

Биология моря, 2022, T. 48, № 4, стр. 273-282
Морфологическая изменчивость японской малоротой корюшки Hypomesus nipponensis Mcallister, 1963 (Osmeriformes, Osmeridae) Дальнего Востока
Н. С. Романов *
Национальный научный центр морской биологии им. А.В. Жирмунского (ННЦМБ) ДВО РАН
690041 Владивосток, Россия
* E-mail: n_romanov@inbox.ru
Поступила в редакцию 13.12.2021
После доработки 15.03.2022
Принята к публикации 24.03.2022
- EDN: PZFZQK
- DOI: 10.31857/S013434752204009X
Аннотация
В 10 выборках японской малоротой корюшки Hypomesus nipponensis McAllister, 1963 исследовали внутри- и межпопуляционную изменчивость числа ветвистых лучей в грудных и брюшных плавниках, а также количества заглазничных и подглазничных костей. Для анализа в дополнение к средним значениям признаков использовали дисперсию флуктуирующей асимметрии и коэффициент вариации признаков, благодаря чему число различий между выборками увеличилось в несколько раз. Это позволило лучше оценить изменчивость не только внутри популяций, но и между ними, а также определить уровень морфологических отличий каждой выборки, подчеркнув ее уникальность. Предполагается, что главная причина большого числа различий между выборками по дисперсии флуктуирующей асимметрии и коэффициенту вариации заключается в несогласованности их величин между признаками в большинстве выборок.
Японская малоротая корюшка Hypomesus nipponensis, описанная Мак Алистером (McAllister, 1963) из оз. Онума (южная часть о-ва Хоккайдо), обитает в Японском море от низовьев р. Амур до Японии и Северной Кореи, в Охотском море – в прибрежных водах, озерах и реках о-ва Сахалин, а также южных Курильских островов (Клюканов, 1966, 1970; Гриценко, Чуриков, 1983; Saruwatari et al., 1997; Гриценко, 2002; Атлас …, 2003; Сафронов, Никифоров, 2003; Заварзина, 2005) и в северных заливах Желтого моря (Chanxin, Kejing, 1987). Это самый мелкий представитель рода Hypomesus. Максимальный возраст H. nipponensis варьирует от 3+ в водах Приморья (Гриценко, 2002; Гавренков, Платошина, 2003) до 5+ на Сахалине (Заварзина, 2005) и южных Курильских островах (Сидоров, Пичугин, 2004). Длительность эмбриогенеза японской малоротой корюшки вместе с этапом предличинки при температуре воды 10 ± 2°С составляет 21.6 сут (Шадрин, 1989).
Морфология малоротых корюшек, в том числе японской, описана многими авторами (Клюканов, 1970, 1975; Гриценко, Чуриков, 1983; Парпура, Колпаков, 2001; Черешнев и др., 2001; Шедько, 2001; Гриценко, 2002; Ву, Картавцев, 2017, и др.), но популяционные аспекты в данных работах освещены лишь на уровне значений признаков. Известно, что в качестве популяционного показателя возможно использование флуктуирующей асимметрии признаков, которая является неспецифическим показателем условий развития; анализ этой величины применяется как в популяционных, так и мониторинговых исследованиях (Суле, 1984; Захаров, 1987; Parsons, 1992; Биотест…, 1993; Romanov, 1995; Романов, 2001; Романов, Скирин, 2011; Zakharov et al., 2020; Graham, 2021). По заключению Яблокова (1966, 1968) надежным показателем изменчивости служит также коэффициент вариации признаков, применяемый в популяционных исследованиях (Яблоков, 1966, 1968; Яблоков, Этин, 1968; Тимофеев-Ресовский и др., 1973).
Цель настоящей работы – проанализировать у японской малоротой корюшки внутри- и межпопуляционную изменчивость средних значений, дисперсий флуктуирующей асимметрии и коэффициентов вариации четырех легко учитываемых билатеральных признаков, чтобы доказать, что сравнение этих показателей даже по небольшому числу признаков демонстрирует уникальность каждой популяции.
МАТЕРИАЛ И МЕТОДИКА
Материалом для работы послужили сборы японской малоротой корюшки в зал. Пильтун, озерах Тунайча и Большой Вавай в Сахалинской области, а также в оз. Пресное и реках Аввакумовка, Литовка, Артемовка, Раздольная и Тесная в Приморском крае (рис. 1; табл. 1). Анализировали легко учитываемые билатеральные признаки: число ветвистых лучей в грудных (P) и брюшных (V) плавниках, а также заглазничных (porb) и подглазничных (iorb) костей, по которым проходит подглазничный канал сейсмосенсорной системы головы. Подробно методика сбора и первичной обработки материала описана ранее (Романов, 2009, 2010).
Рис. 1.
Карта-схема района исследований и места сбора проб японской малоротой корюшки Hypomesus nipponensis: 1 – зал. Пильтун, 2 – оз. Тунайча, 3 – оз. Большой Вавай, 4 – оз. Пресное, 5 – р. Аввакумовка, 6 – р. Литовка, 7 – р. Артемовка, 8 – р. Раздольная, 2001 г., 9 – р. Раздольная, 2018 г., 10 – р. Тесная.
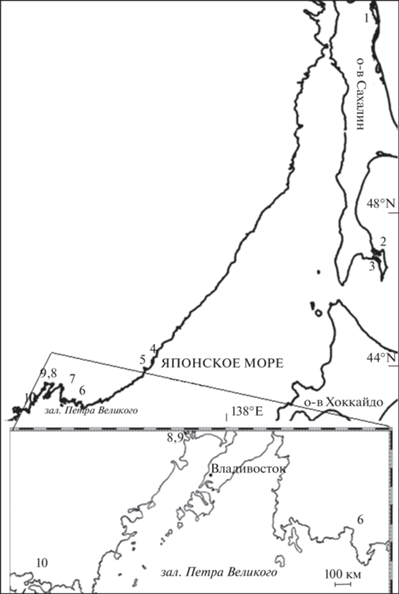
Таблица 1.
Средние значения (M ± m) (над чертой) и колебания значений признаков (под чертой) у японской малоротой корюшки Hypomesus nipponensis
| Водоем | N, экз. | P | V | porb | iorb |
|---|---|---|---|---|---|
| 1. Зал. Пильтун | 40 | $\frac{{11.13 \pm 0.05}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.03 \pm 0.018}}{{7{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.96 \pm 0.02}}{{3{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.00 \pm 0.00}}{2}$ |
| 2. Оз. Тунайча | 100 | $\frac{{11.29 \pm 0.05}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.01 \pm 0.007}}{{7{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{4.01 \pm 0.02}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 3. Оз. Большой Вавай | 85 | $\frac{{11.86 \pm 0.04}}{{11{\kern 1pt} --{\kern 1pt} 14}}$ | $\frac{{7.00 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.99 \pm 0.016}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 4. Оз. Пресное | 100 | $\frac{{11.51 \pm 0.04}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.00 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.99 \pm 0.016}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 5. Р. Аввакумовка | 88 | $\frac{{11.73 \pm 0.04}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.01 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.98 \pm 0.02}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.01 \pm 0.007}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 6. Р. Литовка | 100 | $\frac{{11.82 \pm 0.05}}{{10{\kern 1pt} --{\kern 1pt} 14}}$ | $\frac{{6.99 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.98 \pm 0.01}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.01 \pm 0.01}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 7. Р. Артемовка | 100 | $\frac{{11.48 \pm 0.05}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.01 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.99 \pm 0.02}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 8. Р. Раздольная, 2001 г. | 100 | $\frac{{11.60 \pm 0.04}}{{9{\kern 1pt} --{\kern 1pt} 12}}$ | $\frac{{7.01 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.95 \pm 0.02}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| 9. Р. Раздольная, 2018 г. | 100 | $\frac{{11.56 \pm 0.04}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.00 \pm 0.01}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{4.01 \pm 0.014}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.03 \pm 0.014}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ |
| 10. Р. Тесная | 100 | $\frac{{11.33 \pm 0.04}}{{10{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7.01 \pm 0.012}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{3.97 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
Флуктуирующую асимметрию оценивали по дисперсии, которую рассчитывали по формуле, предложенной Палмером и Штробеком (Palmer, Strobeck, 1986):
Изменчивость признаков оценивали также по коэффициенту вариации, который вычисляли по формуле: CV = 100σ/M, где σ – среднее квадратическое отклонение, а M – среднее арифметическое значение признака. Ошибку коэффициента вариации вычисляли по формуле:
Достоверность отличий средних значений (M) и коэффициентов вариации признаков определяли по величине t-критерия Стьюдента (Плохинский, 1970). Размах изменчивости показателей рассчитывали как разницу между минимумом и максимумом. Под размерностью того или иного признака понимали величину оценки признака, в данном случае – среднее арифметическое значение для всех использованных особей. Связь между параметрами оценивали с помощью непараметрического коэффициента корреляции Спирмена (rs) (Поллард, 1982), а тесноту связи – по шкале Чеддока (Сиденко и др., 2011). Для обработки данных применяли статистическую программу Systat 5.0 (Wilkinson et al., 1992a, 1992b).
РЕЗУЛЬТАТЫ
Межпопуляционная изменчивость значений признаков
Число ветвящихся лучей в грудных плавниках исследованных рыб варьировало от 9 до 14 (табл. 1). Наименьшее среднее значение отмечено для самой северной популяции зал. Пильтун, этот показатель постепенно увеличивался в выборках корюшки к югу до р. Литовка и заметно уменьшался в выборках рыб из рек Артемовка, Раздольная и Тесная. При попарном сравнении отмечено 37 достоверных различий между выборками корюшки. В наибольшей степени от других выборок отличались сахалинские выборки из зал. Пильтун и оз. Тунайча, а также приморские выборки из рек Аввакумовка и Раздольная (2018 г.) (табл. 2).
Таблица 2.
Различия между выборками японской малоротой корюшки Hypomesus nipponensis по cредним значениям (М) числа ветвистых лучей в грудных плавниках (Р)

Примечание. Номера выборок, как в табл. 1; различия достоверны при p: * – < 0.05, ** – < 0.01, *** – < 0.001; “–” – различия недостоверны.
По остальным трем признакам различия между выборками были недостоверны. В брюшных плавниках корюшки число лучей варьировало от 6 до 8. Количество заглазничных костей изменялось от 2 до 5; наименьшее среднее значение отмечено для выборки из р. Раздольная (2001 г.), а наибольшее – для выборок из оз. Тунайча и р. Раздольная (2018 г.). Число подглазничных костей в большинстве выборок колебалось в диапазоне 2–3, но в выборке 2018 г. из р. Раздольная этот показатель изменялся от 1 до 3, а в выборке из зал. Пильтун у всех исследованных рыб он был равен 2. Минимальное среднее значение отмечено для самой северной выборки (зал. Пильтун), а максимальное – для южной выборки из р. Раздольная в 2018 г. (табл. 1).
Размах изменчивости значений признаков был разным и составил 4 для числа лучей в грудных плавниках и 2 – брюшных; 3 – для числа заглазничных костей и 2 – для подглазничных. Сравнение этих величин с числом отмеченных различий между популяциями демонстрирует положительную связь (rs = 0.950, p < 0.05, теснота связи весьма высокая), так как, чем больше размах изменчивости, тем выше вероятность достоверных различий между выборками.
Изменчивость флуктуирующей асимметрии
Анализ средних значений дисперсии флуктуирующей асимметрии исследованных признаков показал, что минимальное значение характерно для числа лучей в брюшных плавниках, по остальным признакам этот показатель достоверно увеличивается в ряду: число лучей в грудных плавниках – число заглазничных костей – число подглазничных костей. Подобная закономерность характеризует выборки из оз. Большой Вавай и рек Артемовка, Раздольная (2018 г.) и Тесная. Для остальных выборок отмечены особенности: выборка из зал. Пильтун показала нулевое значение дисперсии флуктуирующей асимметрии числа подглазничных костей; выборки из озер Тунайча и Пресное, а также из р. Раздольная (2001 г.) не имели достоверных различий по числу заглазничных и подглазничных костей; в выборке из р. Аввакумовка дисперсии числа лучей в грудных и брюшных плавниках достоверно не различались, а дисперсия числа заглазничных костей была достоверно больше, чем числа подглазничных; эта же особенность отмечена и у корюшки из р. Литовка (табл. 3).
Таблица 3.
Дисперсия флуктуирующей асимметрии/103 некоторых признаков у японской малоротой корюшки Hypomesus nipponensis
| Водоем | P | V | porb | iorb | Сумма |
|---|---|---|---|---|---|
| 1. Зал. Пильтун | 1.398 | 0.777 | 5.292 | 0.000 | 7.467 |
| 2. Оз. Тунайча | 1.168 | 0.162 | 5.127 | 5.573 | 12.030 |
| 3. Оз. Большой Вавай | 1.320 | 0.441 | 4.766 | 6.509 | 13.036 |
| 4. Оз. Пресное | 1.089 | 0.737 | 5.758 | 5.573 | 13.157 |
| 5. Р. Аввакумовка | 0.852 | 0.671 | 6.056 | 3.261 | 10.840 |
| 6. Р. Литовка | 1.330 | 0.894 | 6.097 | 2.873 | 11.194 |
| 7. Р. Артемовка | 1.390 | 0.621 | 6.429 | 15.367 | 23.807 |
| 8. Р. Раздольная, 2001 г. | 1.469 | 0.376 | 10.759 | 8.221 | 20.825 |
| 9. Р. Раздольная, 2018 г. | 1.116 | 0.745 | 4.235 | 13.389 | 19.485 |
| 10. Р. Тесная | 1.344 | 0.737 | 6.114 | 13.078 | 21.273 |
| Среднее значение | 1.248 | 0.616 | 6.005 | 7.874 | – |
Примечание. Обозначения признаков, как в табл. 1.
Минимальное значение (0.852) дисперсии количества лучей в грудных плавниках отмечено для выборки из р. Аввакумовка (табл. 3), которая по этому показателю достоверно отличалась от семи выборок (табл. 4). Различия между выборками по дисперсии флуктуирующей асимметрии числа лучей в брюшных плавниках была намного больше (23 достоверных различия). По этому показателю больше всего от других выборок отличались выборки из озер Тунайча и Большой Вавай, а также из р. Раздольная (2001 г.); остальные выборки отличались незначительно. Различия по дисперсии флуктуирующей асимметрии числа заглазничных костей между выборками корюшки были значительно меньше и достоверны в 13 случаях. Больше всего от других выборок достоверно отличалась выборка из р. Раздольная (2001 г.), а меньше всего отличались выборки из зал. Пильтун и озер Тунайча и Большой Вавай. Дисперсия флуктуирующей асимметрии числа подглазничных костей изменялась от 0 в выборке из зал. Пильтун до 15.367 в выборке из р. Артемовка (табл. 3) и показала максимальные различия между выборками. От наибольшего числа выборок достоверно отличались выборки из зал. Пильтун и рек Аввакумовка, Литовка и Раздольная (2001 г.), а от наименьшего количества выборок отличалась выборка из оз. Большой Вавай (табл. 4).
Таблица 4.
Значимые различия между выборками японской малоротой корюшки Hypomesus nipponensis по дисперсии флуктуирующей асимметрии (DFA) и коэффициентам вариации признаков (CV)
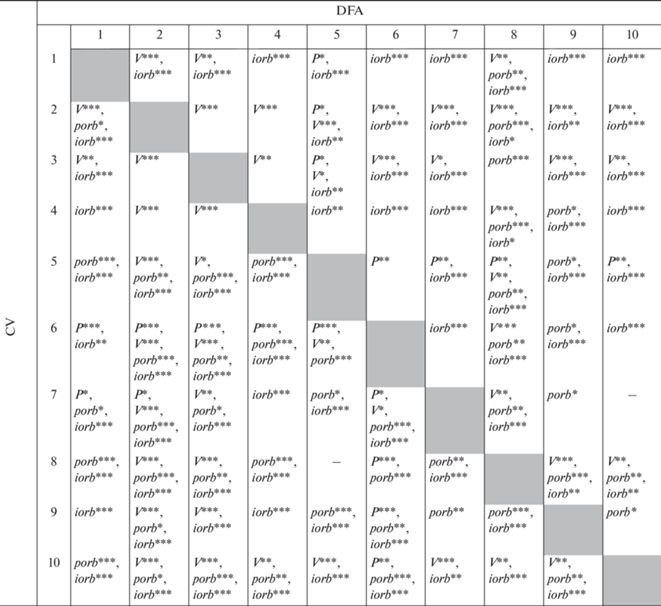
По дисперсии флуктуирующей асимметрии хотя бы одного признака достоверно различались между собой девять выборок, но между выборками из рек Артемовка и Тесная достоверные различия по этому параметру отсутствовали (табл. 4). По сумме отличий по дисперсии по всем признакам наибольшим числом достоверных отличий от других выборок выделялись выборки из рек Раздольная (2001 г.) и Аввакумовка, а наименьшим – выборки из оз. Пресное и р. Тесная (табл. 4).
Количество различий между выборками по дисперсии флуктуирующей асимметрии числа лучей в грудных плавниках было наименьшим (7), заглазничных костей – 13, лучей в брюшных плавниках – 23 и подглазничных костей – 37. Сравнивая эти показатели с размерностью признаков, получаем обратную зависимость (rs = –0.800, р < 0.05, сила связи высокая). От других выборок меньше всего отличалась выборка из оз. Пресное – 12 отличий, а больше всего – выборка из р. Раздольная (2001 г.), для которой отмечены 26 достоверных отличий (табл. 4).
Изменчивость коэффициентов вариации признаков
По результатам анализа средних значений коэффициентов вариации признаков минимальное значение этого параметра характерно для числа ветвистых лучей в брюшных плавниках (табл. 5); для числа лучей в грудных плавниках и заглазничных костей этот показатель достоверно больше, но различия между средними значениями коэффициентов вариации этих двух признаков недостоверны. Наибольшее среднее значение коэффициента вариации отмечено для числа подглазничных костей. Такая закономерность характерна для выборок из озер Большой Вавай и Пресное, а также рек Артемовка и Раздольная (2018 г.). Однако выборка из зал. Пильтун отличается нулевым значением коэффициента вариации для числа подглазничных костей; в выборках из оз. Тунайча и рек Тесная, Аввакумовка и Раздольная (2001 г.) коэффициент вариации числа ветвистых лучей в грудных плавниках достоверно меньше коэффициента для числа заглазничных костей. Кроме того, в выборках из рек Аввакумовка и Раздольная (2001 г.) коэффициент вариации числа заглазничных костей больше, чем числа подглазничных; в выборке из р. Литовка коэффициент для числа ветвистых лучей в грудных плавниках больше, чем коэффициенты для числа подглазничных и заглазничных костей (табл. 5).
Таблица 5.
Коэффициенты вариации значений некоторых признаков у японской малоротой корюшки Hypomesus nipponensis
| Водоем | P | V | porb | iorb | Сумма |
|---|---|---|---|---|---|
| 1. Зал. Пильтун | 4.40 ± 0.35 | 2.23 ± 0.18 | 4.80 ± 0.38 | 0.00 ± 0.00 | 11.43 |
| 2. Оз. Тунайча | 4.58 ± 0.23 | 1.01 ± 0.05 | 5.86 ± 0.29 | 6.93 ± 0.35 | 18.38 |
| 3. Оз. Большой Вавай | 4.79 ± 0.26 | 1.56 ± 0.08 | 5.09 ± 0.28 | 7.52 ± 0.41 | 18.96 |
| 4. Оз. Пресное | 4.95 ± 0.25 | 2.00 ± 0.10 | 5.51 ± 0.28 | 6.93 ± 0.35 | 19.39 |
| 5. Р. Аввакумовка | 4.93 ± 0.26 | 1.87 ± 0.10 | 7.20 ± 0.39 | 5.28 ± 0.28 | 19.28 |
| 6. Р. Литовка | 6.47 ± 0.32 | 2.26 ± 0.11 | 3.98 ± 0.20 | 4.96 ± 0.25 | 17.67 |
| 7. Р. Артемовка | 5.46 ± 0.30 | 1.90 ± 0.10 | 6.09 ± 0.33 | 10.54 ± 0.58 | 23.99 |
| 8. Р. Раздольная 2001 г. | 4.90 ± 0.25 | 2.01 ± 0.10 | 7.70 ± 0.39 | 5.03 ± 0.25 | 19.64 |
| 9. Р. Раздольная 2018 г. | 4.76 ± 0.24 | 2.00 ± 0.10 | 4.99 ± 0.25 | 9.85 ± 0.50 | 21.60 |
| 10. Р. Тесная | 5.24 ± 0.26 | 2.47 ± 0.12 | 6.85 ± 0.34 | 13.08 ± 0.67 | 27.64 |
| Среднее значение | 5.05 ± 0.18 | 1.93 ± 0.11 | 5.81 ± 0.37 | 7.01 ± 0.39 | – |
Коэффициент вариации числа лучей в грудных плавниках корюшки колеблется в небольших пределах, наименьшее его значение отмечено для самой северной выборки (зал. Пильтун), а наибольшее – для одной из южных выборок (р. Литовка). По этому коэффициенту выборки корюшки достоверно различаются между собой в 11 случаях: от всех других отличается выборка из р. Литовка, а выборка из р. Артемовка отличается от выборок из зал. Пильтун и оз. Тунайча (табл. 4). Размах изменчивости коэффициента вариации числа ветвистых лучей в брюшных плавниках корюшки несколько меньше. Минимальное значение отмечено для одной из самых северных выборок (оз. Тунайча), а максимальное – для самой южной выборки из р. Тесная (табл. 5). По этому коэффициенту выборки корюшки достоверно различаются между собой в 24 случаях. Выборки из озер Тунайча и Большой Вавай демонстрируют по девять различий, а выборка зал. Пильтун показывает два различия (табл. 4). Размах изменчивости коэффициента вариации числа заглазничных костей находится в диапазоне от 3.9 до 7.7 (табл. 5); наименьшим (р. Литовка) и наибольшим (р. Раздольная, 2001 г.) значениями характеризуются южные выборки корюшки. Зафиксировано 30 достоверных различий между выборками. Выборка из р. Литовка отличается от всех остальных выборок, за исключением выборки из зал. Пильтун; от семи из исследованных выборок отличаются выборки из рек Аввакумовка и Раздольная (2001 г.). Выборки разных лет из р. Раздольная также различаются по этому параметру (табл. 4). Наибольший размах изменчивости коэффициента вариации отмечен для числа подглазничных костей − от нулевого значения (зал. Пильтун) до максимального в выборке из р. Тесная, самой южной из исследованных популяций (табл. 5). В 38 случаях выборки корюшки достоверно различались между собой по коэффициенту вариации этого признака. Выборки из зал. Пильтун и р. Тесная показали по девять, из рек Артемовка и Раздольная (2018 г.) – по восемь, а все остальные выборки – по семь различий. Как и в предыдущем случае, выборки разных лет из р. Раздольная различались между собой.
Почти все исследованные выборки достоверно различались между собой по коэффициентам вариации хотя бы одного признака; исключение составили выборки из рек Аввакумовка и Раздольная (2001 г.), различия между которыми оказались недостоверны. По сумме коэффициентов вариации всех признаков выборка из зал. Пильтун демонстрирует минимальное, а из р. Тесная – максимальное значение этого показателя; выборки из р. Раздольная за разные годы не различаются (табл. 5). Сопоставление размерности признаков с числом различий между выборками по коэффициентам вариации признаков обнаруживает обратную зависимость между ними (rs = −1.00, р < 0.01, сила связи функциональная).
ОБСУЖДЕНИЕ
Между выборками японской малоротой корюшки из разных популяций отмечено 36 достоверных различий по средним значениям числа лучей в грудных плавниках. В девяти попарных сравнениях выборки не различались между собой. Применение для анализа морфологической изменчивости не только средних значений признаков, но и дисперсий флуктуирующей асимметрии признаков (80 различий) и коэффициентов их вариации (103 различия) увеличило количество различий между выборками до 219. Главная причина этого заключается в несогласованности дисперсий асимметрии и коэффициентов вариации между признаками в большинстве выборок, что служит подтверждением уникальности каждой выборки. Анализ дисперсии флуктуирующей асимметрии и коэффициента вариации признаков показал, что только в четырех случаях из 10 соотношение этих внутрипопуляционных показателей соответствует соотношению, характерному для средних (видовых) значений. Только для трех выборок эти соотношения схожи, как для дисперсии, так и для коэффициента вариации. Для остальных выборок отмечена несогласованность по обоим показателям, которая подтверждается отсутствием статистически значимой связи на популяционном уровне между средними значениями признаков и значениями дисперсий асимметрии и коэффициентов вариации. Статистически значимая связь не была обнаружена и при попытке найти зависимость между значениями дисперсии асимметрии или коэффициентом вариации признаков и числом различий по этим показателям.
Таким образом, применение дисперсии флуктуирующей асимметрии и коэффициента вариации в дополнение к средним значениям признаков позволяет лучше оценить как внутри-, так и межпопуляционную изменчивость, т.е. уровень морфологических отличий каждой выборки, подчеркнув тем самым ее уникальность. Нельзя не отметить следующую закономерность: чем больше размах изменчивости того или иного показателя, тем больше достоверных различий обнаруживается между выборками, что согласуется с мнением некоторых авторов, высказанным ранее в отношении коэффициентов вариации признаков (Яблоков, 1966; Соболевский, 1988).
Сравнение суммы дисперсий всех проанализированных признаков выборки (табл. 3) показало высокий уровень этого показателя у малоротой корюшки из самых южных водоемов – рек Артемовка, Раздольная (2001, 2018 гг.) и Тесная, находящихся под мощным антропогенным прессом (Шулькин, Семыкина, 2005; Никулина, 2006; Кику, Ковековдова, 2008). Для северных местообитаний значения суммы дисперсий значительно меньше. Это может указывать на более высокую стабильность развития корюшки северных популяций, обеспеченную оптимальными условиями эмбрионального и раннего постэмбрионального развития. Выборки корюшки разных лет из р. Раздольная по величине данного показателя не различаются, что может говорить об одинаковом общем уровне воздействия на эмбриогенез и ранний постэмбриогенез корюшки выборок 2001 и 2018 гг.
Однако достоверные различия, отмеченные между выборками корюшки из р. Раздольная в разные годы по дисперсии флуктуирующей асимметрии и коэффициенту вариации анализируемых признаков (по трем и двум признакам соответственно), хорошо согласуются с заключением А.В. Яблокова о том, что “изменчивость даже одного и того же признака оказывается различной в разные сезоны, годы, в разных популяциях вида” (Яблоков, 1968, с. 1755). Каждая популяция адаптирована к определенным условиям обитания, которые могут меняться в разные годы, и реакция развивающихся организмов на эти условия может быть разной, что находит отражение в использованных для анализа параметрах.
Список литературы
Атлас пресноводных рыб России. М.: Наука. 2003. Т. 1. 379 с.
Биотест: интегральная оценка здоровья экосистем и отдельных видов. М.: Московское отд. МФ “Биотест”. 1993. 68 с.
Ву К.Т., Картавцев Ю.Ф. Морфометрические различия малоротых корюшек Hypomesus japonicus (Brevoort, 1856) и H. nipponensis (McAllister, 1963) (Pisces, Osmeridae) из северо-западной части Японского моря // Биол. моря. 2017. Т. 43. № 6. С. 403–412.
Гавренков Ю.И., Платошина Л.К. Биология и экология размножения малоротых корюшек рода Hypomesus (Osmeridae) Приморья // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука. 2003. Вып. 2. С. 425–435.
Гриценко О.Ф. Проходные рыбы острова Сахалин. Сис-тематика, экология, промысел. М.: Изд-во ВНИРО. 2002. 247 с.
Гриценко О.Ф., Чуриков А.А. Систематика малоротых корюшек рода Hypomesus (Salmoniformes, Osmeridae) Азиатского побережья Тихого океана // Зоол. журн. 1983. Т. 52. Вып. 4. С. 553–563.
Заварзина Н.К. О методике определения возраста и размерно-возрастных особенностях малоротых корюшек рода Hypomesus острова Сахалин // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука. 2005. Вып. 3. С. 585–593.
Захаров В.М. Асимметрия животных. M.: Наука. 1987. 216 с.
Кику Д.П., Ковековдова Л.Т. Сравнительная оценка содержания микроэлементов в двустворчатых моллюсках из Уссурийского и Амурского заливов // Уссурийский залив: современное экологическое состояние, ресурсы и перспективы природопользования. Материалы международ. научно-практ. конф. Владивосток: Изд-во ДВГУ. 2008. С. 94–98.
Клюканов В.А. Новые данные о распространении малоротых корюшек в водах СССР // Докл. АН СССР. 1966. Т. 166. № 4. С. 990–991.
Клюканов В.А. Морфологические основы систематики корюшек рода Hypomesus (Osmeridae) // Зоол. журн. 1970. Т. 49. Вып. 10. С. 1534–1541.
Клюканов В.А. Систематика и родственные отношения корюшек родов Osmerus и Hypomesus (Osmeridae) и их расселение // Зоол. журн. 1975. Т. 54. Вып. 4. С. 590–595.
Никулина Т.В. Оценка экологического состояния р. Раздольная по составу индикаторных видов водорослей // Вестн. ДВО РАН. 2006. № 6. С. 71–78.
Парпура И.З., Колпаков Н.В. Биология и внутривидовая дифференциация корюшек Приморья // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука. 2001. Вып. 1. С. 284–295.
Плохинский Н.А. Биометрия. М.: Изд-во МГУ. 1970. 367 с.
Поллард Дж. Справочник по вычислительным методам статистики. М.: Финансы и статистика. 1982. 344 с.
Романов Н.С. Флуктуирующая асимметрия лососей заводского и естественного воспроизводства // Чтения памяти В.Я. Леванидова. Владивосток: Дальнаука. 2001. Вып. 1. С. 328–335.
Романов Н.С. Флуктуирующая асимметрия некоторых признаков у симы Oncorhynchus masou (Salmonidae) // Вестн. Северо-Восточ. науч. центра. 2009. № 1. С. 62–67.
Романов Н.С. Флуктуирующая асимметрия некоторых признаков у морской малоротой корюшки Hypomesus japonicus (Osmeridae) // Вестн. Северо-Восточ. науч. центра. 2010. № 3. С. 37–43.
Романов Н.С., Скирин В.И. Морфологическая изменчивость некоторых осетровых рыб и их искусственных гибридов // Изв. ТИНРО. 2011. Т. 165. С. 283–296.
Сафронов С.Н., Никифоров С.Н. Список рыбообразных и рыб пресных и солоноватых вод Сахалина // Вопр. ихтиологии. 2003. Т. 43. № 1. С. 42–53.
Сиденко А.В., Вишняков В.В., Исаев С.М. Теория статистики. М.: МАКС-Пресс. 2011. 343 с.
Сидоров Л.К., Пичугин М.Ю. Морфологические особенности озерных форм малоротой корюшки рода Hypomesus (Salmoniformes) южных Курильских островов // Вопр. ихтиологии. 2004. Т. 44. № 4. С. 484–495.
Соболевский Е.И. Популяционная морфология ластоногих. Изменчивость и пространственная структура вида. М.: Наука. 1988. 216 с.
Суле М. Алломерическая изменчивость: теория и следствие // Журн. общ. биол. 1984. Т. XLV. № 1. С. 16–26.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. М.: Наука. 1973. 278 с.
Черешнев И.А., Шестаков А.В., Фролов С.В. К систематике малоротых корюшек рода Hypomesus (Osmeridae) залива Петра Великого Японского моря // Биол. моря. 2001. Т. 27. № 5. С. 340–346.
Шадрин А.М. Эмбрионально-личиночное развитие корюшковых (Osmeridae) Дальнего Востока. IV. Hypomesus nipponensis // Вопр. ихтиологии. 1989. Т. 29. № 6. С. 960–972.
Шедько С.В. О видовом составе корюшек (Osmeridae) в водах Приморья // Вопр. ихтиологии. 2001. Т. 41. № 2. С. 261–264.
Шулькин В.М., Семыкина Г.В. Сезонная и многолетняя изменчивость содержания и выноса биогенных соединений р. Раздольной (Приморский край) // Вод. ресурсы. 2005. Т. 32. № 5. С. 575–583.
Яблоков А.В. Изменчивость млекопитающих. М.: Наука. 1966. 364 с.
Яблоков А.В. Популяционная морфология животных // Зоол. журн. 1968. Т. 47. № 12. С. 1749–1765.
Яблоков А.В., Этин В.Я. Изменчивость меристических признаков нутрии (Myocastor coypus Mol.) Азербайджана и Таджикистана // Зоол. журн. 1968. Т. 47. № 1. С. 116–121.
Chanxin L., Kejing Q. Fauna Liaoningica. Pisces. Shenyang: Liaoning Science and Technology Press. 1987. 552 p.
Graham J.H. Fluctuating asymmetry and developmental instability, a guide to best practice // Symmetry. 2021. V. 13. Art. ID 9. https://doi.org/10.3390/sym13010009
Graham J., Freeman D.C., Emlen J. Antisymmetry, directional asymmetry, and dynamic morphogenesis // Genetica. 1993. V. 89. № 1–3. P. 121–137.
McAllister D.E. A revision of the smelt family, Osmeridae // Bulletin of the National Museum of Canada № 191. Ottawa: National Museum of Canada. 1963.
Palmer A.R., Strobeck C. Fluctuating asymmetry: measurement, analysis, patterns // Annu. Rev. Ecol. Syst. 1986. № 17. P. 391–421.
Parsons P.A. Fluctuating asymmetry: a biological monitor of environmental and genomic stress // Heredity. 1992. V. 68. P. 361–364.
Romanov N.S. Fluctuating asymmetry in chum salmon, Oncorhynchus keta, from the Maritime Province // J. Ichthyol. 1995. V. 35. № 9. P. 171–182.
Saruwatari T., Lopez J.A., Pietsch T.W. A revision of the osmerid genus Hypomesus Gill (Teleostei: Salmoniformes), with the description of a new species from the Southern Kuril Islands // Species Diversity. 1997. V. 2. № 1. P. 59–82.
Sokal R.R., Rohlf F.J. Biometry. New York: Freeman & Co. 1981. 859 p.
Wilkinson L., Hill M.-A., Welna J.P., Birkenbeuel G.K. SYSTAT for Windows: statistics, Version 5. Evanston, Ill.: Systat Inc. 1992a. 750 p.
Wilkinson L., Hill M.-A., Miceli S. et al. SYSTAT for Windows: graphics. Version 5. Evanston, Ill.: Systat Inc. 1992b. 636 p.
Zakharov V.M., Shadrina E.G., Trofimov I.E. Fluctuating asymmetry, developmental noise and developmental stability: future prospects for the population developmental biology approach // Symmetry. 2020. 12(8). 1376. https://doi.org/10.3390/sym12081376
Дополнительные материалы отсутствуют.