

Биология моря, 2021, T. 47, № 2, стр. 95-107
Состав, структура и распределение фауны донных остракод (Ostracoda: Myodocopida) в Субантарктике
В. Г. Чавтур 1, А. Г. Башманов 1, *
1 Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: b0000@list.ru
Поступила в редакцию 05.03.2020
После доработки 21.09.2020
Принята к публикации 29.09.2020
Аннотация
Проведен анализ литературных данных по фауне донных остракод Myodocopida Субантарктики, включающей 97 видов. Исследованная фауна характеризуется высокой степенью эндемизма, но его низким рангом. Основу фауны составляют виды широко распространенных родов. По числу видов и родов лидируют остракоды семейств Cylindroleberididae и Cypridinidae. С увеличением глубины видовое богатство остракод и количество их родов уменьшаются, наблюдается перестройка таксономической и батиметрической структур. Сходство субантарктической и антарктической фаун Myodocopida проявляется в небольшом разнообразии видов, в высокой степени и низком ранге эндемизма, в соотношении количества видов и родов в семействах. Различие фаун связано со значительной изоляцией, с характером вертикального распределения, а также с таксономической структурой миодокопид. Субантарктическая фауна Myodocopida изолирована от бореальной, превышает ее по числу видов и родов, а также сопоставима с ней по числу эндемичных видов. Обе фауны характеризуются высокой степенью, но низким уровнем эндемизма. Видовое богатство данных фаун определяют разные семейства. С увеличением глубины количество видов миодокопид в обеих фаунах уменьшается, и ниже 5500 м миодокопиды не проникают.
Первые сведения о донных остракодах отряда Myodocopida в водах Субантарктической структуры появились в XIX столетии (Baird, 1850; Thomson, 1879; Brady, 1898). В современной мировой литературе из 42 публикаций по миодокопидам Южного океана в 21 публикации содержатся сведения о донных остракодах его умеренно-холодных вод (табл. 1). Изученность Myodocopida в разных океанических секторах Субантарктики неравномерна. Так, для тихоокеанского сектора имеется 17 публикаций, для атлантического – 6, а для индоокеанского – всего 4 публикации.
Таблица 1.
Использованный в работе материал
| № | Публикации | Район (сектор) субантарктической зоны | ||
|---|---|---|---|---|
| тихоокеанский | индоокеанский | атлантический | ||
| 1 | Baird, 1850 | + | − | − |
| 2 | Thomson, 1879 | + | − | − |
| 3 | Brady, 1880 | + | + | − |
| 4 | Brady, 1898 | + | − | − |
| 5 | Stebbing, 1901 | − | + | + |
| 6 | Brady, 1902 | ? + | − | − |
| 7 | Müller, 1906 | + | − | − |
| 8 | Skogsberg, 1920 | + | − | + |
| 9 | Poulsen, 1962 | + | − | − |
| 10 | Poulsen, 1965 | + | − | − |
| 11 | Hartmann-Shröder, Hartmann, 1965 | + | − | + |
| 12 | Kornicker, 1969 | − | + | + |
| 13 | Moguilevsky, Ramires, 1970 | − | − | + |
| 14 | Kornicker, 1975 | + | + | + |
| 15 | Kornicker, 1979 | + | − | − |
| 16 | Kornicker, 1994 | + | − | − |
| 17 | Kornicker, 1995 | + | − | − |
| 18 | Kornicker, Poore, 1996 | + | − | − |
| 19 | Kornicker, McKenzie, 1976 | − | + | − |
| 20 | Karanovic, 2010 | + | − | − |
| 21 | Karanovic, Lörz, 2012 | + | − | − |
Настоящая работа является продолжением исследования фауны донных Myodocopida Южного океана. Ранее Чавтур (2016) на основе литературных и оригинальных данных проанализировал их фауну в водах антарктической зоны (рис. 1). Изученность этих остракод в субантарктических водах (нотальной зоне) ограничена описанием отдельных видов и указанием районов их обнаружения (табл. 1). Особенное место занимает классический двухтомный труд американского зоолога Корникера (Kornicker, 1975) “Antarctic Ostracoda (Myodocopina)”. В этой работе приведено описание большинства миодокопид Антарктики и Субантарктики, кратко обсуждается характер их широтного расселения вдоль шельфа Аргентины и Чили, дана общая для Южного океана схема вертикального распределения миодокопид, показана зависимость количества омматидиев в латеральных глазах остракод от глубины обитания и количества яиц у самок от размеров их раковины. Таким образом, представленные в литературе данные позволяют провести анализ структуры и распределения фауны Myodocopida для умеренно-холодных вод в Южном полушарии в целом, а также сопоставить субантарктическую фауну с антарктической в Южном полушарии и с бореальной фауной миодокопид в Северном полушарии.

