

Биология внутренних вод, 2020, № 6, стр. 538-549
Структура фенетического разнообразия и систематика красноперки Scardinius erythrophthalmus (Cypriniformes, Cyprinidae) водоемов и водотоков России и сопредельных стран
А. В. Кожара a, *, А. С. Маврин a, А. Н. Мироновский a, b
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
* E-mail: akozhara@mail.ru
Поступила в редакцию 19.11.2019
После доработки 10.12.2019
Принята к публикации 26.03.2020
Аннотация
Методами популяционной морфологии изучена структура фенетического разнообразия красноперки Scardinius erythrophthalmus (L.) на территории РФ и ряда сопредельных стран. Использованы собственные коллекционные и литературные данные по счетным признакам внешней морфологии, осевого скелета, сейсмосенсорной системы, а также формуле глоточных зубов. В пределах изученной части ареала выделены две фенетически четко обособленные популяционные системы – западная, охватывающая Верхнюю и Среднюю Волгу и водные объекты к юго-западу и западу от волжского бассейна, и юго-восточная, включающая красноперок Нижней Волги, Прикаспия и бессточных рек и озер Казахстана вплоть до рек Сырдарья и Чу. Основные признаки, вносящие вклад в межпопуляционную дифференциацию – число чешуй в боковой линии, общее число позвонков, распределение позвонков по отделам позвоночника и число жаберных тычинок. Показано, что тесная положительная корреляция первых двух признаков с географической широтой мест обитания связана, в первую очередь, не с прямым воздействием физических факторов, функционально зависящих от широты, а с процессами формообразования в речных бассейнах. Полученные результаты не согласуются с существующим представлением о красноперке как о мономорфном виде и позволяют поставить вопрос о таксономической ревизии этого вида в его современном объеме.
ВВЕДЕНИЕ
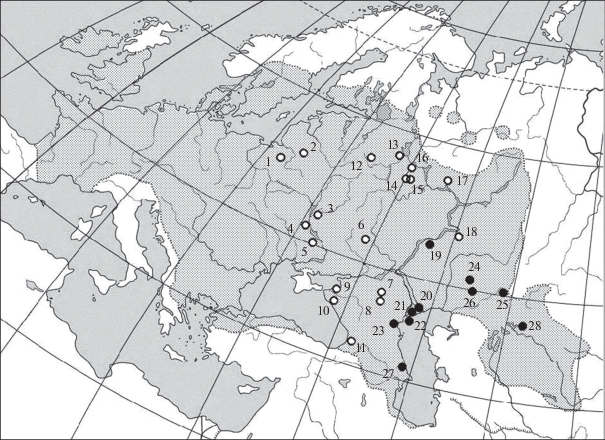
Обыкновенная красноперка Scardinius erythrophthalmus (L.) относится к широко распространенным видам семейства карповых рыб. В настоящее время в роде Scardinius выделяют десять аллопатрических видов, из которых восемь обитают локально в южной Европе (преимущественно на Апеннинском полуострове и на Балканах) и еще один в южной Анатолии (Kottelat, Freyhof, 2007; Fricke et al., 2007). Ареал обыкновенной красноперки охватывает почти всю Европу к северу от Пиренеев и Альп – от Британских островов до Урала. На юге и юго-востоке он включает восточную и северо-восточную Грецию с прилегающей частью Турции (бассейн Эгейского моря), Малую Азию (бассейны Мраморного и Черного морей), западное и восточное Закавказье и часть Средней Азии (бассейны рек Амударья, Сырдарья и бывших притоков последней – реки Сарысу и Чу). Красноперка отсутствует в Крыму, а также на севере Европы: отмечена только в южной части Скандинавии, а ее распространение в Ледовитоморском бассейне ограничивается несколькими озерами бассейнов верхнего течения рек Онега и Северная Двина, где обитают изолированные реликтовые популяции (Атлас …, 2003; Бознак, 2008; Дворянкин, 2014; Kottelat, Freyhof, 2007) (рис. 1). Эта бентопелагическая фитофильная рыба предпочитает эвтрофированные озера и медленные участки рек с богатой водной растительностью, встречается также в распресненных частях лиманов внутренних морей, достигая местами большой численности (Атлас …, 2003; Kottelat, Freyhof, 2007).
Рис. 1.
Схема ареала красноперки (показан пунктирной линией с заливкой) по совокупности литературных данных и места сбора материала (1–28). Описание см. в табл. 1. ⚪ – западная (“типичная”) форма красноперки, ⚫ – юго-восточная форма.
В настоящее время вид S. erythrophthalmus считается мономорфным. Для него не описаны подвиды или сколь-нибудь фенотипически своеобразные формы в отдельных регионах или речных бассейнах. Красноперки, которым в свое время присваивался подвидовой статус в рамках вида S. erythrophthalmus, сегодня рассматриваются как самостоятельные виды. Например, статус красноперки из южной Анатолии, изначально описанной Богуцкой (1997) в качестве подвида S. erythrophthalmus elmaliensis, в дальнейшем был повышен до видового – S. elmaliensis (Fricke et al., 2007). Внутривидовая изменчивость обыкновенной красноперки изучена плохо. До сих пор не проводилось масштабного популяционно-морфологического исследования данного вида с целью получить целостную картину внутривидовой подразделенности и выявить структуру фенетического и генетического разнообразия. Все выполненные к настоящему моменту работы носят локальный либо региональный характер, затрагивая лишь отдельные водные объекты или бассейны (Митрофанов и др., 1987; Мовчан, Смiрнов, 1981; Mavrin et al., 2017; Ilhan, 2019).
Цель работы – проанализировать СФР красноперки в пределах России и сопредельных стран и выявить основные тенденции ее географической и экологической изменчивости.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Материалом для исследования послужили полевые сборы красноперки в период с 1983 по 2014 гг. из водоемов и водотоков Российской Федерации, Казахстана, Украины, Грузии, Азербайджана и Литвы (табл. 1). Взрослых особей отлавливали ставными сетями, 7-метровой мальковой волокушей, 30- или 50-метровым неводом и удочкой. После измерений и подсчетов на свежем материале и последующей обработки рыб горячей водой кости черепа и осевые скелеты очищали от мягких тканей, тщательно промывали в теплой воде и высушивали (Яковлев и др., 1981). Анализировали следующие наружные и остеологические признаки: D, A, ll, CPMd, CPMp, CSOf, CSTp, Va, Vi, Vc, Vt, D. ph1 и D. ph2.
Таблица 1.
Статистические характеристики изменчивости наружных и остеологических счетных признаков красноперки
| № | Водоем | Год | n | D | A | ll | Va | Vi | Vc | Vt | CPMp | CPMd | CSOf | CSTp | L, с.ш. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Оз. Виштынецкое | 1986 | 13 | $\frac{{8.23 \pm 0.17}}{{7 - 9}}$ | $\frac{{10.62 \pm 0.18}}{{10 - 12}}$ | $\frac{{41.31 \pm 0.35}}{{39 - 43}}$ | $\frac{{19.62 \pm 0.21}}{{19 - 21}}$ | $\frac{{2.54 \pm 0.14}}{{2 - 3}}$ | $\frac{{17.62 \pm 0.14}}{{17 - 18}}$ | $\frac{{39.77 \pm 0.23}}{{38 - 41}}$ | $\frac{{8.27 \pm 0.29}}{{6 - 10}}$ | $\frac{{4.69 \pm 0.05}}{{4 - 6}}$ | $\frac{{5.27 \pm 0.08}}{{3 - 8}}$ | $\frac{{1.93 \pm 0.13}}{{0 - 3}}$ | 54.41 |
