

Биология внутренних вод, 2019, № 2-2, стр. 61-67
Реконструкция естественного хода природных процессов по данным комплексного изучения донных отложений оз. Орон (Восточная Сибирь)
1 Лимнологический институт Сибирского отделения Российской академии наук
664033 Иркутск, Улан-Баторская, д. 3, Россия
* E-mail: deschampsia@yandex.ru
Поступила в редакцию 19.10.2017
После доработки 22.06.2018
Принята к публикации 01.08.2018
Аннотация
На основе радиометрического, палинологического анализа и изучения останков личинок хирономид (Insecta: Diptera) донных отложений горного оз. Орон проведена реконструкция изменения абиотических и биотических условий в озере и на сопредельных территориях. Высказано предположение, что ~800 лет исследованной истории оз. Орон смены “холодных” и “теплых” фаз происходили в 200-летнем природном цикле.
ВВЕДЕНИЕ
Биотические компоненты высокогорных озер очень чувствительны к изменениям климата, антропогенной нагрузке и связанным с ними изменением окружающей среды. Донные осадки таких озер наиболее пригодны для палеоклиматических реконструкций с разрешением в год–десятилетие [14, 16]. Объектом наших исследований послужило оз. Орон, внесенное в 2002 г. в перечень объектов ЮНЕСКО, подлежащих изучению в Евразии. Несмотря на это, информация о нем недостаточна.
Цель работы – реконструкция локальных и региональных сукцессий биологических, экологических, климатических условий на основе проведения исследования пыльцы и макроостанков личинок хирономид, содержащихся в горизонтах осадочного чехла озера.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Озеро Орон – одно из наиболее крупных озер, расположенное в северо-восточной части Байкальской рифтовой зоны, в среднем течении р. Витим, на высоте 353 м над у. м. (рис. 1). Его территория находится на стыке Станового и Байкало-Патомского нагорий, входит в состав Саяно-Байкальской горной области (расположена в пределах хребта Кодар) [3]. Озеро имеет тектоническое происхождение. Его длина 24 км, ширина 2.5–6.0 км, максимальная глубина 184 м. Донные отложения в основном представлены илами. Песчаные грунты, а также грунты, сформированные растительным детритом, распространены в северной и южной частях озера [1]. Главные притоки озера – реки Сыгыкта, Култушная и Каменная, берущие начало в Кодарском хребте. Кроме этих рек в озеро впадает >10 небольших речек и ключей.
Рис. 1.
Географическое положение оз. Орон (слева) и схема грунтов оз. Орон (справа). 1 – ил, 2 – песок, 3 – заиленный песок, 4 – водная растительность, 5 – заиленный песок с растительностью. ⚫ – место отбора пробы озерных осадков.
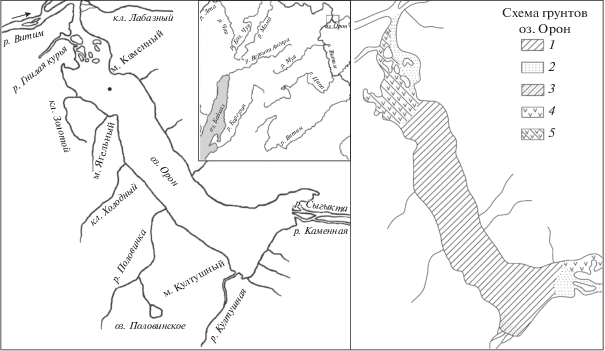
Согласно геоботаническому районированию Иркутской обл. [1], озеро расположено в Делюн-Уранском горно-таежно-гольцовом округе Патомской кедрово-стланниково-лиственнично-горно-таежной провинции. Основная часть территории занята каменистыми россыпями и скалами. Наибольшую площадь занимают лиственничные леса. Еловые, еловые с пихтой леса распространены в долине р. Сыгыкта и ключа Лабазный. В северо-западной оконечности озера, примыкающей к правому берегу р. Витим, отмечены участки леса из сосны обыкновенной.
В марте 2013 г. в оз. Орон нами взята проба донных отложений. Осадок отбирали с глубины 9 м в мелководной зоне в створе мыса Каменный (57°09′14″ с.ш., 116°27′59″ в.д.) с помощью пробоотборника Uwitec-Corer. Длина отобранного керна была 73 см, исследовали верхние 30 см.
Датировку горизонтов проводили по распределению 210Pb и 137Cs. Методы радиоизотопного датирования горизонтов, использованные для отобранного керна, подробно описаны в работе [18]. Подготовку, палинологический анализ полученного материала и изучение погребенных макроостатков личинок хирономид осуществляли стандартными методами [7, 10].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Согласно полученным данным датирования, верхние 10 см керна формировались не ранее 1870 г. Датировки для нижележащих 20 см керна – результат экстраполяции установленных дат.
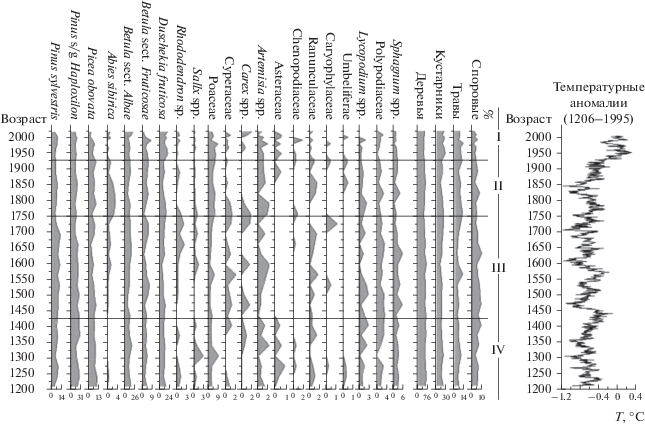
На основе проведенного спорово-пыльцевого анализа отобранного керна построена диаграмма (рис. 2), на которой выделены четыре палинозоны (Пз IV–I), отражающие изменения локальной и региональной растительности за последние 800 лет.
Рис. 2.
Спорово-пыльцевая диаграмма 30 см керна донных отложений оз. Орон и сопоставление результатов исследования с данными по изменению среднегодовых температур в Северном полушарии [12]. IV–I – палинозоны (Пз).
Как по биомассе, так и по таксономическому составу, личинки хирономид – одна из доминирующих групп в сообществах бентосных организмов. Они уже давно используются в лимнологии как индикаторы различных типов местообитаний и палеоклиматических изменений [9, 13, 16, 18 и др.].
В настоящее время в фауне оз. Орон зарегистрировано 43 вида и личиночных форм хирономид, относящихся к 26 родам из пяти подсемейств: Tanypodinae, Diamesinae, Prodiamesinae, Orthocladiinae и Chironominae [6]. Последние два подсемейства представлены наибольшим числом видов. По останкам, содержащимся в донных отложениях оз. Орон, автором выявлено 39 видов и личиночных форм хирономид, относящихся к пяти подсемействам. Среди них также преобладали Orthocladiinae и Chironominae – 19 и 15 таксонов соответственно.
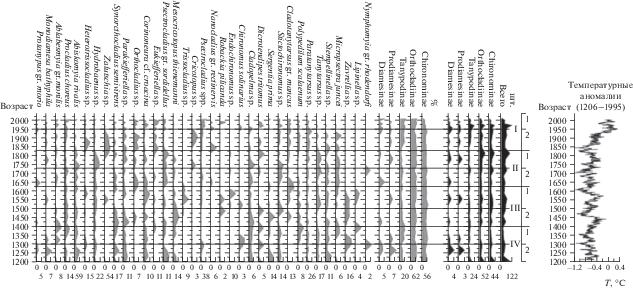
На основе полученных данных построена диаграмма (рис. 3), отражающая изменения таксономического состава личинок хирономид оз. Орон в ответ на изменяющиеся условия окружающей среды; выделено четыре зоны, каждая из которых делится на подзоны, соответствующие “холодным” и “теплым” этапам истории озера.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Прямым применением палинологического метода являются реконструкции растительного покрова. Спорово-пыльцевая диаграмма – это вполне надежный документ, дающий представление о составе растительного покрова определенной территории в определенное время. Рассмотрим диаграмму на рис. 2.
