

Биология моря, 2020, T. 46, № 2, стр. 91-99
Морфологическое описание и биология малоизвестных глубоководных скатов рода Bathyraja Ishiyama, 1958 (Rajidae) из северо-западной части Тихого океана
В. Н. Долганов *
Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: vndolganov@mail.ru
Поступила в редакцию 27.03.2019
После доработки 12.09.2019
Принята к публикации 03.10.2019
Аннотация
Дано морфологическое описание и приведены сведения о биологии редких глубоководных скатов Bathyraja fedorovi, B. andriashevi и B. tzinovskii из северо-западной части Тихого океана. Описано строение неврокраниума, скапулокоракоида и птеригоподиев, а также приведены данные о количестве туловищных и хвостовых позвонков, рядов зубов и оборотов спирального клапана кишечника.
В 1985 г. из северо-западной части Тихого океана было описано пять новых видов скатов рода Bathyraja (см.: Долганов, 1985), из которых три вида – B. andriashevi (голотип и один несохранившийся экземпляр), B. fedorovi (голотип и паратип) и B. tzinovskii (голотип) – были известны только по единичным экземплярам первоописания, а данные по их биологии отсутствовали. Из-за ограниченности материала к моменту описания видов возможности для изготовления морфологических препаратов не было, поэтому такие важные для таксономии скатов признаки, как строение птеригоподиев, неврокраниума и скапулокоракоида, были неизвестны. Во время экспедиции НПС “Дарвин” в 1989 г. на материковом склоне Охотского моря автором был собран материал, позволяющий значительно дополнить недостающие сведения о морфологии и биологии этих редких видов скатов.
МАТЕРИАЛ И МЕТОДИКА
Возраст скатов определяли по шипам срединного ряда, на которых после очистки хорошо видны годичные кольца (Долганов, 1998). Степень созревания особей определяли не только по развитию гонад, но и по степени развития ректальной железы самок и эректильных колючек самцов. Сбор и обработку желудков проводили в соответствии с “Методическим пособием…” (1974). Количественно-весовым методом были обработаны желудки 57 экз. Bathyraja fedorovi, 5 экз. B. andriashevi и 4 экз. B. tzinovskii.
Сокращенные обозначения признаков: TL – общая длина, DW – ширина диска, DL – длина диска, lT – длина хвоста от середины ануса, c – длина головы до начала позвоночной пластины, ao – длина рыла до орбит, o – наибольший диаметр орбиты, io – межорбитальное расстояние, s – длина брызгальца, is – расстояние между брызгальцами, an – длина рыла до ноздрей, in – расстояние между ноздрями, ar – длина рыла до ротовой щели, r – ширина рта, lID и lIID – длина оснований соответственно первого и второго спинных плавников, hID и hIID – их высота, lcl – длина птеригоподиев взрослых самцов; P – количество лучей в грудном плавнике, vt. tr. – туловищных позвонков, vt. prd. – предорсальных позвонков, int. v. – витков в спиральном клапане, dent. – поперечных рядов (серий) зубов в верхней и нижней челюсти.
Номенклатура строения неврокраниума, скапулокоракоида, птеригоподиев и их скелета, используемая в работе, соответствует общепринятой (Ishiyama, 1958; Stehmann, 1970; McEachran, Compagno, 1982).
МОРФОЛОГИЧЕСКОЕ ОПИСАНИЕ
Bathyraja fedorovi Dolganov, 1985 – скат Федорова (рис. 1)
Рис. 1.
Bathyraja fedorovi. а – голотип, взрослая самка TL 733 мм; б – голова, вид снизу (Долганов, 1985); в–ж – взрослый самец TL 673 мм: в – неврокраниум (RA – боковые ростральные отростки, RS – рострум, AF – передняя фонтанель, BF – базальные отверстия); г – скапулокоракоид (ADF – переднее дорзальное отверстие, AVF – переднее вентральное отверстие, PDF – задние дорзальные отверстия, PVF – задние вентральные отверстия); д – тазовая пластина; е – внешний вид развернутого птеригоподия (PS – псевдосифон, HP – гипопиле, SL – выемка, PJ – отросток, SP – острие); ж – вентральный и дорзальный вид скелета птеригоподиев (AX – осевой хрящ; DM – дорзальный осевой хрящ; VM – вентральный осевой хрящ; DT 1, DT 2 и DT 3 – 1-й, 2-й и 3-й дорзальные щитки; AT – добавочный хрящ; VT – вентральный щиток).
