

Биология моря, 2021, T. 47, № 5, стр. 350-354
Первое обнаружение в водах России малого маслюка Pholis nea Peden et Hughes, 1984 (Pisces: Zoarcoidei: Pholidae)
В. В. Земнухов 1, *, А. А. Баланов 1, О. А. Радченко 2, А. В. Петровская 2
1 Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
690041 Владивосток, Россия
2 Институт биологических проблем Севера ДВО РАН
685000 Магадан, Россия
* E-mail: zemnukhov1@ya.ru
Поступила в редакцию 30.06.2020
После доработки 10.11.2020
Принята к публикации 01.04.2021
Аннотация
Приведены данные о первых находках Pholis nea Peden et Hughes, 1984 в российских водах Японского моря (Татарский пролив) с описанием морфологии вида. Проведена генетическая идентификация на основе анализа участка гена COI митохондриальной ДНК (мтДНК). Для Pholis nea предложено новое русское название − малый маслюк.
В списке ихтиофауны российских вод Японского моря исследователи неоднократно указывали разукрашенного маслюка Pholis ornata (Girard, 1854) (Павленко, 1910; Солдатов, Линдберг, 1930; Линдберг, Красюкова, 1975). Долгое время считали, что этот вид имеет обширный ареал, охватывающий всю Северную Пацифику (Hubbs, 1927; Yatsu, 1981; The fishes of…, 1984), хотя П.Ю. Шмидт определенно указывал на отсутствие экземпляров данного вида из Охотского моря (Шмидт, 1950).
Ясность в этот вопрос внесли канадские ихтиологи А.Е. Педен и Г.В. Хьюз (Peden, Hughes, 1984). Проанализировав около 5 тыс. рыб рода Pholis из Северной Пацифики, они установили, что P. ornata встречается только на западном побережье Северной Америки. В данной работе по материалам с южного побережья о-ва Хоккайдо был описан новый вид – Pholis nea Peden et Hughes, 1984. Авторы упомянули, что имеющиеся в литературе указания на обитание в Японском море P. ornata, скорее всего, относятся к P. nea. Однако ни одна достоверная находка P. nea в российских водах до сих пор не описана. Цели настоящей работы − восполнение этого пробела, а также получение более подробной информации о виде, обнаруженном на некотором удалении от типового местообитания.
МАТЕРИАЛ И МЕТОДИКА
Материал собран в экспедиции Национального научного центра морской биологии им. А.В. Жирмунского ДВО РАН на НИС “Профессор Гагаринский” в Татарском проливе (Японское море) на литорали и в верхней сублиторали (до глубины 0.5 м) в июле–августе 2017 г. Исследованные рыбы хранятся в коллекции музея центра (МИМБ, Владивосток) и в Лаборатории ихтиологии Института биологических проблем Севера ДВО РАН (Магадан).
Всего для анализа использовали 21 экз. Pholis nea (табл. 1): МИМБ 40 058, 1 экз. SL 86 мм, 27.07.17, зал. Рында (MW115 530); МИМБ 40 059, 1 экз. SL 84 мм (MW115 531); МИМБ 40 060, 1 экз. SL 85 мм (MW115 532), 27.07.2017, зал. Рында, 44°47′8 N, 136°23′6 E; МИМБ 40061, 1 экз. SL 100 мм, 08.08.2017, зал. Александровский, 50°53′7 N, 142°07′1 E (MW115534); МИМБ 40062, 1 экз. SL 74 мм (MW115533), 09.08.2017, зал. Чихачева, 51°29′1 N, 140°49′3 E; МИМБ 40063, 11 экз. SL 81−111 мм, зал. Чихачева, 51°25′5 N, 140°48′9 E; МИМБ 40 064, 1 экз. SL 81 мм, 21.08.2017, зал. Рында, 44°48′1 N, 136°23′6 E; МИМБ 40 065, 4 экз. SL 81−111 мм, зал. Чихачева, 51°25′5 N, 140°48′9 E. В скобках указан номер в GenBank/NCBI.
Таблица 1.
Пластические и меристические признаки малого маслюка Pholis nea
| Признак | Cобственные данные (N = 21 экз.) | По: Peden, Hughes, 1984 (N = 11 экз.) | |
|---|---|---|---|
| голотип | паратипы | ||
| Длина SL, мм | 74–111 (92.7) | 85.3 | 36.8–92.2 |
| В % от SL | |||
| Высота тела на вертикали начала анального плавника | 11.7 (10.7–13.0) | 10 | 10–12 |
| Антедорсальное расстояние | 11.6 (10.8–12.5) | 12 | 11–16 |
| Антеанальное расстояние | 57.1 (54.3–60.7) | 58 | 56–62 |
| Длина основания спинного плавника | 86.2 (84.2–87.8) | – | – |
| Длина основания анального плавника | 40.1 (37.8–42.6) | – | – |
| Длина грудного плавника | 5.2 (4.6–6.1) | 5 | 4–5 |
| Длина головы | 12.2 (11.4–13.0) | 12 | 12–15 |
| В % от длины головы | |||
| Длина рыла | 18.1 (16.0–20.8) | 17 | 16–18 |
| Горизонтальный диаметр глаза | 21.5 (16.7–26.1) | 20 | 19–25 |
| Заглазничное расстояние | 66.3 (61.1–68.2) | – | – |
| Длина грудного плавника | 42.5 (36.4–47.8) | – | 27–41 |
| Длина хвостового плавника | 7.8 (6.7–8.5) | 7 | 6–8 |
| Счетные признаки | |||
| D | 73–78 | 74–77 | |
| A | 35–38 | 34–37 | |
| P | 10–12 | 11 | 11–12 |
| Туловищных позвонков | 38–40 | – | – |
| Хвостовых позвонков | 42–46 | – | – |
| Всего позвонков | 80–84 | 81–84 | |
Все измерения и подсчеты выполнены в соответствии со стандартными методиками (Hubbs, Lagler, 1958; Правдин, 1966), измененными с учетом особенностей морфологии изучаемого вида. Количество лучей в плавниках и позвонков подсчитано по рентгенограммам. Уростиль принимали за последний позвонок. Поры сейсмосенсорной системы головы анализировали по схеме, предложенной ранее (Макушок, 1958).
Сокращения, принятые в работе: SL – стандартная длина, мм; D, A, P, C – количество лучей в спинном, анальном, грудном и хвостовом плавниках соответственно. Указаны размеры маслюков, фиксированных в формалине и хранящихся в растворе изопропилового спирта.
Для генетического анализа использовали участок гена COI мтДНК длиной 582 пары нуклеотидов пяти видов рода Pholis с привлечением данных из GenBank/NCBI (www.ncbi.nlm.nih.gov). Выделение, амплификацию и секвенирование мтДНК проводили на основе предложенных ранее методик (Радченко и др., 2009). Последовательности мтДНК депонировали в GenBank/NCBI; инвентарные номера указаны в табл. 2. Выравнивание последовательностей, расчет р-дистанций, построение филогенетического дерева с использованием метода максимального правдоподобия (ML) выполнены в программе MEGA 6 (Tamura et al., 2013). Для оценки устойчивости топологии ML-дерева использовали бутстрэп-анализ в 1000 псевдослучайных выборок.
Таблица 2.
Значения p-дистанций (в %) между нуклеотидными последовательностями участка гена COI мтДНК некоторых видов рода Pholis (*данные из GenBank/NCBI)
| Вид (ген. номер; номер в GenBank/NCBI) | Место сбора | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pholis crassispina (1189; FJ932604) | О-в Кунашир | – | – | – | – | – | – | – | – | – | – |
| P. picta (1981; MW115535) | Татарский пролив, зал. Чихачева | 1.20 | – | – | – | – | – | – | – | – | – |
| P. picta (1987; MW115536) | Там же | 1.04 | 0 | – | – | – | – | – | – | – | – |
| P. nea (2100; MW115530) | Северное Приморье, зал. Рында | 2.58 | 2.75 | 2.60 | – | – | – | – | – | – | – |
| P. nea (2101; MW115531) | Там же | 2.58 | 2.75 | 2.60 | 0.34 | – | – | – | – | – | – |
| P. nea (2102; MW115532) | Там же | 2.75 | 2.92 | 2.77 | 0.86 | 0.86 | – | – | – | – | – |
| P. nea (2104; MW115533) | Татарский пролив, зал. Чихачева | 2.58 | 2.75 | 2.60 | 0.69 | 0.69 | 0.86 | – | – | – | – |
| P. nea (2103; MW115534) | Татарский пролив, Александровск-Сахалинский | 2.43 | 2.60 | 2.44 | 0.35 | 0.35 | 0.87 | 0.69 | – | – | – |
| P. fasciata (HQ712713)* | Аляска, Чукотское море | 2.06 | 1.20 | 1.21 | 2.41 | 2.41 | 2.58 | 2.41 | 2.43 | – | – |
| P. fasciata (HQ712711)* | Аляска, Берингов пролив | 2.06 | 1.20 | 1.21 | 2.41 | 2.41 | 2.58 | 2.41 | 2.43 | 0 | – |
| P. ornata (FJ165016)* | Канада, Британская Колумбия | 6.36 | 6.19 | 5.89 | 6.19 | 6.20 | 6.01 | 5.84 | 6.24 | 6.01 | 6.01 |
РЕЗУЛЬТАТЫ
Pholis nea Peden et Hughes, 1984 – малый маслюк (рис. 1). D LXXIII−LXXIX, A II 35–38, P 10–12, V I 1, vert 38–40 + 42–46 = 80–84 (табл. 1)
Рис. 1.
Общий вид и прижизненная окраска малого маслюка Pholis nea МИМБ 40 061, 98 мм SL (а); вариации окраски головы МИМБ 40063, 94 мм SL (б) и МИМБ 4 0063, 103 мм SL (в).

Тело удлиненное, сильно сжатое с боков. Длина рыла равна или меньше горизонтального диаметра глаза и составляет около 1/5 от длины головы. Зубы на челюстях удлиненные, конические, среднего размера; на верхней челюсти 24–26 зубов, на нижней – 18–20.
Каналы сейсмосенсорной системы головы открываются постоянным количеством пор: 2 носовых, 6 подглазничных, 6 заглазничных, 3 затылочных, 4 нижнечелюстных и 5 предкрышечных. Межглазничная пора отсутствует.
Антедорсальное расстояние приблизительно равно высоте тела на вертикали начала анального плавника и укладывается в длине тела около 10 раз. Антеанальное расстояние составляет чуть больше половины длины тела. Грудной плавник сравнительно мал, менее половины от длины головы; его лучи ветвятся 2–3 раза. Спинной плавник длинный, занимает в длине тела в среднем 86.2%; в плавнике присутствуют лишь короткие толстые колючки (табл. 1). Толщина первых 4–6 шипов спинного плавника заметно меньше, чем последующих. В начале анального плавника 2 толстых шипа; лучи мягкой части плавника членистые, ветвятся один раз. Спинной и анальный плавники соединены перепонками с хвостовым плавником. Брюшной плавник состоит из толстого шипа, а также редуцированного неветвящегося и нечленистого мягкого луча, длина которого составляет 2/3 от длины шипа.
Чешуйный покров на голове отсутствует; чешуя не заходит за границы заглазничного канала и затылочной комиссуры, на теле располагается равномерно, у хвостового плавника более редкая. Прижизненная окраска: общий фон тела красно-коричневый с многочисленными мелкими сероватыми пятнами; на спинном плавнике 11–16 выделяющихся узких белых вертикальных полосок, слегка заходящих на его основание; на анальном плавнике до 14 светлых пятен, не заходящих на его основание; на голове под глазом характерная только для этого вида белая полоса (рис. 1). Спиртовые экземпляры темно-желтые, окраска плавников в значительной мере выцветает.
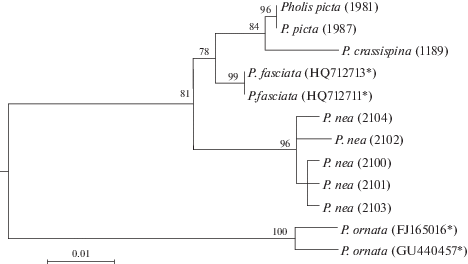
По результатам молекулярно-генетических исследований P. nea и P. ornata демонстрируют четкие видовые отличия. Гаплотипы P. ornata занимают позицию в основании дерева (рис. 2), отличаясь от таковых остальных видов рода Pholis более чем на 6%, в том числе от галотипов P. nea на 6.2% (табл. 2). Нуклеотидные последовательности всех исследованных экземпляров P. nea образуют один кластер, в пределах которого значения р-дистанций составляют 0.34–0.87%. Основные узлы ветвления филогенетического дерева поддержаны достоверными значениями бутстрэп-оценок. Несмотря на небольшую длину участка гена COI, этот маркер надежно дифференцирует изученные виды рода Pholis.
Рис. 2.
Максимально правдоподобное (ML) дерево видов рода Pholis, полученное на основе анализа нуклеотидных последовательностей участка гена COI мтДНК. Цифры в скобках – номера в GenBank/NCBI (*данные о нуклеотидной последовательности участка гена COI взяты из GenBank/NCBI); цифры над ветвями – бутстрэп-поддержки (в % от 1000 реплик бутстрэп-анализа).
ОБСУЖДЕНИЕ
Сравнительные замечания. Исследованные экземпляры относятся к Pholis nea Peden et Hughes, 1984, поскольку соответствуют диагнозу, данному авторами, описавшими вид (Peden, Hughes, 1984): межглазничная пора отсутствует, грудной плавник сравнительно небольшой, присутствуют небольшие заметные полосы на нижней части головы и отчетливые белые пятна на анальном плавнике. Помимо этого исследованные нами экземпляры соответствуют описанию вида почти по всем пластическим и счетным признакам (табл. 1, рис. 1). Имеющиеся различия (длина головы и грудного плавника), скорее всего, являются следствием возрастной изменчивости. Средняя длина (SL) исследованных нами особей 92.7 мм, в то время как в работе канадских коллег длина самого крупного экземпляра составляла 92.2 мм (Peden, Hughes, 1984). Новый максимальный размер для P. nea − 111 мм SL (МИМБ 40063).
По счетным признакам и наличию характерных полос ниже глаза к P. nea наиболее близок разукрашенный маслюк P. ornata, обитающий у западного побережья США и не встречающийся к северу от о-ва Ванкувер (Peden, Hughes, 1984). P. nea отличается от него меньшими размерами, окраской тела, головы и особенно анального плавника. У P. ornata анальный плавник светлый и без полос, у P. nea – темный со светлыми пятнами (Peden, Hughes, 1984).
От P. picta, P. fasciatus и Rhodimenichthys dolichogaster вид P. nea отличается меньшим количеством позвонков (80–84 против 87–102 у остальных видов), лучей в спинном плавнике (73–78 против 80–95 у остальных) и мягких лучей в анальном плавнике (35–38 против 38–51). Некоторое внешнее сходство P. nea имеет с видами P. nebulosa и P. crassispina, а отличается от них отсутствием чешуи на голове и характерной окраской головы, спинного и анального плавников.
Молекулярно-генетические данные также показывают четкие видовые различия между P. nea и P. ornata (рис. 2, табл. 2).
Этимология. Вместо употреблявшегося ранее другими отечественными авторами названия вида “разукрашенный маслюк” (Парин и др., 2014) мы предложили новое русское название вида P. nea – “малый маслюк”. Название “разукрашенный маслюк” давно закреплено за видом P. ornata и его использование может вносить путаницу. Название “малый маслюк” отражает особенности морфологии P. nea, так как среди маслюков, обитающих в водах дальневосточных морей России, он имеет наименьшие предельные размеры.
Список литературы
Линдберг Г.У., Красюкова З.В. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Л.: Наука. 1975. Ч. 4. 463 с.
Макушок В.М. Морфологические основы системы стихеевых и близких к ним семейств рыб (Stichaeoidae, Blennioidei, Pisces) // Тр. ЗИН АН СССР. 1958. Т. 25. С. 3–129.
Павленко М.Н. Рыбы залива Петр Великий. Типолитография Императорского университета. Казань. 1910. 95 с.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. Рыбы морей России: аннотированный каталог. М.: Товарищество науч. изд. КМК. 2014. 733 с.
Правдин И.Ф. Руководство по изучению рыб, преимущественно пресноводных. М.: Пищ. пром-ть. 1966. 377 с.
Радченко О.А., Черешнев И.А., Петровская А.В. Родственные отношения и дивергенция некоторых таксонов подсемейства Lycodinae (Zoarcidae, Pisces) по молекулярно-генетическим и морфологическим данным // Вопр. ихтиологии. 2009. Т. 49. № 5. С. 603–616.
Солдатов В.К., Линдберг Г.У. Обзор рыб дальневосточных морей // Изв. Тихоокеан. НИИ рыб. хоз-ва и океаногр. 1930. Т. 5. 576 с.
Шмидт П.Ю. Рыбы Охотского моря. М.; Л.: Изд-во АН СССР. 1950. 379 с.
Hubbs C.L. Notes on the blennioid fishes of western North America // Pap. Mich. Acad. Sci., Arts Lett. 1927. V. 7. P. 351–394.
Hubbs C.L., Lagler K.F. Fishes of the Great Lakes region. Ann Arbor, Mich.: Univ. of Michigan Press. 1958. 213 p.
The fishes of the Japanese Archipelago. 2 vols. Masuda H., Amaoka K., Araga C., Eds. Tokyo: Tokai Univ. Press. 1984. 437 p.
Peden A.E., Hughes G.W. Distribution, morphological variation, and systematic relationship of Pholis laeta and P. ornata (Pisces: Pholididae) with a description of the related form P. nea n. sp. // Can. J. Zool. 1984. V. 62. № 2. P. 291−305.
Tamura K., Stecher G., Peterson D. et al. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0 // Mol. Biol. Evol. 2013. V. 30. P. 2725−2729.
Yatsu A. A revision of the gunnel family Pholididae (Pisces, Blennioidei) // Bull. Natl. Sci. Mus., Ser. A (Tokyo). 1981. V. 7. № 4. P.165−190.
Дополнительные материалы отсутствуют.