Пз IV (интервал 30–22 см). Во время формирования 30–22 см осадочной толщи оз. Орон имело место постепенное похолодание и возрастание сухости климата. В это время большая часть подгольцового пояса была занята зарослями кедрового стланика; значительна лесообразующая роль сосны обыкновенной у верхней границы леса. В горных долинах заметное развитие получали еловые и елово-березовые леса с подлеском из ольховника. На границе с высокогорными лугами господствующее положение занимали кустарниковые заросли из березы и ивы. За пределами прирусловой части долин на плоских водоразделах господствовали древесные березы, на торфяно-мерзлотных почвах подгорных шлейфов и надпойменных террас были распространены кустарниковые ассоциации из ерника и ивы. На открытых участках формировались полынные и полынно-маревые сообщества степного облика.
Пз III (интервал 22–12 см). Вместе с небольшим повышением температуры условия среды становятся влажнее. В долинах рек и на заболоченных участках широкое распространение получали ассоциации из ерника и ивы. Избыточное увлажнение пойм привело к господству прибрежно-водных (Cyperaceae) и лугово-болотных растений (Ranunculaceae, Caryophylaceae, Carex spp.). Значительная часть подгольцового пояса занята стланиковыми зарослями. Во влажных понижениях рельефа могли произрастать сосново-березовые леса с участием кедра (сосны кедровой) и подлеском из ольховника, а в каньонообразных горных долинах – еловые и елово-пихтовые леса с участием древесных берез. Наиболее прогреваемые скально-каменистые и щебнистые горные склоны окружающих водораздельных хребтов были заняты сосновыми лесами. Здесь же могли формироваться сухие полынные сообщества с незначительным участием злаков. Широкое распространение на затененных склонах получили сосняки с подлеском из рододендрона.
Пз II (интервал 12–7 см). Последующее повышение среднегодовой температуры и влажности способствовало возрастанию роли пихты в сложении елово-пихтовых лесов с незначительным участием древесных берез и ерника в горных долинах. Заметно снизилась лесообразующая роль сосны обыкновенной и кедрового стланика у верхней границы леса; сократились площади сосняков рододендроновых. Вероятно, в это время частые лесные пожары оказывали большое влияние на распределение растительности по склонам гор, окружающих озеро. В нижнем и верхнем тундрово-альпийском поясе гор широкое распространение получили сообщества степного облика. В сложении растительности пойм заметно уменьшилось участие влаголюбивых травянистых, возрастала роль злаков. Складывались оптимальные по температуре и влажности условия, увеличивалась роль атмосферных осадков в летние месяцы.
Пз I (интервал 7–0 см). Дальнейшее потепление и увеличение сухости привело к заметным изменениям в составе растительности. Значительно снизилась роль пихты и ели. В группе травянистых растений увеличилась роль степных элементов (Asteraceae, Chenopodiaceae). На осушенных пойменных лугах Cyperaceae, Ranunculaceae, Caryophylaceae уступили место полынным группировкам с участием злаков (Poaceae), сложноцветных (Asteraceae) и маревых (Chenopodiaceae).
В систематическом составе фауны ископаемых хирономид оз. Орон прослеживается значительное сходство с биотой озер, расположенных в тектонических впадинах Байкальской рифтовой зоны [4]. Отмечено, что количественное развитие и структурная организация зообентоса оз. Орон характеризуется устойчивым и стабильным состоянием, что в целом характерно для горных озер независимо от их морфометрии и генезиса [6]. Однако если для такой оценки использовать хирономид отдельно от других групп, слагающих макрозообентос оз. Орон, этот вывод не подтверждается.