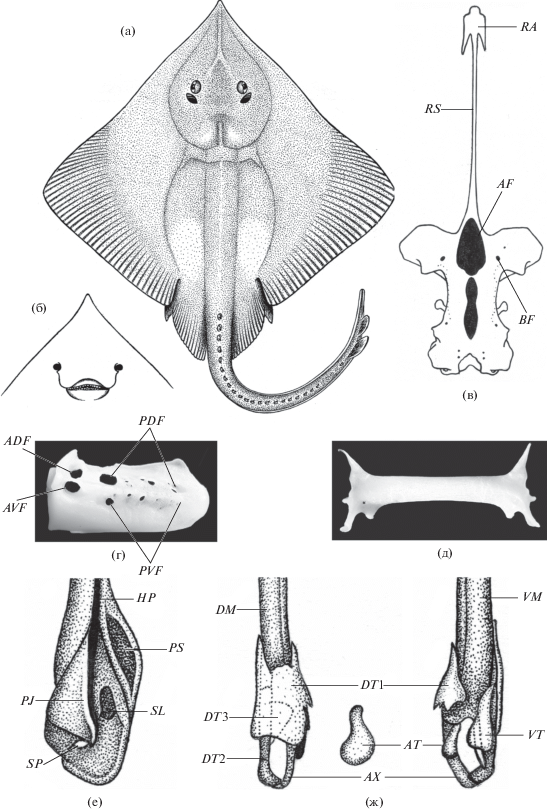
Материал. Голотип: взрослая самка TL 733 мм, DW 420 мм, ЗИН № 46190, НПС “Геракл”, 9 августа 1975 г., Охотское море, 44°41′ с.ш., 146°12′ в.д., глубина лова 1370–1550 м. Коллектор В.В. Федоров. Паратип: самка TL 698 мм, DW 417 мм, музей ТИНРО № 750, НПС “Мыс Дальний”, 9 февраля 1981 г., тихоокеанское побережье Японии, 37°58′ с.ш., 142°48′ в.д., глубина лова 1400 м. Коллектор Б.А. Шейко. Дополнительный материал: 28 самок TL 292–727 мм, DW 180–410 мм, 14 самцов TL 267–699 мм, DW 168–417 мм, Охотское море, май–сентябрь 1989 г., НПС “Дарвин”, донный трал, глубина лова 1110–2025 м. Коллектор В.Н. Долганов.
P 69–74; vt. tr. 27–31, vt. prd. 58–73, int. v. 9–11, dent. 17–26/15–22.
Описание. Рыло острое, образует угол менее 90°. Глаза маленькие, намного меньше межорбитального расстояния, их длина около 5 раз содержится в длине рыла до глаз. Брызгальце в любом возрасте меньше диаметра глаза и лишь изредка равно ему у наиболее крупных экземпляров. Носовой клапан с неглубокой выемкой. Диск ромбовидный, с заостренными внешними и задними углами грудных плавников; его длина составляет 76.3–92.0 (80.9)% ширины. Длина хвоста от середины ануса больше предхвостового расстояния. Размеры спинных плавников одинаковы; хвостовой плавник в виде невысокой складки. Хвост с боковыми кожными складками по всей длине.
Зубы с маленькими заостренными вершинами, околосимфизные крупнее боковых. Глотка и жаберные дуги без шипиков. Шипы на диске обычно отсутствуют. У молодых особей шипы срединного ряда иногда имеются. Хвостовая серия шипов срединного ряда начинается далеко позади тазовых бугров и состоит из 20–31 (24.3) шипа. Междорзальный шип обычно отсутствует. Эректильные (аларные) колючки взрослых самцов расположены в 21–23 сериях по 5–7 колючек в каждой. Сверху диск и хвост покрыты мелкими шипиками. Нижняя поверхность голая, кроме двух полос шипиков, расположенных вдоль боковых складок хвоста.
Измерения B. fedorovi в % DW (в скобках среднее значение признака): TL 157.5–181.8 (163.8); DL 76.3–92.0 (80.9); c 29.3–36.5 (33.0); ao 19.3–26.2 (23.1); o 4.0–6.0 (4.5); io 4.8–7.2 (6.0); s 3.3–4.6 (3.8); is 10.6–12.1 (11.1); an 15.0–20.5 (17.9); in 11.8–14.9 (13.5); ar 19.8–28.2 (24.2); r 10.7–13.8 (11.9); lT 80.7–97.9 (85.7); lcl 26.6–29.6 (28.1); lID 4.9–7.4 (6.1); lIID 4.9–7.4 (5.9); hID 1.8–4.1 (2.8); hIID 1.8–4.4 (2.8).
