

Биология моря, 2023, T. 49, № 6, стр. 417-428
Различия в питании репродуктивно изолированных экотипов косаток (Orcinus orca Linnaeus, 1758) в морях Дальнего Востока России
О. А. Филатова 1, 2, *, И. Д. Федутин 1, 2, О. А. Белонович 3, Е. А. Борисова 1, Е. В. Волкова 4, Т. В. Ивкович 5, М. Е. Исмаил 1, 6, И. Г. Мещерский 7, О. В. Титова 7, С. В. Фомин 8, О. В. Шпак 9
1 Биологический факультет Московского государственного университета им. М.В. Ломоносова
119234 Москва, Россия
2 Department of Biology, University of Southern Denmark
5230 Odense, Denmark
3 ФГБУ “Национальный парк “Командорские острова” им. С.В. Маракова
684500 с. Никольское, Камчатский край, Россия
4 ФГБУ “Кроноцкий государственный природный биосферный заповедник”
684000 Елизово, Камчатский край, Россия
5 Школа KIT
191123 Санкт-Петербург, Россия
6 Faculty of Science, Port-Said University
42526 Egypt
7 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
8 Камчатский филиал Тихоокеанского института географии ДВО РАН
683000 Петропавловск-Камчатский, Россия
9 Независимый исследователь
Харьков, Украина
* E-mail: alazorro@gmail.com
Поступила в редакцию 31.10.2022
После доработки 30.04.2023
Принята к публикации 08.06.2023
- EDN: HCZUKX
- DOI: 10.31857/S0134347523060037
Аннотация
Проведен систематический анализ видового состава добычи косатки Orcinus orca Linnaeus, 1758. Обобщены результаты наблюдений за охотой косаток на разные виды добычи и данные анализа содержимого их желудков; видовая принадлежность жертв сопоставлена с принадлежностью хищников к R- или Т-типу на основе генетического анализа. Показано, что косатки дальневосточных морей России имеют выраженную пищевую специализацию, которая коррелирует с гаплотипом контрольного региона митохондриальной ДНК. Косаток с гаплотипами NT1 и GAT, описанными ранее для плотоядных косаток T-типа из северо-восточной Пацифики (называемых также косатками Бигга), наблюдали охотящимися на морских млекопитающих, но не на рыбу. Косатки с гаплотипом SR, описанным ранее для рыбоядных косаток R-типа из северо-восточной Пацифики, охотились только на рыбу. Обнаружены два новых гаплотипа косаток Т-типа; животных с этими гаплотипами наблюдали охотящимися на крупных усатых китов. Отмечена важность традиций и социального обучения в дифференциации экологических ниш у китообразных. Передающаяся от матери к детенышам специализация к охоте на определенную добычу позволяет косаткам разных экотипов избежать пищевой конкуренции и приобрести морфологические и поведенческие адаптации, облегчающие охоту на конкретный тип добычи.
Хищники верхнего трофического уровня оказывают сильное влияние на экосистемы за счет каскадных эффектов (Pace et al., 1999). Косатка Orcinus orca – хищник верхнего трофического уровня в морских экосистемах. Ее рацион очень разнообразен: от 30-сантиметровой сельди (Similä et al., 1996) до 20-метрового синего кита (Totterdell et al., 2022). Однако для отдельных популяций или группировок характерна дифференциация экологической ниши, основанная на питании определенным типом добычи. Например, в северной части Тихого океана (Филатова и др., 2014; Ford et al., 1998; Saulitis et al., 2000) и в Антарктике (Pitman, Ensor, 2003) описаны отдельные популяции косаток, специализирующиеся на питании либо рыбой, либо морскими млекопитающими и птицами, тогда как в северной Атлантике в одной популяции могут встречаться рыбоядные группы и группы, в рацион которых входит как рыба, так и морские млекопитающие (Vongraven, Bisther, 2014; Jourdain et al., 2017; Samarra et al., 2018).
Для экотипов косаток северной части Тихого океана (R-типа, часто по традиции называемого резидентным, и Т-типа, называемого транзитным или косаткой Бигга)11 неоднократно показаны поведенческие, морфологические и генетические различия и доказана репродуктивная изоляция между экотипами, несмотря на их симпатричность на существенной части ареала (Филатова и др., 2014; Hoelzel, Dover, 1991; Hoelzel et al., 1998; Barrett-Lennard, 2000; Hoelzel et al., 2007; Morin et al., 2010; Parsons et al., 2013; Foote et al., 2016). Данные экотипы имеют существенные различия в морфологии (Baird, Stacey, 1988; Kotik et al., 2023), поведении (Morton, 1990), акустической коммуникации (Deecke et al., 2005), социальной структуре (Baird, Dill, 1996) и других аспектах биологии. В частности, недавнее исследование с помощью дронов показало, что косатки Т-типа в среднем на 7% длиннее, чем косатки R-типа той же половозрастной категории (Kotik et al., 2023). Многие авторы предлагали выделить эти экотипы в разные подвиды или даже виды на основании морфологических, поведенческих и генетических различий (Baird, Stacey, 1988; Reeves et al., 2004; Morin et al., 2010).
Для российских вод показано наличие специфичных аллелей в микросателлитных локусах, которые характерны для косаток либо R-, либо Т-типа (Филатова и др., 2014), что является однозначным свидетельством отсутствия потока генов между этими двумя формами. Пищевая специализация этих форм в российских водах описана на основе анализа стабильных изотопов, показавшего, что трофический уровень косаток Т-типа в среднем выше, чем трофический уровень косаток R-типа (Филатова и др., 2014; Borisova et al., 2020), но систематический анализ видового состава добычи косаток в морях Дальнего Востока России до сих пор не проводился.
Цели данной работы – оценить различия в питании косаток R- и Т-типов по результатам прямых наблюдений и на основе анализа содержимого желудков, а также обобщить результаты наблюдений за охотой косаток на разные виды животных, сопоставляя видовую принадлежность жертвы с принадлежностью хищников к R- или Т-типу на основе генетического анализа.
