

Биология внутренних вод, 2020, № 3, стр. 253-259
Структурная организация Nelumbo nucifera (Nelumbonaceae) на северной границе ареала
Н. П. Савиных a, И. А. Коновалова a, *, М. Н. Шаклеина a, Е. В. Лелекова a
a Вятский государственный университет
Киров, Россия
* E-mail: S-dulcamara@yandex.ru
Поступила в редакцию 18.04.2019
После доработки 23.05.2019
Принята к публикации 22.07.2019
Аннотация
На основе биоморфологического анализа описана структурная организация Nelumbo nucifera Gaertn. Побеговая система представлена полурозеточными поликарпическими вегетативно-генеративными осями; модель побегообразования – моноподиальная полурозеточная. Анатомическое строение стебля, черешка и цветоноса N. nucifera соответствует представителям однодольных растений.
ВВЕДЕНИЕ
Nelumbo nucifera Gaertn. (лотос орехоносный) – представитель сем. Лотосовые (Nelumbonaceae) класса Двудольные. Согласно классификации И.Г. Серебрякова (Серебряков, 1964), он гидрофит, относится к типу плавающих и подводных трав, хорошо переносит временное понижение уровня воды, оказываясь на суше (Папченков, 1985, 2003). Растению свойственна гетерофиллия: наряду с подводными сидячими чешуевидными у него присутствуют плавающие и поднимающиеся за счет длинных черешков высоко над водой листья с округло-щитковидными листовыми пластинками. Чешуевидные листья плотно охватывают молодые почки, выполняют функцию специализированной почечной чешуи, после развертывания почки в побег отмирают (разрушаются). Цветки крупные, одиночные, пазушные, обоеполые, ярко-розовые.
N. nucifera встречается в Китае, Индокитае, на п‑ове Индостан, в Индии, на о. Цейлон, в южной Японии, на о-вах Малайского архипелага, Филиппинских о-вах, в Австралии, Республике Азербайджан, Иране (Крюкова, Снигиревская, 2008; Куприянова, 2016).
Территория России включается в северную часть ареала N. nucifera. В европейской части страны лотос распространен по берегам Каспийского моря (Гроссгейм, 1940; Снигиревская, 1988), в дельте р. Волга (Лабутина, Балдина, 2009). На Дальнем Востоке он растет в бассейне среднего и нижнего течения р. Амур, в дельте р. Зея, в старицах рек Бурея, Тунгуска, Уссури, на Приханкайской низменности и на о. Путятин (Качура, 1981; Крюкова, 2000; Кудрин, 1990, 2003; Куренцова, 1968; Харкевич и др., 1995). В результате интродукции N. nucifera появился в ряде лиманов в дельте р. Кубань и на побережье Азовского моря, где к настоящему времени образует обширные заросли (Лисицына, Папченков, 2000). В Краснодарском крае лотос орехоносный растет в Темрюкском районе, в Долине лотосов (Куприянова, 2016).
N. nucifera культивируется в ботанических садах г. Санкт-Петербург (Санкт-Петербургский государственный университет), г. Краснодар (Кубанский госуниверситет) и в дендрарии г. Сочи (Растения…, 2005). Он занесен в Красную книгу РФ (Крюкова, Снигиревская, 2008) со статусом III, региональные Красные книги Астраханской (Красная…, 2004) и Амурской областей (Красная книга…, 2009), Еврейской АО (Красная…, 2006), Хабаровского края (Красная…, 2008) и др. Охраняется на территориях заповедников Астраханский, Хинганский, Ханкайский и ряда памятников природы (Троицкий, 1953; Шехов, 1998).
N. nucifera в природе предпочитает заболоченные водоемы с проточной водой и медленным течением (Куренцова, 1968). Встречается по мелководьям (глубиной ≤2–2.5 м) рек и озер, имеющих мощный слой илистых донных отложений. Корневища лотоса располагаются в непромерзающем иле, что позволяет растению выносить морозы до –30°С (Куприянова, 2016). При благоприятных условиях лотос орехоносный часто образует монодоминантные заросли и выступает в качестве эдификатора сообществ водных растений.