Туловищных позвонков 27–31 (29.5), предорзальных 58–73 (65.0); лучей в грудном плавнике 69–74 (71.1). В верхней челюсти 17–26 (22.2) серий зубов, в нижней – 15–22 (18.3). Складка спирального клапана образует 9–11 (10.1) полных оборотов.
Неврокраниум сравнительно узкий. Ростральный хрящ цельный, очень слабый и длинный; он намного длиннее половины длины черепа. Боковые ростральные хрящи маленькие, их длина около 5 раз содержится в длине рострума; задние края хрящей плоские в сечении. Передняя фонтанель широкая, открывается у начала обонятельных капсул и простирается до уровня начала орбит. Базальные отверстия носовых капсул маленькие (рис. 1в). Скапулокоракоид невысокий. Имеется перемычка (anterior bridge), разделяющая переднюю фонтанель на дорзальную и вентральную части. Постдорзальных и поствентральных отверстий 7–8 (рис. 1г). Передний и задний края тазовой пластины прямые (рис. 1д).
Птеригоподии некрупные, около 1/3 длины хвоста. Псевдосифон (pseudosiphon) крупный. Внутренняя поверхность дорзальной доли птеригоподия с неглубокой выемкой (slit). Дорзальная и вентральная доли разделены срединной впадиной – гипопиле (hypopyle). C вентральной стороны к срединной впадине примыкают вытянутый к дистальному краю птеригоподия изогнутый отросток (projection) и примыкающий к дорзальной доле небольшой выступ – острие (spike), покрытый складкой кожи (рис. 1е).
Скелет птеригоподия B. fedorovi состоит из осевого хряща (axial), к которому продольно прикреплены дорзальный (dorsal marginal) и вентральный (ventral marginal) осевые хрящи. Дорзальный осевой хрящ в дистальной части дифференцирован на три дорзальных щитка (dorsal terminal 1, 2, 3), а вентральный осевой хрящ – на овальный добавочный хрящ (accessory terminal) и крупный вентральный щиток (ventral terminal) (рис. 1ж).
Окраска взрослых скатов сверху и снизу темно-коричневая. Верхняя поверхность молоди более светлая, чем нижняя.
Распространение. Глубокие воды Охотского моря и тихоокеанского побережья Японии (1100–2025 м).
Bathyraja andriashevi Dolganov, 1985 – скат Андрияшева (рис. 2)
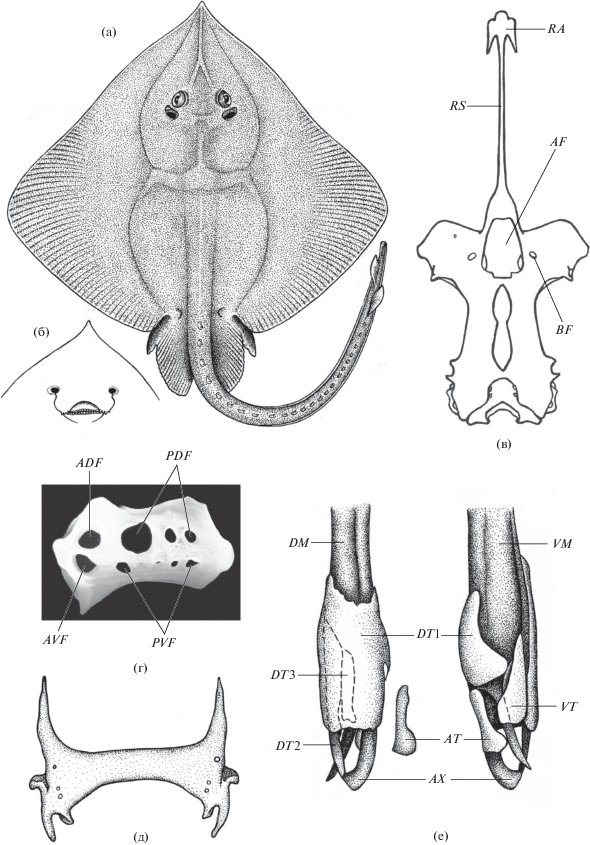
Рис. 2.