Работа основана на литературных данных (табл. 1); материал по миодокопидам, собранный российскими исследователями во время экспедиций в Южном океане, ограничен антарктической зоной (рис. 1). В статье приняты следующие границы Субантарктики: южная граница соответствует положению антарктической дивергенции (примерно до 65° ю.ш.) (Северный…., 1985), а положение северной границы заимствовано из опубликованных работ (Лаппо, Лебедев, 2005; Vierros et al., 2009). Следует отметить, что субантарктическая зона несколько выходит за границы Южного океана у южных берегов Австралии, Африки и Южной Америки (Физическая география, 1980). В зарубежной литературе (см., например: Kornicker, 1975) нижним пределом батиальной зоны (материкового свала) определена глубина 2000 м, тогда как в схеме вертикальной зональности бентали, разработанной российскими учеными (Беляев и др., 1973), она простирается до глубины 3000 м. Мы следуем этой схеме.
СРАВНЕНИЕ СУБАНТАРКТИЧЕСКОЙ И АНТАРКТИЧЕСКОЙ ФАУН ОСТРАКОД
Фауна донных остракод отряда Myodocopida Субантарктики представлена 97 видами в составе 34 родов и 5 семейств (табл. 2); это всего 10% от всей мировой фауны данного отряда. Наблюдается различие в количестве видов остракод по океаническим секторам: для тихоокеанского сектора известно 68 видов (30 родов), для атлантического – 29 видов (16 родов) и для индоокеанского – всего 7 видов (3 рода). Такое соотношение исключает возможность характеристики фаун отдельных секторов, поэтому мы анализируем фауну в целом для всей Субантарктики. Несмотря на относительно небольшое разнообразие фауны, ее специфичность здесь достаточно высокая. Степень эндемизма достигает 74%. По числу видов субантарктическая фауна несколько превышает антарктическую (70 видов), но уступает ей по количеству эндемов (83%) (Чавтур, 2016). Данные фауны содержат всего 16 общих видов, что свидетельствует о значительной разобщенности фаун, хотя они занимают соседние пространства. Если степень эндемизма фаун высокая, то ранг низкий. Лишь один род Dolasterope Poulsen, 1965, описанный по единственному виду D. johanseni Poulsen, 1965 из прибрежных вод Новой Зеландии, по-видимому, является эндемичным для Субантарктики, а в целом для Южного океана в этом качестве зарегистрирован только род Anarthron Kornicker, 1975.
Таблица 2.
Видовой состав остракод отряда Myodocopida и их встречаемость в водах Субантарктики
| № | Таксон | Район (сектор) | **Биогеографическая характеристика | ***Батиметрическая характеристика | Глубина, м | ||
|---|---|---|---|---|---|---|---|
| атлантический | индоокеанский | *тихоокеанский | |||||
| Семейство Cypridinidae | |||||||
| 1 | Bathyvargula walfordi Poulsen, 1962 | 14 | ?Н | С-Б | 10, 610 | ||
| 2 | Cypridinodes concentrica Kornicker, 1979 | 15 | ?Н | Б | 384 | ||
| 3 | Cypridinodes reticulata Poulsen, 1962 | 9 | ?Н | С-Б | 10, 610 | ||
| 4 | Doloria (Dolorietta) pectinata Skogsberg, 1920 | 8, 14 | 14 | Н | С-Б | 21–439 | |
| 5 | Doloria (Dolorietta) levinsoni Kornicker, 1975 | 14 | Н-Ан | Б | 424–857, 2453 | ||
| 6 | Doloria (Dolorietta) septenaria Kornicker, 1975 | 14 | Н | Б | 384–494 | ||
| 7 | Doloria (Dolorietta) mawsoni Kornicker, 1975 | 14 | Н | С | 69 | ||
| 8 | Hadacypridina bruuni Poulsen, 1962 | 9, 14 | юСбтр-Н | А | 5340 | ||
| 9 | Metavargula adinothrix Kornicker, 1975 | 14 | 14 | Н-Ан | Б-А | 1709–1803 | |
| 10 | Metavargula mazeri Kornicker, 1979 | 15 | ?Н | Б | 384 | ||
| 11 | Metavargula spadix Kornicker, 1996 | 18 | юСбтр-Н | Б | 1264 | ||
| 12 | Metavargula currax Kornicker, 1996 | 18 | ?Н | Б | 720 | ||
| 13 | Rheina relax Kornicker, 1996 | 18 | юСбтр-Н | С-Б | 124–478 | ||
| 14 | Paradoloria pugnax Kornicker, 1996 | 18 | ?Н | Б | 720 | ||
| 15 | Paradoloria species A (по Kornicker, 1996) | 18 | ?Н | Б | 720 | ||
| 16 | Pterocypridina tressleri Kornicker, 1996 | 18 | юСбтр-Н | Б | 800 | ||
| 17 | Vargula antarctica (Müller, 1908) | 14 | Ан | Б-А | 300 | ||
| 18 | Vargula hamata Kornicker, 1975 | 14 | Н-Ан | Б | 429–1212 | ||
| 19 | Vargula subantarctica Kornicker, 1975 | 14 | Н | С-Б | 119–572 | ||
| 20 | Vargula ? danae (Brady, 1880) | 3 | Н | ? Б | 219 | ||
| 21 | Vargula dentata Kornicker, 1975 | 14 | Н | Б | 429–1978 | ||
| 22 | Vargula lusca Kornicker, 1975 | 14 | Н | Б | 910–915 | ||
| 23 | Vargula ascensus Kornicker, 1979 | 15 | ?Н | Б | 549 | ||
| 24 | Vargula stranx Kornicker, 1994 | 16 | юСбтр-Н | Б | 720 | ||
| 25 | Vargula rapax Kornicker, 1994 | 16 | Н | Б | 400 | ||
| 26 | Rugosidoloria serrata Kornicker, 1975 | 14 | ?Н | С | 44 | ||
| 27 | Isocypridina quatuorsetae Kornicker, 1975 | 14 | Н-Ан | Б-А | 2818–3197 | ||
| Семейство Philomedidae | |||||||
| 28 | Philomedes eugeniae Skogsberg, 1920 | 8 | 8 | Н | С-Б | 7−270 | |
| 29 | Philomedes tetrathrix Kornicker, 1975 | 14 | Н-Ан | Б-А | 516, 1276, 3219 | ||
| 30 | Philomedes subantarctica Kornicker, 1975 | 14 | Н | С-Б | 129–572 | ||
| 31 | Philomedes minys Kornicker, 1975 | 14 | Н | С | 31−108 | ||
| 32 | Philomedes ramus Kornicker, 1975 | 14 | Н | А | 3219 | ||
| 33 | Philomedes lofthousae Kornicker, 1975 | 14 | Н-Ан | С | 25–130 | ||
| 34 | Philomedes cubitum Kornicker, 1975 | 14, 19 | 14 | Н | С | 20–40 | |
| 35 | Philomedes fortax Kornicker, 1994 | 14 | ? Н | Б | 720 | ||
| 36 | Pleoshisma agilis (Thomson, 1879) | 2, 14 | юСбтр-Н | С | Поверхность | ||
| 37 | Scleroconcha solox Kornicker, 1995 | 17 | юСбтр-Н | Б | 920–1130 | ||
| 38 | Scleroconcha arcuata Poulsen, 1962 | 14 | ? Н | С | 1.8–146 | ||
| 39 | Scleroconcha flexilis (Brady, 1898) | 4 | Н | С | 1.8–9.1 | ||
| 40 | Scleroconcha sculpta (Brady, 1898) | 4 | Н | С | Поверхность | ||
| 41 | Scleroconcha frons Kornicker, 1975 | 14 | ?Н | Б | 957–1263 | ||
| 42 | Scleroconcha wolffi Kornicker, 1975 | 14 | Н | С | 11 | ||
| 43 | Anarthron reticulata (Hartmann, 1965) | 11, 14 | С-Б | 110–295 | |||
| 44 | Anarthron chilensis (Hartmann, 1965) | 11, 14 | Н | С | 100–112 | ||
| 45 | Anarthron ditrix Kornicker, 1975 | 14 | 14 | С-Б | 71–119, 1146−1201 | ||
| 46 | Anarthron pholion Kornicker, 1975 | 14 | Н | С-Б | 82–284 | ||
| 47 | Anarthron evexum Kornicker, 1975 | 14 | Н | С-Б | 79–284 | ||
| 48 | Euphilomedes tasmanicus Karanovic, 2010 | 20 | ?Н | С | 13 | ||
| 49 | Igene walleni Kornicker, 1975 | 14 | Н-Ан | А | 4303 | ||
| Семейство Cylindrolaberididae | |||||||
| 50 | Empoulsenia pentathrix (Kornicker, 1971) | 14 | Н | Б-А | 278–280, 1812–2145 | ||
| 51 | Archasterope bulla Kornicker, 1975 | 14 | Н-Ан | Б | 429, 1212 | ||
| 52 | Archasterope efficas Kornicker, 1996 | 17 | юСбтр-Н | Б | 400 | ||
| 53 | Archasterope apex Kornicker, 1996 | 17 | ?Н | Б | 500 | ||
| 54 | Archasterope verax Kornicker, 1996 | 17 | юСбтр-Н | Б | 1264 | ||
| 55 | Diasterope grisea (Brady, 1898) | 4, 15 | Н | С | 14 | ||
| 56 | Dolasterope Johanseni Poulsen, 1965 | 10 | Н | С | 26 | ||
| 57 | Parasterope longiseta (Skogsberg, 1920) | 8, 14 | 14 | Н-Ан | С | 12–108 | |
| 58 | Parasterope quadrata (Brady, 1898) | 4 | ?Н | С | 1.8−9.1 | ||
| 59 | Parasterope quadrata? (Brady, 1898) | 4 | ?Н | С | Поверхность | ||
