration? // Program and abstracts of 3rd international symposium on the early life history of fishes and 8th annual larval fish conference. University of British Columbia, Vancouver, British Columbia, 6−10 May, 1984. P. 20.
Okiyama M. Bathypelagic capture of a metamorphosing juvenile of Ipnops agassizi (Ipnopidae, Myctophiformes) // Jpn. J. Icthyol. 1986. V. 32. № 4. P. 443–445.
Parr A.E. Deepsea fishes of the order Iniomi from the waters around the Bahama and Bermuda Islands: With annotated keys to the Sudididae, Myctophidae, Scopelarchidae, Evermannellidae, Omosudidae, Cetomimidae and Rondeletidae of the world // Bull. Bingham Oceanogr. Collect. 1928. V. 3. № 3. P. 1–193.
Sulak K.J. The systematics and biology of Bathypterois (Pisces, Chlorophthalmidae) with the revised classification of benthic myctophiform fishes // Galathea Rep. 1977. V. 14. P. 49–108.
Sulak K.J. Family No. 76: Chlorophthalmidae // Smith’s Sea Fishes. M.M. Smith, Ph.C. Heemstra (eds.). Berlin: Springer-Verlag. 1986. P. 261–265.
Theisen B. On the cranial morphology of Ipnops murrayi Günther, 1878 with special reference to the relations between the eyes and the skull // Galathea Rep. 1965. V. 8. P. 7–20.
Дополнительные материалы отсутствуют.