Bathyraja andriashevi. а – голотип, взрослая самка TL 1032 мм; б – голова, вид снизу (Долганов, 1985); в–д – ювенильный самец TL 550 мм (в – неврокраниум, г – скапулокоракоид, д – тазовая пластина); е – взрослый самец TL 1200 мм (внешний вид развернутого птеригоподия). Условные обозначения, как на рис. 1.

Материал. Голотип: самка TL 1032 мм, DW 681 мм, ЗИН № 46191, НПС “Мыс Юноны”, 23 января 1980 г., тихоокеанское побережье Японии, 36°24′ с.ш., 141°29′ в.д., глубина лова 1470–1480 м. Коллектор В.Н. Долганов. Дополнительный материал: взрослый самец TL 1200 мм, DW 756 мм, тихоокеанское побережье Японии, 36°25′ с.ш., 141°28′ в.д., 25 января 1980 г., НПС “Мыс Юноны”, глубина лова 1390–1400 м; самец TL 390 мм, DW 235 мм, Охотское море, 48°08′ с.ш., 148°05′ в.д., 24 мая 1989 г., НПС “Дарвин”, глубина 1960–1980 м; самец TL 550 мм, DW 348 мм, Охотское море, 48°46′ с.ш., 151°08′ в.д., 31 мая 1989 г., НПС “Дарвин”, глубина 1978–2004 м; две сильно поврежденные взрослые самки TL 132 и 141 см, Охотское море, 48°07′ с.ш., 149°11′ в.д., 25 мая 1989 г., НПС “Дарвин”, глубина 1948–1998 м. Коллектор В.Н. Долганов.
P 93–97; vt. tr. 40–43, vt. prd. 75–79, int. v. 11, dent. 29–30/25–28.
Описание. Рыло широкое, тупое; образует угол 120–125°. Глаза очень маленькие, 58.5–80.2% межорбитального расстояния и 13.1–22.7% длины рыла до глаз. Брызгальце большое, у взрослых особей 127.3–158.3% диаметра глаза, у молоди – 83.1–107.6%. Носовой клапан со слабой выемкой. Диск широкий, округлый; его длина составляет 75.3–79.6% ширины. Внешние и задние углы грудных плавников плавно закруглены, передний край прямой или слегка выпуклый. Длина хвоста от середины ануса составляет 104.1–106.4% предхвостового расстояния у взрослых особей и 125.4–129.3% у молоди. Первый спинной плавник немного крупнее второго, хвостовой плавник сильно редуцирован. Боковая складка хвоста начинается у внутренней пазухи брюшного плавника.
Зубы мелкие, с небольшими острыми вершинами; у взрослых самцов вершины более высокие и массивные. Глотка и жаберные дуги без шипиков. Шипы на диске отсутствуют. Хвостовая серия шипов начинается далеко позади тазовых бугров и состоит из 25–31 некрупного шипа. Междорзальный шип обычно имеется. Эректильные колючки взрослого самца расположены в 20 сериях; количество колючек в серии достигает 5. Имеется небольшое количество недоразвитых колючек; это показывает, что серии могут быть более многочисленными. Сверху диск и хвост полностью покрыты очень мелкими шипиками, но у взрослых самцов центральная часть грудных плавников голая. Нижняя поверхность голая.
Измерения B. andriashevi в % DW: TL 151.5–166.0; DL 75.3–79.6; c 31.5–33.2; ao 22.1–24.7; o 3.2–5.1; io 4.6–6.4; s 4.0–5.0; is 8.3–9.8; an 17.2–18.1; in 11.2–12.6; ar 21.2–25.1; r 8.5–10.3; lT 78.1–93.6; lcl 20.9; lID 5.6–7.2; lIID 4.7–5.5; hID 3.2–3.5; hIID 3.2–3.5.
Туловищных позвонков 40–43, предорзальных 75–79; лучей в грудном плавнике 93–97. В верхней челюсти 29–30 серий зубов, в нижней – 25–28. Складка спирального клапана образует 11 полных оборотов.
Неврокраниум сравнительно широкий. Ростральный хрящ цельный, тонкий; его длина немного более половины длины черепа. Боковые ростральные хрящи очень маленькие, их длина около 5 раз содержится в длине рострума; задние края хрящей плоские в сечении. Передняя фонтанель широкая, открывается примерно на уровне начала обонятельных капсул и простирается до начала орбит. Базальные отверстия носовых капсул небольшие (рис. 2в). Скапулокоракоид умеренно высокий. Имеется перемычка, разделяющая переднюю фонтанель на дорзальную и вентральную части. Постдорзальных и поствентральных отверстий 5–6 (рис. 2г). Передний край тазовой пластины почти прямой, задний край сильно вогнут (рис. 2д).
Дорзальная поверхность птеригоподиев с крупным псевдосифоном. На вентральной доле внутренней поверхности имеются хорошо выраженный удлиненный выступ и расположенный под ним округлый хрящ с острым краем – острие. На дорзальной доле находится неглубокая выемка. Вентральная и дорзальная доли птеригоподиев разделены глубокой продольной впадиной – гипопиле (рис. 2е).
Прижизненная окраска верхней поверхности диска и хвоста взрослых самок светло-сиреневая с неясными серыми пятнами разных размеров; взрослый самец серый с небольшими светлыми пятнами. Молодые скаты однотонные, серо-фиолетовые или коричнево-серые. Нижняя поверхность светлая с широкой темной окантовкой грудных и брюшных плавников.
Распространение. Известен у тихоокеанского побережья о-ва Хонсю на глубинах 1390–1480 м и в Курильской котловине Охотского моря на глубинах 1960–2004 м.
Bathyraja tzinovskii Dolganov, 1985 – скат Циновского (рис. 3)
Рис. 3.
Bathyraja tzinovskii. а – голотип, взрослая самка TL 714 мм; б – голова, вид снизу (Долганов, 1985); в –е – взрослый самец TL 730 мм (в – неврокраниум, г – скапулокоракоид, д – тазовая пластина, е – вентральный и дорзальный вид скелета птеригоподиев). Условные обозначения, как на рис. 1.
Материал. Голотип: взрослая самка TL 714 мм, DW 415 мм; ЗИН № 46192, 23 июня 1972 г., тихоокеанское побережье Японии, 40°12′ с.ш., 143°35′ в.д., глубина лова 2500 м. Коллектор В.Д. Циновский. Дополнительный материал: два взрослых самца TL 743 и 730 мм, DW 440 и 457 мм, Охотское море, 48°02′ с.ш., 146°49′ в.д., 22 мая 1989 г., НПС “Дарвин”, глубина лова 2025–2100 м; созревающий самец TL 682 мм, DW 392 мм, Охотское море, 48°06′ с.ш., 147°09′ в.д., 22 мая 1989 г., НПС “Дарвин”, глубина лова 1745–1766 м. Коллектор В.Н. Долганов.
P 70–72; vt. tr. 30–31, vt. prd. 64–69, int. v. 8–9, dent. 26–32/23–32.
Описание. Рыло образует угол около 100°. Глаза сравнительно небольшие, 50.2–74.9% межорбитального расстояния и 16.2–21.1% длины рыла до глаз. Брызгальце составляет 85.0–117.2% диаметра глаза. Носовой клапан с неглубокой выемкой. Диск ромбовидный, со слабо заостренными внешними и задними углами грудных плавников; его длина составляет 78.3–86.0% ширины. Длина хвоста от середины ануса в любом возрасте больше предхвостового расстояния. Первый спинной плавник немного длиннее второго; хвостовой плавник выражен слабо. Боковые складки хвоста начинаются от основания брюшных плавников и постепенно расширяются назад.
Зубы мелкие, с направленными назад вершинами. Глотка и жаберные дуги без шипиков. Шипы на диске отсутствуют. Срединный ряд шипов представлен только хвостовой серией из 21–24 некрупных шипов. Эректильные колючки взрослого самца образуют до 23 серий по 6 колючек в каждой. Диск и хвост полностью покрыты густо сидящими мелкими шипиками; нижняя поверхность голая.
Измерения B. tzinovskii в % DW: TL 159.7–174.0; DL 78.3–86.0; c 32.2–34.9; ao 21.7–23.6; o 3.7–4.8; io 6.4–7.4; s 4.3–4.8; is 10.5–11.5; an 17.3–18.6; in 13.4–14.5; ar 23.6–25.0; r 11.8–13.5; lT 83.6–93.1; lcl 25.9–27.8; lID 6.6–8.0; lIID 6.0–6.8; hID 3.3–4.3; hIID 3.3–4.3.
Туловищных позвонков 30–31, предорзальных 64–69; лучей в грудном плавнике 70–72. В верхней челюсти 26–32 серий зубов, в нижней – 23–32. Складка спирального клапана образует 8–9 полных оборотов.
Неврокраниум сравнительно узкий. Ростральный хрящ цельный, без сочленения; его длина немного более половины длины черепа. Длина боковых ростральных хрящей около 5 раз содержится в длине рострума; задние края хрящей в сечении плоские. Передняя фонтанель широкая, открывается у начала обонятельных капсул и простирается до уровня начала орбит. Базальные отверстия носовых капсул маленькие (рис. 3в). Скапулокоракоид невысокий, с перемычкой, разделяющей переднюю фонтанель на дорзальную и вентральную части. Постдорзальных и поствентральных отверстий 3–5 (рис. 3г). Передний край тазовой пластины почти прямой, задний край вогнут (рис. 3д).
Скелет птеригоподия B. tzinovskii состоит из осевого хряща с прикрепленными к нему продольно дорзальным и вентральным осевыми хрящами. Дорзальный осевой хрящ в дистальной части дифференцирован на три дорзальных щитка, а вентральный осевой хрящ – на добавочный хрящ и вентральный щиток. Первый дорзальный щиток очень крупный, его значительная часть заходит на вентральную сторону птеригоподиев. Задний конец 2-го дорзального щитка прилегает к концу осевого хряща. Дистальные концы добавочного хряща и вентрального щитка расширены и закруглены (рис. 3е).
Окраска спинной поверхности фиксированного голотипа светло-кремовая, однотонная; брюшная поверхность белая. Прижизненная окраска верхней стороны диска от фиолетово-розовой до темно-серой. Нижняя поверхность светлая, но края грудных плавников с широкой и четко отграниченной темной полосой.
Распространение. Известен по голотипу, пойманному у тихоокеанского побережья о-ва Хонсю на глубине 2500 м и в Охотском море на глубинах 1745–2100 м.
БИОЛОГИЯ
Скаты Bathyraja andriashevi, B. fedorovi и B. tzinovskii относятся к наиболее глубоководным скатам мира. В 1989 г. во время полной съемки материкового склона Охотского моря на НПС “Дарвин” (300–2000 м) их встречаемость увеличивалась с глубиной. Это позволяет предположить, что основная зона обитания данных видов – гораздо большие глубины. Вместе со скатами Bathyraja abyssicola (710–2903 м), B. spinosissima (1250–2400 м) и Amblyraja hyperborea (890–2400 м) они принадлежат к батибентальному ихтиоцену северной части Тихого океана, представители которого обитают на глубинах от 1000 до 1500–3000 м (Парин, 1988). Два вида этого ихтиоцена – Raja hyperborea и Bathyraja spinosissima, являются космополитами, B. abyssicola широко распространен в северной Пацифике, а ареалы B. andriashevi, B. fedorovi и B. tzinovskii тяготеют к побережью Азии (Долганов, 1999).
Характеристики глубинных вод в течение года практически не изменяются (Шунтов, 2001). Этим обусловлена крайняя стенотермность скатов батибентального ихтиоцена, которые во время съемки 1989 г. встречались только при температуре 1.95–2.32°С. Вследствие однородности глубинных водных масс данные скаты характеризуются широким диапазоном распространения, слабо выраженными сезонными миграциями и отсутствием сезонности в размножении.
Размножение скатов семейства Rajidae довольно своеобразное. У этих видов внутреннее оплодотворение, и они откладывают на дно яйца, облаченные в яйцевую капсулу. Половой диморфизм выражен очень сильно: у взрослых самцов имеются крупные птеригоподии для внутреннего оплодотворения, образованные из внутренних лучей брюшных плавников, а для остановки и удержания самки – более острые, чем у самок, зубы, сильно вогнутые передние края грудных плавников и острые эректильные колючки. С наступлением половозрелости самки круглогодично в течение жизни откладывают яйца, которые постоянно находятся в яичниках на разной стадии созревания – от мельчайших безжелтковых до готовых к вымету. Созревшие яйца из яичников по яйцепроводам попадают в скорлуповые железы, где оплодотворяются и покрываются капсулой (Долганов, 1998). Яйцевая капсула имеет четыре роговидных выроста по углам, служащих для прикрепления к субстрату. Для этой же цели на задних более коротких рогах формируется по пучку длинных тонких фибриллярных нитей. Поверхность капсулы шероховатая, с множеством мелких выростов разной формы, которые расположены правильными рядами. В боковой части каждого роговидного выроста имеется узкая щель, через нее происходит обмен с внешней средой (Ishiyama, 1958). Размер яйцевой капсулы зависит от видовой принадлежности и размера самки. Так, у крупного вида B. andriashevi длина яйцевой капсулы достигает 122–130 мм, а ширина – 55–58 мм. У более мелких B. fedorovi и B. tzinovskii длина яйцевой капсулы составляет соответственно 75–86 и 85 мм, а ширина – 40–48 и 49 мм. У всех скатов батибентального ихтиоцена в капсуле развивается только один эмбрион. Соотношение полов эмбрионов из яйцевых капсул приблизительно 1 : 1, как и в естественных популяциях, в которых самки преобладают лишь в старших возрастных группах (Долганов, 1998).
Известные к настоящему времени максимальные размеры и возраст составляют для B. fedorovi 69.9 см (7 лет) у самцов и 73.3 см (9 лет) у самок, для B. andriashevi 120.0 см (14 лет) у самцов и 141.0 см (16 лет) у самок, для B. tzinovskii соответственно 74.3 см (8 лет) и 71.4 см (8 лет). Судя по появлению эректильных колючек и развитию гонад, половозрелыми самцы B. fedorovi становятся при длине 56.3–61.2 см в возрасте 4–5 лет, самки – при длине 62.0–64.8 см в возрасте 5–6 лет; самцы B. andriashevi – при длине около 105 см (10 лет), а самцы B. tzinovskii – при длине около 68 см в возрасте 6 лет.
Скаты батибентального ихтиоцена обладают сходными морфологическими особенностями строения пищеварительной системы. Характерной чертой является большой нижний рот с выдвижными челюстями, при раскрытии которого объем ротовой полости значительно увеличивается. Многочисленные мелкие зубы, расположенные на челюстях, облегчают удержание захваченной добычи. Коронки зубов скатов представляют собой сплющенные сверху овалы с острой конусовидной вершиной, направленной назад. Имеются хорошо выраженные возрастные различия в форме зубов: у молодых скатов вершина коронки маленькая, у взрослых особей она намного крупнее и острее. Увеличение вершины коронки с возрастом функционально связано с переходом на питание более крупным бентосом и рыбой.
По способу питания B. fedorovi является бентоихтиофагом. Этот вид питается преимущественно креветками (55.3%), амфиподами (8.9%), молодью крабов (7.1%), мелкой рыбой (светящимися анчоусами – 12.2%) и др. (7.4%). Второстепенной пищей являются головоногие моллюски (кальмары и осьминоги – 7.0%) и полихеты (2.1%). Несмотря на небольшое количество исследованных желудков B. andriashevi и B. tzinovskii, по-видимому, они тоже являются бентоихтиофагами, так как в их желудках приблизительно в равной пропорции обнаружены креветки и рыбы.
Список литературы
Долганов В.Н. Новые виды скатов семейства Rajidae из северо-западной части Тихого океана // Вопр. ихтиологии. 1985. Т. 25. № 3. С. 415–425.
Долганов В.Н. Размножение скатов семейства Rajidae дальневосточных морей России // Изв. ТИНРО. 1998. Т. 124. С. 495–498.
Долганов В.Н. Географическое и батиметрическое распространение скатов семейства Rajidae в дальневосточных морях России и сопредельных водах // Вопр. ихтиологии. 1999. Т. 39. № 3. С. 428–430.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. М.: Наука. 1974. 254 с.
Парин Н.В. Рыбы открытого океана. М.: Наука. 1988. 272 с.
Шунтов В.П. Биология дальневосточных морей России. Владивосток: ТИНРО-центр. 2001. Т. 1. 580 с.
Ishiyama R. Studies on the rajid fishes (Rajidae) found in the waters around Japan // J. Shimonoseki Coll. Fish. 1958. V. 7. № 2, 3. P. 193–394.
McEachran J.D., Compagno L.J.V. Interrelationships of and within Breviraja based on anatomical structures (Pisces, Rajoidei) // Bull. Mar. Sci. 1982. V. 32. № 2. P. 399–425.
Stehmann M. Vergleichend morphologische und anatomische Untersuchungen zur Neuordnung der Systematik der nordostatlantischen Rajidae (Chondrichthyes, Batoidei) // Arch. Fischereiwiss. 1970. V. 21. № 2. P. 73–163.
Дополнительные материалы отсутствуют.