К лимитирующим факторам, ограничивающим существование N. nucifera, относят загрязнение поверхностных вод, гидротехническое строительство, мелиоративные работы в поймах рек, выпас, рекреационные нагрузки, а также применение удобрений на площади возделывания риса, что ведет к нарушению естественных местообитаний. Лотос страдает от сбора цветков, плодов и подземных органов, которые поедают кабан и ондатра. Изменения условий среды, приводящие к полному осушению озер, также сокращают его ареал.
N. nucifera – один из древнейших представителей цветковых растений, реликт третичной флоры (Куренцова, 1968). Известен с древних времен своей пищевой и фармакологической ценностью (Куприянова, 2016). В связи с этим в настоящее время большое внимание уделяется вопросам генетики, интродукции и акклиматизации N. nucifera, мониторинга его распространения, влияния на состояние водоемов, практического применения (Кудрин, 1990, 2003; Куприянова, 2016; Лабутина, Балдина, 2009; Троицкий, 1953; Чуйков, 2013; Шехов, 1998; Ahn et al., 2014; Chaudhuri, Singh, 2013; Diao et al., 2016; Ki et al., 2009; Rho, Yoon, 2017).
Работы по биоморфологии и анатомии неизвестны. Однако именно такие данные позволяют определять запасы, состояние, возможное использование, а также охрану, воспроизведение ценопопуляций и создание искусственных плантаций вида с сохранением основных экосистемных функций естественных водоемов.
Цель работы – изучить структуру побеговой системы и анатомию органов N. nucifera на северной границе ареала в европейской части России.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Побеговые системы N. nucifera исследованы в посадках в дельте р. Волги в окрестностях с. Самосделка Астраханской обл. в августе 2017 г. В лабораторных условиях проведен анализ побеговых систем, промежуточных почек и поперечных срезов стебля, черешка и цветоноса. В результате натурных исследований изучено 32 особи, для камеральной обработки изъято десять побеговых систем.
За основу исследования принят разработанный И.Г. Серебряковым (Серебряков, 1952, 1954) на базе сравнительно-морфологического метода способ описания конкретных структур, модель побегообразования оценена в соответствии с подходами Т.И. Серебряковой (Серебрякова, 1977, 1981), модульная организация – согласно представлениям Н.П. Савиных (Савиных, 2002). Жизненная форма охарактеризована в соответствии с классификацией И.Г. Серебрякова (Серебряков, 1964), экологическая группа – по В.Г. Папченкову (Папченков, 1985, 2003). Для лучшего микроскопирования анатомические срезы окрашивали по методу Мейле – перманганатная реакция (Барыкина и др., 2004). В дальнейшем их анализировали, используя микроскоп Motic BA300 со встроенным видеоокуляром.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
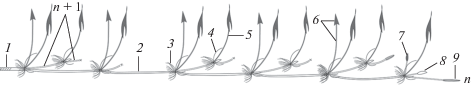
Из грунта было изъято несколько побеговых систем. Они состояли из оси n-го порядка длиной до 7 м с сформировавшимися центрами закрепления в каждом узле и отходящими от них боковыми осями длиной до 3 м (рис. 1). Междоузлия главной оси длинные (до 1 м) и толстые (до 1.8 см). У боковых осей они того же строения, но короче и тоньше. В побеговой системе N. nucifera закономерно и последовательно чередуются два элементарных модуля. Первый образован длинным междоузлием, узлом с чешуевидным листом и генеративной пазушной почкой или без нее (у боковых осей). Он обеспечивает расселение и репродукцию растения. Далее следует второй элементарный модуль из короткого междоузлия, узла с листом срединной формации и почкой или побегом замещения в его пазухе. Этот модуль служит в основном для ассимиляции, ветвления, заполнения пространства, занятого исходной осью. Структура из модулей повторяется ритмично и закономерно в течение всего вегетационного сезона, соответствует элементарному побегу (Грудзинская, 1960), формирующемуся за один период роста.
Рис. 1.
Фрагмент побеговой системы N. nucifera: n, n + 1 – порядки ветвления побегов; 1 – отмирающая часть побега; 2 – часть побега текущего года; 3 – придаточные корни; 4 – чешуевидный лист; 5 – воздушный лист; 6 – цветок; 7 – генеративная почка; 8 – вегетативная почка; 9 – вегетативно-генеративная почка.
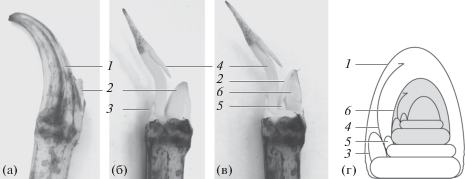
Развитие побеговых систем определяется строением почек лотоса. Мы изучили почки побегов всех порядков ветвления в собранных побеговых системах. Они размером 5 × 1.8 см, имеют на поверхности мясистый чешуевидный лист (рис. 2а). В его пазухе находится генеративная почка (рис. 2б). Далее располагается зачаток воздушного листа с особой структурой из двух почек, закрытых общей чешуей (рис. 2в). Меньшая по размеру почка представляет собой пазушную почку листа срединной формации. В будущем она реализуется в побег замещения, выполняющий функцию ветвления и расселения (у этих побегов мы не наблюдали цветков и плодов). Более крупная почка – апикальная, вегетативно-генеративная, в ней заложены структуры двух элементарных модулей следующего кванта роста и конус нарастания (рис. 2г).
Рис. 2.
Промежуточная почка N. nucifera: а – с чешуевидным листом, б – без чешуевидного листа, в – продольный разрез через структуры, г – схема строения (серым цветом отмечен один квант роста). 1 – чешуевидный лист, 2 – почки, закрытые чешуевидным листом, 3 – генеративная почка, 4 – воздушный лист, 5 – вегетативная почка, 6 – вегетативно-генеративная почка.
Развитие апикальной почки начинается с формирования длинного междоузлия и развертывания воздушного листа. Чешуевидный лист при этом разрывается, верхушка оси отдаляется от исходного центра за счет формирования длинного междоузлия первого элементарного модуля следующего кванта роста. Позднее одновременно развиваются обе боковые структуры: цветок и побег замещения. Образуются узловые придаточные корни. Так формируется центр закрепления N. nucifera (рис. 3а). Он состоит из двух сближенных узлов с разрушившимся чешуевидным листом и листом срединной формации, оснований цветоножки, бокового побега замещения и длинного междоузлия следующего элементарного побега, узловых придаточных корней (рис. 3б). У таких структур нет почек для формирования побегов в будущем. По-видимому, основная функция их – это закрепление, минеральное питание и репродукция. В них все реализуется здесь и сейчас.
Рис. 3.
Центр закрепления N. nucifera: а – внешний вид; б – схема; условные обозначения см. на рис. 1.

К концу вегетационного сезона нарастание осей прекращается, в стебле апикальных почек откладывается крахмал, формируются специализированные диаспоры, подобные турионам, которые обеспечивают перезимовку и вегетативное размножение. Именно эти структуры используются в пищу.
Морфологическая целостность особи длится не более двух лет и может прекратиться в конце первого вегетационного сезона в ходе морфологической дезинтеграции при разрушении длинных междоузлий и отделении парциальных структур в виде центров закрепления и диаспор. Таким образом, N. nucifera расселяется за счет длинных междоузлий, а размножается вегетативно путем их разрушения (неспециализированная морфологическая дезинтеграция) и специализированными диаспорами.
Традиционно это растение в определителях и флорах называют многолетним. Детальный морфологический анализ показал, что многолетние части в структуре особей у N. nucifera отсутствуют из-за раннего морфологического распада особей. Поэтому бóльшую часть своей жизни растение существует в виде клона, состоящего из отдельных недолго живущих особей вегетативного происхождения. Заросли N. nucifera формируются за счет высокой энергии вегетативного размножения с образованием потомков с многочисленными длинными придаточными корнями. Это обеспечивает длительное существование растения на занятой территории. На основании вышеизложенного жизненную форму лотоса определяем как поликарпик; летне-зеленый вегетативно подвижный полицентрический малолетник с моноподиальной полурозеточной моделью побегообразования и ранней неспециализированной морфологической дезинтеграцией.
Специфические особенности N. nucifera определены и на тканевом уровне. Согласно общепринятым представлениям, в строении стебля выделяют три анатомо-топографические зоны: покровную ткань, первичную кору и центральный цилиндр.
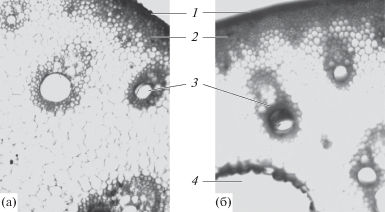
Покровная ткань представлена эпидермой, ее клетки на поперечном срезе имеют цилиндрическую форму. Клеточные стенки утолщены с наружной и боковых сторон, что обеспечивает их прочность и противостояние на разрыв (рис. 4а).
Рис. 4.
Эпидерма с первичной корой (а) и фрагмент поперечного среза стебля (б) N. nucifera. 1 – клетки эпидермы, 2 – округлые паренхимные клетки первичной коры, 3 – уплощенные паренхимные клетки первичной коры, 4 – проводящие пучки, 5 – склеренхима, 6 – флоэма, 7 – сосуды ксилемы, 8 – древесинная паренхима, 9, 10 – воздухоносные полости.

Первичная кора занимает небольшой объем стебля, слагается пятью-шестью слоями клеток округлой или неправильной формы, близкой к ней (рис. 4а). Некоторые из них уплощаются и приобретают неправильную прямоугольную либо близкую к шиловидной форму (рис. 4а). Кора представлена только паренхимными клетками без включения механических элементов, видимо, из-за условий местообитания растения: ось располагается горизонтально, а цветки и листья поддерживаются выталкивающей силой воды.
Глубже расположена стела, не отделенная от первичной коры эндодермой, так как последняя не выражена. Проводящие пучки, в отличие от типичных для двудольных растений, располагаются в несколько концентрических кругов. Ближе к центральной полости их размеры увеличиваются, в тканях паренхимы между пучками образуются крупные межклетники (рис. 4б). Паренхимные клетки содержат много крахмальных зерен, что обеспечивает запас питательных веществ и в стебле.
Сосудисто-волокнистые пучки коллатеральные закрытые, что несвойственно для двудольных. Они имеют небольшую механическую обкладку из склеренхимы (рис. 4б). Проводящие ткани представлены ксилемой и флоэмой, камбий между ними отсутствует.
Флоэма составлена ситовидными трубками, клетками спутницами и лубяной паренхимой (рис. 4б, 6). Ксилема включает два крупных сосуда, у мелких пучков может быть один (рис. 4б, 7). Также в состав пучка входит воздухоносная полость.
Таким образом, строение сосудисто-волокнистого пучка N. nucifera типично для проводящих пучков однодольных растений.
Цветоножка и черешок ассимилирующего листа имеют одинаковое строение и расположение тканей (рис. 5). Покровная ткань, как и в стебле, представлена эпидермой идентичного строения. Под ней расположен слой механической ткани – склеренхимы, который не выражен в стебле. Появление этой ткани в структуре органов связано с их вертикальным, а также с внутриводным положением и обеспечивает поддержание листьев с крупными листовыми пластинками и цветка. Дальнейшее строение внутренней структуры идентично описанному в стебле.
Рис. 5.
Фрагмент поперечного среза цветоножки (а) и черешка листа (б) N. nucifera. 1 – покровная ткань, 2 – склеренхима, 3 – проводящие пучки, 4 – воздухоносная полость.
Внутреннее строение стебля, черешка воздушного листа и цветоноса имеет строение, типичное для однодольных растений, что противоречит принадлежности N. nucifera к классу двудольных. Неравномерное утолщение клеточных стенок эпидермы, наличие склеренхимы вокруг проводящих пучков стебля и в первичной коре черешка и цветоноса, крупные воздухоносные полости обеспечивают успешное произрастание лотоса в воде.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Побегообразование лотоса аналогично квантированному росту, описанному для наземных древесных и травянистых растений (Серебрякова, 1971; Шилова, 1988; Шмидт, 1968), а также водных трав (Лапиров, 2008). Квантированность роста у лотоса определяется, видимо, чередованием процессов расселения/цветения и ассимиляции, когда цветки образуются в пазухе чешуевидного листа модуля с длинным междоузлием каждый раз после развертывания ассимилирующего листа. В результате побеговая система N. nucifera состоит из полурозеточных поликарпических (из-за многократного разновременного заложения цветков в структуре элементарных побегов в течение вегетационного сезона) вегетативно-генеративных осей, развивающихся по моноподиальной полурозеточной модели побегообразования.
Побегообразование N. nucifera с периодическим формированием ритмически развивающихся репродуктивных структур, беспрерывным образованием за вегетационный период нескольких следующих друг за другом побегов ветвления и растянутым периодом семенной репродукции аналогично итеративному ветвлению (термин по: Troll, 1964) водных трав. Сходство состоит в ритмичном образовании побегов и их типе. Отличие наблюдается в способе нарастания побеговых систем: моноподиальное с образованием элементарных побегов у лотоса и симподиальное с формированием побегов замещения – при типичном итеративном ветвлении. Более того у побеговых систем лотоса с образованием элементарных побегов не увеличивается число функционально единых осей. По пути типичного итеративного ветвления других водных трав у лотоса образуются побеги замещения. Они формируются одновременно с нарастанием главной оси, в то время как при симподиальном нарастании главная ось прекращает свое развитие. В результате за один вегетационный сезон формируются побеговые системы в виде моноподиев нескольких порядков ветвления, а растение быстро не только занимает значительные пространства, но и заполняет их за счет развития боковых осей.
Выводы. N. nucifera – древнейший представитель цветковых растений из числа вторично водных трав, сохранивший исходные признаки этой группы. В их числе – квантированное моноподиальное нарастание с образованием элементарных побегов в сочетании с итеративным ветвлением, за счет чего растение не только занимает пространство, но и осваивает его. По-видимому, структурные единицы из двух элементарных модулей, возникающих ритмически и закономерно в ходе моноподиального нарастания, у N. nucifera можно признать промежуточными модулями (термин по: Бобров, 2008), еще одной их категорией наряду с элементарными и универсальными (одноосными побегами). Анатомическое строение стеблей, черешков и цветоносов лотоса с типичной структурой таковых у эволюционно более продвинутых растений из класса Однодольные демонстрирует гетерохронию – разновозрастность признаков в пределах одного таксона. Поскольку настоящее исследование проведено на небольшом объеме материала в виду охраны вида, требуется дальнейшее изучение N. nucifera, в частности его сезонного развития и онтоморфогенеза.
Список литературы
Барыкина Р.П., Веселова Т.Д., Девятов А.Г. и др. 2004. Справочник по ботанической микротехнике. Основы и методы. Москва: Изд-во МГУ.
Бобров Ю.А. 2008. Промежуточный модуль – вспомогательный уровень морфологического анализа // Вестник Тверского государственного университета. Сер. биология и экология. № 9. С. 16.
Гроссгейм А.А. 1940. Лотос в СССР // Бот. материалы Гербария Бот. ин-та АН СССР. Т. 8. Вып. 4–9. С. 130.
Грудзинская И.А. 1960. Летнее побегообразование у древесных растений и его классификация // Бот. журн. Т. 45. № 7. С. 968.
Качура Н.Н. 1981. Редкие виды растений советского Дальнего Востока и их охрана. Москва: Наука.
Красная книга Амурской области: Редкие и находящиеся под угрозой исчезновения виды животных, растений и грибов. 2009. Благовещенск: Изд-во Белорус. гос. пед. ун-та.
Красная книга Астраханской области: Редкие и находящиеся под угрозой исчезновения объекты животного и растительного мира. 2004. Астрахань: Нижневолжский центр экологического образования.
Красная книга Еврейской автономной области. Редкие и находящиеся под угрозой исчезновения виды растений и грибов. 2006. Новосибирск: АРТА.
Красная книга Хабаровского края. Редкие и находящиеся под угрозой исчезновения виды растений и животных. 2008. Хабаровск: Издательский дом “Приамурские ведомости”.
Крюкова М.В. 2000. Флористические находки на Среднеамурской низменности // Бот. журн. Т. 85. № 11. С. 122.
Крюкова М.В., Снигиревская Н.С. 2008. Семейство Лотосовые – Nelumbonaceae // Красная книга Российской Федерации (растения и грибы). Москва: Товарищество научных изданий КМК. С. 348.
Кудрин С.Г. 1990. Водные реликты Хинганского государственного заповедника // Бот. журн. Т. 75. № 10. С. 1461.
Кудрин С.Г. 2003. Динамика восстановления Nelumbo komarovii (Nelumbonaceae) на юго-востоке Амурской области // Бот. журн. Т. 88. № 8. С. 83.
Куприянова Н.Б. 2016. Краснодарский лотос: успешность интродукции и акклиматизации // Ресурсы региона: культурно-историческое развитие в контексте науки и образования. Славянск-на-Кубани: Кубанский государственный университет. С. 248.
Куренцова Г.Э. 1968. Реликтовые растения Приморья. Ленинград: Наука.
Лабутина И.А., Балдина Е.А. 2009. Мониторинг распространения лотоса в дельте Волги // Вестник Московского университета. Сер. 5: География. № 9. С. 27.
Лапиров А.Г. 2008. Явление квантированности роста в метамерной организации побега некоторых представителей семейства Аlismataceae Vent. // Вестник Тверского государственного университета. Серия “Биология и экология”. Вып. 9. С. 122.
Лисицына Л.И., Папченков В.Г. 2000. Флора водоемов России: Определитель сосудистых растений. Москва: Наука.
Папченков В.Г. 1985. О классификации макрофитов водоeмов // Экология. № 6. С. 8.
Папченков В.Г. 2003. О классификации растений водоeмов и водотоков // Гидроботаника: методология, методы: Матер. Школы по гидроботанике. Рыбинск: Дом печати. С. 23.
Растения Красной книги России в коллекциях ботанических садов и дендрариев. 2005. Москва: Гл. бот. сад РАН; Тула: Гриф и К.
Савиных Н.П. 2002. Модули у растений // Тез. докл. II Междунар. конф. по анат. и морфол. растений. Санкт-Петербург: Ботанический ин-т им. В.Л. Комарова РАН. С. 95.
Серебряков И.Г. 1952. Морфология вегетативных органов высших растений. Москва: Сов. наука.
Серебряков И.Г. 1954. О методах изучения ритмики сезонного развития растений в стационарных геоботанических исследованиях // Уч. записки МГПИ им. В.П. Потемкина. Т. 37. Вып. 2. С. 3.
Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение // Полевая геоботаника. Т. 3. Москва: Наука. С. 148.
Серебрякова Т.И. 1971. Морфогенез побегов и эволюция жизненных форм злаков. Москва: Наука.
Серебрякова Т.И. 1977. Об основных “архитектурных моделях” травянистых многолетников и модусах их преобразования // Бюл. МОИП. Отд. биол. Т. 82. № 5. С. 112.
Серебрякова Т.И. 1981. Жизненные формы и модели побегообразования наземно-ползучих многолетних трав // Жизненные формы, структура, спектры, эволюция. Москва: Наука. С. 161.
Снигиревская Н.С. 1988. Лотос орехоносный. Nelumbo nucifera Gaertn. // Красная книга РСФСР (растения). Москва: Росагропромиздат. С. 291.
Троицкий С.К. 1953. Лотос в Кубанских лиманах // Природа. № 9. С. 119.
Харкевич С.С., Старченко В.М., Дарман Г.Ф. и др. 1995. Редкие и исчезающие растения Амурской области. Благовещенск: Изд-во Амур. бот. сада НЦ ДВО РАН.
Чуйков Ю.С. 2013. Лотос орехоносный в дельте Волги – охрана и возможное использование // Астраханский Вестник экологического образования. № 3(25). Астрахань: Нижневолжский экоцентр. С. 145.
Шехов А.Г. 1998. Интродукция лотосов на Кубани // Природа. № 6. Москва: Наука. С. 37.
Шилова Н.В. 1988. Ритмы роста и пути структурной адаптации тундровых растений. Ленинград: Наука.
Шмидт В.М. 1968. О квантированности роста и органогенеза у растений // Вестн. ЛГУ. Вып. 1. № 3. С. 99.
Ahn J.H., Kim S.B., Kim E.S. et al. 2014. A new flavolignan flom Nelumbo nucifera leaves // Chem. Nat. Compd. V. 50. № 6. P. 998.
Chaudhuri P.K., Singh D. 2013. A new triterpenoid from the rhizomes of Nelumbo nucifera // Natural Product Research. V. 27. № 6. P. 532.
Diao Y., Li G.L., Yu A.Q. et al. 2016. Cloning and characterization of the UBC gene from lotus (Nelumbo nucifera) // Genet. Mol. Res. V. 15. № 3. P. 1.
Ki H.K., Sang W.C., Shi Y.R. et al. 2009. Phytochemical constituents of Nelumbo nucifera // Nat. Prod. Sci. V. 15. № 2. P. 90.
Rho T., Yoon K.D. 2017. Chemical Constituents of Nelumbo nucifera Seeds // Nat. Prod. Sci. V. 23. № 4. P. 253.
Troll W. 1964. Die Infloreszenzen. B. 1. Jena: Fischer.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод