

Биология моря, 2022, T. 48, № 6, стр. 380-387
Осенний бактериопланктон северо-восточной части Баренцева моря
М. П. Венгер 1, В. Г. Дворецкий 1, *, Т. М. Максимовская 1, Т. Г. Ишкулова 1, А. В. Ващенко 1, В. В. Водопьянова 1
1 Мурманский морской биологический институт РАН (ММБИ)
183010 Мурманск, Россия
* E-mail: v-dvoretsky@yandex.ru
Поступила в редакцию 28.03.2022
После доработки 24.06.2022
Принята к публикации 28.06.2022
- EDN: XQTVYZ
- DOI: 10.31857/S0134347522060146
Аннотация
Впервые изучено осеннее состояние бактериопланктона в северо-восточной части Баренцева моря. В районе исследований выделены теплые трансформированные атлантические, холодные арктические, промежуточные и придонные воды. В верхнем слое содержание минеральных биогенных элементов и кислорода было низким, максимальные концентрации биогенных элементов зарегистрированы в придонном слое. Численность бактерий изменялась от 220 до 1068 тыс. кл./мл, биомасса – от 2.7 до 12.5 мг С/м3, средний объем клеток варьировал от 0.020 до 0.136 мкм3. Горизонтальное распределение бактериопланктона характеризовалось мозаичностью, более явно был выражен вертикальный тренд – снижение количества бактерий и увеличение размеров их клеток с глубиной. В составе бактериопланктона доминировали одиночные мелкие клетки. Общее количество и биомасса бактериопланктона, а также численность некоторых размерных групп бактерий обратно коррелировали с концентрацией биогенных элементов (нитратов, фосфатов, силикатов) и с соленостью. Численность и биомасса бактерий демонстрировали слабую тенденцию к росту при увеличении температуры воды и содержания хлорофилла а в поверхностном слое. Проведено сравнение полученных результатов с литературными данными по другим сезонам и арктическим районам. Обсуждаются факторы, определяющие особенности распределения морских бактерий в осенний период.
С конца 20-го века Северный Ледовитый океан подвергается существенным климатическим изменениям (Climatic atlas …, 2004). Отмечен выраженный тренд на потепление, снижение общей площади льда и удлинение безледного периода (ICES, 2021). В свою очередь, это оказывает большое влияние на все компоненты морских экосистем – от микробов до китообразных. Регистрируются изменения потоков органического углерода в пелагических трофических сетях, повышается активность микроорганизмов и снижается эффективность передачи энергии по пищевым цепям (Копылов, Косолапов, 2011).
Баренцево море – крупнейший шельфовый арктический район, который служит транзитной зоной между Атлантическим и Северным Ледовитым океанами (Ecosystem …, 2009). Состояние экосистем Баренцева моря во многом определяется интенсивностью притока теплых высокосоленых вод атлантического происхождения из Норвежского моря, а в северных районах основное влияние оказывают холодные воды (Жизнь …, 1985; Wassmann et al., 2006; Ecosystem …, 2009). Здесь наблюдается высокая пространственно-временная вариабельность абиотических и биотических факторов, которая создает основу для существенной пространственной и сезонной вариации функционирования микробных трофических сетей (Жизнь …, 1985; Планктон морей …, 1997).
Микробиологические исследования в Баренцевом море проводятся в течение многих лет, большая часть данных получена для весенне-летнего периода (Планктон морей …, 1997; Howard-Jones et al., 2002; Sturluson et al., 2008; Tammert et al., 2008; Венгер, 2011; Венгер и др., 2012, 2016), преимущественно для южных, центральных и западных районов. Для северной части моря сведений о состоянии бактериопланктона очень мало, они относятся в основном к периоду массового цветения фитопланктона (Sturluson et al., 2008; Tammert et al., 2008). Однако для понимания сезонной динамики населения пелагиали необходимы данные о состоянии компонентов планктонных сообществ и в другие периоды. Исследований, анализирующих осеннее и зимнее состояние сообществ морских микробов, относительно немного, причем в них рассматриваются прибрежные и центральные районы моря (Широколобова и др., 2016).
Цель настоящей работы – изучить особенности пространственного распределения бактериопланктона в северо-восточной части Баренцева моря в осенний период, провести анализ размерно-морфологического состава сообщества бактерий и выявить внешние факторы, влияющие на обилие бактериопланктона.
МАТЕРИАЛ И МЕТОДИКА
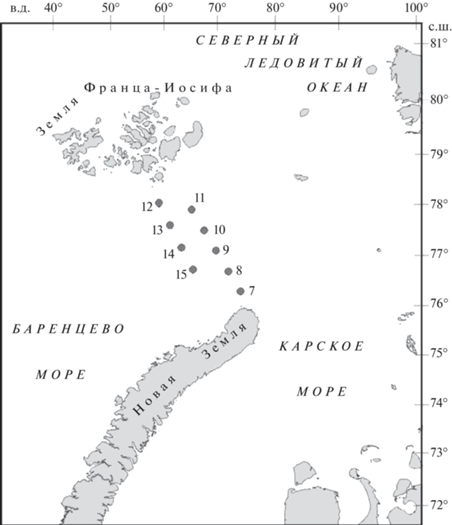
В работе использовали данные, полученные в экспедиции ММБИ РАН на НИС “Дальние Зеленцы” в первой декаде октября 2020 г. (рис. 1). Вертикальные профили температуры воды и солености получены с помощью зонда СТД-SEACAT SBE 19plus V2. Глубина на станциях в районе исследований варьировала от 190 до 370 м. Пробы воды для гидрохимического и микробиологического анализа в зависимости от глубины станции отбирали на 5–6 горизонтах (0, 10, 25, 50, 100 м, дно). Для отбора проб использовали комплекс Rosette, снабженный пластиковыми батометрами Нискина.
Концентрацию растворенного кислорода определяли по методу Винклера. Насыщенность вод кислородом рассчитывали по стандартным формулам (Таблицы …, 1976). Фосфаты, силикаты, нитриты и нитраты определяли стандартными методами (Руководство …, 1993). Для анализа содержания хлорофилла a отбирали пробы объемом по 5 л с поверхностного горизонта (0, 10 м). Всего для анализа фитопигментов было отобрано 13 проб воды. Воду фильтровали под вакуумом (перепад давления 0.15–0.20 атм), использовали мембраны “Владипор” типа МФАС с диаметром пор 0.6 мкм. После гомогенизации фильтров хлорофилл экстрагировали 90%-ным ацетоном. Определение осуществляли на спектрофотометре Nicolett Evolution 500. Концентрацию хлорофилла а (мг/м3) вычисляли по стандартным формулам (ГОСТ…, 2001).
Воду для микробиологического анализа фиксировали 40%-ным формалином (конечная концентрация в пробе 2%). Количественный учет микроорганизмов проводили методом эпифлуоресцентной микроскопии; препараты просматривали при увеличении ×1000 в свете УФ-лучей под микроскопом Olympus BX 53 с системой анализа изображения CellSensStandart (Япония).
Численность бактерий определяли с использованием флуорохрома DAPI и ядерных фильтров с диаметром пор 0.2 мкм (Porter, Feig, 1980). При подсчете бактерий просматривали 20–40 полей зрения. В каждой пробе просчитывали 400 и более клеток. Средний объем клеток рассчитывали на основе промеров не менее 100 бактерий из каждой пробы. Углеродную биомассу бактерий рассчитывали по уравнению (Norland, 1993): С = 120 × V0.72, где C – содержание углерода, фг/кл.; V – средний объем клетки, мкм3. По размерно-морфологическому составу выделяли мелкие одиночные клетки (диаметр и длина менее 2 мкм), среди которых выделяли ультрамикроорганизмы (объем клетки менее 0.04 мкм3); крупные палочковидные клетки (размер более 2 мкм); нитевидные клетки (соотношение длины к ширине более 10) и агрегированные бактерии на частицах детрита.
Распределение бактерий, а также гидрохимических и океанологических показателей рассматривали для четырех горизонтов: 0–10 м – поверхностный горизонт, 25−50 м – подповерхностный, 100 м – промежуточный и 190–365 м – придонный.
Для выявления различий между горизонтами отбора проб при нормальном распределении данных применяли однофакторный дисперсионный анализ, в остальных случаях − непараметрический тест Краскела−Уоллиса. Гомогенность данных проверяли при помощи модифицированного теста Левене. Для оценки возможных связей между численностью бактериопланктона, его биомассой, средним размером клеток и внешними факторами использовали метод ранговой корреляции Спирмена. Различия считали достоверными при p < 0.05. Средние величины представлены со стандартным отклонением.
РЕЗУЛЬТАТЫ
Температура воды в поверхностном слое изменялась от 0.06°С на севере до 2.7°С на юге, соленость варьировала от 33.49 до 34.52‰ (табл. 1). Отмечены теплые трансформированные атлантические (ТАВ), холодные арктические (АРВ), промежуточные и придонные воды. Потоки ТАВ в слое от 25 до 75 м представлены ветвью Новоземельского течения, а в слое от 100 до 200 м – ветвью заглубленного течения желоба Святой Анны. На состояние верхнего слоя водной толщи определяющее влияние оказывал ветер, под воздействием которого происходило смешение ТАВ и АРВ в слое 0–25 м. Глубина залегания пикноклина (слоя скачка плотности воды) менялась в небольших пределах: от 30–45 м на северных станциях, до 40–60 м на южных. Ниже, под пикноклином, располагались воды с отрицательной температурой от –0.2 до –1.5°С и соленостью 34.50–34.70‰ (табл. 1). Между слоями воды выявлены достоверные различия по температуре и солености (p < 0.001).
Таблица 1.
Гидрологические параметры, концентрация биогенных элементов, хлорофилла а и микробиологические показатели в разных водных горизонтах северо-восточной части Баренцева моря (октябрь 2020 г.)
| Параметр | Горизонты | |||
|---|---|---|---|---|
| поверхностный (0–10 м) | подповерхностный (25–50 м) | промежуточный (100 м) | придонный | |
| Температура, °С | $\frac{{0.06{\kern 1pt} --{\kern 1pt} 2.41}}{{1.29 \pm 0.69}}$ | $\frac{{\left( { - 1.48} \right){\kern 1pt} --{\kern 1pt} 2.51}}{{0.24 \pm 1.26}}$ | $\frac{{\left( { - 1.65} \right){\kern 1pt} --{\kern 1pt} 1.18}}{{\left( { - 0.18} \right) \pm 1.02}}$ | $\frac{{\left( { - 1.3} \right){\kern 1pt} --{\kern 1pt} \left( { - 0.76} \right)}}{{\left( { - 0.99} \right) \pm 0.18}}$ |
| Соленость, ‰ | $\frac{{33.49{\kern 1pt} --{\kern 1pt} 34.27}}{{33.91 \pm 0.25}}$ | $\frac{{33.51{\kern 1pt} --{\kern 1pt} 34.7}}{{34.24 \pm 0.38}}$ | $\frac{{34.65{\kern 1pt} --{\kern 1pt} 34.81}}{{34.74 \pm 0.05}}$ | $\frac{{34.73{\kern 1pt} --{\kern 1pt} 34.84}}{{34.81 \pm 0.04}}$ |
| Хлорофилл а, мг/м3 | $\frac{{0.18{\kern 1pt} --{\kern 1pt} 0.36}}{{0.28 \pm 0.06}}$ | – | – | – |
| Растворенный кислород, мл/л | $\frac{{7.62{\kern 1pt} --{\kern 1pt} 8.01}}{{7.89 \pm 0.13}}$ | $\frac{{7.05{\kern 1pt} --{\kern 1pt} 8.2}}{{7.75 \pm 0.29}}$ | $\frac{{6.81{\kern 1pt} --{\kern 1pt} 8.2}}{{7.09 \pm 0.42}}$ | $\frac{{6.81{\kern 1pt} --{\kern 1pt} 7.0}}{{6.9 \pm 0.06}}$ |
| Насыщение воды кислородом, % | $\frac{{100{\kern 1pt} --{\kern 1pt} 101}}{{100 \pm 0}}$ | $\frac{{87{\kern 1pt} --{\kern 1pt} 101}}{{96 \pm 5}}$ | $\frac{{84{\kern 1pt} --{\kern 1pt} 98}}{{88 \pm 4}}$ | $\frac{{82{\kern 1pt} --{\kern 1pt} 84}}{{83 \pm 1}}$ |
| Концентрация биогенных элементов, мкг моль/л | ||||
| РО4 | $\frac{{0.03{\kern 1pt} --{\kern 1pt} 0.17}}{{0.1 \pm 0.05}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.7}}{{0.23 \pm 0.21}}$ | $\frac{{0.13{\kern 1pt} --{\kern 1pt} 0.85}}{{0.63 \pm 0.22}}$ | $\frac{{0.25{\kern 1pt} --{\kern 1pt} 0.89}}{{0.63 \pm 0.18}}$ |
| NO2 | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.09}}{{0.02 \pm 0.02}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.27}}{{0.1 \pm 0.09}}$ | $\frac{{0.01{\kern 1pt} --{\kern 1pt} 0.13}}{{0.05 \pm 0.04}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.07}}{{0.03 \pm 0.02}}$ |
| NО3 | $\frac{{0.1{\kern 1pt} --{\kern 1pt} 1.37}}{{0.87 \pm 0.39}}$ | $\frac{{0.1{\kern 1pt} --{\kern 1pt} 10.45}}{{3.06 \pm 3.57}}$ | $\frac{{5.06{\kern 1pt} --{\kern 1pt} 17.26}}{{12.87 \pm 4.53}}$ | $\frac{{14.97{\kern 1pt} --{\kern 1pt} 18.65}}{{17.46 \pm 1.08}}$ |
| SiO3 | $\frac{{0.39{\kern 1pt} --{\kern 1pt} 1.94}}{{0.9 \pm 0.45}}$ | $\frac{{0.4{\kern 1pt} --{\kern 1pt} 1.95}}{{1.13 \pm 0.5}}$ | $\frac{{1.95{\kern 1pt} --{\kern 1pt} 5.44}}{{3.66 \pm 1.19}}$ | $\frac{{4.34{\kern 1pt} --{\kern 1pt} 8.6}}{{6.03 \pm 1.5}}$ |
| Биомасса бактерий, мг С/м3 | $\frac{{3.6{\kern 1pt} --{\kern 1pt} 12.5}}{{6.31 \pm 2.46}}$ | $\frac{{3.4{\kern 1pt} {\kern 1pt} --{\kern 1pt} 12}}{{7.1 \pm 2.17}}$ | $\frac{{3{\kern 1pt} --{\kern 1pt} 5.4}}{{4.36 \pm 0.89}}$ | $\frac{{2.7{\kern 1pt} --{\kern 1pt} 8.2}}{{4.4 \pm 1.78}}$ |
| Средний объем клетки, мкм3 | $\frac{{0.02{\kern 1pt} --{\kern 1pt} 0.074}}{{0.038 \pm 0.014}}$ | $\frac{{0.025{\kern 1pt} --{\kern 1pt} 0.085}}{{0.04 \pm 0.014}}$ | $\frac{{0.037{\kern 1pt} --{\kern 1pt} 0.071}}{{0.048 \pm 0.011}}$ | $\frac{{0.031{\kern 1pt} --{\kern 1pt} 0.136}}{{0.054 \pm 0.032}}$ |
| Численность бактерий, тыс. кл./мл | ||||
| Общая численность | $\frac{{333{\kern 1pt} --{\kern 1pt} 925}}{{563 \pm 143}}$ | $\frac{{406{\kern 1pt} --{\kern 1pt} 1068}}{{622 \pm 172}}$ | $\frac{{230{\kern 1pt} --{\kern 1pt} 392}}{{328 \pm 57}}$ | $\frac{{220{\kern 1pt} --{\kern 1pt} 409}}{{313 \pm 71}}$ |
| Группа 1 | $\frac{{239{\kern 1pt} --{\kern 1pt} 698}}{{385 \pm 120}}$ | $\frac{{189{\kern 1pt} --{\kern 1pt} 729}}{{418 \pm 124}}$ | $\frac{{142{\kern 1pt} --{\kern 1pt} 284}}{{198 \pm 48}}$ | $\frac{{86{\kern 1pt} --{\kern 1pt} 283}}{{178 \pm 63}}$ |
| Группа 2 | $\frac{{51{\kern 1pt} --{\kern 1pt} 391}}{{170 \pm 88}}$ | $\frac{{63{\kern 1pt} --{\kern 1pt} 454}}{{197 \pm 89}}$ | $\frac{{85{\kern 1pt} --{\kern 1pt} 196}}{{127 \pm 40}}$ | $\frac{{63{\kern 1pt} --{\kern 1pt} 214}}{{130 \pm 60}}$ |
| Группа 3 | $\frac{{0.2{\kern 1pt} --{\kern 1pt} 29.4}}{{4 \pm 7}}$ | $\frac{{0.3{\kern 1pt} --{\kern 1pt} 23.8}}{{4 \pm 7}}$ | $\frac{{0.4{\kern 1pt} --{\kern 1pt} 6.7}}{{2 \pm 2}}$ | $\frac{{0.3{\kern 1pt} --{\kern 1pt} 13.5}}{{3 \pm 4}}$ |
| Группа 4 | $\frac{{0.5{\kern 1pt} --{\kern 1pt} 9.5}}{{4 \pm 2}}$ | $\frac{{0.4{\kern 1pt} --{\kern 1pt} 5.8}}{{2 \pm 1}}$ | $\frac{{0.4{\kern 1pt} --{\kern 1pt} 3.8}}{{1 \pm 1}}$ | $\frac{{1.1{\kern 1pt} --{\kern 1pt} 5.4}}{{2 \pm 1}}$ |
| Группа 5 | $\frac{{0{\kern 1pt} --{\kern 1pt} 2}}{{0.38 \pm 0.53}}$ | $\frac{{0.1{\kern 1pt} --{\kern 1pt} 1.3}}{{0.48 \pm 0.33}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.6}}{{0.24 \pm 0.19}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 0.5}}{{0.2 \pm 0.17}}$ |
| Группа 6 | – | $\frac{{0{\kern 1pt} --{\kern 1pt} 2}}{{0.03 \pm 0.01}}$ | $\frac{{0{\kern 1pt} --{\kern 1pt} 1.7}}{{0.07 \pm 1}}$ | – |
Примечание. Здесь и в табл. 2: группа 1 – ультрамикроорганизмы; группа 2 – мелкие клетки; группа 3 – крупные клетки (палочки); группа 4 – клетки, агрегированные на детрите; группа 5 – нитевидные клетки; группа 6 – микроколонии. PO4 – фосфаты, NO2 – нитриты, NO3 – нитраты, SiO3 – силикаты. Над чертой – диапазон значений, под чертой – среднее.
Верхний водный слой характеризовался низкими концентрациями минеральных биогенных элементов (табл. 1), при этом насыщение воды кислородом и содержание О2 были высокими. Минимальные концентрации фосфатов, нитритов и нитратов и максимальное содержание кремния определены в АРВ. Отмечены невысокие концентрации растворенного кислорода (табл. 1). От побережья Новой Земли (ТАВ) в северном направлении (АРВ) на глубине 50 м наблюдалась область повышенного содержания нитритного азота, которая образовалась в результате окисления органического вещества, скопившегося над скачком плотности. Максимальные концентрации биогенных веществ регистрировали в придонных водах. Различия показателей на разных горизонтах были статистически значимыми (p < 0.05). Концентрация хлорофилла а в поверхностном слое была низкой и колебалась в узких пределах (табл. 1).
Численность бактерий изменялась от 220 до 1068 тыс. кл./мл, а биомасса – от 2.7 до 12.5 мг С/м3 при среднем объеме клеток 0.020–0.136 мкм3 (табл. 1). В поверхностном и подповерхностном слоях численность, биомасса и средний объем клеток варьировали в более узких диапазонах. В промежуточном слое наблюдалось снижение обилия и биомассы бактерий, а минимальные значения отмечены в придонном горизонте. Различия между слоями были статистически значимыми (p < 0.05). Для бактериальных клеток среднего объема отмечена обратная тенденция, наибольшими размеры клеток были в глубоководных слоях. В верхней части водной толщи количественные показатели бактериопланктона достоверно не различались, при этом диапазоны значений в АРВ были значительно шире, чем в ТАВ. В глубинных водах разброс величин был незначительным (табл. 1).
Качественный состав бактериопланктона был представлен в основном одиночными клетками, агрегированный бактериопланктон встречался редко, микроколонии и бактериальные нити в пробах воды практически отсутствовали (табл. 1). Основу микробного сообщества формировали клетки длиной менее 2 мкм и шириной 0.2–0.6 мкм; их доля в общей численности достигала 94–100%, в общей биомассе – 76–100%. Доля ультрамикроорганизмов в указанной размерной фракции в среднем составляла 69 ± 3%, снижаясь в донных водах у Земли Франца-Иосифа и Новой Земли до 53 ± 4%. Вклад крупных палочковидных клеток и агрегированного бактериопланктона в общую численность и биомассу был невысоким: в водах у Земли Франца-Иосифа их доля не превышала 5% от общей численности и 23% от общей биомассы. Отмечена тенденция к увеличению среднего объема клеток по мере продвижения с юга на север, выделялись ТАВ с доминированием мелких бактерий, а также донные воды Земли Франца-Иосифа, в которых бактериопланктон был представлен более крупными клетками (табл. 1).
Отмечена обратная корреляция общего количество бактериопланктона и численности некоторых размерных групп бактерий с концентрацией биогенных элементов (нитраты, фосфаты, силикаты), а также с соленостью (табл. 2); выявлена прямая корреляция с концентрацией растворенного кислорода. Средний объем клетки возрастал при снижении температуры воды и увеличении солености (табл. 2). Численность и биомасса бактерий демонстрировали слабую тенденцию (p > 0.05) к увеличению по мере роста температуры воды и содержания хлорофилла а в поверхностном слое (табл. 2).
Таблица 2.
Результаты корреляционного анализа между микробиологическими параметрами и показателями внешней среды в северо-восточной части Баренцева моря (октябрь 2020 г.)
| Характеристика внешней среды | Численность бактерий, тыс. кл./мл | Биомасса бактерий, мг С/м3 |
Средний объем клеток, мкм3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| общая | группа 1 | группа 2 | группа 3 | группа 4 | группа 5 | ||||
| Т, °С | 0.29 | 0.27 | –0.01 | –0.15 | 0.21 | –0.22 | 0.00 | –0.39 | |
| S, ‰ | –0.64 | –0.64 | –0.17 | –0.25 | –0.41 | –0.26 | –0.38 | 0.38 | |
| Хл а, мг/м3 | 0.08 | 0.04 | –0.10 | 0.13 | 0.02 | 0.16 | 0.13 | 0.15 | |
| Растворенный O2, мл/л | 0.62 | 0.58 | 0.15 | 0.22 | 0.27 | 0.24 | 0.35 | –0.39 | |
| Насыщение воды O2, % | 0.58 | 0.55 | 0.1 | 0.14 | 0.32 | 0.06 | 0.29 | –0.41 | |
| Концентрация, мкг моль/л |
PO4 | –0.54 | 0.27 | –0.01 | –0.15 | 0.21 | –0.22 | 0.00 | –0.39 |
| NO2 | –0.02 | –0.64 | –0.17 | –0.25 | –0.41 | –0.26 | –0.38 | 0.38 | |
| NO3 | –0.65 | 0.04 | –0.10 | 0.13 | 0.02 | 0.16 | 0.13 | 0.15 | |
| SiO3 | –0.57 | 0.58 | 0.15 | 0.22 | 0.27 | 0.24 | 0.35 | –0.39 | |
Примечание. T – температура воды; S – соленость; Хл а – концентрация хлорофилла а в поверхностном слое. Жирным шрифтом отмечены статистически значимые коэффициенты ранговой корреляции Спирмена (p < 0.05). Объем выборки n = 50 для всех показателей, кроме концентрации хлорофилла а; для хлорофилла а объем выборки n = 13. Для группы 6 анализ не проводили из-за малого объема выборки.
ОБСУЖДЕНИЕ
Сравнение средней температуры воды с многолетними данными 1958–1998 гг. (Climatic Atlas …, 2004) показало, что в 2020 г. в поверхностном слое температура воды была выше примерно на 1°С. Этот результат закономерен и соответствует ранее отмеченной тенденции к потеплению, которое регистрируется в Баренцевом море с начала 2000-х гг. (ICES, 2021). Таким образом, наши данные по распределению микробов относятся к более теплому периоду.
На исследованной акватории гидрохимические параметры варьировали в широком диапазоне. Присутствие водных масс разного генезиса, их ветровое смешение и формирование пикноклина обусловили различие в содержании биогенных элементов как в соседних локациях, так и в пределах одной станции. Распределение биогенных элементов было характерным для конца вегетационного периода: высокие концентрации фосфатов, нитратов и силикатов, но низкие – нитритов и кислорода (Жизнь …, 1985; Ecosystem …, 2009). Значительное насыщение воды кислородом в фотическом слое при низком содержании хлорофилла а указывало на преимущественное влияние физико-химических и атмосферных процессов. Повышенное содержание некоторых биогенных веществ в придонных водах во многом было связано с замедленной скоростью водообмена в глубоководных горизонтах, приводящей к накоплению нитратов, фосфатов и силикатов. Кроме того, наличие пикноклина препятствовало поступлению биогенов в поверхностные слои из-за вертикальной конвекции. Данная закономерность была отмечена ранее и в других исследованиях (Wassmann et al., 2006). Низкое содержание хлорофилла а в поверхностном слое типично для осени, так как основные продукционные процессы в пелагиали уже закончились. Ранее было показано, что динамика микроводорослей в Баренцевом море имеет ярко выраженный сезонный характер. Обилие микроводорослей и концентрация хлорофилла а в фотическом слое достигают максимума весной. К концу лета количественные показатели фитопланктона снижаются, а в осенне-зимний период содержание хлорофилла а и количество микроводорослей опускаются до минимальных значений (Ecosystem …, 2009; Макаревич, Дружкова, 2010).
Горизонтальное распределение бактериопланктона в исследуемом районе характеризовалось мозаичностью. Наблюдалась слабая тенденция к увеличению количества бактерий в АРВ. В то же время прослеживался явный тренд к снижению количества бактериопланктона с глубиной. Подобные результаты получены и для других арктических районов (Howard-Jones et al., 2002; Сажин и др., 2010; Венгер и др., 2016; Мошарова и др., 2017). Данная структура вертикального распределения микроорганизмов характерна для морей с выраженными градиентами температуры и солености. В нашем случае на большинстве станций был отмечен ярко выраженный пикноклин. Температура воды в глубоководных и поверхностных горизонтах различалась на 1.5–4.0°С. Скорее всего, именно различием температуры воды обусловлены существенные различия размерного состава морских бактерий. В придонном горизонте средний объем клеток был существенно выше, чем в поверхностном и подповерхностном слоях, располагавшихся над пикноклином. Схожая картина описана ранее для южных, центральных и восточных районов Баренцева моря (Венгер и др., 2012, 2016; Широколобова и др., 2016).
Численность и биомасса бактериопланктона в Баренцевом море и сопредельных водах варьируют в широких пределах и сильно зависят от сезона. Сопоставление опубликованных ранее сведений о количестве бактерий в Баренцевом и Карском морях с нашими данными (табл. 3) выявило следующие черты распределения бактериопланктона. Во-первых, численность бактерий в осенний период 2020 г. была сравнима с показателями, отмеченными в Баренцевом море в другие осенние периоды. Во-вторых, в летний и весенний сезоны количество бактерий по сравнению с полученными нами показателями было выше. В-третьих, в северной части Карского моря в районе, граничащем с северо-восточной частью Баренцева моря, обилие и биомасса бактериопланктона были значительно ниже. Различие весенне-летних и осенних количественных показателей бактерий связано с различием гидрологического режима, а также состояния сообществ пелагических микроводорослей (Жизнь …, 1985; Ecosystem …, 2009). Отличительной чертой Карского моря является олиготрофность его глубоководных северных районов (желоб Св. Анны), что приводит к низким количественным показателям бактериопланктона (Сажин и др., 2010; Мошарова и др., 2017).
Таблица 3.
Численность и биомасса бактериопланктона в Баренцевом и Карском морях
| Район | Период | Численность бактерий | Биомасса бактерий | Ссылка | ||
|---|---|---|---|---|---|---|
| мин–макс | средняя | мин–макс | средняя | |||
| Баренцево море | ||||||
| Северо-восток | Октябрь, 2020 г. | 220–1068 | 497 ± 27 | 9.3–51.4 | 20.5–1.3 | Наши данные |
| Юг | Ноябрь, 2013 г. | 480–1140 | 724 ± 127 | – | – | Широколобова и др., 2016 |
| Юго-восток | Август, 2010 г. | 900–4400 | 2000 ± 400 | – | – | Венгер и др., 2012 |
| Центральная часть | Июнь/июль, 1998 г. | 410–4100 | – | – | 20–33 | Howard-Jones et al., 2002 |
| Центральная часть | Август, 2010 г. | 400–4000 | 1500 ± 300 | – | – | Венгер и др., 2012, 2016 |
| Центральная часть | Ноябрь, 2013 г. | 400–760 | 523 ± 24 | – | – | Широколобова и др., 2016 |
| Восток | Август, 2010 г. | 400–1800 | 900 ± 200 | – | – | Венгер и др., 2016 |
| Северо-запад | Июль, 2003–2004 гг. |
200–3000 | – | 3.7–63.5 | 8.6–22.7 | Tammert et al., 2008 |
| Северо-запад | Май–август, 2003–2005 гг. | 80–910 | 370 ± 29 | 1.5–18.1 | 7.4 ± 5.8 | Sturluson et al., 2008 |
| Север | Август, 2010 г. | 400–2100 | 1000 ± 200 | – | – | Венгер и др., 2012 |
| Северо-восток | Август, 2009–2010 гг. |
250–3400 | 681–2434 | 4–39.1 | 13.4–30.7 | Венгер, 2011 |
| Карское море | ||||||
| Желоб Св. Анны | Сентябрь, 2007 г. | 9.8–150 | – | 0.11–7.51 | – | Сажин и др., 2010 |
| Северо-запад | Сентябрь, 2011 г. | 8.9–104.88 | 35.91 | – | – | Мошарова и др., 2017 |
| Желоб Св. Анны | Сентябрь, 2011 г. | 19.4–549.2 | 55.8–94.8 | 0.38–8.97 | 1.53–1.97 | Копылов и др., 2015 |
Корреляционный анализ показал, что численность и биомасса морских бактерий тесно связаны с концентрацией основных биогенных элементов, более высоким концентрациям которых соответствовало меньшее количество бактерий. Подобные взаимосвязи характерны и для других районов Баренцева моря (Howard-Jones et al., 2002; Sturluson et al., 2008; Ecosystem …, 2009), наиболее явно они выражены в период “цветения” фитопланктона. Как правило, обратная корреляция количества бактерий и биогенных элементов реализуется следующим образом: во время массового развития микроводорослей происходит активное поглощение нитратов, фосфатов и силикатов, что приводит к резкому снижению их концентраций в фотическом слое. Рост обилия фитопланктона ведет к соответствующему увеличению численности бактерий. В нашем случае также отмечена слабая прямая корреляции между содержанием хлорофилла а и количеством бактериопланктона.
По-видимому, дополнительным источником органического вещества как субстрата для роста и размножения бактерий в осенний период выступали другие организмы. Ранее сообщалось, что в арктических районах важную роль в течение осенне-зимнего затухания функциональной активности фитопланктона играют фотосинтетические пикоэукариоты рода Micromonas (Lovejoy et al., 2007; Terrado et al., 2008, 2011). Более мелкие виды фитопланктона лучше “справляются” с низкой доступностью питательных веществ в олиготрофных условиях (Raven, 1998). Скорее всего, гетеротрофные простейшие и копеподы также могли быть поставщиками органического вещества для бактерий. Важность протозоопланктона как источника автохтонного и аллохтонного органического вещества и потребителя бактериальной биомассы отмечена для ряда арктических районов (Sherr et al., 1997, 2003). Показано, что мелкие копеподы также значительно влияют на обилие микробных сообществ (Turner, 2004). Ранее было установлено, что в составе зоопланктона в сентябре–январе мелкие планктонные ракообразные являются доминирующей группой (Дворецкий, Дворецкий, 2015). Многие из них активны в течение всего года (Дворецкий, Дворецкий, 2015), поэтому можно предположить, что осенью они играют существенную роль в трофических цепях пелагиали, являясь источником органического углерода и прочих биогенов для морских бактерий.
Примечательно, что температура воды существенно не влияла на горизонтальное распределение бактериопланктона, однако прослеживалась значимая обратная корреляция с ее соленостью. Наблюдаемую картину можно объяснить вертикальной структурой водной толщи – более плотные и соленые воды располагались у дна, где количество бактерий было минимальным. В других исследованиях показано, что температура слабо влияла на обилие бактериопланктона в Баренцевом море независимо от сезона (Howard-Jones et al., 2002; Sturluson et al., 2008.). Это еще раз подчеркивает, что фитопланктон и другие простейшие, а также зоопланктон оказывают большее влияние на пространственно-временную динамику сообществ морских бактерий в Арктике.
По результатам работы можно сделать выводы: горизонтальное распределение бактериопланктона характеризовалось мозаичностью; отмечен явный вертикальный тренд – снижение количества бактерий и увеличение их размеров от поверхностного слоя к придонному; численность и биомасса бактериопланктона были сопоставимы с таковыми в других районах Баренцева моря в осенний период; распределение бактериопланктона определялось содержанием биогенных элементов в воде, а также структурой водных масс.
Список литературы
Венгер М.П. Бактериопланктон северо-восточной части Баренцева моря // Вестн. ЮНЦ РАН. 2011. Т. 7. № 4. С. 55–60.
Венгер М.П., Широколобова Т.И., Макаревич П.Р., Водопьянова В.В. Вирусы в пелагиали Баренцева моря // Докл. РАН. 2012. Т. 446. № 3. С. 345–349.
Венгер М.П., Копылов А.И., Заботкина Е.А., Макаревич П.Р. Влияние вирусов на бактериопланктон открытой и прибрежной части Баренцева моря // Биол. моря. 2016. Т. 42. № 1. С. 19–26.
ГОСТ 17.1.4.02-90. ВОДА. Методика спектрофотометрического определения хлорофилла а // Государственный контроль качества воды. М.: ИПК Изд-во стандартов. 2001. С. 551–563.
Дворецкий В.Г., Дворецкий А.Г. Экология зоопланктонных сообществ Баренцева моря и сопредельных вод. СПб.: Реноме. 2015. 736 с.
Жизнь и условия ее существования в пелагиали Баренцева моря. Апатиты: Изд-во Кольск. фил. АН СССР. 1985. 218 с.
Копылов А.И., Косолапов Д.Б. Микробная “петля” в планктонных сообществах морских и пресноводных экосистем. Ижевск: КнигоГрад. 2011. 332 с.
Копылов А.И., Сажин А.Ф., Заботкина Е.А., Романова Н.Д. Вириопланктон Карского моря: влияние вирусов на смертность гетеротрофных бактерий // Океанология. 2015. Т. 55. № 4. С. 620–631.
Макаревич П.Р., Дружкова Е.И. Сезонные циклические процессы в прибрежных планктонных альгоценозах северных морей. Ростов н/Д: Изд-во ЮНЦ РАН. 2010. 280 с.
Мошарова И.В., Мошаров С.А., Ильинский В.В. Особенности распределения бактериопланктона с активным метаболизмом в водах желоба Святой Анны (Карскоe море) осенью 2011 г. // Океанология 2017. Т 57. № 1. С. 128–136.
Планктон морей Западной Арктики. Апатиты: Изд-во КНЦ РАН. 1997. 352 с.
Руководство по химическому анализу морских вод. Руководящий документ. Л.: Гидрометеоиздат. 1993. 263 с.
Сажин А.Ф., Романова Н.Д., Мошаров С.А. Бактериальная и первичная продукция в водах Карского моря // Океанология. 2010. Т. 50. № 5. С. 801−808.
Таблицы растворимости кислорода в морской воде / Под ред. З.И. Мироненко. Л.: Гидрометеоиздат. 1976. 165 с.
Широколобова Т.И., Жичкин А.П., Венгер М.П. и др. Бактерии и вирусы свободной ото льда акватории Баренцева моря в период наступления полярной ночи // Докл. РАН. 2016. Т. 469. № 3. С. 383–387.
Climatic Atlas of the Arctic Seas 2004: Part I. Database of the Barents, Kara, Laptev, and White Seas – Oceanography and Marine Biology. NOAA Atlas NESDIS 58. Washington: U.S. Gov. Print. Off. 2004. 148 p.
Ecosystem Barents Sea. Trondheim: Tapir Acad. Press. 2009. 587 p.
Howard-Jones M.H., Ballard V.D., Allen A.E. et al. Distribution of bacterial biomass and activity in the marginal ice zone of the central Barents Sea during summer // J. Mar. Syst. 2002. V. 38. P. 77–91.
ICES. Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Sci. Rep. 2021. V. 3. № 77. 236 p.
Lovejoy C., Vincent W.F., Bonilla S. et al. Distribution, phylogeny, and growth of cold-adapted picoprasinophytes in arctic seas // J. Phycol. 2007. V. 43. P. 78–89.
Norland S. The relationships between biomass and volume of bacteria // Handbook of Methods in Aquatic Microbial Ecology. Boca Raton: Lewis Publ. 1993. P. 303–308.
Porter K.G., Feig Y.S. The use of DAPI for identifying and counting of aquatic microflora // Limnol. Oceanogr. 1980. V. 25. P. 943–948.
Raven J.A. Small is beautiful: the picophytoplankton // Funct. Ecol. 1998. V. 12. P. 503–513.
Sherr E.B., Sherr B.F., Fessenden L. Heterotrophic protists in the Central Arctic Ocean // Deep-Sea Res. Part II. 1997. V. 44. P. 1665–1682.
Sherr E.B., Sherr B.F., Wheeler P.A., Thompson K. Temporal and spatial variation in stocks of autotrophic and he-terotrophic microbes in the upper water column of the central Arctic Ocean // Deep-Sea Res. Part I. 2003. V. 50. P. 557–571.
Sturluson M., Nielsen T.G., Wassmann P. Bacterial abundance, biomass and production during spring blooms in the northern Barents Sea // Deep-Sea Res. Part II. 2008. V. 55. P. 2186–2198.
Tammert H., Olli K., Sturluson M., Hodal H. Bacterial biomass and activity in the marginal ice zone of the nor-thern Barents Sea // Deep-Sea Res. Part II. 2008. V. 55. P. 2199–2209.
Terrado R., Lovejoy C., Massana R., Vincent W.F. Microbial food web responses to light and nutrients beneath arctic sea ice during the winter-spring transition // J. Mar. Syst. 2008. V. 74. P. 964–977.
Terrado R., Medrinal E., Dasilva C. et al. Protist community composition during spring in an Arctic flaw lead poly-nya // Polar Biol. 2011. V. 34. P. 1901–1914.
Turner J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs // Zool. Stud. 2004. V. 43. P. 255–266.
Wassmann P., Reigstad M., Haug T. et al. Food webs and carbon flux in the Barents Sea // Prog. Oceanogr. 2006. V. 71. P. 232–287.
Дополнительные материалы отсутствуют.