

Биология моря, 2023, T. 49, № 2, стр. 127-134
Модель стохастического роста минтая Gadus chalcogrammus (Pallas, 1814)
1 Национальный научный центр морской биологии им. А.В. Жирмунского (ННЦМБ) ДВО РАН
690041 Владивосток, Россия
2 Дальневосточный федеральный университет
690001 Владивосток, Россия
* E-mail: vsukhan@mail.ru
Поступила в редакцию 27.09.2022
После доработки 24.11.2022
Принята к публикации 24.11.2022
- EDN: DVWGFD
- DOI: 10.31857/S0134347523020080
Аннотация
Предлагается математическая модель, описывающая возрастную динамику вектора средних и ковариационной матрицы признаков особей сахалинской популяции минтая Gadus chalcogrammus (Pallas, 1814). Модель базируется на уравнениях Берталанффи и Гомпертца. Ковариационная матрица слагается из двух частей: шумовой (вызванной быстрыми случайными флюктуациями условий внешней среды) и структурной (обусловленной внутрипопуляционной изменчивостью параметров, входящих в уравнения роста). Модель неплохо воспроизводит возрастную динамику распределения рыб по количественным признакам особей. Описывается возрастное увеличение, прохождение через максимум в молодом возрасте, последующее снижение дисперсий и их стабилизация на низких уровнях у длины и массы тела взрослых рыб. Объясняется возрастное снижение корреляции между длиной и массой тела.
Изучение изменчивости признаков у организмов составляет одно из важных направлений теоретической биологии. Организмы растут в процессе взросления. В этом процессе изменчивость их признаков также демонстрирует закономерную динамику. Количественное описание таких закономерностей представляется важным как в теоретическом плане (выявление общих законов, управляющих онтогенетической изменчивостью), так и в прикладных аспектах (более точные расчеты популяционной структуры, управление искусственным отбором по размерам особей).
Полезную роль здесь должны играть математические модели стохастического роста организмов. С их помощью большое количество фактической информации можно свернуть в малый набор модельных параметров, а затем работать уже не с исходными эмпирическими таблицами, а с аппроксимирующими их модельными уравнениями. Одна из моделей, претендующих на такую роль, предлагается в настоящей статье. Эта модель представляет собой дальнейшее развитие идей, изложенных в ранней работе автора (Суханов, 1980).
Построение модели
Случайный процесс роста особи опишем системой стохастических дифференциальных уравнений следующего вида:
Здесь t – время (возраст поколения); xi – значение i-го элемента в векторе, включающем m количественных признаков особи; fi и gi – функции от xi, которые будут определены далее; ni(t) – белый гауссовский шум в смысле Ито (Тихонов, Миронов, 1977) с нулевой средней, отражающий влияние быстрых случайных флюктуаций среды на рост данного i-го признака.
Согласно модели, средняя скорость роста i-го признака, описываемая функцией fi(xi), не зависит от значений остальных признаков. Это существенное упрощение модели вводится в самом начале для более понятного изложения. Материалы по росту минтая не дают оснований для построения более общей модели (Sukhanov, 2019) со взаимовлияниями признаков.
Несмотря на отсутствие детерминированных взаимодействий, признаки могут быть сцеплены друг с другом посредством статистических взаимосвязей. Один из важных типов таких взаимосвязей проявляется в коррелированности между белыми шумами ni(t) и nj(t) для признаков xi и xj. Данное обстоятельство заставляет рассматривать совокупность представленных стохастических уравнений именно как взаимосвязанную систему. Этой системе соответствует следующее уравнение Фоккера-Планка-Колмогорова:
(1)
$\frac{{\partial P}}{{\partial t}} = - \sum\nolimits_{i = 1}^m {\frac{\partial }{{\partial {{x}_{i}}}}} \left[ {{{f}_{i}}\left( {{{x}_{i}}} \right)P - \frac{1}{2}} \right.\left. {\,\sum\nolimits_{j = 1}^m {\frac{\partial }{{\partial {{x}_{j}}}}} \left( {{{\rho }_{{ij}}}{{\beta }_{i}}{{\beta }_{j}}{{g}_{i}}\left( {{{x}_{i}}} \right){{g}_{j}}\left( {{{x}_{j}}} \right)P} \right)} \right].$Здесь P = P(x1, …, xm, t) – совместная вероятностная плотность распределения признаков x1, …, xm в момент времени $t$, постоянная $\beta _{i}^{2}$ = const – интенсивность белого шума для i-го признака, ${{\rho }_{{ij}}}$= const – коэффициент корреляции между шумами для i-го и j-го признаков, причем ${{\rho }_{{ii}}} = 1$.