| 60 | Parasterope pseudoquadrata (Hartmann, 1965) | 11 | ?Н | С | 12 | ||
| 61 | Parasterope crinita Kornicker, 1975 | 14 | ?Н | С | 51 | ||
| 62 | Parasterope anommata Kornicker, 1975 | 14 | Н | Б | 424–578 | ||
| 63 | Parasterope styx Kornicker, 1975 | 14 | Н-Ан | А | 4303 | ||
| 64 | Parasterope prolixa Kornicker, 1975 | 14 | Н | С | 20–40 | ||
| 65 | Parasterope micrommata Kornicker, 1975 | 14 | Н | С | 86–124 | ||
| 66 | Synasterope dimorpha (Hartmann, 1965) | 11, 14 | Н | С-Б | 70–210, 2470–2657 | ||
| 67 | Synasterope empoulseni Kornicker, 1975 | 14 | юСбтр-Н | С | 10–15 | ||
| 68 | Synasterope arnaudi Kornicker, 1975 | 14 | Н | C | 50 | ||
| 69 | Synasterope mystax Kornicker, 1975 | 14 | Н-Ан | Б-А | 4006 | ||
| 70 | Synasterope brachythrix Kornicker, 1975 | 14 | Н | Б | 214–460 | ||
| 71 | Skogsbergiella spinifera (Skogsberg, 1920) | 14 | Н-Ан | С-Б | 70–119 | ||
| 72 | Skogsbergiella macrothrix Kornicker, 1975 | 14 | 14 | Н | С-Б | 70–1201 | |
| 73 | Skogsbergiella plocus Kornicker, 1975 | 14 | Н | С | 112 | ||
| 74 | Skogsbergiella pax Kornicker, 1975 | 14 | ?Н | Б | 1475 | ||
| 75 | Homasterope micra Kornicker, 1975 | 14 | Н | С | 112 | ||
| 76 | Bathyleberis grossmani Kornicker, 1975 | 14 | Н-Ан | А | 4303 | ||
| 77 | Bathyleberis monothrix Kornicker, 1975 | 14 | Н | С | 71−82 | ||
| 78 | Bathyleberis okulata Kornicker, 1975 | 14 | ?Н | С | 117 | ||
| 79 | Asteropella rotundicostata (Hartmann, 1965) | 11 | Н | С | 12 | ||
| 80 | Asteropella species A (по: Kornicker, 1975) | 14 | Н | С | 44–57 | ||
| 81 | Cycloleberis zealandica (Baird, 1850) | 1 | ?Н | С | 14 | ||
| 82 | Cycloleberis ovulum (Brady, 1898) | 4 | Н | ||||
| 83 | Microasteropteron chathamensis Karanovic, Lörz, 2012 | 21 | Н | Б | 1023–1103 | ||
| Семейство Sarsiellidae | |||||||
| 84 | Eusarsiella bex Kornicker, 1994 | 16 | Н | Б | 720 | ||
| 85 | Eusarsiella iayx Kornicker, 1994 | 16 | Н | Б | 400 | ||
| 86 | Spinacopia bisetula Kornicker, 1969 | 12 | Б | ||||
| 87 | Spinacopia variabilis Kornicker, 1969 | 12 | Н | С-Б | 185, 1475 | ||
| 88 | Cimbicopia zealandica (Poulsen, 1965) | 10 | Н | С | 10–15 | ||
| 89 | Cimbicopia hanseni (Brady, 1898) | 4 | Н | С | 1.8–12 | ||
| 90 | Cimbicopia hispida (Brady, 1898) | 4 | Н | С | 1.8–12 | ||
| 91 | Cimbicopia brevicosta Kornicker, 1975 | 14 | Н | С | 1.8–11 | ||
| Семейство Rutidermatidae | |||||||
| 92 | Rutiderma herdhartmanni Kornicker, 1975 | 14 | Н | С | 12 | ||
| 93 | Rutiderma ovata Kornicker, 1975 | 14 | ?Н | С | 176–192 | ||
| 94 | Rutiderma species A (по: Kornicker, 1975) | 14 | Н | С | 12 | ||
| 95 | Rutiderma species B (по: Kornicker, 1975) | 14 | Н | С | 1–97 | ||
| 96 | Scleraner chacaoi (Hartmann, 1965) | 11 | Н | С | 21.3−240 | ||
| 97 | Adelta theta Kornicker, 1975 | 14 | юСбтр-Н | C | 15 | ||
Примечание. *Номер соответствует порядковому номеру публикации в табл. 1; **Н – нотальный, Н-Ан – нотально-антарктический, юСбтр-Н – южносубтропическо-нотальный, Ан – антарктический; ***С – сублиторальный, Б – батиальный, А – абиссальный, С-Б – сублиторально-батиальный, Б-А – батиально-абиссальный.
Говорить о биогеографической структуре субантарктической фауны Myodocopida можно лишь приближенно, поскольку некоторые виды известны по единичным сборам (в табл. 2 они указаны с вопросительным знаком). Чаще других в субантарктических водах встречаются Doloria pectinata, Vargula hamata, Philomedes assimilis, Anarthron dithrix, Parasterope longiseta, Skogsbergiella macrothrix и Empoulsenia pentathrix.
В пределах Субантарктики отмечено 34 рода донных остракод отряда Myodocopida, что составляет 37% от их мировой фауны (92 рода) и в 2 раза больше, чем известно для Антарктики (17 родов) (Чавтур, 2016). Основу в Субантарктике, как и в умеренно-холодных и холодных водах, составляют такие широко распространенные роды (65%), как Philomedes Lilljeborg, 1853; Vargula Skogsberg, 1920; Scleroconcha Skogsberg, 1920; Doloria Skogsberg, 1920 и Bathyleberis Kornicker, 1975. По количеству видов доминирует семейство Cylindroleberididae (35%), затем следуют семейства Cypridinidae (28%), Philomedidae (23%), Sarsiellidae (9%) и Rutidermatidae (5%) (рис. 2a). Примерно такое же соотношение отмечено и для антарктической фауны, однако в полярных водах доля видов в семействах Cypridinidae и Philomedidae равная, а остракоды семейства Rutidermatidae отсутствуют (Чавтур, 2016). В Субантарктике по количеству родов, как и в Антарктике, лидируют семейства Cylindroleberididae (32%) и Cypridinidae (33%), а количество родов в остальных семействах не отличается от такового в Антарктике (рис. 2б).
Рис. 2.
Относительное содержание видов (а) и родов (б) в составе субантарктической фауны донных остракод отряда Myodocopida.

При увеличении глубины (рис. 3) в субантарктической зоне изменяются количество видов, состав и структура фауны Myodocopida. Максимум видового богатства миодокопид отмечен в верхнем отделе шельфа (0–50 м), которое формируется видами Bathyvargula walfordi, Cypridinodes reticulata, Philomedes eugeniae, P. minis, P. loftousae, P. cubitum, P. agilis, Scleroconcha arcuata, S. flexilis, S. sculpta, S. wolffi, Euphilomedes tasmanicus, Diasteroe grisea, Dolasterope johanseni, Parasterope longiseta, P. quadrata, P. pseudoquadrata, P. prolix, Synasterope empoulseni, Asteropella rotundicostata, A. species A (по: Kornicker, 1975), Cycloleberis zealandica, Cymbicopia zealandica, C. hanseni, C. hispida, C. brevicosta, Rutiderma herdhartmanni, R. species A (по: Kornicker, 1975), R. species B (по: Kornicker, 1975), Scleraner chacaoi и Adelta theta.
Рис. 3.
Вертикальное распределение донных остракод отряда Myodocopida в водах Субантарктики (цифры под колонками соответствуют порядковым номерам видов в табл. 2).

С глубиной разнообразие миодокопид уменьшается, но в верхней части материкового свала (200–500 м) оно вновь незначительно возрастает за счет появления здесь глубоководных видов: Cypridinodes concentric, Doloria (Dolorietta) levensoni, D. (D.) septenaria, Metavargula mazeri, Rheina relax, Vargula antarctica, V. hamata, V. subarcuata, V. danae, V. dentata, V. rapax, Anarthron reticulate, A. chilensis, Archasterope bulla, A. pentathrix, A. efficas, A. apex, Parasterope anommata, Synasterope dimorpha, S. brachythrix, Eusarsiella iayx и Spinacopia variabilis. А глубже 1000 м и до абиссальных глубин наблюдается резкая убыль миодокопид. Ниже глубины 5500 м остракоды отряда Myodocopida не обнаружены. Такое распределение миодокопид заметно отличается от их распределения в антарктических водах, где с увеличением глубины число видов, напротив, возрастает, достигая максимума в верхней батиали на глубинах 200–500 м (Чавтур, 2016).
Характер вертикального распределения остракод отдельных семейств различается (рис. 4). Так, количество видов семейств Philomedidae и Cylindroleberididae с увеличением глубины монотонно убывает от верхней сублиторали до абиссали (до 4500 м). Иначе распределены остракоды семейства Cypridinidae: в пределах шельфа отмечено всего 6 видов, тогда как в верхней части материкового свала на глубинах 200–500 и 500–1000 м их количество наибольшее – соответственно 14 и 13 видов (рис. 3). Глубже разнообразие остракод этого семейства резко уменьшается, и ниже 3500 м (на глубине 5340 м) обнаружен лишь один вид Hadacypridina bruuni Poulsen, 1962. Семейства Sarsiellidae и Rutidermatidae в фауне Субантарктики представлены небольшим количеством видов, распространение которых ограничено шельфовой зоной и верхней частью материкового свала до глубины 1500–2000 м. В антарктических водах распределение миодокопид характеризуется тем, что видовое богатство семейств Philomedidae и Cylindroleberididae с увеличением глубины до 200–500 м возрастает, а глубже заметно снижается (Чавтур, 2016).
Рис. 4.
Вертикальное распределение миодокопид отдельных семейств и в целом их фауны в водах Субантарктики.
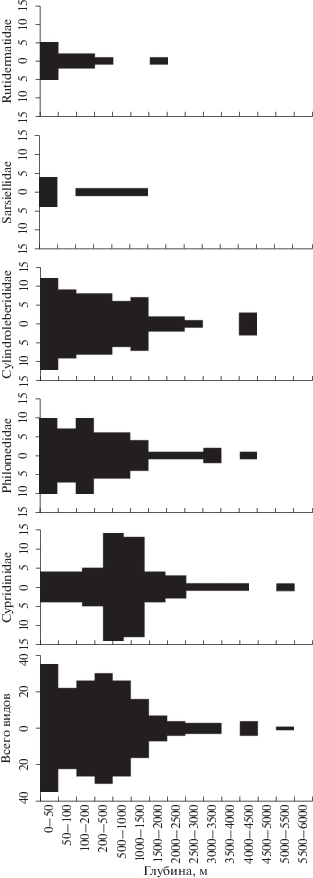
С увеличением глубины изменяется и батиметрическая структура фауны Myodocopida (табл. 3). В прибрежной зоне (до глубины 50 м) зарегистрирован максимум сублиторальных видов, количество которых с глубиной уменьшается, и за нижнюю границу шельфа они почти не проникают. Сублиторально-батиальных видов с увеличением глубины, напротив, становится больше, особенно у нижней границы шельфа и в верхней батиали на глубинах 200–500 м, ниже их число убывает. Максимум батиальных видов приходится на глубины 500–1000 м. Батиально-абиссальная и абиссальная группы содержат небольшое число остракод. Примерно такие же изменения отмечены в структуре антарктической фауны Myodocopida (Чавтур, 2016).
Таблица 3.
Изменение с глубиной числа видов остракод отряда Myodocopida в водах Субантарктики
| Глубина, м | Виды | |||||
|---|---|---|---|---|---|---|
| сублиторальные | сублиторально-батиальные | батиальные | батиально-абиссальные | абиссальные | всего | |
| 0–50 | 31 | 4 | – | – | – | 35 |
| 50–100 | 12 | 10 | – | – | – | 22 |
| 100–200 | 11 | 14 | 1 | – | – | 26 |
| 200–500 | 1 | 14 | 14 | 2 | – | 31 |
| 500–1000 | – | 8 | 17 | 2 | – | 27 |
| 1000–1500 | – | 4 | 10 | 2 | – | 16 |
| 1500–2000 | – | 1 | 3 | 3 | – | 7 |
| 2000–2500 | – | 1 | 1 | 2 | – | 4 |
| 2500–3000 | – | 1 | – | 2 | – | 3 |
| 3000–3500 | – | – | – | 2 | 1 | 3 |
| 3500–4000 | – | – | – | 0 | 0 | 0 |
| 4000–4500 | – | – | – | 1 | 3 | 4 |
| 4500–5000 | – | – | – | 0 | 0 | 0 |
| 5000–5500 | – | – | – | 0 | 1 | 1 |
С глубиной изменяется и число родов (рис. 5). В данном случае также наблюдается два количественных максимума – в верхней сублиторали (0–50 м) и в верхней батиали (200–500 и 500–1000 м), тогда как в антарктических водах максимум числа родов один и он приходится на глубины 200–500 м (Чавтур, 2016). В Субантарктике в шельфовой зоне зарегистрировано больше родов (24), чем в батиали (22) и абиссали (6), а в Антарктике их больше в батиали (16), но меньше на шельфе (11) и в абиссали (11), что обусловлено чрезвычайно суровыми температурными условиями в верхних слоях полярных вод (Чавтур, 2016). Наиболее обычными на шельфе являются представители родов Philomedes, Scleroconcha, Parasterope и Synasterope, в батиали – Vargula, Doloria, Metavargula, Philomedes, Scleroconcha и Archasterope, а в зоне абиссальных глубин зарегистрированы лишь единичные особи родов Philomedes, Syphonostra, Bathyleberis, Isocypridina, Igene и Hadacypridina, из них три последних рода отмечены только в пределах этой зоны (рис. 5). В субантарктических водах наиболее эврибатными среди миодокопид являются роды Philomedes, Syphonostra и Bathyleberis, вертикальный диапазон обитания которых простирается от прибрежья до абиссальных глубин (рис. 5).

СРАВНЕНИЕ СУБАНТАРКТИЧЕСКОЙ И БОРЕАЛЬНОЙ ФАУН ОСТРАКОД
При сравнении фаун Myodocopida в умеренно-холодных водах в Южном и Северном полушариях предварительно отметим, что широтный диапазон, площади шельфа и материкового свала бореальной зоны значительно больше, тогда как почти вся акватория нотальной зоны расположена над абиссальными глубинами. Кроме этого, континенты в Северном полушарии создают полную изоляцию соответствующих вод в Атлантике и Пацифике, чего не наблюдается в Южном полушарии. Это безусловно определяет степень разнообразия и специфичности их фаун.
Субантарктическая и бореальная фауны миодокопид изолированы на видовом уровне (Чавтур, 1992). Они не содержат даже викариантных видов. Число видов и родов в субантарктической фауне немного больше, чем в бореальной: соответственно 97 и 90 видов, 34 и 28 родов, из них 17 родов общие (Чавтур, 1992). По количеству эндемичных видов данные фауны сопоставимы: в субантарктических водах их доля составляет 74%, а в бореальных – 75%. При этом в бореальной части Тихого океана эндемизм достигает 87%, а в Атлантике составляет всего 59% (Чавтур, 1992). Если в сравниваемых фаунах степень эндемизма высокая, то его ранг чрезвычайно низкий: в бореальных водах он не превышает видового, а в субантарктических водах известен лишь один эндемичный монотипический род Dolasterope, найденный вблизи субтропической зоны (Новая Зеландия, 44°34′ ю.ш. – 167°18′ в.д., глубина 136 м).