МАТЕРИАЛ И МЕТОДИКА
Данные были собраны в ходе экспедиций в разные районы Дальнего Востока. Экспедиции на п-ов Камчатка, в Чукотский автономный округ, северную часть Охотского моря и на западное побережье о-ва Беринга (Командорские о-ва) проходили в рамках Дальневосточного проекта по косатке (FEROP); на Северном и Северо-западном лежбищах северных морских котиков на о-ве Беринга – в рамках экспедиции КамчатНИРО; в западной части Охотского моря – в рамках экспедиции Института проблем экологии и эволюции им. Северцова РАН. В подавляющем большинстве случаев экспедиции базировались в полевом лагере на берегу, а выходы в море для исследования китообразных осуществлялись в светлое время суток на небольшой лодке (4−7 м) с подвесным мотором. Наблюдения за поеданием косатками останков малого полосатика в Карагинском заливе сделаны в ходе исследовательского рейса на 10-метровой яхте; за нападением на ларгу в Тауйской губе наблюдали с 14-метрового катера.
Регистрация фактов охоты
Под охотой косаток на определенный вид добычи понимали успешную охоту с визуально зарегистрированным убийством жертвы и ее поеданием; наблюдение фрагментов поедаемой косатками добычи (куски мяса, жира, внутренности, чешуя) или обнаружение добычи в желудке косатки. Если косатки находились в непосредственной близости от морских млекопитающих или скоплений рыбы и совершали действия, похожие на охоту, но убийство и поедание жертвы не были достоверно зарегистрированы, то такие случаи не учитывались.
Генетический анализ
Пробы кожи косаток для генетического анализа отбирали методом дистанционной биопсии с помощью арбалета и плавающей стрелы с пробоотборником. Полученные кусочки кожи сохраняли в 96% этаноле. Генетический анализ проводили в Кабинете методов молекулярной диагностики Института проблем экологии и эволюции им. Северцова РАН. ДНК косаток выделяли с помощью набора Diatom DNA Prep (лаборатория Изоген, Россия). Для амплификации контрольного региона митохондриальной ДНК использовали пары праймеров L15926 (O’Corry-Crowe et al., 1997) + + beluga_R1int (Meschersky et al., 2018) и beluga_intF2 [5'-AAATCACATAATATGCATGCTC-3'] + + MTCRr (Hoelzel, Green, 1998), ограничивающие два частично перекрывающихся между собой участка мтДНК длиной 416 и 734 п.н. соответственно. ДНК жертв косаток также выделяли с помощью набора Diatom DNA Prep и (в одном случае) с помощью набора QIAamp DNA Investigator Kit. Для амплификации контрольного региона митохондриальной ДНК усатых китов использовали праймеры MT4 и H00034 (LeDuc et al., 2002), ограничивающие фрагмент гена tRNA_Pro, длиной 65 п.н. и участок самого контрольного региона длиной 556 п.н. Видовую принадлежность кольчатой нерпы определяли по гену цитохрома-B митохондриальной ДНК. Для амплификации его фрагмента длиной 450 п.н. использовали праймеры L14724 и H15149 (Irwin et al., 1991). Терминирующую реакцию ставили с использованием набора BigDye Terminator v.3.1 с каждым из указанных праймеров. Нуклеотидные последовательности определяли на автоматическом анализаторе Applied Biosystems 3130 Genetic Analyzer. Выравнивание и совмещение последовательностей, а также сопоставление результирующей последовательности с вариантами, представленными в Генбанке, выполнены в программе BioEdit 7.0.5.3 (Hall, 1999) с визуальным контролем.
Определение вида рыбы по чешуе
Сразу после успешной охоты косаток на рыбу исследовательская лодка подходила к месту охоты, чтобы собрать плавающие на поверхности фрагменты добычи. В подавляющем большинстве случаев это была чешуя рыб, иногда также кусочки кожи, мускулов и других тканей. Чешуя тихоокеанских лососей (Oncorhynchus spp.) обладает видоспецифичными особенностями, которые можно использовать для идентификации вида (Mosher, 1969). В сомнительных случаях для подтверждения видовой принадлежности использовали генетические методы (Volkova et al., 2019).
РЕЗУЛЬТАТЫ
В качестве добычи косаток зарегистрированы 10 видов морских млекопитающих: финвал Balaenoptera physalis, белокрылая морская свинья Pho-coenoides dalli, малый полосатик Balaenoptera acutorostrata, гренландский кит Balaena mysticetus, акиба Pusa hispida, ларга Phoca largha, лахтак Erignathus barbatus, сивуч Eumetopias jubatus, северный морской котик Callorhinus ursinus и калан Enhydra lutris; 5 видов рыб: кижуч Oncorhynchus kisutch, кета O. keta, нерка O. nerka, чавыча O. tshawytscha и северный одноперый терпуг Pleurogrammus monopterygius, а также неопределенные до вида кальмары (табл. 1, рис. 1).
Таблица 1.
Добыча косаток в дальневосточных морях России
| Вид добычи | Метод определения добыча | Месяц, год | Регион | H* |
|---|---|---|---|---|
| Финвал Balaenoptera physalis | Генетически | Август 2021 | П-ов Камчатка, Авачинский залив | EKT |
| Белокрылая морская свинья Phocoenoides dalli | Визуально | Июль 2013 | Командорские о-ва, о-в Беринга | Нет |
| Малый полосатик Balaenoptera acutorostrata | Генетически | Август 2015 | П-ов Камчатка, Карагинский залив | Нет |
| Малый полосатик B. acutorostrata | Визуально | Июль 2010 | П-ов Камчатка, Авачинский залив | NT1 |
| Малый полосатик B. acutorostrata | Визуально | Июнь 2017 | П-ов Камчатка, Авачинский залив | Нет |
| Гренландский кит Balaena mysticetus | Визуально | Сентябрь 2015, август 2016 | Охотское море, Ульбанский залив | OST |
| Акиба Pusa hispida | Генетически | Август 2017 | Чукотский АО, прол. Сенявина | NT1 |
| Акиба P. hispida | Визуально | Август 2013 | Охотское море, Ульбанский залив | NT1 |
| Ларга Phoca largha | Визуально | Июль 2016 | Магаданская область, Тауйская губа | NT1 |
| Ларга P. largha | Визуально | Июль 2021 | П-ов Камчатка, Авачинский залив | NT1 |
| Ларга P. largha | Визуально | Август 2022 | П-ов Камчатка, Авачинский залив | Нет |
| Лахтак Erignathus barbatus | Визуально | Июль 2011 | Охотское море, Ульбанский залив | Нет |
| Калан Enhydra lutris, кальмары | Содержимое желудка (визуально) | Апрель 2020 | Командорские о-ва, о-в Беринга | GAT |
| Сивуч Eumetopias jubatus, кальмары | Содержимое желудка (визуально) | Июнь 2021 | Командорские о-ва, о-в Беринга | GAT |
| Северный морской котик Callorhinus ursinus | Визуально | Июль 2009, июнь–август 2011–2012, июль 2013–2014 | Северное лежбище, о-в Беринга | Нет |
| Северный морской котик C. ursinus | Визуально | Июль 2015, 2019 | Северо-западное лежбище, о-в Беринга | Нет |
| Кижуч Oncorhynchus kisutch (N = 51) | Морфология чешуи | Август 2011–2018 | П-ов Камчатка, Авачинский залив | SR |
| Кета O. keta (N = 18) | Морфология чешуи | Июль–август 2011–2018 | П-ов Камчатка, Авачинский залив | SR |
| Нерка O. nerka (N = 7) | Морфология чешуи | Июль–август 2012, июль 2014, 2016 | П-ов Камчатка, Авачинский залив | SR |
| Чавыча O. tshawytscha (N = 6) | Морфология чешуи | Август 2011–2014, 2016 | П-ов Камчатка, Авачинский залив | SR |
| Северный одноперый терпуг Pleurogrammus monopterygius | Морфология чешуи | Июль 2012 | П-ов Камчатка, Авачинский залив | SR |
| Северный одноперый терпуг P. monopterygius | Визуально | Август 2007–2009, август 2019 | П-ов Камчатка, Авачинский залив | SR |
Рис. 1.
Карта мест охоты косаток на разные виды добычи (показаны значками) в дальневосточных морях России. Гаплотип косаток, если он был определен, обозначен цветным полупрозрачным кругом.
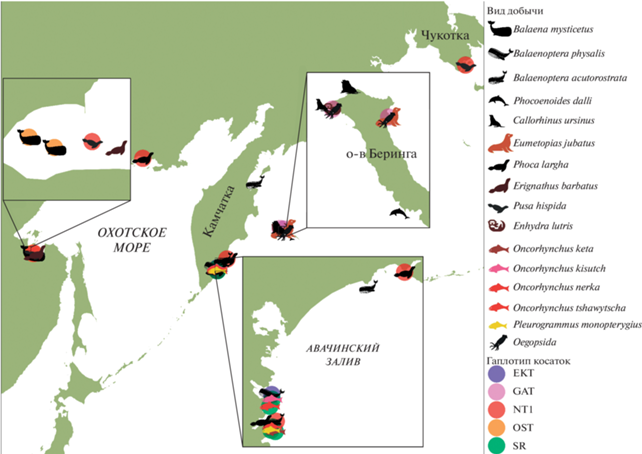
Чукотка
В прол. Сенявина на Чукотке несколько раз визуально наблюдали охоту косаток на неопределенную жертву под водой, после чего в воде оставались кровавые следы и фрагменты добычи. В одном случае из такого фрагмента удалось выделить ДНК и определить принадлежность добычи к виду кольчатая нерпа (акиба). Добывший ее одиночный самец косатки имел гаплотип NT1, номер в Генбанке DQ399075.1. Здесь и далее, если нет дополнительной ссылки, номера последовательностей в Генбанке даются по Zerbini et al. (2007), и все известные названия гаплотипов соответствуют Barrett-Lennard (2000).
п-ов Камчатка
В ходе долговременных наблюдений за косатками в Авачинском заливе (юго-восток п-ва Камчатка) неоднократно наблюдали охоту косаток как на рыбу (разные виды лосося Oncorhynchus sp. и северный одноперый терпуг), так и на морских млекопитающих: ларгу, малого полосатика и финвала.
Охотясь на лосося, косатки ныряли в разных направлениях и нередко в конце погони совершали “финальный бросок”, выпрыгивая из воды на большой скорости. При этом часто можно было увидеть рыбу, за которой гналась косатка. В отдельных случаях удавалось снять на видео, как косатка гонится за рыбой и затем поедает ее. После успешной охоты на лосося в воде оставалась отслоившаяся чешуя, по которой определяли видовую принадлежность рыб. В большинстве случаев это были кижуч (N = 51) или кета (N = 18), реже – нерка (N = 7) или чавыча (N = 6). Кроме того, неоднократно наблюдали охоту на северного одноперого терпуга в районах его нереста (мысы Опасный и Кекурный), когда рыба была видна сквозь воду, а в некоторых случаях обнаруживалась в пасти косатки (рис. 2). Все охотившиеся на рыбу косатки, у которых удалось взять биопсию (N = 50), имели гаплотип SR (номер в Генбанке DQ399077.1).
Рис. 2.
Фотографии охот косаток на разные виды добычи. а – северный одноперый терпуг; б – ларга; в, д – лосось; г – малый полосатик; е – северный морской котик.

Охоту косаток на морских млекопитающих наблюдали также в Авачинском заливе и на севере п-ва Камчатка в Карагинском заливе. В Карагинском заливе обнаружили группу косаток, поедающих неопределенную крупную добычу, при этом в толще воды плавали крупные образцы (до 30 × 30 см) ткани жертвы. Из одного такого образца взяли пробу для генетического анализа, позволившего выяснить, что добычей был кит малый полосатик. Пробу биопсии у косаток в этом случае, к сожалению, взять не удалось.
Охоту на малого полосатика наблюдали в Авачинском заливе дважды. В 2010 г. вид добычи установлен визуально по отснятым и собранным частям тела убитого кита; гаплотип косаток был определен как NT1. В 2017 г. охоту удалось наблюдать целиком: косатки таранили и топили кита, затем одна из них вырвала ему язык, и в конце кита разорвали на части. Взять пробу биопсии у этих косаток не удалось. В Авачинском заливе наблюдали охоту и на значительно более крупного усатого кита – финвала. При этом изначально охоту на кита наблюдали с туристических катеров, к моменту прибытия исследовательской лодки мертвый финвал уже был затоплен, а косатки поедали куски жира и мяса. Вид кита определили генетически по пробе одного из образцов ткани. Гаплотип этих косаток ранее не был описан и отличался на одну замену от гаплотипа GAT2 (номер в Генбанке DQ399080) из северо-восточной части Тихого океана. Этот новый гаплотип получил название EKT и загружен в GenBank под номером OP729197.
В Авачинском заливе несколько раз наблюдали охоту косаток на ларгу. В июле 2021 г. косатки ныряли в разные стороны в полосе прибоя немного южнее мыса Шипунского. Самца, прошедшего мимо лодки с добычей в пасти, сфотографировали. Гаплотип этих косаток определен как NT1. В августе 2022 г. охоту на ларгу дважды наблюдали в районе б. Русская. Косатки начинали крутиться на одном месте. В первом случае в один из таких моментов удалось сфотографировать добычу, а во втором увидеть объект охоты удалось только благодаря съемке с дрона (рис. 2б). Косатки несколько раз хватали ларгу за задние ласты, ныряли с ней, а затем отпускали. В группе присутствовал детеныш, который также участвовал в охоте. Момент гибели ларги снять не удалось. Косатки таскали останки ларги и делили их между собой.
Индивидуальная идентификация косаток по фотографиям показала, что охотившиеся на рыбу семьи косаток с гаплотипом SR никогда не взаимодействовали с группами косаток, охотившихся на морских млекопитающих и имевших гаплотипы NT1 и EKT.
Командорские острова
На Командорских о-вах визуально наблюдать охоту на рыбу не удалось, однако неоднократно отмечали случаи, когда косатки, имевшие гаплотип SR, на 3−4 мин ныряли на глубину, издавая при этом большое количество эхолокационных щелчков, что могло свидетельствовать об охоте на рыбу (морских млекопитающих при этом поблизости не наблюдалось).
В этом регионе визуально отмечена охота косаток на белокрылую морскую свинью и на северного морского котика. Охоту на белокрылую морскую свинью наблюдали у западного побережья о-ва Беринга напротив б. Лисинская. Во время охоты косатка атаковала морскую свинью снизу, полностью вылетая вертикально из воды. Затем две косатки совместно поедали добычу, сняв кожу с жировым слоем (даже мелон был аккуратно выкушен из головы) и выдрав позвоночник с ребрами и плотью. Голову и внутренности не поедали. Собрав остатки трапезы косаток, мы определили, что морская свинья была молодой, так как ее зубы едва показались из десен.
Напротив Северного и Северо-западного лежбищ северного морского котика неоднократно наблюдали охоту косаток на этих млекопитающих. Косатки подбрасывали морских котиков в воздух ударами хвоста, что зафиксировано на фотографиях (рис. 2е). К сожалению, пробы биопсии ни у этих косаток, ни у группы, охотившейся на белокрылую морскую свинью, взять не удалось. Интересно, что по результатам фотоидентификации только три из 24 косаток, идентифицированных возле Северного и Северо-Западного лежбищ, повторно встречены у юго-западного побережья о-ва Беринга, хотя там, в общей сложности, были определены 18 косаток Т-типа.
У двух косаток, найденных мертвыми на побережье о-ва Беринга, исследовано содержимое желудков. В желудке косатки, обнаруженной в апреле 2020 г. недалеко от мыса Западный, найдены останки шести каланов (еще один калан застрял в пищеводе), а также клювы и глаза кальмаров (подробнее об этом случае см. Fomin et al. (2023). В кишечнике косатки, обнаруженной в б. Столбовая в июне 2021 г., найдены клювы кальмаров и шерсть сивуча (видовая принадлежность определена по окраске шерсти). Обе косатки имели гаплотип GAT (номер в Генбанке DQ399081.1).
Охотское море
В северной и западной частях прибрежной зоны Охотского моря косатки охотились только на морских млекопитающих: акибу, ларгу, лахтака и гренландского кита. Достоверной охоты на рыбу не наблюдали.
Охоту на ларгу наблюдали в центральной части Тауйской губы недалеко от берега напротив длинной песчаной косы. Косатка схватила плававшую возле исследовательской лодки ларгу и утащила ее под воду, где разорвала жертву на куски.
Охоту на акибу, лахтака и гренландского кита наблюдали в Ульбанском заливе. При наблюдениях охоты на акибу и лахтака, косаток заметили, когда оба животных уже были убиты и разорваны на части. Их видовую принадлежность удалось определить по окраске шкуры (акиба) и форме головы (лахтак). Во время атак на гренландских китов косатки таранили жертву и пытались ее утопить.
Косатки, охотившиеся на ластоногих, имели гаплотип NT1, а охотившиеся на гренландских китов – гаплотип, не описанный в северо-восточной части Тихого океана и отличавшийся от гаплотипа NT1 на одну замену (он получил название OST и загружен в GenBank под номером OP729198). Отмечено, что при поедании убитого гренландского кита группа добывших кита косаток с гаплотипом OST взаимодействовала с подошедшей позже группой косаток с гаплотипом NT1. Модальность этих взаимодействий до конца не ясна: при появлении чужаков косатки-китобои сначала быстро двинулись им навстречу, но “чужие” прошли место кормежки, и “хозяева” вернулись к киту, затем “чужие” также подошли к месту, где находилась туша кита. Через некоторое время между группой, добывшей кита, и подошедшими особями, наблюдались активные социальные взаимодействия, в том числе плавание брюхом к брюху, что в некоторых случаях интерпретируется как попытка спаривания.
ОБСУЖДЕНИЕ
Специализация является одним из главных направлений биологической эволюции (Северцов, 1987). Мы показали, что косатки, обитающие в дальневосточных морях России, имеют выраженную специализацию по пищевым предпочтениям, которая коррелирует с гаплотипом контрольного региона митохондриальной ДНК. Косаток с гаплотипами NT1 и GAT, описанными ранее для плотоядных косаток T-типа из северо-восточной Пацифики (называемых также косатками Бигга), наблюдали охотящимися на морских млекопитающих, но не на рыбу. Напротив, косаток с гаплотипом SR, описанным ранее для рыбоядных косаток R-типа из того же района, наблюдали охотящимися только на рыбу. Кроме того, мы обнаружили два новых гаплотипа (в Авачинском заливе и в Охотском море), близких к известным гаплотипам плотоядных косаток T-типа; животные с этими гаплотипами охотились на крупных усатых китов (финвала и гренландского).
В предыдущих работах, посвященных изучению питания косаток в дальневосточных морях России, доказательством охоты на определенную добычу нередко считался сам факт нахождения косаток в непосредственной близости от морских млекопитающих или скоплений рыбы (например, Томилин, 1957; Болтнев и др., 2017). Это породило ошибочное мнение, что косатки дальневосточных морей не имеют определенной пищевой специализации и свободно переходят от питания рыбой к питанию морскими млекопитающими. Такие сообщения, как правило, опирались на экспертные оценки наблюдателей, которые, например, наблюдали косаток в устье нерестовой реки, где в это время присутствовали скопления лосося и тюлени, и из этого следовал вывод, что косатки охотятся и на тех, и на других. Мы не наблюдали ни одного факта питания одних и тех же косаток и рыбой, и морскими млекопитающими. При этом показано, что в диету плотоядных косаток входят головоногие моллюски, так как клювы и глаза кальмаров были найдены нами в желудках косаток вместе с останками каланов и сивучей. В литературе также неоднократно описаны подобные случаи (Rice, 1968; Yamada et al., 2007; Hanson, Walker, 2014).
Согласно литературным данным, в водах Чукотки косатки могут нападать на серого кита Eschrichtius robustus, белуху Delphinapterus leucas, моржа Odobenus rosmarus (см. Крюкова и др., 2012; Melnikov, Zagrebin, 2005), у побережья о-ва Сахалин – на ларгу (Бобков и др., 2015), в водах о-ва Медный они охотились на морских котиков (Мамаев, Бурканов, 2006; Рязанов и др., 2011), а в Охотском море объедали палтуса Reinhardtius hippoglossoides с ярусов (Корнев и др., 2014; Belonovich et al., 2021). Однако не описано ни одного достоверного случая, когда в северной части Тихого океана одни и те же косатки добывали и поедали и рыбу, и морских млекопитающих, как не отмечено случаев одновременного обнаружения в их желудках останков рыб и морских млекопитающих. Например, Зенкович (1938) упоминает только наличие останков дельфинов и морских котиков в желудке косатки у о-вов Прибылова и останков моржей и лахтаков в желудках двух косаток в Анадырском заливе. Томилин (1957) также не упоминает об одновременном присутствии в желудке косаток северной Пацифики рыб и морских млекопитающих. Иванова (1961) и Бетешева (1961) обнаружили в желудках 21 косатки из района Курильских о-вов только фрагменты кальмаров и рыб.
Японские исследователи представили отчет о содержимом желудков тихоокеанских косаток, проанализировав 364 желудка косаток, добытых в ходе китобойного промысла в Японии в 1948−1957 гг. (Nishiwaki, Handa, 1958). К сожалению, в этой работе приводятся только общие сведения о видовом составе добычи, и нет данных по содержимому отдельных желудков, поэтому неясно, были ли обнаружены останки рыб и морских млекопитающих в одних и тех же желудках. На основании данных о длине животных и содержимом желудков высказано предположение, что молодые косатки могут питаться рыбой или кальмарами, а более взрослые – дельфинами или любой другой добычей (Nishiwaki, Handa, 1958). Однако приведенный в этом отчете рисунок (рис. 6 ) указывает на небольшой объем выборки (желудки всего семи особей косаток длиной менее 4.3 м), что может объяснять отсутствие среди добычи тюленей, которых было немного и в желудках косаток других размерных категорий, питавшихся в том числе и дельфинами. Доли разных типов добычи в желудках более крупных молодых животных несущественно отличались от содержимого желудков взрослых косаток.
Нам известна единственная работа, в которой упоминается одновременное присутствие останков рыб и морских млекопитающих в желудке северотихоокеанской косатки (Hanson, Walker, 2014). В желудках косаток из северо-восточной части Тихого океана эти авторы обнаружили только останки кальмаров и морских млекопитающих, и лишь в желудке одной, убитой в 1988 г. в водах Японии косатки найдены кальмары, три вида рыб и останки белокрылой морской свиньи. Возможно, в японских водах действительно существует экотип, диета которого включает и рыб, и морских млекопитающих. Однако, учитывая, что содержимое желудка было собрано инспектором-наблюдателем на промысле за 26 лет до публикации и переслано из Японии в США одному из авторов, к этим данным следует относиться с осторожностью. Известно, что желудки косаток, затертых льдами в водах северной Японии, содержали только останки тюленей и кальмаров (Yamada et al., 2007).
В результатах исследований, проведенных в северо-восточной части Тихого океана, также нет упоминаний об одновременном присутствии останков рыб и морских млекопитающих в желудке одной и той же косатки. Анализ желудков восьми косаток с Аляски и из Калифорнии показал, что два из них содержали только останки рыб, а шесть – лишь останки морских млекопитающих и кальмаров (Rice, 1968). При исследовании содержимого желудков 14 косаток, найденных мертвыми на побережье Британской Колумбии с 1973 по 1989 гг., в восьми желудках обнаружены останки рыб, а в трех – останки морских млекопитающих (Ford et al., 1998). С 1990 по 2001 гг. на побережье Аляски обнаружены 12 косаток, содержимое желудков которых также описано (Heise et al., 2003). Один из желудков был пуст, в семи содержались останки морских млекопитающих, рыба в них не обнаружена. В одном желудке найдены клювы головоногих моллюсков и кайра, в остальных – обнаружены останки рыб и несъедобные предметы. В желудке косатки, найденной на Аляске в 2003 г., идентифицированы останки калана, обыкновенного тюленя, речной выдры, морских птиц и осьминога, а также камни, раковины моллюсков и водоросли (Vos et al., 2006). Останки рыб в желудке не обнаружены.
Корреляция между пищевыми предпочтениями косаток и гаплотипом митохондриальной ДНК указывает на то, что навыки охоты на определенную добычу передаются по материнской линии. Поскольку митохондриальная ДНК передается потомкам только через яйцеклетку, но не через сперматозоид, то и гаплотипы митохондриальной ДНК наследуются по материнской линии. Известно, что семьи косаток основаны на родстве по материнской линии. У косаток R-типа социальная структура исключительно стабильна – детеныши обоих полов всю жизнь остаются в материнской группе (Bigg et al., 1990; Ivkovich et al., 2010). У косаток Т-типа состав групп более пластичен, однако детеныши также остаются с матерью как минимум до достижения 10–15 лет (Baird, Dill, 1996). Охотничьи навыки детеныши косаток перенимают от матери и других членов своей семьи (Guinet, 1991). Это способствует дифференциации экологической ниши, когда часть обитающих в определенном регионе косаток питается одним типом добычи, а часть – другим.
Дифференциация экологической ниши приводит к снижению конкуренции. Кроме того, специализация на определенном типе добычи позволяет охотиться более эффективно, так как для охоты на морских млекопитающих и рыбу необходимы противоположные морфологические и поведенческие адаптации. Плотоядным косаткам выгоднее быть более крупными и плотно сложенными, а рыбоядным – более легкими и маневренными. Показано, что плотоядные косатки Британской Колумбии и штата Вашингтон имели более широкую голову и в среднем были длиннее, чем обитающие в том же районе рыбоядные (Kotik et al., 2023). Различия в социальной структуре и поведении также связывают с типом добычи. Плотоядные косатки молчаливы и чаще держатся небольшими группами, которые жертве сложнее заметить, и лишь иногда собираются большими скоплениями для охоты на крупных китов или социального поведения. Рыбоядные косатки передвигаются большими семьями и часто кричат, так как рыбы не в состоянии оценить присутствие хищников по этим признакам (Baird, Dill, 1996). В северной Атлантике разделение косаток по пищевым предпочтениям проходит не на межпопуляционном уровне, как в Тихом океане, а внутри популяций. Однако семьи, которые регулярно охотятся на тюленей, демонстрируют те же поведенческие особенности, что и тихоокеанские косатки Т-типа: передвигаются молча и небольшими группами (Jourdain et al., 2017).
Определенный уровень экологической диверсификации существует, по-видимому, и внутри экотипов. В частности, мы обнаружили, что группа косаток-“китобоев”, которых неоднократно наблюдали охотившимися на гренландских китов в Ульбанском заливе, имела отдельный гаплотип контрольного региона митохондриальной ДНК – OST, не зарегистрированный в северо-восточной части Тихого океана и описанный ранее нашей группой в рамках работы по анализу географического распределения гаплотипов полных митохондриальных геномов косаток Тихого океана (Filatova et al., 2018, номер в Генбанке MH062792). При этом косатки с гаплотипом OST свободно взаимодействовали с другими косатками Т-типа с распространенным гаплотипом NT1 и, очевидно, принадлежали к одной с ними популяции. По-видимому, склонность к охоте на китов в данном случае совпала с отличающимся от других косаток в популяции гаплотипом и независимо, но параллельно с ним, передавалась по материнской линии (гаплотип – генетически, а традиция охоты – путем социального обучения).
Интересен также факт обнаружения косаток с гаплотипом GAT на Командорских о-вах. Согласно результатам нашего анализа биопсий и данным Парсонс с соавторами (Parsons et al., 2013), ранее в этом районе встречались только косатки с гаплотипом NT3 (номер в Генбанке GU187157), а гаплотип GAT встречался преимущественно на восточных Алеутских о-вах, в заливе Аляска и на центральных Алеутах. Две косатки с этим гаплотипом были найдены выброшенными на берег о-ва Беринга в весенний период, в их желудках обнаружены останки морских млекопитающих и кальмаров. Возможно, такое различие связано с сезонностью присутствия косаток разных матрилиний: косатки с гаплотипом NT3 нагуливались в водах Командорских о-вов в летний период, когда проходил наш полевой сезон, а группы с гаплотипом GAT посещали этот район весной.
В целом, наша работа иллюстрирует важность традиций и социального обучения в дифференциации экологических ниш у китообразных. Передающаяся от матери к детенышам специализация к охоте на определенную добычу позволяет косаткам разных экотипов избежать пищевой конкуренции и приобрести морфологические и поведенческие адаптации, облегчающие охоту на конкретный тип животных. По-видимому, такая специализация адаптивна лишь в тех случаях, когда биомасса добычи каждого типа достаточно высока, поэтому она описана только в продуктивных водах высоких широт (северная часть Тихого океана и Антарктика, в меньшей степени северная Атлантика). В то же время в других районах, особенно в тропических и субтропических водах, косатки нередко потребляют и рыбу, и морских млекопитающих, не формируя определенной пищевой специализации (de Bruyn et al., 2013). Наличие пищевой специализации у косаток дальневосточных морей России необходимо учитывать при разработке мер охраны и устойчивого использования их популяций, а также при расчете потребляемой ими биомассы разных типов добычи в моделях морских трофических сетей.
Список литературы
Бетешева Е.И. Питание промысловых китов Прикурильского района // Китообразные дальневосточных морей: Тр. Ин-та морфологии животных им. А.М. Северцова. 1961. Т. 34. С. 7–12.
Бобков А.В., Стародымов С.П., Иваненко С.Ю. Нападения косаток на морских млекопитающих у северо-восточного побережья острова Сахалин // Морские млекопитающие Голарктики. Сборник научных трудов по материалам VIII международной конференции. 2015. Т. 1. С. 61−70.
Болтнев А.И., Жариков К.А., Сомов А.Г., Сальман А.Л. Спутниковое слежение за косатками в Охотском море в летне-осенний период 2015 г. // Тр. ВНИРО. 2017. Т. 168. С. 62–73.
Зенкович Б.А. О косатке или ките-убийце (Grampus orca Lin.) // Природа. 1938. Т. 4. С. 109–112.
Иванова Е.И. О тихоокеанской косатке (Orcinus orca L.) // Тр. Ин-та морфологии животных им. А.М. Северцова АН СССР. 1961. Т. 34. С. 205–215.
Корнев С.И. Белонович О.А., Никулин С.В. Косатки (Orcinus orca) и промысел черного палтуса (Reinhardtius hippoglossoides) в Охотском море // Сб. науч. тр. Камчат. НИИ рыб. хоз-ва и океанографии. 2014. Т. 34. С. 3550–3561.
Крюкова Н.В., Крученкова Е.П., Иванов Д.И. Охота косаток (Orcinus orca) на моржей (Odobenus rosmarus divergens) в районе косы Рэткын, Чукотка // Зоол. журнал. 2012. Т. 91. С. 734−734.
Мамаев Е.Г., Бурканов В.Н. Косатки (Orcinus orca) и северные морские котики (Callorhinus ursinus) Командорских о-вов: формирование пищевой специализации? // Морские млекопитающие Голарктики. Сборник научных трудов по материалам IV международной конференции. 2006. С. 347–351.
Рязанов С.Д., Мамаев Е.Г., Бородавкина М.В. и др. Наблюдения за активностью хищных косаток Orcinus orca в районе о. Медного (Командорские острова) в 2011 г. // Сохранение биоразнообразия Камчатки и прилегающих морей. Тезисы докладов XII международной научной конференции, посвященной 300-летию со дня рождения С.П. Крашенинникова. 2011. С. 261−263.
Северцов А.С. Основы теории эволюции // М.: Изд-во МГУ. 1987. 320 с.
Томилин А.Г. Китообразные, род косатки // Звери СССР и прилежащих стран. Т. 9. М: Изд-во АН СССР. 1957. С. 643–665.
Филатова О.А., Борисова Е.А., Шпак О.В. и др. Репродуктивно изолированные экотипы косаток Orcinus orca в морях Дальнего Востока России // Зоол. журнал. 2014. Т. 93. С. 1345–1353.
Baird R.W., Dill L.M. Ecological and social determinants of group size in transient killer whales // Behav. Ecol. 1996. V. 7. P. 408–416.
Baird R.W., Stacey P.J. Variation in saddle patch pigmentation in populations of killer whales (Orcinus orca) from British Columbia, Alaska, and Washington State // Can. J. Zool. 1988. V. 66. P. 2582–2585.
Barrett-Lennard L.G. Population structure and mating patterns of killer whales (Orcinus orca) as revealed by DNA analysis // Ph.D. Diss. Univ. of British Columbia. Vancouver, British Columbia. 2000.
Belonovich O.A., Agafonov S.V., Matveev A.A., Kalugin A.A. Killer whale (Orcinus orca) depredation on longline groundfish fisheries in the northwestern Pacific // Polar Biol. 2021. V. 44. P. 2235–2242.
Bigg M.A., Olesiuk P.F., Ellis C.M. et al. Social organization and genealogy of resident killer whales (Orcinus orca) in the coastal waters of British Columbia and Washington State // Rep. Int. Whaling Comm. 1990. Spec. issue 12. P. 383–405.
Borisova E.A., Filatova O.A., Fedutin I.D. et al. Ecotype and geographical variation in carbon and nitrogen stable isotope values in western North Pacific killer whales (Orcinus orca) // Mar. Mamm. Sci. 2020. V. 36. P. 925–938.
de Bruyn P.J.N., Tosh C.A., Terauds A. Killer whale ecotypes: is there a global model? // Biol. Rev. Cambridge Philos. Soc. 2013. V. 88. P. 62−80.
Deecke V.B., Ford J.K., Slater P.J. The vocal behaviour of mammal-eating killer whales: communicating with costly calls // Anim. Behav. 2005. V. 69. P. 395−405.
Filatova O.A., Borisova E.A., Meschersky I.G. et al. Coloni-zing the wild west: low diversity of complete mitochondrial genomes in western North Pacific killer whales suggests a founder effect // J. Hered. 2018. V. 109. P. 735–743.
Fomin S.V., Fedutin I.D., Borisova E.A. et al. Sea otters (Enhydra lutris) found in stomach of a stranded killer whale (Orcinus orca) in the Commander Islands, Western North Pacific // Aquat. Mamm. 2023. V. 49. P. 462–467.
Foote A.D., Vijay N., Ávila-Arcos M.C. et al. Genome-culture coevolution promotes rapid divergence of killer whale ecotypes // Nat. Commun. 2016. V. 7. Art. № 11693.
Ford J.K., Ellis G.M., Barrett-Lennard L.G. et al. Dietary specialization in two sympatric populations of killer whales (Orcinus orca) in coastal British Columbia and adjacent waters // Can. J. Zool. 1998. V. 76. P. 1456–1471.
Guinet C. Intentional stranding apprenticeship and social play in killer whales (Orcinus orca) // Can. J. Zool. 1991. V. 69. P. 2712–2716.
Hall T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT // Nucleic Acids Symp. Ser 1999. V. 41. P. 95–98.
Hanson M.B., Walker W.A. Trans-Pacific consumption of cephalopods by North Pacific killer whales (Orcinus orca) // Aquat. Mamm. 2014. V. 40. P. 274–284.
Heise K., Barrett-Lennard L.G., Saulitis E. et al. Examining the evidence for killer whale predation on Steller sea lions in British Columbia and Alaska // Aquat. Mamm. 2003. V. 29. P. 325–334.
Hoelzel A.R., Dahlheim M., Stern S.J. Low genetic variation among killer whales (Orcinus orca) in the eastern North Pacific and genetic differentiation between foraging specialists // J. Hered. 1998. V. 89. P. 121–128.
Hoelzel A.R., Dover G.A. Genetic differentiation between sympatric killer whale populations // J. Hered. 1991. V. 66. P. 191–195.
Hoelzel A.R., Green A. PCR protocols and population ana-lysis by direct DNA sequencing and PCR-based DNA fingerprinting // Molecular genetic analysis of populations, a practical approach: 2nd ed. 1998. Oxford: Oxford Univ. Press. P. 201–235.
Hoelzel A.R., Hey J., Dahlheim M.E. et al. Evolution of po-pulation structure in a highly social top predator, the killer whale // Mol. Biol. Evol. 2007. V. 24. P. 1407–1415.
Irwin D.M., Kocher T.D., Wilson A.C. Evolution of the cytochrome b gene of mammals // J. Mol. Evol. 1991. V. 32. P. 128–144.
Ivkovich T.V., Filatova O.A., Burdin A.M. et al. The social organization of resident-type killer whales (Orcinus orca) in Avacha Gulf, Northwest Pacific, as revealed through association patterns and acoustic similarity // Mamm. Biol. 2010. V. 75. P. 198–210.
Jourdain E., Vongraven D., Bisther A., Karoliussen R. First longitudinal study of seal-feeding killer whales (Orcinus orca) in Norwegian coastal waters // PLoS One. 2017. V. 12. e0180099.
Kotik C., Durban J.W., Fearnbach H., Barrett-Lennard L.G. Morphometrics of mammal-eating killer whales from drone photogrammetry, with comparison to sympatric fish-eating killer whales in the eastern North Pacific // Mar. Mamm. Sci. 2023. V. 39. P. 42–58.
LeDuc R.G., Weller D.W., Hyde J. et al. Genetic differences between western and eastern gray whales (Eschrichtius robustus) // J. Cetacean Res. Manag. 2002. V. 4. P. 1–5.
Melnikov V.V., Zagrebin I.A. Killer whale predation in coas-tal waters of the Chukotka Peninsula // Mar. Mamm. Sci. 2005. V. 21. P. 550–556.
Meschersky I.G., Chernetsky A.D., Krasnova V.V. et al. Mitochondrial lineages of the beluga whale Delphinapterus leucas in the Russian Arctic // Biol. Bull. 2018. V. 45. P. 147–154.
Morin P.A., Archer F.I., Foote A.D. et al. Complete mitochondrial genome phylogeographic analysis of killer whales (Orcinus orca) indicates multiple species // Genome Res. 2010. V. 20. P. 908–916.
Morton A.B. A quantitative comparison of the behavior of resident and transient forms of the killer whale off the central British Columbia coast // Rep. Int. Whaling Comm. 1990. Spec. issue 12. P. 245–248.
Mosher K.H. Identification of Pacific salmon and steelhead trout by scale characteristics // U.S. Bur. Commer. Fish. Circ. 317. 1969.
Nishiwaki M., Handa C. Killer whales caught in the coastal waters of Japan for recent 10 years // Sci. Rep. Whales Res. Inst. (Tokyo). 1958. V. 13. P. 53–83.
O’Corry-Crowe G.M., Suydam R.S., Rosenberg A. et al. Phylogeography, population structure and dispersal patterns of the beluga whale Delphinapterus leucas in the western Nearctic revealed by mitochondrial DNA // Mol. Ecol. 1997. V. 6. P. 955–970.
Pace M.L., Cole J.J., Carpenter S.R., Kitchell J.F. Trophic cascades revealed in diverse ecosystems // Trends Ecol. Evol. 1999. V. 14. P. 483–488.
Parsons K.M., Durban J.W., Burdin A.M. et al. Geographic patterns of genetic differentiation among killer whales in the northern North Pacific // J. Hered. 2013. V. 104. P. 737–754.
Pitman R.L., Ensor P. Three forms of killer whales (Orcinus orca) in Antarctic waters // J. Cetacean Res. Manag. 2003. V. 5. P. 131–140.
Reeves R.R., Perrin W.F., Taylor B.L. et al. Report of the workshop on shortcomings of cetacean taxonomy in relation to needs of conservation and management; April 30–May 2, 2004, La Jolla, California. 2004.
Rice D.W. Stomach contents and feeding behaviour of killer whales in the eastern North Pacific // Nor. Hvalfangst-Tid. 1968. V. 57. P. 35–38.
Samarra F.I., Bassoi M., Béesau J. et al. Prey of killer whales (Orcinus orca) in Iceland // PloS One. 2018. V. 13. e0207287.
Saulitis E., Matkin C., Barrett-Lennard L. et al. Foraging strategies of sympatric killer whale (Orcinus orca) po-pulations in Prince William Sound, Alaska // Mar. Mamm. Sci. 2000. V. 16. P. 94–109.
Similä T., Holst J.C., Christensen I. Occurrence and diet of killer whales in northern Norway: seasonal patterns re-lative to the distribution and abundance of Norwegian spring-spawning herring // Can. J. Fish. Aquat. Sci. 1996. V. 53. P. 769–779.
Totterdell J.A., Wellard R., Reeves I.M. et al. The first three records of killer whales (Orcinus orca) killing and eating blue whales (Balaenoptera musculus) // Mar. Mamm. Sci. 2022. V. 38. P. 1286–1301.
Volkova E.V., Ivkovich T.V., Shitova M.V. et al. The summer diet of fish-eating killer whales in Avacha Gulf of Kamchatka: are there any preferences? // Mamm. Biol. 2019. V. 97. P. 72–79.
Vongraven D., Bisther A. Prey switching by killer whales in the north-east Atlantic: observational evidence and experimental insights // J. Mar. Biol. Assoc. U.K. 2014. V. 94. P. 1357–1365.
Vos D.J., Quakenbush L.T., Mahoney B.A. Documentation of sea otters and birds as prey for killer whales // Mar. Mamm. Sci. 2006. V. 22. P. 201–205.
Yamada T.K., Uni Y., Amano M. et al. Biological indices obtained from a pod of killer whales entrapped by sea ice off northern Japan // Pap. IWC Sci. Comm. Meet. Anchorage, Alaska. 2007. 15 p.
Zerbini A.N., Waite J.M., Durban J.W. et al. Estimating abundance of killer whales in the nearshore waters of the Gulf of Alaska and Aleutian Islands using line-transect sampling // Mar. Biol. 2007. V. 150. P. 1033–1045.
Дополнительные материалы отсутствуют.
Инструменты
Биология моря