Для нахождения приближенного решения уравнения (1) воспользуемся методом интегральных моментов (Коздоба, 1975; Тихонов, Миронов, 1977). В результате умножения правой и левой части этого уравнения на сомножители вида ${{x}_{i}},~~x_{i}^{2},~~{{x}_{i}}{{x}_{j}}$ и последующего интегрирования по пространству состояний (x1, …, xm) исходное уравнение (1) в частных производных заменяется системой обыкновенных дифференциальных уравнений. Эти уравнения описывают возрастную динамику вектора средних арифметических и динамику ковариационной матрицы у распределения P. Для нашей модели (1) данная система имеет вид:
(2)
$\begin{gathered} \frac{{d{{E}_{i}}}}{{dt}} = {{f}_{i}}\left( {{{E}_{i}}} \right); \\ \frac{{d{{v}_{{ij}}}}}{{dt}} = {{\rho }_{{ij}}}{{\beta }_{i}}{{\beta }_{j}}{{g}_{i}}\left( {{{E}_{i}}} \right){{g}_{j}}\left( {{{E}_{j}}} \right) + \\ + \,\,\left( {\frac{{\partial {{f}_{i}}\left( {{{E}_{i}}} \right)}}{{\partial {{E}_{i}}}} + \frac{{\partial {{f}_{j}}\left( {{{E}_{j}}} \right)}}{{\partial {{E}_{j}}}}} \right){{v}_{{ij}}}, \\ \end{gathered} $где i, j = 1, …, m; Ei – средняя арифметическая для i-го признака: vij – ковариация между i-м и j-м признаками; функция fi(Ei) имеет своим аргументом “свою” среднюю арифметическую Ei, то же относится и к функциям ${{g}_{i}}\left( {{{E}_{i}}} \right)$; параметры ${{\rho }_{{ij}}},~{{\beta }_{i}},~{{\beta }_{j}}$ сохраняют прежний смысл.
В большинстве практически важных случаев распределение P можно аппроксимировать m-мерным нормальным законом. Тогда система (2) вместе с заданными начальными условиями исчерпывающе определяет возрастную динамику такого распределения. Отметим, что нормальное распределение стремится к нулю при $~{{x}_{i}} \to \pm \infty $. Тем самым обеспечивается выполнение естественных граничных условий, использованных при интегрировании уравнения (1).
Таким образом, быстрые случайные шумовые флюктуации условий внешней среды, действующие на растущий организм, приводят к возникновению особого рода изменчивости признаков. Назовем ее шумовой изменчивостью (рис. 1а). Кроме нее в дисперсионной матрице присутствует еще один компонент. Он обусловлен генетической изменчивостью и разнообразием местообитаний популяции. Подобного рода изменчивость воплощается в вариабельности параметров, входящих в уравнения роста признаков. Такую изменчивость мы будем называть параметрической или структурной (рис. 1б). Перейдем к ее модельному описанию.
Рис. 1.
Схема формирования шумовой (а) и структурной (б) изменчивости в размерах тела. Абсциссы t – возраст, ординаты x – размер.

Пусть в функции x1(t), …, xm(t), описывающие траектории признаков во времени, входит вектор параметров роста p1, …, pn. Численные значения этих параметров (коэффициентов функций) варьируют от особи к особи. Пусть Q означает ковариационную матрицу, описывающую их популяционную изменчивость. В соответствии с Томовичем и Вукобратовичем (1972), отклик величины признака xi на малые вариации l-го параметра pl назовем коэффициентом чувствительности и обозначим как ${{\varphi }_{{il}}} = {{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}.$ Выведем уравнения возрастной динамики для коэффициентов ${{\varphi }_{{il}}}$.
Вернемся к уравнению роста ${{d{{x}_{i}}} \mathord{\left/ {\vphantom {{d{{x}_{i}}} {dt}}} \right. \kern-0em} {dt}} = $ $ = {{f}_{i}}\left( {{{x}_{i}},{{p}_{1}}, \ldots {{p}_{n}}} \right)$, в котором опустим за ненадобностью слагаемое ${{g}_{i}}\left( {{{x}_{i}}} \right)$, поскольку шумовую изменчивость мы уже описали. Продифференцируем правую и левую части этого уравнения по параметру pl: ${{\partial ({{d{{x}_{i}}} \mathord{\left/ {\vphantom {{d{{x}_{i}}} {dt}}} \right. \kern-0em} {dt}})} \mathord{\left/ {\vphantom {{\partial ({{d{{x}_{i}}} \mathord{\left/ {\vphantom {{d{{x}_{i}}} {dt}}} \right. \kern-0em} {dt}})} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}} = \left( {{{\partial {{f}_{i}}} \mathord{\left/ {\vphantom {{\partial {{f}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}} \right)$ + $({{\partial {{f}_{i}}} \mathord{\left/ {\vphantom {{\partial {{f}_{i}}} {\partial {{x}_{i}}}}} \right. \kern-0em} {\partial {{x}_{i}}}})({{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}})$. Здесь использовано цепное правило. Изменим порядок дифференцирования левой части этого равенства: ${{\partial ({{d{{x}_{i}}} \mathord{\left/ {\vphantom {{d{{x}_{i}}} {dt}}} \right. \kern-0em} {dt}})} \mathord{\left/ {\vphantom {{\partial ({{d{{x}_{i}}} \mathord{\left/ {\vphantom {{d{{x}_{i}}} {dt}}} \right. \kern-0em} {dt}})} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}$ $ = {{d\left( {{{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}} \right)} \mathord{\left/ {\vphantom {{d\left( {{{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}} \right)} {dt}}} \right. \kern-0em} {dt}}$. С учетом обозначения ${{\varphi }_{{il}}} = {{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}$ получаем:
(3)
$\frac{{d{{\varphi }_{{il}}}}}{{dt}} = \frac{{\partial {{f}_{i}}}}{{\partial {{p}_{l}}}} + \frac{{\partial {{f}_{i}}}}{{\partial {{x}_{i}}}}{{\varphi }_{{il}}};\,\,\,~~i = 1,\,\, \ldots ,\,\,m;~~l = 1,\,\, \ldots ,\,\,n.$Эта система уравнений определяет возрастную динамику коэффициентов чувствительности. Дифференцированием по параметрам начальных условий для xi при t = 0 можно задать начальные условия для ${{\varphi }_{{il}}}$, так что ${{\left. {{{\varphi }_{{il}}}} \right|}_{{t = 0}}} = {{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{p}_{l}}}}} \right. \kern-0em} {\partial {{p}_{l}}}}$. Таким образом, решение уравнений (3) определяет возрастную динамику коэффициентов чувствительности ${{\varphi }_{{il}}}$.
Пусть нам известна ковариационная матрица параметров Q = ||qlh||; l, h = 1, …, n. Воспользуемся формулой переноса ошибок (Бард, 1979) и вычислим структурную ковариацию Sij между признаками xi и xj, задающую параметрическую изменчивость в кривых роста особей:
(4)
${{S}_{{ij}}} = \mathop \sum \limits_{l = 1}^n \left[ {{{\varphi }_{{il}}}{{\varphi }_{{jl}}}{{q}_{{ll}}} + \mathop \sum \limits_{h = l + 1}^n {{q}_{{lh}}}\left( {{{\varphi }_{{il}}}{{\varphi }_{{jh}}} + {{\varphi }_{{ih}}}{{\varphi }_{{jl}}}} \right)} \right].$Общая ковариационная матрица ||$\sigma _{{ij}}^{2}$|| слагается из двух компонентов: шумовой матрицы ||${{v}_{{ij}}}$|| и структурной (параметрической) матрицы ||${{S}_{{ij}}}$||:
Как отдельные слагаемые в правой части этого равенства, так и их сумма изменяются в соответствии с полученными выше уравнениями (2, 3 и 4).
Подведем итоги данного раздела. Мы разделили параметры модели на две группы. В первую группу входят параметры роста, быстро и случайно флюктуирующие во времени. Они не фигурируют в модельных уравнениях детерминированного роста явно, а задаются в форме белых гауссовских шумов. Эти параметры порождают описанную выше шумовую изменчивость в признаках организма.
Во вторую группу входят параметры, медленно изменяющиеся во времени – так, что в течение жизни особи их можно считать постоянными. Естественная вариабельность этих параметров у особей, входящих в одновозрастное поколение, вызвана генетическим и средовым разнообразиями. Она порождает структурный компонент в общей изменчивости признаков, описанный выше.
Существует и третья группа параметров роста, динамика которых по своим характерным скоростям вполне сопоставима со скоростями роста fi(Ei). В качестве примера вспомним периодические колебания в скорости роста организмов, вызванные сезонными изменениями факторов внешней среды, в первую очередь температуры и пищевой обеспеченности. Компактное модельное представление таких случайных процессов непросто. Тем не менее если их включение в модель обязательно, то это приведет к неавтономным дифференциальным уравнениям роста. Тогда внешние “среднескоростные” неслучайные или случайные возмущения должны описываться явно заданными функциями от времени. В данной работе мы не касаемся этой особой задачи.
Многомерное стохастическое обобщение модели Берталанффи
Уравнения нашей модели представлены в общем виде. Для решения реальных задач их нужно конкретизировать. Остановимся на самой популярной и простой модели Берталанффи. В этой модели скорость роста признака описывается функцией f(x) = k(a – x), где a = const – дефинитивная величина признака, к которой стремятся старые, взрослые организмы; k = const – относительная скорость затухания роста.
Функцию $g\left( x \right)$ из системы (2) зададим в следующем виде: $g\left( x \right) = {{f}^{\gamma }}\left( x \right)$, где $\gamma = {\text{const}} \geqslant 0$. Эта связь означает, что, согласно развиваемой модели, амплитуда шумовых флюктуаций у скорости роста тем выше, чем больше сама эта скорость роста. При γ = 0 шум аддитивен, и его амплитуда не зависит от f, при γ = 1 шум мультипликативен, и эта амплитуда пропорциональна f. При 0 < γ < 1 шум влияет промежуточным образом. Тогда систему (2) можно представить в следующей форме:
Решение этой системы с начальными условиями Ei = ci = const, vij = v0ij = const при t = 0 имеет следующий вид:
(7)
$\begin{gathered} {{v}_{{ij}}} = {{v}_{{0ij}}}{{e}^{{ - {{K}_{{ij}}}t}}} + \frac{{{{\rho }_{{ij}}}{{\beta }_{i}}{{\beta }_{j}}{{{\left[ {{{k}_{i}}\left( {{{a}_{i}} - {{c}_{i}}} \right){{k}_{j}}\left( {{{a}_{j}} - {{c}_{j}}} \right)} \right]}}^{\gamma }}}}{{\left( {1 - \gamma } \right){{K}_{{ij}}}}} \times \\ \times \,\,\left[ {{{e}^{{ - \gamma {{K}_{{ij}}}t}}} - {{e}^{{ - {{K}_{{ij}}}t}}}} \right]. \\ \end{gathered} $Здесь для компактности записи обозначено Kij = = ki + kj .
Теперь систему уравнений (3) для коэффициентов чувствительности также можно конкретизировать:
В этой записи коэффициентов чувствительности вместо подстрочного индекса l, обозначающего в уравнениях (3) номер того или иного параметра модели, использовано непосредственно само буквенное обозначение этого параметра pl. Так, например, ${{\varphi }_{{i{{k}_{i}}}}}$ представляет собой частную производную ${{\partial {{x}_{i}}} \mathord{\left/ {\vphantom {{\partial {{x}_{i}}} {\partial {{k}_{i}}}}} \right. \kern-0em} {\partial {{k}_{i}}}}$. Решения выписанных уравнений имеют вид:
(8)
${{\varphi }_{{i{{k}_{i}}}}} = \left( {{{a}_{i}} - {{c}_{i}}} \right)t{{e}^{{ - {{k}_{i}}t}}};\,\,\,\,~{{\varphi }_{{i{{a}_{i}}}}} = 1 - {{e}^{{ - {{k}_{i}}t}}};~\,\,\,\,~{{\varphi }_{{i{{c}_{i}}}}} = {{e}^{{ - {{k}_{i}}t}}}.$При этом$~~{{\varphi }_{{i{{k}_{j}}}}} = {{\varphi }_{{i{{a}_{j}}}}} = {{\varphi }_{{i{{c}_{j}}}}} = 0,$ если i ≠ j.
Окончательное представление модели сформировано из следующих элементов. Вектор средних арифметических определяется уравнениями (6). Общая ковариационная матрица согласно (5), слагается из шумового компонента (7) и структурного компонента. Последний вычисляется подстановкой коэффициентов чувствительности (8) вместе с матрицей параметрических ковариаций ||${{S}_{{ij}}}$|| в уравнение (4).
Подгонка параметров модели по данным роста минтая
Модель иллюстрируется материалами по стохастическому росту сахалинской популяции минтая Gadus chalcogramma. Они представляют собой данные массовой промысловой статистики, собранные в научно-поисковой экспедиции Сахалинского отделения ТИНРО в 1966 г. Данные представлены измерениями длины AC и общей массы тела по 1228 особям в возрасте от 3 до 8 лет. Самцы и самки не отличались достоверно друг от друга по изучавшимся характеристикам роста, поэтому материалы по обоим полам были объединены. Связь между длиной и массой тела в исходных единицах измерения показана на рис. 2. Надо подчеркнуть, что в логарифмическом масштабе эмпирические точки группируются вдоль прямой линии. Это говорит о том, что в первоначальных координатах длины и массы тела минтая эта регрессия описывается классической степенной зависимостью.
Рис. 2.
Связь между длиной (мм) и массой (г) тела минтая. Оси координат представлены в логарифмической шкале. Каждая точка – отдельная особь.

Проведем конкретизацию модели. Обозначим через x1 натуральный логарифм длины тела особи (мм, по Смиту), через x2 – натуральный логарифм общей массы тела (г). Средние арифметические, среднеквадратические отклонения и коэффициент корреляции между этими переменными обозначим соответственно через E1, E2, σ1, σ2, R. В качестве базовой модели для исходных признаков (непосредственно длины и массы тела) возьмем уравнение Гомпертца, которое логарифмированием зависимой переменной превращается в нужное нам уравнение Берталанффи.
Итоговая модель после завершения подгонки описывается следующими уравнениями:
(9)
$\begin{gathered} {{E}_{1}} = {{a}_{1}}(1 - {{e}^{{ - kt}}}) + {{c}_{1}}{{e}^{{ - kt}}};\,\,\,\,{{E}_{2}} = {{a}_{2}}(1 - {{e}^{{ - kt}}}) + {{c}_{2}}{{e}^{{ - kt}}}; \\ {{\sigma }_{1}} = {{\left[ {\beta _{1}^{2}{{{\left( {{{a}_{1}} - {{c}_{1}}} \right)}}^{{2\gamma }}}F + A_{1}^{2}{{{(1 - {{e}^{{ - kt}}})}}^{2}} + D_{1}^{2}{{e}^{{ - 2kt}}}} \right]}^{{\frac{1}{2}}}}; \\ {{\sigma }_{2}} = {{\left[ {\beta _{2}^{2}{{{\left( {{{a}_{2}} - {{c}_{2}}} \right)}}^{{2\gamma }}}F + A_{2}^{2}{{{(1 - {{e}^{{ - kt}}})}}^{2}}} \right]}^{{\frac{1}{2}}}}; \\ R = {{\beta }_{1}}{{\beta }_{2}}{{\left( {{{a}_{1}} - {{c}_{1}}} \right)}^{\gamma }}{{\left( {{{a}_{2}} - {{c}_{2}}} \right)}^{\gamma }}\frac{F}{{{{\sigma }_{1}}{{\sigma }_{2}}}}; \\ F = \frac{{{{k}^{{2\gamma - 1}}}}}{{2\left( {1 - \gamma } \right)}}({{e}^{{ - 2\gamma kt}}} - {{e}^{{ - 2kt}}}). \\ \end{gathered} $Модель имеет 11 параметров: k – константа относительной скорости затухания роста, оказавшаяся одинаковой для обоих признаков; γ – степенной параметр в функции $g\left( x \right) = {{f}^{\gamma }}\left( x \right)$ из системы (2); a1 и a2 – дефинитивные значения средних; c1 и c2 – начальные значения средних при t = 0; $\beta _{1}^{2},\,\,~\beta _{2}^{2}{\text{\;\;}}--$ интенсивности белого шума для 1-го и 2-го признаков; $A_{1}^{2},~A_{2}^{2}$ – структурные дисперсии параметров a1 и a2 (дисперсии дефинитивных значений признаков). Последний параметр $D_{1}^{2}$ является суммой двух слагаемых. Первое слагаемое − это начальная шумовая дисперсия, то есть параметр v011 в уравнении (7) для признака x1. Второе слагаемое − структурная дисперсия для параметра c1. Характер использованного материала не позволил оценить каждое слагаемое этой суммы по отдельности.
Слагаемое $D_{2}^{2}{{e}^{{ - 2kt}}}$ отсутствует в уравнении для ${{\sigma }_{2}}$ в системе (9). Оно не отличалось достоверно от нуля по материалам для сахалинской популяции минтая. Кроме того, в представленном списке модельных коэффициентов не фигурируют также и другие структурные дисперсии и ковариации, указанные в первоначальном полном описании модели (уравнения 4–8). Их оценки, полученные на промежуточных этапах идентификации модели, оказались плохо определены, то есть с большими ошибками. Поэтому такие коэффициенты были приравнены к нулю. Коэффициент корреляции ${{\rho }_{{12}}}$ между шумами для признаков x1 и x2 не отличался достоверно от единицы и был принят равным ей, поэтому в уравнении для R в системе (9) он отсутствует явно.
В процессе идентификации применялось последовательное упрощение модели. Исходная модель (4−8) строилась как бы “на вырост” и поэтому сначала включала в себя избыточное количество подгоняемых параметров. Исходя из фактического материала, имевшегося в нашем распоряжении, некоторые из этих параметров были определены с неприемлемо большими ошибками, а они влияют на доверительные интервалы самой модели. Поэтому на следующей итерации эти параметры из модели исключались: заменялись нулями или очевидными не подгоняемыми константами. В результате следующая версия модели упрощалась, и это приводило к уменьшению ошибок у оставшихся для оценивания параметров (это хорошо). При этом качество подгонки модели под материал, разумеется, несколько снижалось (это плохо). После достижения субъективного компромисса между этими двумя противоречащими друг другу целями, процесс идентификации модели завершился.
Подгонка параметров модели (9) проводилась с помощью градиентной процедуры Марквардта (Бард, 1979) с использованием штрафных функций для соблюдения естественных ограничений, запрещающих получение отрицательных значений у всех параметров. В качестве целевой функции, подлежавшей минимизации, выбрана остаточная сумма квадратов – сразу по всем пяти возрастным рядам переменных E1, E2, σ1, σ2, R. Такая процедура сложения здесь допустима, поскольку все переменные имеют одинаковую размерность, точнее, они безразмерны. Напомним, что признаки x1 и x2 представляют собой натуральные логарифмы длины и массы тела.
Попутно заметим, что если “обезразмеривание” комплекса признаков нецелесообразно или невозможно, стоит воспользоваться обобщенным алгоритмом для системы модельных уравнений, описанным в книге Барда (1979). При этом в качестве целевой функции полезно использовать многомерное обобщение понятия дисперсии (Животовский, 1984).
Не останавливаясь на конкретных этапах многошаговой процедуры идентификации, приведем окончательный список оптимальных оценок для всех одиннадцати параметров модели:
(10)
$\begin{gathered} k = 0.2876 \pm 0.0284,\,\,\,\,{{\beta }_{1}} = 0.04859 \pm 0.01307, \\ {{\beta }_{2}} = 0.1426 \pm 0.0188,\,\,\,\gamma = 0.7148 \pm 0.03277, \\ {{a}_{1}} = 6.339 \pm 0.040,\,\,\,{{a}_{2}} = 6.896 \pm 0.096, \\ {{c}_{1}} = {\text{ }}4.826 \pm 0.117,\,\,\,{{c}_{2}} = {\text{ }}2.627 \pm 0.272, \\ {{D}_{1}} = 0.04602 \pm 0.02564,\,\,\,{{A}_{1}} = 0.02031 \pm 0.0291, \\ {{A}_{2}} = 0.1220 \pm 0.0406. \\ \end{gathered} $Здесь после знаков ± указаны среднеквадратические ошибки оценок.
Анализ представленных результатов подгонки свидетельствует о следующем. Оценки коэффициентов k, γ, a1, a2, c1, c2 и β2 получены с неплохой точностью. Оценки для β1, A2 и D1 определены хуже: их ошибки составляют десятки процентов от самих оценок параметров. Коэффициент A1 определен плохо. Несмотря на большую ошибку, мы оставили его в списке коэффициентов модели, так как без него возрастной ряд для сигмы σ1 заметно хуже описывался модельной кривой.
Качество подгонки модели для средних арифметических оказалось неплохим. Остаточная дисперсия для переменной E1 составляет всего лишь 0.5% от общей эмпирической дисперсии и 0.6% для переменной E2. Качество подгонки для сигм несколько хуже: остаточная дисперсия равна 6.3% для σ1 и 15.4% для σ2. Возрастной ряд для коэффициента корреляции R приемлемо аппроксимируется моделью: остаточная дисперсия равна 2.0%.
Возрастная динамика модельных кривых имеет следующие характерные особенности. Средние E1 и E2 с замедлением растут во времени. При этом относительная скорость роста (параметр k) у них одинакова, а различия в углах наклона кривых вызваны разницей в начальных значениях c1 и c2 этих средних. Среднеквадратические отклонения σ1 и σ2 снижаются во времени, причем для σ2 это снижение идет более круто. Для старших поколений рыб стандартные отклонения обоих признаков стабилизируются на сравнительно невысоких дефинитивных уровнях.
Нужно еще раз напомнить, что признаки x1 и x2 представляют собой логарифмы длины (мм) и массы (г) тела рыб. Монотонное возрастное снижение дисперсий этих признаков вовсе не означает, что и в исходных шкалах (миллиметры и граммы) будет сохраняться такая же тенденция. Дисперсии непосредственно длины тела exp(x1) и массы тела exp(x2) вначале относительно быстро растут во времени и только затем, перевалив через свои максимумы, снижаются и, в конце концов, устанавливаются на своих низких дефинитивных предельных уровнях. Такая же картина обнаружена и для размеров раковины у приморского гребешка Mizuchopecten yessoensis (Jay, 1857) (Суханов, Селин, 2018).
Возрастная динамика связи длины и массы тела у минтая
На примере с ростом минтая покажем необычный и редко обсуждаемый феномен. Оказывается, что связь между признаками организма не остается постоянной во времени, но демонстрирует закономерные возрастные изменения. Модель воспроизводит такие изменения и дает объяснение этому явлению.
Коэффициент корреляции R между логарифмами длины и массы тела для сеголетков (возраст 0+) близок к единице, а затем монотонно уменьшается во времени. Согласно модельным оценкам асимптотическое (дефинитивное) значение R можно считать равным нулю.
Эта тенденция наглядно изображена на рис. 3. Здесь представлен возрастной ряд корреляционных эллипсов для x1и x2, каждый из которых охватывает суммарную вероятность, равную 90%. Отметим неплохое сходство между эмпирическим и модельным рядами этих корреляционных эллипсов.
Рис. 3.
Возрастная последовательность 90%-х корреляционных эллипсов для связи между логарифмами длины (ось x1) и логарифмами массы тела (ось x2) минтая. Сплошные эллипсы – эмпирика, пунктирные – модель. Центры эллипсов: маленькие кружки – эмпирика, крестики – модель. Возраст увеличивается снизу слева, вверх направо от 3 до 8 лет.
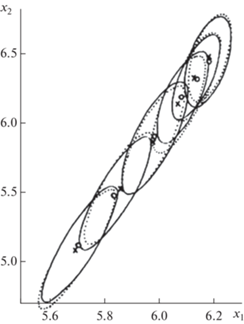
Центры эллипсов лежат на одной прямой. Эта важная особенность является следствием того, что параметр k один и тот же для обоих признаков. Точнее, различия между величинами этого параметра для обоих признаков оказались статистически недостоверными и поэтому были проигнорированы. Именно по этой причине в качестве исходной базовой модели здесь выбрано уравнение Гомпертца, а длина и масса тела представлены в логарифмических шкалах. В первоначальных арифметических шкалах эта простая линейная взаимозависимость была бы искажена нелинейностью степенной связи между непосредственно длиной и массой тела.
По мере взросления рыб корреляционные эллипсы движутся вверх направо, уменьшаются в размерах и становятся все более округлыми (рис. 3). Таким образом, в процессе роста и развития особей относительная изменчивость каждого из признаков снижается. Параллельно с этим сила связи между признаками слабеет.
Высокое значение коэффициента корреляции R на начальных этапах роста рыб и последующее его снижение объясняется следующим. Коэффициент корреляции ${{\rho }_{{12}}}$ между шумами, “раскачивающими” траектории роста признаков, практически равен единице. Это означает, что и на длину, и на массу тела рыб фактически воздействует один и тот же источник шума. По всей видимости, он непосредственно связан со случайными вариациями потока ассимилированной пищи, идущего на приросты длины и массы тела. По мере взросления рыб скорость роста каждого из этих признаков уменьшается. Вследствие этого снижается и амплитуда флюктуаций у скорости роста, поскольку параметр γ близок к единице (см. список 10). В связи с этим величина ковариации между признаками также снижается к нулю. Вместе с тем даже при полном прекращении роста обе величины дисперсий остаются ненулевыми: ${{\sigma }_{1}} \to {{A}_{1}},~\,\,{{\sigma }_{2}} \to {{A}_{2}}~$. Наличие структурных (параметрических) компонентов в общей дефинитивной изменчивости признаков, а также снижение шумовой ковариации между признаками во времени, вызванное затуханием роста приводят к возрастному падению коэффициента корреляции R.
Организм представляет собой как бы жестко образованную конструкцию, реагирующую на шумовые флюктуации внешней среды как единое целое. Эта целостность особенно ярко проявляется у быстрорастущих организмов: скорости роста отдельных признаков флюктуируют около своих средних скоростей взаимно согласованно. По мере снижения скоростей роста амплитуды этих флюктуаций снижаются, и на первый план выступает не шумовая, а структурная изменчивость параметров a1 и a2. При этом по крайней мере для нашего примера дефинитивная структурная коррелированность у взрослых особей практически отсутствует. Это позволяет прояснить механизм онтогенетической изменчивости и коррелированности признаков у организмов.
Величину коэффициента корреляции R можно рассматривать как меру цельности, связности организма как системы. Тогда естественно заключить, что молодые особи являются высокоинтегрированными системами с хорошо работающим механизмом гомеостаза. С возрастом целостность организмов и, вероятно, их регуляторные способности постепенно ослабевают в онтогенезе.
Похожий результат обнаружен и на примере двустворчатого моллюска приморского гребешка (Суханов, Селин, 2018). Использованная там версия модели оказалась несколько проще, чем описанная здесь. Тем не менее у приморского гребешка также обнаружено возрастное снижение коэффициента корреляции между признаками.
ЗАКЛЮЧЕНИЕ
Можно напомнить одну очевидную закономерность. Каждое нетривиальное развитие модели приводит к усложнению ее описания, а также к росту усилий по оцениванию ее параметров.
Простейшая детерминированная модель Берталанффи требует нескольких строк для своего описания. Для оценивания ее параметров методом Форда-Уолфорда (Мина, Клевезаль, 1976) нужно рассчитать простую линейную регрессию методом наименьших квадратов.
Для построения уравнений в первой версии модели одномерного стохастического роста (Суханов, 1980) хватило одной страницы рукописи. Кроме того, оценивание параметров потребовало обобщения метода Форда-Уолфорда, хотя и осталось в рамках метода линейной регрессии.
При построении данной стохастической многомерной версии модели понадобилось более 6 страниц. При оценивании ее параметров потребовался весьма непростой алгоритм нелинейного оценивания, который был реализован автором при помощи программы на языке Free Pascal. Изложение результатов модельной подгонки растянулось более чем на 3 страницы, и это без описания самого алгоритма расчетов, здесь он опущен за ненадобностью.
С новыми шагами количество усилий увеличивалось в геометрической прогрессии. На каждом этапе трудоемкость кратно возрастала, в среднем примерно в четыре раза.
Аналогичную закономерность автор обнаружил ранее при работе над дискретной моделью роста личинок мотыля Chironomus thummi (Суханов, Лопатин, 1988). Исходная модель – степенное уравнение роста с двумя параметрами, предложенное еще Шмальгаузеном (1935) – развернулась в систему из 6 уравнений. Их вывод занял около 10 страниц рукописи. Подгонка параметров также потребовала привлечения методов нелинейного оценивания.
Все это приводит к ограничению сложности моделей, которые можно создать “в одной голове”. Как выйти за пределы такого барьера – пока неясно.
Нельзя забывать и про доступность понимания читателями новых версий моделей. При попытках экологов полноценно их освоить требуются знания из несмежных областей науки. Недаром говорят, что каждая формула в научной публикации уменьшает количество потенциальных читателей вдвое. Тем самым затрудняется вхождение в научный арсенал новых, более сложных моделей.
Дадим резюме по обсуждаемой модели. Основанная на уравнениях Берталанффи-Гомпертца модель многомерного стохастического роста организмов проверена на данных по росту длины и массы тела сахалинской популяции минтая. Проверка показала неплохое соответствие между моделью и реальными данными.
Сформулированы два главных нетривиальных вывода. Во-первых, дисперсии длины и массы тела достигают своих максимумов в молодом возрасте. Это обусловлено разной возрастной динамикой у шумовой и структурной изменчивости признаков. Шумовая изменчивость признаков проходит максимум, а потом идет к нулю. Структурная изменчивость признаков постепенно возрастает. Во-вторых, сила межпризнаковых корреляций монотонно падает в процессе старения. Это явление, также объяснимое с помощью модели, представляется важным для теории онтогенеза и, возможно, геронтологии.
Список литературы
Бард Й. Нелинейное оценивание параметров. М.: Статистика. 1979. 349 с.
Животовский Л.А. Интеграция полигенных систем в популяциях. М.: Наука. 1984. 182 с.
Коздоба Л.А. Методы решения нелинейных задач теплопроводности. М.: Наука. 1975. 227 с.
Мина М.В., Клевезаль Г.А. Рост животных. М.: Наука. 1976. 291 с.
Суханов В.В. Стохастическая модель роста рыб // Вопр. ихтиологии. 1980. Т. 20. № 4. С. 615−624.
Суханов В.В., Лопатин О.Е. Математическое моделирование роста и развития Chironomus thummi // Деп. в ВИНИТИ 30.06.88 № 5244-В88. Владивосток. 1988. 57 с.
Суханов В.В., Селин Н.И. Модель стохастического многомерного роста Mizuhopecten yessoensis (Jay, 1857) (Bivalvia: Pectinidae) // Биол. моря. 2018. Т. 44. № 5. С. 1−8.
Тихонов В.И., Миронов М.А. Марковские процессы. М.: Сов. радио. 1977. 488 с.
Томович Р., Вукобратович М. Общая теория чувствительности. Серия “Библиотека технической кибернетики”. М.: Сов. радио. 1972. 240 с.
Шмальгаузен И.И. Определение основных понятий и методика исследования роста. Л.: Биомедгиз. 1935. 53 с.
Sukhanov V.V. Generalization of Bertalanffy-Gompertz model for multidimensional stochastic growth of organisms // Marine Biodiversity for a Healthy Ocean – Biodiversity, Functional Groups and Ocean Health. Proc. of the Russia-China Bilateral Workshop. October 10−11, 2019. Vladivostok. Russia. P. 130−132.
Дополнительные материалы отсутствуют.