Наблюдаются различия в таксономической структуре сравниваемых фаун. По числу видов в субантарктической фауне лидируют остракоды семейств Cylindroleberididae и Cypridinidae, а в бореальной − семейств Philomedidae и Cylindroleberididae. Кроме этого, в составе бореальной фауны отсутствуют миодокопиды семейства Rutidermatidae. Характер вертикального распределения обеих фаун в основном сходный: с увеличением глубины число видов Myodocopida уменьшается и глубже 5500 м они не проникают (Igene curtus Chavtur, 1983–5240 м, район Курило-Камчатской впадины; Hadacypridina bruuni Poulsen, 1962–5340 м, Субантарктика). Наличие в верхней части шельфа субантарктической зоны большой доли сублиторальных видов свидетельствует о принадлежности этого района (южной оконечности Южной Америки, о-ва Тасмания и Южного острова Новой Зеландии) к низкосубантарктической подзоне субантарктической зоны. Это же отмечено и для низкобореальной подзоны бореальной зоны, где по числу видов лидируют сублиторальные остракоды, в отличие от высокобореальной подзоны, где они встречаются лишь в нижних отделах шельфа.
ЗАКЛЮЧЕНИЕ
Таким образом, фауна миодокопид Субантарктики представлена 97 видами, что составляет 10% от их мировой фауны. Она характеризуется высокой степенью, но низким уровнем эндемизма. Ее основу составляют виды широко распространенных родов. По числу видов и родов лидируют остракоды семейств Cylindroleberididae и Cypridinidae. Максимумы видового богатства и количества родов зарегистрированы в прибрежной зоне (0–50 м). С увеличением глубины их показатели снижаются, однако в верхней части материкового свала (200–500 м) образуется второй, но меньший максимум. Ниже глубины 1000 м число видов и родов резко снижается, и глубже 5500 м миодокопиды не проникают. С изменением глубины наблюдается перестройка таксономической структуры фауны миодокопид на уровне родов и семейств, а батиметрической структуры на уровне видов.
При сравнении с антарктической фауной Myodocopida сходство наблюдается в относительно небольшом разнообразии видов, в высокой степени и низком ранге эндемизма, а также в соотношении количества видов и родов в семействах. Различия связаны со значительной изоляцией фаун миодокопид в Субантарктике и Антарктике, имеющих лишь 10 и 50% общих видов и родов соответственно. С увеличением глубины видовое богатство и количество родов в субантарктических водах уменьшаются, а в антарктических водах увеличиваются, достигая максимума в верхней части батиали. Различия в вертикальном распределении остракод проявляются также на примере отдельных семейств. Так, в Субантарктике число видов в семействах Cylindroleberididae и Philomedidae с увеличением глубины уменьшается, а в семействе Cypridinidae – увеличивается, достигая максимума в верхней батиали, тогда как в Антарктике видовое богатство всех семейств с глубиной увеличивается, достигая максимальных значений у нижней границы шельфа и в верхней части материкового свала.
При сравнении нотальной и бореальной фаун Myodocopida следует учитывать их широтную разобщенность, а также существенные различия в размерах площадей шельфа и верхней батиали в пределах умеренно-холодных вод Южного и Северного полушарий, что влияет на степень разнообразия и специфичности их фаун. Эти фауны абсолютно изолированы на видовом уровне и значительно на родовом. По числу видов и родов субантарктическая фауна немного превышает бореальную и сопоставима с ней по числу эндемичных видов. Обе фауны характеризуются высокой степенью, но низким рангом эндемизма. По видовому богатству в сравниваемых фаунах преимущество имеют разные семейства. Характер вертикального распределения в основном сходный: с увеличением глубины число видов Myodocopida уменьшается, и ниже 5500 м они не проникают.
Список литературы
Беляев Г.М., Виноградова Н.Г., Левенштейн Р.Я. и др. Закономерности распределения глубоководной донной фауны в свете развития идеи о биологической структуре океана // Океанология. 1973. Т. 13, вып. 1. С. 149–157.
Лаппо С.С., Лебедев В.Л. Географическая зональность в водах Мирового океана // Горизонты географии. К 100-летию К.К. Маркова. М.: Географический факультет МГУ. 2005. С. 229–237.
Северный Ледовитый и Южный океаны. Л.: Наука. 1985. С. 271–458. (Серия “География Мирового океана”).
Физическая география Мирового океана. Л.: Наука. 1980. С. 313–316. (Серия “География Мирового океана”).
Чавтур В.Г. Состав, структура и распределение донной и пелагической фаун остракод надотряда Myodocopa умеренных и холодных вод Северного полушария. Владивосток: ИБМ ДВО РАН. 1992. 155 с.
Чавтур В.Г. Состав, структура и распределение фауны донных остракод (Ostracoda Myodocopida) в водах Антарктики // Биол. моря. 2016. Т. 42. № 6. С. 407–418.
Baird W. Description of several new species of Entomostraca // Proc. Zool. Soc. London. 1850. V. 18. P. 254–257.
Brady G.S. Report on the Ostracoda dredged by H.M.S. Challenger during the Years 1873–1876 // Report on the scientific results of the Voyage of the H.M.S. Challenger during the years 1873–76. Zoology. V. 1. 1880. London. P. 1–84.
Brady G.S. On new or imperfectly-known species of Ostracoda, chiefly from New Zealand // Trans. Zool. Soc. London. 1898. V. 14. № 8. P. 429–452.
Brady G.S. On new or imperfectly-known Ostracoda, chiefly from a Collection in the Zoological Museum, Copenhagen // Trans. Zool. Soc. London. 1902. V. 16. № 4. P. 179–210.
Hartmann-Schröder G., Hartmann G. Ostracoden des Sublitorals, in Zur kenntnis des sublitorals der chilenischen küste unter besonderer Berücksichtigung der Polychaeten und Ostracoden // Mitt. Hamburg. Zool. Mus. Inst. 1965. V. 62. P. 307–380.
Karanovic I. A new Euphilomedes Kornicker, 1967 (Myodocopida: Philomedidae) from Tasmania with a key to the species of the genus // Mar. Biodiversity. 2010. V. 40. № 3. P. 219–236.
Karanovic I., Lörz A.-N. A new subfamily and species of Ostracoda (Cylindroleberididae, Myodocopa) from the Chatham Rise (New Zealand) // N. Z. J. Zool. 2012. V. 39. № 4. P. 265–290.
Kornicker L.S. Morphology, ontogeny, and intraspecific variation of Spinacopia, a new genus of myodocopid ostracod (Sarsiellidae) // Smithson. Contrib. Zool. 1969. № 8. P. 1–50.
Kornicker L.S. Antarctic Ostracoda (Myodocopina). Pt. 1, 2 // Smithson. Contrib. Zool. 1975. № 163. https://doi.org/10.5479/si.00810282.163
Kornicker L.S. The marine fauna of New Zealand benthic Ostracoda (Suborder Myodocopina) // N. Z. Oceanogr. Inst. Memoir. 1979. V. 82. P. 5–58.
Kornicker L.S. Ostracoda (Myodocopina) of the SE Australian Continental Slope, Part 1 // Smithson. Contrib. Zool. 1994. V. 553. P. 1–200.
Kornicker L.S. Ostracoda (Myodocopina) of the SE Australian Continental Slope, Part 2 // Smithson. Contrib. Zool. 1995. V. 562. P. 1–97.
Kornicker L.S., McKenzie K.G. Redescription of Eurypylus petrosus Brady, 1869 and a key to the genera of Sarsiellidae (Myodocopina: Ostracoda) // Proc. Biol. Soc. Washington. 1976. V. 89. № 27. P. 347–352.
Kornicker L.S., Poore C.B. Ostracoda (Myodocopina) of the SE Australian Continental Slope, Part 3 // Smithson. Contrib. Zool. 1996. V. 573. P. 1−186. https://doi.org/10.5479/si.00810282.573
Moguilevsky A., Ramires F.C. Cycloleberis poulseni especie nueva (Ostracoda, Asteropidae) crustaceo bentonico del area de Mar del Plata, Argentina // Physis. 1970. V. 29. № 79. P. 461–471.
Müller G.W. Ostracoda from the Siboga Expedition // Siboda-Expeditie: Uitkomsten op Zoologisch, Botanisch, Oceanographisch en Geologisch Gebied verzameld in Nederlandsch Oost-Indië 1899−1900. 1906. V. 30. P. 1–40.
Poulsen E.M. Ostracoda−Myodocopa. Pt. I: Cypridiniformes – Cypridinidae, Dana Report Carlsberg Foundation. 1962. V. 57. 414 pp.
Poulsen E.M. Ostracoda−Myodocopa. Pt. II: Cypridiniformes – Rutidermatidae, Sarsiellidae and Asteropinae. Dana-Report. Copenhagen: Andr. Fred. Host & Son. 1965. V. 65. 484 p.
Thomson G.M. On the New Zealand Entomostraca // Trans. Proc. N. Z. Inst. 1879. V. 11. P. 251–263.
Skogsberg T. Studies on marine ostracods. Pt. 1 (Cypridinids, halocyprids and polycopids) // Zool. Bidr. Uppsala Suppl. 1920. V. 1. № 1. 784 p.
Stebbing T.R.R. Giant Ostracoda: old and new // Knowledge. 1901. V. 24. № 187. P. 100.
Vierros M., Cresswell I., Escobar B. et al. Global open oceans and deep seabed (GOODS): biogeographic classification, Paris: UNESCO-IOC 77; IOC Tech. Ser. 2009. № 84. P. 26.
Дополнительные материалы отсутствуют.