Насколько стабильна структура хирономид озера в многолетнем плане, можно судить по диаграмме на рис. 3. Из пяти подсемейств лишь для двух (Diamesinae и Prodiamesinae) состояние количественного развития и структурной организации в той или иной мере может характеризоваться как стабильное. Для трех других подсемейств отмечена тенденция постепенного возрастания биомассы (на 12–5 см керна, ~1750–1961 гг. – Chironominae и Orthocladiinae; на 9–5 см, ~1850–1961 гг. – Tanypodinae) и последующего ее снижения (5–0 см). На представленной диаграмме максимуму Diamesinae соответствует минимум Tanypodinae. С уменьшением процентного содержания останков личинок Diamesinae в горизонтах осадка рассматриваемой зоны роль Tanypodinae возрастает. Аналогичным образом пикам Orthocladiinae соответствует спад Chironominae. С возрастанием роли Chironominae участие Orthocladiinae в горизонтах осадка снижается. Отчетливые пики процентного содержания Chironominae с соответствующими им спадами Orthocladiinae зафиксированы на 28, 20, 14 и 7 см керна, т.е. через каждые ~7 см. Таким образом, изученный керн отражает смену фаз, которая в период накопления 30-сантиметровой толщи осадочного чехла происходила в ~200-летнем природном цикле. Смена фаз обусловлена изменениями среднегодовой температуры в Северном полушарии и, вероятно, значительными колебаниями расхода рек и ручьев, впадающих в оз. Орон.
Зона IV (интервал 30–23 см). Горизонты осадка, соответствующие рассматриваемой зоне, отличается очень низким содержанием макроостанков личинок хирономид, в процентном отношении характеризуется значительным (по сравнению с III и II зонами) участием представителей подсем. Tanypodinae, Diamesinae и Prodiamesinae.
Подзона IV2 (интервал 30–26 см). Обращают на себя внимание пики Abiskomyia rivalis Makarchenko E. et Makarchenko M., Thiemanniella sp., Sergentia prima Proviz V. et Proviz L., Stempellinella sp. В то же время роль Parakiefferiella sp. в подзоне постепенно снижается от нижележащих горизонтов осадка к вышележащим. Среди приведенных таксонов хирономид Abiskomyia rivalis, Thiemanniella sp., Sergentia prima, Stempellina sp. – наиболее холодолюбивые. Личинки Abiskomyia встречаются в холодных олиготрофных арктических и субарктических озерах по всей Голарктике [10]. Селятся в замшелых участках русел небольших водотоков [11]. Очевидно, вид Abiskomyia rivalis не обитал в самом озере, а останки его личинок были привнесены в осадки оз. Орон ручьями и реками. Наибольшее количество головных капсул личинок A. rivalis отлагалось в слоях осадка, сформировавшихся в наиболее холодное время, когда ключи, впадающие в озеро, характеризовались малым расходом. Такие условия благоприятствовали формированию в каменистых руслах ручьев мохового покрова, где личинки этого вида жили и развивались в большом количестве. Периодически их макроостатки смывались в озеро течением ручьев.
Также следует отметить находку останков личинок Nymphomyia gr. rhodendorfi [2]. Это высокоспециализированные двукрылые насекомые, по-видимому, являющиеся филогенетическими и географическими реликтами [5]. В настоящее время в мировой фауне насчитывается семь видов нимфомийид, встречающихся на территории Российского Дальнего Востока, в Японии, Канаде, США, Индии, Западной Бенгалии и Гонконге. Личинки населяют предгорные и горные реки с каменистыми гравийно-галечными грунтами, быстрым течением и высоким содержанием кислорода в воде.
Вероятно, во время формирования горизонтов осадка, которым соответствует рассматриваемая подзона, на территории озера и сопредельных территориях складывались “холодные” и “сухие” условия. Лето характеризовалось низкими температурами и малым количеством осадков, реки и ручьи, впадающие в озеро, – низким стоком.
Подзона IV1 (интервал 26–23 см). Заметно возрастает число останков личинок таких видов, как Ablobesmyia gr. monilis, Procladius choreus Mg., Cladopelma sp., Lipinella sp., а также Polipedillum scalaenum Schrank, Parakiefferiella sp. и Micropsectra juncei (Meigen). В то же время уменьшается роль видов Abiskomyia rivalis, Thiemanniella sp., Psectrocladius gr. sordidellus, Sergentia prima. Представители рода Cladopelma sp. относятся к достаточно теплолюбивым [19], стенотермным видам, однако они не особенно терпимы к условиям высокой трофности, характерным для мезотрофных озер [9, 17]. Обитают в литоральной зоне, на илистых и песчаных/каменистых субстратах [15, 20]. Представители Polipedillum служат индикаторами умеренных климатических условий [10]. Обитают в литорали эвтрофных озер [9, 15], некоторые виды встречаются среди водной растительности [8].
Вероятно, во время формирования горизонтов осадочного чехла, соответствующего подзоне IV1, климат на рассматриваемой территории был теплее. С активным таянием снежников и ледников, вызванным повышением среднегодовой температуры, возрастал водорасход ручьев и рек, впадающих в оз. Орон. Бурные потоки вычищали каменистые русла от моховых обрастаний. С сокращением местообитаний, пригодных для развития личинок Abiskomyia rivalis в руслах рек и ручьев, впадающих в оз. Орон, уменьшался смыв их останков в озеро. В летнее время выпадало достаточное количество осадков, увеличивался сток, наполнялись курьи и протоки. Это давало возможность для развития фауны литоральных видов.
Зона III (интервал 23–16 см). Резко снизилось количество личинок Ablabesmyia gr. monilis. Уменьшается и роль Procladius choreus. Однако этот вид, хотя и в небольшом количестве, все же отмечается во всех горизонтах рассматриваемой зоны. Аналогичная картина наблюдается у Abiskomyia rivalis, Cladopelma sp., Polypedilum scalaenum. Встречаемость других видов (Hydrobaenus sp., Paratanytarsus sp.) в слоях зоны III осталась такой же, как и в нижележащей зоне. Появляются новые таксоны, не отмечавшиеся в нижележащих горизонтах осадка: Zalutschia sp., Trissocladius sp., Robackia pilicauda Saether, Endochironomus sp.
Подзона III2 (интервал 23–20 см) соответствует “холодному” и “сухому” периоду. Условия осадконакопления складывались по тому же принципу, как и для подзоны IV2. Однако рассматриваемый период отличался более суровыми условиями. В пользу этого говорят низкие показатели биомассы хирономид в горизонтах осадка, а также значительное участие в составе фауны погребенных личинок хирономид холодолюбивых Mesocricotopus thienemanni Goetgh, Sergentia prima, Stictochironomus sp. Отметим, что Mesocricotopus thienemanni – один из наиболее холодолюбивых, стенотермных видов; останки его личинок в донных осадках служат надежным показателем холодных условий прошлого [10].
Подзона III1 (интервал 20–16 см). Количество Procladius choreus, Abiskomyia rivalis, Cladopelma sp. в слоях осадка осталось тем же, что и в подзоне III2. Резко снизилось число Mesocricotopus thienemanni, Sergentia prima и Stictochironomus sp., появились литоральные Zalutschia sp., Endochironomus sp., Cladotanytarsus gr. mancus, Lipinella sp. Представители рода Endochironomus предпочитают эвтрофные водоемы, обычны в растительном детрите на глубине 1-2 м, либо являются минерами макрофитов. Среди видов рода Zalutschia (Z. zalutschicola Lipina) есть виды, обитающие на грубом детрите эвтрофных и дистрофных озер. Таким образом, таксономический состав хирономид данной подзоны свидетельствует о существовании “теплых” и “влажных” условий на территории озера в период формирования 16–20 см озерных осадков. Картина событий этого времени аналогична таковой при анализе подзоны IV1.
Зона II (интервал 16–10 см). Заметно возрастает количество макроостатков личинок хирономид в горизонтах осадка. Наблюдается резкое увеличение численности Hydrobaenus sp., Zalutschia sp., Tanytarsus spp. и снижение Cladotanytarsus gr. mancus и Endochironomus sp. Единично отмечены Nymphomyia gr. rhodendorfi, Mesosmittia sp., Cricotopus sp., Nanocladius gr. rectinervis, исчезает Sergentia prima. Слои осадка рассматриваемой зоны формировались в более теплых условиях, чем таковые, залегающие ниже. И все же в зоне еще можно выделить подзоны, соответствующие более “холодному” и более “теплому” периодам времени, в которое формировались данные горизонты осадка.
Подзона II2 (интервал 16–13 см). Возрастает роль Procladius choreus, Ablobesmyia gr. monilis. Резко увеличивается количество Abiskomyia rivalis, Hydrobaenus sp., Stictochironomus sp.; падает численность Cladopelma sp. Таксономический состав и количественное соотношение отдельных групп хирономид свидетельствуют о “холодных” климатических условиях.
Подзона II1 (интервал 13–10 см). В горизонтах осадка резко снижается количество макроостатков Tanypodinae (Procladius choreus, Ablobesmyia gr. monilis), Abiskomyia rivalis, Hydrobaenus sp., Stictochironomus sp., а также Stempellinella sp. и Micropsectra juncei. В то же время возрастает роль Zalutschia sp., Parakiefferiella sp., Psectrocladius gr. sordidellus. Условия формирования горизонтов осадочного чехла, соответствующих рассматриваемой подзоне, можно охарактеризовать как “теплые” и “сухие”.
Зона I (интервал 10–0 см). Накопление осадков приходится на конец Малого Ледникового периода, время последнего глобального потепления (Recent Warming, 1850–1860) и длительного периода регионального потепления климата с 1900 по 1960 гг. [17]. Зона отражает постепенное становление современных (или близких к ним) условий среды.
Выводы. Проведена реконструкция изменения состава хирономидофауны оз. Орон в ответ на изменяющиеся условия окружающей среды за ~800-летний период. Рассмотрена возможная картина преобразования ландшафтов на прилегающих к озеру территориях. Установлено, что значительное влияние на изменение таксономического состава хирономид в макрозообентосе оз. Орон оказывало колебание водорасхода рек и ручьев, впадающих в озеро. Во время похолодания климата сток в озеро уменьшался за счет снижения активности таяния ледников, снежников и сезонного снежного покрова Кодарского хребта – деградация снежников не превышала их прироста. Это позволило охарактеризовать такие периоды истории озера как “холодные” и “сухие”. В результате глобального увеличения температур в Северном полушарии ледники и снежники постепенно сокращали свои площади – сток в озеро возрастал. Таким образом, вместе с потеплением климата наступал “влажный” период, когда наряду с растительными сообществами степного облика на наиболее прогреваемых склонах широкое распространение в понижениях рельефа получали мезофитные сообщества.
Изученный керн отражает смену “теплых” и “холодных” фаз в истории озера. Каждая смена в рассматриваемый период времени происходила в 200-летнем цикле. В настоящее время в истории оз. Орон продолжается вторая, “теплая” фаза цикла, которая началась ~70 лет назад.
БЛАГОДАРНОСТИ
Автор выражает благодарность д.г.-м.н. А.П. Федотову (Лимнологический институт СО РАН, Иркутск), за организацию экспедиции на оз. Орон в 2013 г. и участие в ней; друзьям и коллегам из Государственного природного заповедника “Витимский” (Бодайбо): В.А. Исаеву, ${\text{С }}{\text{.В }}{\text{. Н и к и ш и н у }}$, ${\text{М }}{\text{.А }}{\text{. Р о г о в у }}$, О.А. Замулину, П.И. Старожуку, оказавших содействие во время совместной работы на территории заповедника; друзьям и коллегам д.г.н., проф. В.Б. Выркину (Институт географии им. В.Б. Сочавы СО РАН, Иркутск) и д.б.н. В.В. Тахтееву (ФГБОУ ВО “Иркутский государственный университет, биолого-почвенный факультет”) за критический просмотр рукописи, сделанные замечания и ценные советы.
Список литературы
Атлас. Иркутская область: экологические условия развития. М.; Иркутск, 2004. 90 с.
Енущенко И.В., Макарченко Е.А. Находка останков личинок нимфомийид (Diptera, Nymphomyiidae) в донных осадках озера Орон (Иркутская область) // Жизнь пресных вод. Владивосток: Дальнаука, 2016. С. 180–184.
Карта геоморфологического районирования СССР. Масштаб 1 : 8 000 000. М.: Главное управление геодезии и картографии, 1985. 2 с.
Линевич А.А. Хирономиды Байкала и Прибайкалья. Новосибирск: Наука. Сиб. отд., 1981. 152 с.
Макарченко Е.А. Сем. Nymphomyiidae – Нимфомииды // Определитель насекомых Дальнего Востока России. Т. VI: Двукрылые и блохи. Ч. 4. Владивосток: Дальнаука, 2006. С. 734–739.
Матвеев А.Н., Самусенок В.П., Рожкова Н.А. и др. Биота Витимского заповедника: структура биоты водных экосистем. Новосибирск: Академическое изд-во “Гео”, 2006. 267 с.
Berglund B.E., Ralska-Jasiewiczowa M. Pollen analysis and pollen diagrams // Handbook of Holocene Palaeoecology and Palaeochydrology. N.Y.: John Wiley & Sons, 1986. P. 455–484.
Brodersen K.P., Odgaard B.V., Vestergaard O., Anderson N.J. Chironomid stratigraphyin the shallow and eutrophic Lake Søbygaard, Denmark: chironomid-macrophyte co-occurrence // Freshwater Biol. 2001. № 46. P. 253–267.
Brodin Y.W. The postglacial history of Lake Flarken, southern Sweden, interpreted from subfossil insect remains // Int. Rev. Gesamt Hydrobiol. 1986. № 71. P. 371–432.
Brooks S.J., Langdon P.G., Heiri O. The identification and use of Palaearctic Chironomidae larvae in palaeoecology. Technical Guide No. 10. L.: Qurternary research association. 2007. 276 p.
Cranston P.S., Oliver D.R., Saeter O.A. The adult males of Orthocladiinae (Diptera: Chironomidae) of the Holarctic region – Keys and diagnoses // Entomologica Scandinavica. Suppl. 34. 1989. P. 165–352.
D’Arrigo R., Wilson R., Jacoby G. On the long-term context for late twentieth century warming // J. Geophys. Res. 2006. https://doi.org/10.1029/2005JD006352
Enushchenko I.V., Melgunov M.S., Fedotov A.P. Reconstruction of summer temperatures in East Siberia (Russia) for the last 850 years, inferred from records in lake sediments of non-biting midges (Diptera: Chironomidae) // Int. J. Environ. Stud. 2014. V. 71. № 5. P. 647–655.
Fedotov A.P., Trunova V.A., Zvereva V.V. et al. Reconstruction of glacier fluctuation (East Siberia, Russia) during the last 160 years based on high-resolution geochemical proxies from proglacial lake bottom sediments of the Baikalsky Ridge // Int. J. Environ. Stud. 2012. V. 69. № 5. P. 806–815.
Hofman W. Stratigraphie subfossiler Cladocera (Crustaceae) und Chironomidae (Diptera) in zwei Sedimentprofilen des Meerfelder Maares // Courier Forschungs Institut Senckenberg. 1984. V. 65. P. 67–80.
Jones P.D., Briffa K.R., Osborn T.J. et al. High-resolution palaeoclimatology of the last millennium: a review of current status and future prospects // Holocene. 2009. V. 19. P. 3–49.
Saether O.A. Chironomid communities as water quality indicators // Holarctic Ecol. 1979. V. 2. P. 65–74.
Vorobyeva S.S., Trunova V.A., Stepanova O.G. et al. Impact of glacier changes on ecosystem of proglacial lakes in high mountain regions of East Siberia (Russia) // Environ. Earth Sci. 2015. № 74. P. 2055–2063.
Walker I.R., Smol J.P., Engstrom D.R., Briks H.J.B. An assessment of Chironomidae as quantitative indicators of past climatic change // Can. J. Fish and Aquat. Sci. 1991. № 48. P. 975–987.
Walker I.R., MacDonald G.M. Distributions of Chironomidae (Insecta: Diptera) and other freshwater midges with respect to tree line, Northwest Territories, Canada // Arctic and Alpine Res. 1995. № 3. P. 258–263.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод