

Биология внутренних вод, 2019, № 4-2, стр. 36-43
Семенная продуктивность и особенности плавучести генеративных диаспор некоторых европейских видов рода Sparganium L.
Е. А. Беляков a, b, *, А. Г. Лапиров a, **
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Некоузский р-н, Ярославская обл., пос. Борок, Россия
b Череповецкий государственный университет
Череповец, Россия
* E-mail: Eugenybeliakov@yandex.ru
** E-mail: lapir@ibiw.yaroslavl.ru
Поступила в редакцию 12.03.2019
После доработки 09.04.2019
Принята к публикации 25.05.2019
Аннотация
Изучены фактическая семенная продуктивность и плавательная способность генеративных диаспор различных видов Sparganium L. Установлено, что по мере снижения фактической семенной продуктивности виды Sparganium формируют следующий ряд: S. microcarpum (Neum.) Domin. > S. emersum Rehm. > S. glomeratum (Laest.) Neum. > S. gramineum Georgi. > S. erectum L. > S. hyperboreum Laest. ex Beurl. > S. natans L. Минимальными показателями фактической семенной продуктивности характеризуются представители секции Minima Aschers. et. Graebner. – S. hyperboreum и S. natans, что обусловлено малыми размерами этих видов. Наибольшей плавательной способностью обладают плодики видов подрода Sparganium (по сравнению с представителями подрода Xanthosparganium Holmb.), имеющие более толстую стенку костянки и большее число воздухоносных полостей в околоплоднике. В отличие от остальных видов плодики S. gramineum после отделения от растения погружаются на дно водоема.
ВВЕДЕНИЕ
Семена – важнейшее, а часто и единственное средство размножения и расселения растений (Николаева и др., 1999). При этом, эффект размножения достигается лишь в том случае, если оно приводит к возобновлению вида в ценозе (Левина, 1987). Однако даже самое интенсивное продуцирование семян создает лишь предпосылки для возобновления вида (Левина, 1981, 1987; Middleton et al., 2006; Nathan, Muller-Landau, 2000). Изучение семенной продуктивности и процессов диссеминации (рассеивания семян) различных видов растений на значительные расстояния важно как с теоретической, так и с практической точки зрения (Вайнагий, 1974). Эти сведения полезны не только для понимания адаптационных механизмов приспособления растений к окружающей среде, но и для осознания их роли в фитоценозе (Вайнагий, 1974; Ходачек, 1970).
Известно, что на основе потенциальной и реальной семенной продуктивности можно судить не только о степени соответствия экологических условий современных местообитаний биологическим требованиям вида, но и о жизненном состоянии популяций. Репродуктивные показатели многих видов связаны не только с генетическим потенциалом конкретного вида (Баранова и др., 2012). Они зависят от многих абиотических и биотических факторов (способа и условий опыления, наличия фитофагов, конкретной климатической зоны и др.) (Вайнагий, 1974; Зубаирова, 2013; Левина, 1981; Schopmeyer, 1974). Данные о семенной продуктивности различных видов водных и прибрежно-водных растений в литературе немногочисленны (Беляков и др., 2017; Девятов, 2014; Лапиров и др., 2017; Лебедева, Лапиров, 2013; Чернова, 2013).
С учетом роста антропогенной нагрузки на окружающую среду (Vittoz, Engler, 2007), актуальность работ, связанных с механизмами распространения генеративных и вегетативных диаспор, становится все более очевидным фактом (Девятов, 2014; Chambert, 2006; Couvreur et al., 2005; Müller-Schneider, 1983; Ridley, 1930; Vittoz, Engler, 2007). Для аборигенных видов растений это связано с изменением естественных природных ландшафтов, а также с проблемами, создаваемыми растениями-вселенцами (Couvreur et al., 2005). Сюда можно отнести и активную миграцию растений в ответ на глобальные изменения климатических условий (особенно при прогнозируемом глобальном потеплении) (Vittoz, Engler, 2007). Таким образом, изучение особенностей распространения диаспор растений решает ряд важных задач по сохранению биоразнообразия, пониманию процессов эволюции, объяснению текущего распределения растения в разных ландшафтах и прогнозированию будущих изменений (Sádlo et al., 2018).
Основным агентом по распространению водных и прибрежно-водных видов растений, в том числе различных видов рода Sparganium L., служит вода (гидрохория). Именно поэтому большое влияние на особенность распространения таких растений оказывает плавательная способность генеративных диаспор. Однако, в связи с рядом трудностей, возникающих в процессе исследования, данные по плавучести семян и плодов водных и прибрежно-водных растений в литературе по-прежнему сильно разрозненны и немногочисленны (Andersson et al., 2000; Cook, 1990; Griffith, Forseth, 2002; Hroudová et al., 1997; Pollux et al., 2009; Ridley, 1930; Sádlo et al., 2018; Soomers et al., 2010; Staniforth, Cavers, 1976).
Представители рода Sparganium L. относятся к экологической группе гелофитов (за исключением типичного гидрофита S. gramineum Georgi) и считаются одним из основных компонентов водных экосистем. Выбор этих растений в качестве объекта исследования обусловлен не только особенностями их плодов, но и наличием как массовых (обычных для исследуемой территории), так и редких (реликтовых) видов, например, S. gramineum. Последний представлен в Красных книгах ряда регионов России (Belyakov, Lapirov, 2018). Интересен в этой связи и S. natans L. – в ряде регионов Европейской России его указывают как редкий, уязвимый или нуждающийся в биологическом контроле вид (Belyakov, Philippov, 2018).
Цель работы – сравнительное изучение семенной продуктивности некоторых европейских видов рода Sparganium L. и плавательной способности их генеративных диаспор.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Семенная продуктивность. Материал для исследования ФСП Sparganium собирали в ходе маршрутных исследований на водоемах и водотоках Ярославской, Московской, Вологодской, Тверской, Калужской и Нижегородской областей и Республики Удмуртия. Исследование ФСП осуществляли в соответствие с методикой Вайнагия (1974). Под ФСП мы, вслед за данным автором, понимаем общее число завязавшихся семян на МП. Для подсчета числа семян из каждой исследованной популяции в случайном порядке отбирали по пять МП. Затем в лабораторных условиях подсчитывали число реализованных плодиков в составе каждого соплодия. Всего исследовано 36 МП у S. emersum Rehm., 12 у S. glomeratum (Laest.) Neum., 38 у S. gramineum, 74 у S. natans, 56 у S. erectum L. и 44 у S. microcarpum (Neum.) Domin. Также семенную продуктивность изучали у S. hyperboreum Laest. ex Beurl. (15 МП). Плодики этого растения собирали в мелководном водоeме, расположенном вдоль бетонной дороги в тундре, с. Газ-Сала (67°20′31″ с.ш., 78°56′48″ в.д.; Тазовский р-н Тюменской обл.; сбор 05.VIII 2016 г.).
Плавательная способность плодиков. Для проведения экспериментов генеративныe диаспоры S. emersum собирали на правом берегу р. Корожечна напротив с. Масальское (57°32′28.05″ c.ш., 38°2′13.87″ в.д., Угличский р-н Ярославской обл., сбор 23.VIII 2013 г.); плодики S. natans – в мелиоративной канаве на южном берегу оз. Чашницкое (56°56′12.60″ с.ш., 39°22′32.00″ в.д., Ростовский р-н Ярославской обл., сбор 12.X 2016 г.); плодики S. gramineum – на оз. Рюмниково (56°58′12″ с.ш., 39°23′20″ в.д., Ростовский р-н Ярославской обл., сбор 13.IX 2015 г.), плодики S. erectum – на обсыхающем участке литорали Рыбинского водохранилища в месте впадения р. Сутка и Ильд (58°1′19.11″ с.ш., 38°15′58.51″ в.д., Некоузский р-н, Ярославской обл., сбор 10.IX 2013 г.); S. microcarpum – на р. Урдома (57°56′28.7″ с.ш., 39°29′39.4 в.д., Тутаeвский р-н Ярославской обл., сбор 13.IX 2015 г.).
Для изучения плавательной способности свежесобранные плодики видов Sparganium (по 100 шт.) помещали в индивидуальные стеклянные этикетированные стаканы с отстоявшейся водопроводной водой (в трeх повторностях). В течение первого месяца опустившиеся на дно сосудов плодики учитывали ежедневно, затем – через 2 сут, спустя 8 мес – раз в неделю. Эксперимент продолжался 1.5 года.
После этого плодики помещали в холодильник на влажную холодную стратификацию и содержали в течение 1.5 мес при температуре 3 ± 2°С. Затем плодики переносили в люминостат на 25 сут (освещенность 1868 ± 126 лк, фотопериод 16/8, температура от 18.2 ± 0.9°С (утро) до 28.7 ± 2.9°С (вечером)), где они проращивались в чашках Петри на фильтровальной бумаге, смоченной отстоявшейся водопроводной водой. Все варианты опыта проводили в трех повторностях (по 50 плодиков в каждой). Один раз в сутки в одно и то же время отмечали число проросших плодиков. Для характеристики процесса прорастания применяли показатель Gfin (Shipley, Parent, 1991), соответствующий термину “лабораторная всхожесть” (Николаева и др., 1999; Попцов, Некрасов и др. 1981).
По окончании эксперимента непроросшие плодики каждого вида Sparganium проверяли на жизнеспособность с использованием тетразольного метода (Николаева и др., 1999).
В тексте и таблицах все данные представлены в виде средних значений и их стандартных отклонений.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Семенная продуктивность. Среднее число соплодий на МП у исследованных видов Sparganium различается. У S. microcarpum и S. erectum (подрод Sparganium) на одном МП формируются от 5.4 ± 2.1 до 7.8 ± 1.8 соплодий, соответственно. У представителей подрода Xanthosparganium Holmb. число соплодий на одном МП тоже варьирует: у S. emersum образуется 3.2 ± 0.6 соплодий, у S. gramineum – 4.7 ± 1.5, у S. glomeratum – 4.8 ± 0.5, у S. hyperboreum Laest. ex Beurl. – 2.7 ± 0.7, у S. natans – 2.0 ± 0.4. Средние значения числа плодиков в соплодиях даны в табл. 1.
Таблица 1.
Среднeе число (шт.) и размерно-массовая характеристика плодиков в одном соплодии некоторых представителей рода Sparganium
| Вид | Плодики | Масса, г | Lобщ, мм | Lст, мм | d, мм | ||
|---|---|---|---|---|---|---|---|
| развитые | недоразвитые | всего | |||||
| Подрод Xanthosparganium | |||||||
| Sparganium emersum | 173 ± 69 | 28 ± 19 | 182 ± 47 | 0.556 ± 0.008 | 11.2 ± 1.0 | 3.4 ± 0.6 | 2.0 ± 0.2 |
| S. glomeratum | 36 ± 31 | 98 ± 28 | 134 ± 30 | 0.260 ± 0.003 | 4.8 ± 0.3 | 1.3 ± 0.2 | 1.7 ± 0.2 |
| S. gramineum | 54 ± 30 | 54 ± 31 | 100 ± 36 | 0.351 ± 0.031 | 4.4 ± 1.0 | 1.8 ± 0.2 | 2.0 ± 0.3 |
| S. natans | 34 ± 12 | 10 ± 7 | 44 ± 11 | 0.248 ± 0.030 | 4.1 ± 0.6 | 1.0 ± 0.2 | 2.0 ± 0.3 |
| S. hyperboreum | 50 ± 14 | 29 ± 19 | 79 ± 17 | 0.140 ± 0.002 | 3.4 ± 0.2 | 0.3 ± 0.1 | 1.4 ± 0.1 |
| Подрод Sparganium | |||||||
| S. erectum | 63 ± 20 | 26 ± 14 | 79 ± 22 | 2.540 ± 0.152 | 8.1 ± 1.6 | 3.7 ± 1.0 | 3.7 ± 0.7 |
| S. microcarpum | 93 ± 22 | 29 ± 12 | 115 ± 27 | 0.922 ± 0.022 | 9.7 ± 0.3 | 3.8 ± 0.3 | 2.0 ± 0.0 |
МП у S. erectum нередко несет больше соплодий, чем у S. microcarpum, однако, общее число плодиков в соплодиях первого вида в 1.4–1.5 раз меньше из-за их большого диаметра (табл. 1). Также плодики S. erectum отличаются от плодиков остальных видов значительно бóльшей массой.
Высокая ФСП зарегистрирована на одном МП S. emersum и S. microcarpum (табл. 2). У S. erectum ФСП почти в 1.2 раза ниже по сравнению с S. microcarpum. У остальных представителей подрода Xanthosparganium по мере снижения ФСП образуется следующий ряд: S. glomeratum > S. gramineum > > S. hyperboreum > S. natans. Соотношение числа нормально развитых плодиков на МП к числу недоразвитых плодиков варьирует. У S. erectum, S. microcarpum и S. glomeratum оно находится в пределах 2.4–2.7, у S. hyperboreum составляет 1.6, у S. natans – 2.5. Самое большое значение этого показателя на МП зарегистрировано у S. emersum – 6.1. Отметим, что у S. gramineum число недоразвитых плодиков на МП нередко превышает число нормально развитых (табл. 2).
Таблица 2.
Средние значения фактической семенной продуктивности (ФСП) на монокарпическом побеге представителей рода Sparganium
| Вид | ФСП, шт. | Число плодиков, шт. | |
|---|---|---|---|
| нормально развитых | недоразвитых | ||
| Подрод Xanthosparganium | |||
| Sparganium emersum | 704 ± 290 | 681 ± 427 | 112 ± 67 |
| S. glomeratum | 647 ± 136 | 473 ± 123 | 174 ± 80 |
| S. gramineum | 596 ± 128 | 263 ± 114 | 306 ± 152 |
| S. natans | 73 ± 31 | 56 ± 28 | 16 ± 12 |
| S. hyperboreum | 223 ± 80 | 140 ± 45 | 83 ± 46 |
| Подрод Sparganium | |||
| S. erectum | 568 ± 239 | 548 ± 189 | 224 ± 137 |
| S. microcarpum | 721 ± 268 | 575 ± 238 | 210 ± 103 |
По приблизительным оценкам, у исследованных видов Sparganium ПСП достигает больших величин, по сравнению с ФСП. У S. emersum величина ПСП может быть 732–1360 семязачатков на МП, у S. glomeratum – 716–890, у S. gramineum – 595–813, у S. hyperboreum – 281–385, у S. natans – 114–162, у S. microcarpum – ~1000, у S. erectum – ≥1000.
Плавательная способность генеративных диаспор. Нами установлено, что в лабораторных условиях плодики разных видов Sparganium могут плавать на поверхности воды (в стаканах со стоячей водой) >1.5 лет. Почти у всех исследованных видов погружение на дно части плодиков происходит в первые 2.0–2.5 мес, исключая S. emersum, у которого этот процесс длится 8–9 мес (рис. 1). Затем для каждого из изученных видов процесс стабилизируется до конца эксперимента на разном уровне. Самое малое количество погрузившихся на дно плодиков за весь срок экспозиции (15 мес) отмечено у S. natans (3.0–5.0%). У S. emersum количество таких плодиков было в среднем (5.6–9.3%), у S. erectum – 6.2%, у S. microcarpum – 6.0–7.0%. В отличие от всех исследованных видов Sparganium, плодики S. gramineum почти сразу погружались на дно лабораторных стаканов.
Рис. 1.
Количество плодиков (%) S. emersum (1), S. natans (2), S. microcarpum (3) и S. erectum (4), плавающих на поверхности воды в течение эксперимента.
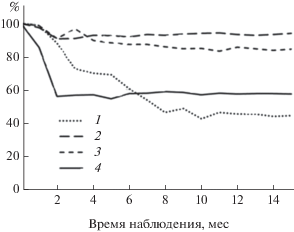
В эксперименте появление проростков в чашках с плодиками, погрузившимися на дно, наблюдали лишь у S. emersum и S. microcarpum. У S. emersum из количества плавающих на поверхности воды плодиков проросло 1.5%, из числа плодиков погрузившихся на дно – 5.1%, у S. microcarpum – 8.8% и 18.0%, соответственно. Отметим, что плодики S. erectum, S. natans и S. gramineum в лабораторных условиях нередко не прорастают, либо прорастают лишь единично.
По окончании эксперимента по прорастанию, проверка на жизнеспособность эмбрионов непроросших плодиков (из групп плавающих и погрузившихся на дно) показала, что у S. erectum, S. emersum и S. natans их количество с отмершим зародышем было ≤2.0–3.0% общей выборки. У плодиков S. microcarpum, плавающих на поверхности воды, почти все зародыши из общей выборки оказались полностью жизнеспособны. Соотношение плодиков с живым и отмершим зародышем в выборке погруженных на дно плодиков этого вида достигало 84.6 и 15.4% соответственно.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате наших исследований показано, что наиболее важные факторы, влияющие на ФСП у Sparganium – число соплодий на побеге и размерные характеристики плодиков. Число реализованных на МП соплодий у Sparganium зависит от конкретных экологических условий, сложившихся в момент цветения (наличия дождливой погоды, подъема уровня воды во время цветения, наличия малого числа насекомых опылителей на затененных участках местообитаний и др.). Между тем, различия в количестве реализованных соплодий, в зависимости от конкретной экологической формы растения, часто оказываются статистически недостоверными.
Размерные характеристики растения также важны. Например, S. hyperboreum и S. natans имеют меньшие размеры, по сравнению с другими видами Sparganium, и, соответственно, они продуцируют меньшее число плодиков, независимо от числа реализованных соплодий. Миниатюризация размеров этих растений, как одна из наиболее важных эволюционных черт у этих видов, могла быть связана с возрастанием экстремальности климатических условий (Хохряков, 1975) в течение прошлых геологических эпох. Заметим, что среднее значение ФСП монокарпического побега S. hyperboreum нередко в ~3.3 раза выше, чем у S. natans.
Сравнивая данные по семенной продуктивности различных видов Sparganium и других видов водных и прибрежно-водных растений, следует отметить, что у последних этот показатель тоже значительно варьирует. Так, семенная продуктивность гидрофита Lobelia dortmanna L. в озерных экосистемах Верхнего Поволжья достигает в среднем 1621 ± 451 семян на один МП (Лапиров и др., 2017), у гидрофитов Ranunculus trichophyllus Chaix, R. circinatus Sibth. и R. kauffmannii Clerc – 300–500 плодиков на МП (Лебедева, Лапиров, 2013). В зависимости от вида у Potamogeton данный показатель также широко варьирует. Например, на одном МП P. berchtoldii Fieber (на территории Ярославской обл.) образуется в среднем 21 ± 16 орешков, у P. alpinus Balb. – 177 ± 38, в то время как у P. perfoliatus L. – 235 ± 108 (данные авторов). Отметим, что при успешном опылении всех цветков семенная продуктивность рдестов была в 2–3 раза больше. У Nuphar lutea L. в малых реках европейской России показатель ФСП лежит в пределах от 106 ± 48 до 363 ± 98 семян на одну ягодообразную многолистовку (Чернова, 2013). У гелофитов (к ним относятся и Sparganium) семенная продуктивность также высока, о чем свидетельствуют представленные выше данные по разным видам Sparganium. По семенной продуктивности им не уступает и гелофит Sagittaria sagittifolia L. (2926 ± 1463 семян), на розеточном побеге которого в течение вегетационного сезона формируется 1–3 цветоносa (данные авторов). По нашим данным у гигрогелофита Scheuchzeria palustris L. ФСП на один МП достигает 16 ± 4 семян, у Lysimachia thyrsiflora L. – 472 ± 284. У Сalla palustris L. количество семян в одном соплодии достигает 164 ± 89 (Беляков и др., 2017). Таким образом, большинство видов водных и прибрежно-водных растений обладают способностью к формированию значительного числа генеративных диаспор.
Определение ПСП, в отличие от ФСП, у Sparganium – задача более сложная. Это связано с наличием у всех изученных нами видов ежеголовников нереализованных головчатых пестичных соцветий в пазухах второго и третьего, реже первого листа цветоносной оси (снизу–вверх). Кроме того, присутствие в некоторых плодиках ежеголовников двухгнездной завязи также усложняет, либо вообще не позволяет, корректно определить ПСП.
Говоря о важности гидрохории, отметим, что у ежеголовников специализация к этому способу распространения, прежде всего, связана с анатомической структурой плодиков. Так, мезокарп Sparganium представлен рыхло расположенными паренхимными клетками и большим количеством воздухоносных полостей (особенно в верхней части плодика) (Зубкова, Шабес, 1983). Твердый эндокарп ежеголовников состоит из мощно развитой механической ткани (Зубкова, Шабес, 1983), надежно защищающей от проникновения воды к эмбриону и в то же время увеличивающей массу плодиков. Кроме того, наружные оболочки плодиков Sparganium плохо смачиваются водой, что, очевидно, тоже отражается на их плавательной способности.
Рядом ученых (Hroudová et al., 1997; Staniforth, Cavers, 1976) отмечена важность сохранения листочков околоцветника, которые на первых этапах способствуют увеличению плавающей способности семян. Аналогичные структуры имеются и при основании плодиков Sparganium. Увеличение продолжительности плавания таких семян и плодов можно объяснить не только ростом площади соприкосновения генеративных диаспор с водой, но и наличием воздушных пространств между плодом и высохшими листочками околоцветника (Staniforth, Cavers, 1976). Подобная особенность может рассматриваться как эволюционная адаптация вида к условиям среды обитания, позволяющая его генеративным диаспорам распространяться на расстояние от нескольких метров, до >100 км (Andersson et al., 2000; Griffith, Forseth, 2002).
Способность держаться на поверхности воды – важный момент для успешного распространения диаспор. Генеративные диаспоры Sparganium, а также ряда других низкотравных гелофитов (Alisma и Sagittaria) способны плавать от 2–10 сут (Чорна, 2006) до 6–15 мес (Ridley, 1930). Все это согласуется с данными, полученными для разных видов ежеголовников. Кроме того, установлено, что семена одного и того же вида растения могут различаться по продолжительности периода плавания на поверхности воды. Эта особенность, названная “бимодальной плавучестью” (Pollux et al., 2009), может быть связана с размерами и массой самого эмбриона, количеством эндосперма и общим объемом воздушных внутренних полостей в экзокарпе. Так, у S. emersum одна часть плодиков (~72% всех плодиков) опускалась на дно в течение 4 нед. (коротко-плавающие), другая (~28%) – сохраняла плавучесть в течение 6 мес. (длительно-плавающие) (Pollux et al., 2009). Плодики, с промежуточными сроками погружения в эксперименте были единичны. Подобное “поведение” плодиков различных видов Sparganium отмечено и авторами. Кроме того, в течение всего эксперимента часть диаспор периодически то погружалась на дно, то всплывала на поверхность воды. Заметим, что плодики S. gramineum, в отличие от таковых других исследованных видов ежеголовников, после отделения от растения не оставались на поверхности воды, а сразу погружались на дно. По нашему мнению, это связано с тем, что в природных условиях соплодия S. gramineum в момент созревания нередко уже полностью или частично оказываются погруженными в воду.
По мнению Поллукса с соавт. (Pollux et al., 2009), коротко-плавающие плодики опускаются на дно в пределах собственных популяций до разрушения надземной биомассы растений. Длительно-плавающие плодики способны распространяться водными потоками вниз по течению водотоков на значительные расстояния, обеспечивая при этом колонизацию новых местообитаний. В условиях стоячих водоемов (прудов и озер) колонизация происходит, как правило, под воздействием ветра. Мы, в свою очередь, склонны полагать, что коротко-плавающие плодики служат страховкой на случай вероятного катастрофического уничтожения родительских популяций (например, в результате сильной засухи или работ по расчистке русел рек).
Важная особенность, повышающая вероятность распространения семян водой – процесс ингибирования их прорастания. Van den Broek et al. (2005) указывают на возможность коэволюции между способностью семян и плодов к удержанию на поверхности воды и ингибированием их прорастания. Так, Кларк и др. (2001) (цит. по: Van den Broek et al., 2005) обнаружили отрицательную корреляцию между плавучестью и скоростью прорастания диаспор у растений мангровых зарослей. Подобная взаимосвязь была подтверждена в настоящей работе, о чем свидетельствует низкий показатель прорастания плавающих на поверхности воды плодиков S. emersum и S. microcarpum.
Вышеизложенное хорошо согласуется с термином “стратегия…” (“dispersal strategies”) – “…ясно выраженные многократно встречающиеся комбинации способов рассеивания, из которых некоторые обычно преобладают у данных видов, в то время как другие менее обычны, но все еще вносят свой вклад в их рассеивание…” (Sádlo et al., 2018). Sádlo et al., (2018, Appendix 1) отнесли различные виды Sparganium к двум типам “стратегий рассеивания”: VII – тип Sparganium (S. emersum, S. erectum subsp. erectum, S. erectum subsp. microcarpum, S. erectum subsp. neglectum, S. erectum subsp. oocarpum) и VIII – тип Wolffia (S. angustifolium и S. natans). К преобладающим “способам рассеивания” (“dispersal mode”) применительно к указанным выше типам стратегий, относятся автохория и гидрохория (автохория не характерна для типа Wolffia). Среди дополнительных способов рассеивания для указанных типов отмечены эндо- и эпизоохория. Кроме того, по мнению этих авторов, тип Sparganium – аналог ветландного типа Wolffia и относится, главным образом, к видам, продуцирующим не только семена с хорошей плавучестью, но и вегетативные диаспоры, играющие важную роль. В стратегии рассеивания типа Wolffia (характерной для водных растений, распространяющихся с помощью плодов, семян и спор) вегетативное размножение доминирует в большинстве случаев, а генеративное – подавлено, либо отсутствует; все типы диаспор – семена, плоды и фрагменты побегов – обычно плавающие (Sádlo et al., 2018).
Выводы. Наиболее высокими показателями ФСП из исследованных видов Sparganium обладают S. emersum и S. erectum, наиболее низкими – представители секции Minima (S. hyperboreum и S. natans). Последнее обусловлено их меньшими размерными характеристиками и, соответственно, меньшим числом плодов в соплодиях. По снижению показателя ФСП виды Sparganium формируют следующий ряд: S. microcarpum > S. emersum > S. glomeratum > S. gramineum > S. erectum > S. hyperboreum > S. natans.
Сложности в изучении ПСП у Sparganium вызывает наличие у разных видов нереализованных пестичных головчатых соцветий и формирование в некоторых костянках двухгнездной завязи, наиболее часто встречающейся у представителей подрода Sparganium – S. erectum и S. microcarpum.
Исследование плавающей способности плодов разных видов ежеголовников показало, что наибольшей плавучестью обладают представители подрода Sparganium, которая обеспечивается большей толщиной стенки костянки и числом воздухоносных полостей в околоплоднике (по сравнению с исследованным представителем подрода Xanthosparganium).
Полученные нами данные подтверждают мнение Pollux et al., (2009), установивших, что коротко-плавающие плодики имеют не только более высокий процент прорастания, но и чуть более высокую скорость прорастания, по сравнению с длительно-плавающими плодиками. Между тем, длительный период гидрохорного распространения плодиков сильно сказывается на их способности к прорастанию и, в меньшей мере, на сохранности зародыша в живом состоянии. Очевидно, что в данном случае, также подтверждается тезис о возможном ингибировании прорастания плодиков, связанном с их высокой способностью к удержанию на поверхности воды (Van den Broek et al., 2005).
Список литературы
Баранова Т.В., Кузнецов Б.И., Моисеева Е.В., Сливкин А.И. 2012. Семенная продуктивность мачка желтого (Glaucium flavum Crantz.) в центральном черноземье // Вестник Воронежского государственного университета. Сер. Химия. Биология. Фармация. № 2. С. 233.
Беляков Е.А., Лапиров А.Г., Лебедева О.А. 2017. Экология прорастания семян и особенности онтогенеза сплавинообразующего гигрогелофита Сalla palustris (Araceae) в лабораторных условиях // Biosystems Diversity. V. 25. № 4. P. 282. https://doi.org/10.15421/011743
Вайнагий И.В. 1974. О методике изучения семенной продуктивности растений // Ботан. журн. Т. 59. № 6. С. 826.
Девятов А.Г. 2014. Репродуктивная экология семенных растений: Учебное пособие к летней практике по ботанике. Москва: МАКС Пресс.
Зубаирова Ш.М. 2013. Особенности семенной продуктивности Hedysarum daghestanicum Boiss. ex Rupr. в природных популяциях // Fundamental Research. № 6. P. 352.
Зубкова И.Г., Шабес Л.К. 1983. Анатомическое строение околоплодника видов Sparganium (Sparganiaceae) // Ботан. журн. Т. 68. № 3. С. 381.
Лапиров А.Г., Беляков Е.А., Лебедева О.А. 2017. Биоморфология и ритм сезонного развития реликтового вида Lobelia dortmanna в олиготрофных озерах Тверской области // Regulatory Mechanisms Biosyst. V. 8. № 3. P. 349. https://doi.org/10.15421/021754
Лебедева О.А., Лапиров А.Г. 2013. О распространении некоторых видов шелковников (Ranunculaceae) в водоемах и водотоках Ярославского Поволжья // Ярославский педагогический вестник. Т. 3. № 2. С. 55.
Левина Р.Е. 1981. Репродуктивная биология семенных растений (обзор проблемы). Москва: Наука.
Левина Р.Е. 1987. Морфология и экология плодов. Ленинград: Наука.
Николаева М.Г., Лянгузова И.В., Поздова Л.М. 1999. Биология семян. Санкт-Петербург: НИИ химии СПбГУ.
Попцов А.В., Некрасов В.И., Иванова И.А. 1981. Очерки по семеноведению. Москва: Наука.
Ходачек Е.А. 1970. Семенная продуктивность и урожай семян растений в тундрах Западного Таймыра // Ботан. журн. Т. 55. № 7. С. 995.
Хохряков А.П. 1975. Соматическая эволюция однодольных. Москва: Наука.
Чернова А.М. 2013. Сравнительный анализ репродуктивной сферы Nyphar lutea (L.) Smith в изменяющихся условиях среды // Вестник Тверского гос. унив. Сер. Биол. и экол. Т. 31. № 23. С. 123.
Чорна Г.А. 2006. Поширення дiаспор гiдрофiтов i гелофiтiв у зв'язку з проблемами синантропiзацiï // Пром. ботан. № 6. С. 60.
Andersson E., Nilsson C., Johansson M.E. 2000. Plant dispersal in boreal rivers and its relation to the diversity of riparian flora // J. Biogeography. V. 27. № 5. P. 1095. https://doi.org/10.1046/j.1365-2699.2000.00481.x
Belyakov E.A., Lapirov A.G. 2018. Morphological and Ecological Cenotic Features of the Relict Species Sparganium gramineum Georgi (Typhaceae) in Waterbodies of European Russia // Inland Water Biology. V. 11. № 4. P. 417. https://doi.org/10.1134/S199508291804003X
Belyakov E.A., Philippov D.A. 2018. The effect of changes in environmental conditions on the morphology of Sparganium natans L. (Typhaceae) in the taiga zone of European Russia // Ecosystem Transformation. V. 1. № 1. P. 29. https://doi.org/10.23859/estr-180326
Chambert S. 2006. Sorting of Seeds by Hydrochory. Göteborg: Department of Civil and Environmental Engineering, Division of Water Environment Technology, Chalmers University Technology.
Cook C.D.K. 1990. Seed dispersal of Nympoides peltata (SG Gmelin) O. Kuntze (Menyanthaceae) // Aquat. Bot. V. 37. № 4. P. 325. https://doi.org/10.1016/0304-3770(90)90019-H
Couvreur M., Cosyns E., Hermy M., Hoffmann M. 2005. Complementarity of epi- and endozoochory of plant seeds by free ranging donkeys // Ecography. V. 28. № 1. P. 37. https://doi.org/10.1111/j.0906-7590.2005.04159.x
Griffith A.B., Forseth I.N. 2002. Primary and secondary seed dispersal of a rare, tidal wetland annual, Aeschynomene virginica // Wetlands. № 22. P. 696.
Hroudová Z., Moravcova L., Zákravský P. 1997. Effect of anatomical structure on the buoyancy of achenes of two subspecies of Bolboschoenus maritimus // Folia Geobotanica et Phytotaxonomica. V. 32. № 4. P. 377. https://doi.org/10.1007/BF02821943
Middleton B., Diggelen R., Jensen K. 2006. Seed dispersal in fens // Applied Vegetation Science. V. 9. № 2. P. 279.
Müller-Schneider P. 1983. Verbreitungsbiologie (Diasporologie) der Blütenpflanzen. Zürich: Geobot. Inst. ETH.
Nathan R., Muller-Landau H.C. 2000. Spatial patterns of seed dispersal, their determinants and consequences for recruitment // Trends in Ecology and Evolution. V. 15. № 7. P. 278. https://doi.org/10.1016/S0169-5347(00)01874-7
Pollux B.J.A., Verbruggen E., Van Groenendael J.M., Ouborg N.J. 2009. Intraspecific variation of seed floating ability in Sparganium emersum suggests a bimodal dispersal strategy // Aquat. Bot. V. 90. № 2. P. 199. https://doi.org/10.1016/j.aquabot.2008.07.002
Ridley H.N. 1930. The Dispersal of Plants throughout the World. Kent: L. Reeve & Co.
Sádlo J., Chytrý M., Pergl J., Pyšek P. 2018. Plant dispersal strategies: a new classification based on the multiple dispersal modes of individual species // Preslia. V. 90. P. 1. https://doi.org/10.23855/preslia.2018.001
Schopmeyer C.S. 1974. Seeds of woody plants in the United States. Washington: U.S. Department of Agriculture, Agriculture Handbook.
Shipley B., Parent M. 1991. Germination responses of 64 wetland species in relation to seed size, minimum time to reproduction and seedling relative growth rate // Functional Ecology. V. 5. № 1. P. 111.
Soomers H., Winkel D.N., Du Y., Wassen M.J. 2010. The dispersal and deposition of hydrochorous plant seeds in drainage ditches // Freshwater Biol. V. 55. № 10. P. 2032. https://doi.org/10.1111/j.1365-2427.2010.02460.x
Staniforth R.J., Cavers P.B. 1976. An experimental study of water dispersal in Polygonum spp. // Can. J. Bot. V. 54. № 22. P. 2587. https://doi.org/10.1139/b76-278
Van den Broek T., Diggelen R., Bobbink R. 2005. Variation in seed buoyancy of species in wetland ecosystems with different flooding dynamics // J. Veget. Sci. V. 16. № 5. P. 579. https://doi.org/10.1111/j.1654-1103.2005.tb02399.x
Vittoz P., Engler R. 2007. Seed dispersal distances: a typology based on dispersal modes and plant traits // Bot. Helvetica. V. 117. № 2. P. 109. https://doi.org/10.1007/s00035-007-0797-8
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод