

Биология моря, 2021, T. 47, № 5, стр. 355-358
Морфология биссуса и прочность прикрепления к субстрату двустворчатого моллюска Arca boucardi Jousseaume, 1894 (Arcidae)
Н. И. Селин *
Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
* E-mail: nikselin@yandex.ru
Поступила в редакцию 15.03.2021
После доработки 21.04.2021
Принята к публикации 22.04.2021
Аннотация
В эксперименте наблюдали за поведением двустворчатого моллюска Arca boucardi Jousseaume, 1894 при повторном прикреплении к оргстеклу. Установлено, что после активного перемещения в течение 0.5–4 ч в поисках подходящего места моллюски начинали формировать биссус, выделяя прозрачный секрет. Через 9–10 ч новый биссус, принципиально не отличавшийся от старого, был сформирован. Показано, что биссус – это монолитный относительно эластичный тяж зеленого цвета, имеющий в дистальной части адгезивную подошву, а в проксимальной части полость с системой септ, которые служат для прикрепления мышц, отвечающих за работу биссуса. Полевые исследования прочности прикрепления моллюсков к субстрату показали, что в зависимости от размера животного она изменялась от нескольких сотен граммов до 23.5 кг. Данные обсуждаются с позиции функциональной морфологии и особенностей пространственного распределения A. boucardi в верхней сублиторали.
Двустворчатые моллюски, во взрослом состоянии ведущие прикрепленный образ жизни с помощью специального образования – биссуса, встречаются в ряде семейств, но наиболее обычны среди Mytilidae и Arcidae. Обширные натурные и экспериментальные наблюдения дают представление о морфофизиологических особенностях биссусного аппарата митилид, что позволяет объяснить процветание моллюсков этой группы в современных условиях. Для семейства Arcidae подобная информация отсутствует; сведения, встречающиеся в литературе, фрагментарны, порой противоречивы или вызывают сомнение. Например, указано, что биссус у Arca boucardi, как и у мидий, включает биссусные нити (Евсеев, Яковлев, 2006). Однако по мнению других исследователей биссус у этого вида имеет вид плоского тяжа, не разделенного на нити (Скарлато, 1981). Двустворчатый моллюск A. boucardi широко распространен от юга Охотского моря до о-ва Тайвань и является массовым в ряде биотопов прибрежных донных мелководий, где, по-видимому, играет важную роль (Скарлато и др., 1967; Скарлато, 1981). В связи с этим цель настоящей работы заключалась в изучении внешней морфологии и динамики образования биссуса у A. boucardi, а также в оценке прочности прикрепления моллюска к субстрату.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в апреле 2020 г. на биостанции “Восток” Национального научного центра морской биологии ДВО РАН в зал. Восток (зал. Петра Великого Японского моря). Образование биссуса изучали экспериментально у 40 взрослых особей с длиной раковины (L) 30–40 мм, которых после акклиматизации к температуре 15°С на 1 сут помещали в прозрачные пластиковые аквариумы с проточной морской водой. Из них 20 экз. тестировали без грунта, полагая, что таковым для животных станет оргстекло аквариума. Для оценки возможности образования моллюсками биссуса на мягком дне остальные 20 экз. были помещены в аквариумы с песком. Высота слоя песка − 10 см, размер частиц – менее 2 мм. Морфологические особенности организации биссуса изучали на нескольких препарированных моллюсках, используя световой бинокуляр. Прочность прикрепления животных оценивали под водой на глубине 3–6 м в месте их обитания в друзах модиолуса Modiolus kurilensis и мидии Грея Crenomytilus grayanus (Селин, 2018а). Определяли силу (F, кг), которую необходимо приложить для отрыва животного от субстрата, по аналогии с исследованиями прочности прикрепления митилид (Price, 1980; Селин, 2018б). В работе использовали пружинные динамометры разной мощности с ценой деления 10, 100 и 500 г.
РЕЗУЛЬТАТЫ
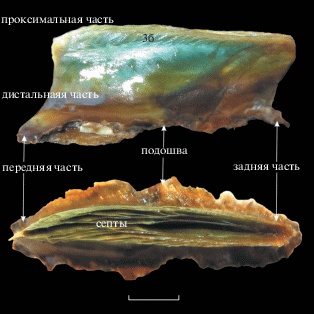
Внешне биссус у Arca boucardi представляет собой гибкий монолитный плоский тяж с отблеском серебристо-зеленого перламутра (рис. 1). У взрослой особи с длиной раковины 68 мм высота тяжа спереди и сзади составляет соответственно 8.3 и 7.9 мм, длина проксимальной части – 28 мм, дистальной – 25.3 мм. Дистальная часть биссуса заканчивается эллипсоидной клейкой подошвой (диском), которой моллюск крепится к субстрату на площади 100.9 мм2. В проксимальной части биссуса находится полость, охватывающая около 1/3 его высоты. Полость несет септы, которые полураскрытым веером распространяются от заднего края к переднему. У крупных моллюсков центральные тяжи обычно соединяются между собой перемычками вблизи переднего края, образуя ячеистую структуру. Крайние латеральные тяжи небольшим бугорком огибают верхнюю часть биссуса и на наружной его части видны как регулярно расположенные наклонные зубчики. К этим зубчикам, как и к септам внутри полости, количество и выраженность которых у моллюсков заметно варьируют, крепятся мышцы, отвечающие за функционирование биссуса.
Рис. 1.
Биссус Arca boucardi. Вверху – вид сбоку, зб – зубчики на внешней латеральной стороне; внизу – проксимальная часть, вид сверху. Масштаб – 5 мм.
Процесс повторного прикрепления A. boucardi к субстрату условно можно разделить на два последовательных этапа. Этап поиска подходящего места начинается с момента отбрасывания старого биссуса, характеризуется перемещением моллюска с помощью хорошо развитой ноги и завершается началом выделения биссусной железой прозрачного секрета в области контакта ноги с субстратом. В эксперименте без грунта продолжительность данного этапа составила от 28 мин (20% подопытных животных) до 4 ч (5%). За это время моллюски, передвигаясь по дну и стенкам аквариумов, в большинстве случаев с короткими остановками проделали путь от 4 до 22 см. Второй этап связан с обильным выделением секрета и его полимеризацией до момента, когда биссус и нога приобретут типичные форму и положение. Сначала формировалась подошва, затем моллюск постепенно поднимался над субстратом и вновь образуемым биссусом. Вместе с этим нога моллюска утолщалась и приобретала ярко-морковный цвет. При отвердении секрета и обозначении контура подошвы биссуса краски ноги блекли, она поджималась в переднюю часть раковины, прозрачность секрета исчезала, а новый биссус сначала приобретал цвет ноги, а почти через 9 ч процесса начинал зеленеть. За 1 сут все 20 особей этой серии эксперимента успешно прикрепились к оргстеклу, а 4 моллюска отбросили новый биссус и образовали другой. Наблюдения за поведением 20 особей A. boucardi, помещенными на песок, показали, что животные, преодолев участок дна с грунтом, прикрепились к стенкам аквариумов, даже не попытавшись сделать это на песке.
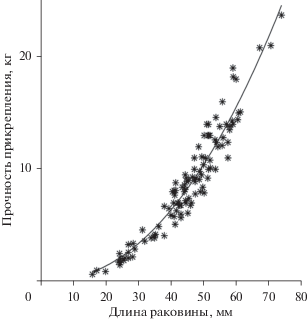
В природных условиях прочность прикрепления моллюсков к субстрату сильно варьирует в зависимости от их размера (рис. 2). Например, чтобы оторвать от субстрата моллюсков размером 30 и 60 мм, необходимо приложить усилие равное соответственно 3.2 и 16.4 кг. Эта зависимость хорошо (Р < 0.001) аппроксимируется степенным уравнением вида F = 0.0011L2.3473 (стандартная ошибка линейного коэффициента SEa равна 0.0006, степенного SEb − 0.0506; коэффициент детерминации равен 95.3%; объем наблюдений – 107 экз.). Как правило, отрыв происходит на участке подошва–субстрат.
ОБСУЖДЕНИЕ
С позиций функциональной морфологии современные представители порядка Arcoida имеют богатую предысторию многократного перехода через неотению от закапывающегося в осадок образа жизни к прикрепленному на поверхности грунта (см., например: Stanley, 1972). Это сопровождалось значительными морфологическими преобразованиями моллюсков, в частности, при переходе к эпибентосному образу жизни раковина стала вытянутой и плоской в вентральной части, где появилось зияние, связанное с формированием особого органа фиксации на субстрате – биссуса. Сходный путь развития прошли моллюски другой обширной группы обладателей биссуса – митилиды. Отличительные черты организации и функционирования биссуса у представителей этих двух родов ярко демонстрируют приспособленность современных форм к обитанию в разных биотопах и объясняют картину пространственного распределения Arca boucardi в контрастных условиях верхней части сублиторали.
Моллюски рода Arca прикрепляются к субстрату с помощью одного мощного тяжа в отличие от моллюсков рода Mytilus, которые используют для этого многочисленные эластичные нити толщиной 0.04–0.33 мм и менее (Селин, Вехова, 2002). Дистальная часть тяжа заканчивается своеобразной прикрепительной подошвой, для образования и функционирования которой необходима сравнительно большая площадь субстрата, характерная для твердых грунтов (скал, валунов). Маленькие адгезивные диски биссусных нитей мидий и модиолусов способны крепиться даже к отдельным песчинкам, что в совокупности придает моллюскам устойчивое (стабильное) положение (Meadows, Shand, 1989; Селин, Вехова, 2004). Поэтому митилиды обитают как на твердых, так и на мягких грунтах, а указания о находках представителей рода Arca на песке и илу, скорее всего, являются артефактом, связанным, в частности, со случайным заносом моллюсков в несвойственный биотоп. Экспериментальные наблюдения за поведением моллюсков в аквариумах с песком служат наглядным подтверждением этого.
Успех и прочность прикрепления моллюсков обеспечиваются не только характером грунта, но и организацией биссуса. У митилид он довольно универсальный благодаря способности нитей обеспечить прикрепление животного по всем направлениям воздействия внешних сил (волн). Моллюски существуют как бы на растяжках, направленных в разные стороны и пружинящих своей эластичной проксимальной частью при нагрузке. Биссус A. boucardi лишен такой возможности и способен противостоять лишь силам, воздействующим фронтально и вверх, создавая сопротивление не меньшее, чем, например, биссус мидии Mytilus unguiculatus (M. coruscus) (Селин, Вехова, 2002). Однако малейшее отклонение арки от вертикального положения в сторону приводит к нарушению связи с субстратом и к смыву. Этим объясняется отсутствие A. boucardi на открытых прибойных участках и обилие в местах, относительно защищенных от прямого воздействия штормовых волн (личные наблюдения), а также во внутренних частях заливов и бухт (Скарлато и др., 1967; Погребов, Кашенко, 1976, и др.).
В заключение следует отметить, что в расселении и процветании A. boucardi существенную роль играет способность моллюска легко отбрасывать биссус, активно переползать в места с благоприятными условиями и быстро формировать новое прикрепление. Не исключено, что небольшая асимметричность раковины и наличие вогнутости обеих створок в районе биссусного зияния у многих взрослых особей являются отражением неоднократной смены биссуса.
Список литературы
Евсеев Г.А., Яковлев Ю.М. Двустворчатые моллюски дальневосточных морей. Владивосток: ПК Поликон. 2006. 120 с.
Погребов В.Б., Кашенко В.П. Донные сообщества твердых грунтов залива Восток Японского моря // Биологические исследования залива Восток. Владивосток: ДВНЦ АН СССР. 1976. С. 63–82.
Селин Н.И. Состав и структура смешанных поселений Crenomytilus grayanus (Dunker, 1853) и Modiolus kurilensis (Bernard, 1983) (Bivalvia: Mytilidae) в заливе Петра Великого Японского моря // Биол. моря. 2018а. Т. 44. № 5. С. 307–316.
Селин Н.И. Онтогенетическая изменчивость прочности прикрепления к субстрату у Modiolus kurilensis F.R. Bernard, 1983 (Bivalvia: Mytilidae) в связи с друзным характером организации // Биол. моря. 2018б. Т. 44. № 6. С. 418–420.
Селин Н.И., Вехова Е.Е. Морфология двустворчатых моллюсков Crenomytilus grayanus и Mytilus coruscus в связи с особенностями их пространственного распределения в верхней сублиторали // Биол. моря. 2002. Т. 28. № 3. С. 228–232.
Селин Н.И., Вехова Е.Е. Влияние условий среды на образование биссусных нитей у некоторых представителей семейства Mytilidae из Японского моря // Биол. моря. 2004. Т. 30. № 5. С. 359–365.
Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана. Л.: Наука. 1981. 479 с.
Скарлато О.А., Голиков А.Н., Василенко С.В. и др. Состав, структура и распределение донных биоценозов в прибрежных водах залива Посьет (Японское море) // Исслед. фауны морей. 1967. Т. 5(13). С. 5–61.
Meadows P.S., Shand P. Experimental analysis of byssus thread production by Mytilus edulis and Modiolus modiolus in sediments // Mar. Biol. 1989. V. 101. № 2. P. 219–226.
Price H.A. Seasonal variation in the strength of byssal attachment of the common mussel Mytilus edulis L. // J. Mar. Biol. Assoc. U.K. 1980. V. 60. № 4. P. 1035–1037.
Stanley S.M. Functional morphology and evolution of byssally attached bivalve mollusks // J. Paleontol. 1972. V. 46. № 2. P. 165–212.
Дополнительные материалы отсутствуют.