| 2 | Оз. Дрингис, Литва | 1985 | 27 | $\frac{{8.19 \pm 0.09}}{{7 - 9}}$ | $\frac{{11.07 \pm 0.11}}{{10 - 12}}$ | $\frac{{41.49 \pm 0.18}}{{40 - 43}}$ | $\frac{{18.96 \pm 0.14}}{{18 - 20}}$ | $\frac{{2.78 \pm 0.10}}{{2 - 4}}$ | $\frac{{17.78 \pm 0.08}}{{17 - 18}}$ | $\frac{{39.48 \pm 0.11}}{{39 - 41}}$ | $\frac{{8.54 \pm 0.18}}{{7 - 10}}$ | $\frac{{5.26 \pm 0.14}}{{4 - 7}}$ | $\frac{{5.94 \pm 0.15}}{{5 - 8}}$ | $\frac{{2.48 \pm 0.04}}{{1 - 5}}$ | 55.5 |
| 3 | Киевское вдхр | 26 | $\frac{{8.04 \pm 0.07}}{{7 - 9}}$ | $\frac{{10.72 \pm 0.12}}{{10 - 12}}$ | $\frac{{41.16 \pm 0.16}}{{39 - 43}}$ | $\frac{{19.13 \pm 0.11}}{{17 - 20}}$ | $\frac{{2.87 \pm 0.07}}{{2 - 3}}$ | $\frac{{17.30 \pm 0.15}}{{16 - 19}}$ | $\frac{{39.30 \pm 0.16}}{{38 - 41}}$ | $\frac{{8.22 \pm 0.12}}{{6 - 10}}$ | $\frac{{4.67 \pm 0.09}}{{3 - 6}}$ | $\frac{{5.70 \pm 0.13}}{{4 - 8}}$ | $\frac{{1.63 \pm 0.14}}{{0 - 3}}$ | 51.04 | |
| 4 | Каневское вдхр | 1988 | 57 | $\frac{{8.30 \pm 0.07}}{{8 - 10}}$ | $\frac{{10.96 \pm 0.09}}{{9 - 12}}$ | $\frac{{40.99 \pm 0.15}}{{37 - 43}}$ | $\frac{{19.18 \pm 0.07}}{{18 - 20}}$ | $\frac{{2.71 \pm 0.06}}{{2 - 3}}$ | $\frac{{17.23 \pm 0.06}}{{16 - 18}}$ | $\frac{{39.14 \pm 0.07}}{{38 - 40}}$ | $\frac{{8.58 \pm 0.09}}{{6 - 11}}$ | $\frac{{4.66 \pm 0.07}}{{3 - 6}}$ | $\frac{{5.83 \pm 0.08}}{{4 - 8}}$ | $\frac{{1.79 \pm 0.06}}{{0 - 4}}$ | 50.0 |
| 5 | Кременчугское вдхр | 1988 | 39 | $\frac{{8.15 \pm 0.06}}{{8 - 9}}$ | $\frac{{10.75 \pm 0.10}}{{10 - 12}}$ | $\frac{{40.86 \pm 0.15}}{{38 - 43}}$ | $\frac{{19.32 \pm 0.09}}{{19 - 20}}$ | $\frac{{2.74 \pm 0.08}}{{2 - 3}}$ | $\frac{{17.10 \pm 0.08}}{{16 - 18}}$ | $\frac{{39.16 \pm 0.11}}{{38 - 40}}$ | $\frac{{8.21 \pm 0.11}}{{7 - 11}}$ | $\frac{{4.52 \pm 0.08}}{{3 - 6}}$ | $\frac{{5.75 \pm 0.11}}{{4 - 7}}$ | $\frac{{1.70 \pm 0.09}}{{0 - 3}}$ | 49.45 |
| 6 | Р. Оскол | 1999–2000 | 16 | $\frac{{8.19 \pm 0.10}}{{8 - 9}}$ | $\frac{{10.63 \pm 0.13}}{{10 - 11}}$ | $\frac{{40.58 \pm 0.24}}{{39 - 43}}$ | $\frac{{19.25 \pm 0.16}}{{19 - 20}}$ | $\frac{{2.75 \pm 0.16}}{{2 - 3}}$ | $\frac{{17.0}}{{17}}$ | $\frac{{39.0}}{{39}}$ | – | – | – | – | 51.0 |
| 7 | Р. Джурак-Сал | 1990 | 16 | $\frac{{8.13 \pm 0.09}}{{8 - 9}}$ | $\frac{{10.69 \pm 0.12}}{{10 - 11}}$ | $\frac{{41.00 \pm 0.22}}{{40 - 43}}$ | $\frac{{19.33 \pm 0.19}}{{18 - 21}}$ | $\frac{{2.88 \pm 0.15}}{{2 - 4}}$ | $\frac{{16.75 \pm 0.11}}{{16 - 17}}$ | $\frac{{38.93 \pm 0.15}}{{38 - 40}}$ | $\frac{{8.69 \pm 0.23}}{{7 - 10}}$ | $\frac{{4.88 \pm 0.20}}{{3 - 6}}$ | $\frac{{5.94 \pm 0.30}}{{4 - 9}}$ | $\frac{{1.78 \pm 0.14}}{{1 - 3}}$ | 46.6 |
| 8 | Р. Чекалда | 1990 | 62 | $\frac{{8.40 \pm 0.06}}{{8 - 9}}$ | $\frac{{10.90 \pm 0.09}}{{9 - 11}}$ | $\frac{{40.79 \pm 0.14}}{{39 - 44}}$ | $\frac{{19.49 \pm 0.07}}{{19 - 21}}$ | $\frac{{2.77 \pm 0.07}}{{2 - 4}}$ | $\frac{{16.46 \pm 0.09}}{{15 - 18}}$ | $\frac{{38.75 \pm 0.08}}{{37 - 40}}$ | $\frac{{8.48 \pm 0.08}}{{6 - 11}}$ | $\frac{{4.61 \pm 0.06}}{{3 - 6}}$ | $\frac{{5.69 \pm 0.08}}{{4 - 8}}$ | $\frac{{1.84 \pm 0.05}}{{0 - 3}}$ | 46.35 |
| 9 | Красный лиман, Азовское море | 1985 | 37 | $\frac{{8.05 \pm 0.04}}{{8 - 9}}$ | $\frac{{10.46 \pm 0.09}}{{9 - 11}}$ | $\frac{{40.17 \pm 0.16}}{{39 - 43}}$ | $\frac{{19.19 \pm 0.10}}{{18 - 21}}$ | $\frac{{2.95 \pm 0.08}}{{2 - 4}}$ | $\frac{{16.51 \pm 0.09}}{{15 - 17}}$ | $\frac{{38.65 \pm 0.11}}{{38 - 41}}$ |
$\frac{{8.33 \pm 0.11}}{{6 - 10}}$ |
$\frac{{4.64 \pm 0.10}}{{3 - 6}}$ | $\frac{{6.11 \pm 0.16}}{{4 - 9}}$ | $\frac{{1.82 \pm 0.08}}{{0 - 4}}$ | 45.97 |
| 10 | Оз. Абрау | 1998 | 22 | $\frac{{8.23 \pm 0.09}}{{8 - 9}}$ | $\frac{{10.55 \pm 0.11}}{{10 - 11}}$ | $\frac{{41.72 \pm 0.20}}{{40 - 43}}$ | $\frac{{19.17 \pm 0.17}}{{18 - 20}}$ | $\frac{{2.67 \pm 0.11}}{{2 - 3}}$ | $\frac{{17.00 \pm 0.11}}{{16 - 18}}$ | $\frac{{38.83 \pm 0.12}}{{38 - 40}}$ | $\frac{{8.78 \pm 0.18}}{{7 - 10}}$ | $\frac{{4.72 \pm 0.15}}{{4 - 6}}$ | $\frac{{5.50 \pm 0.22}}{{3 - 7}}$ | $\frac{{1.90 \pm 0.16}}{{1 - 4}}$ | 44.69 |
| 11 | Оз. Палеостоми, Грузия | 1986 | 10 | $\frac{{8.00 \pm 0.15}}{{7 - 9}}$ | $\frac{{10.50 \pm 0.17}}{{10 - 11}}$ | $\frac{{39.90 \pm 0.23}}{{39 - 41}}$ | $\frac{{19.30 \pm 0.15}}{{19 - 20}}$ | $\frac{{2.70 \pm 0.15}}{{2 - 3}}$ | $\frac{{16.90 \pm 0.18}}{{16 - 18}}$ | $\frac{{38.90 \pm 0.18}}{{38 - 40}}$ | $\frac{{8.50 \pm 0.15}}{{7 - 9}}$ | $\frac{{4.67 \pm 0.21}}{{4 - 7}}$ | $\frac{{5.80 \pm 0.19}}{{5 - 8}}$ | $\frac{{2.15 \pm 0.13}}{{1 - 3}}$ | 42.11 |
| 12 | Оз. Селигер | 1990 | 32 | $\frac{{8.25 \pm 0.08}}{{8 - 9}}$ | $\frac{{11.25 \pm 0.14}}{{9 - 13}}$ | $\frac{{41.87 \pm 0.15}}{{40 - 44}}$ | $\frac{{19.28 \pm 0.09}}{{18 - 20}}$ | $\frac{{2.75 \pm 0.09}}{{2 - 4}}$ | $\frac{{17.69 \pm 0.09}}{{17 - 19}}$ | $\frac{{39.72 \pm 0.09}}{{39 - 41}}$ | $\frac{{7.89 \pm 0.11}}{{5 - 10}}$ | $\frac{{4.56 \pm 0.08}}{{3 - 6}}$ | $\frac{{6.00 \pm 0.10}}{{5 - 7}}$ | $\frac{{1.93 \pm 0.07}}{{1 - 3}}$ | 57.19 |
| 13 | Рыбинское вдхр | 2011–2013 | 107 | $\frac{{8.00 \pm 0.03}}{{7 - 9}}$ | $\frac{{10.95 \pm 0.07}}{{8 - 13}}$ | $\frac{{41.47 \pm 0.13}}{{37 - 44}}$ | $\frac{{19.98 \pm 0.05}}{{18 - 21}}$ | $\frac{{2.30 \pm 0.05}}{{1 - 3}}$ | $\frac{{17.61 \pm 0.06}}{{16 - 19}}$ | $\frac{{39.89 \pm 0.06}}{{37 - 41}}$ | $\frac{{8.01 \pm 0.09}}{{6 - 10}}$ | $\frac{{4.44 \pm 0.07}}{{3 - 6}}$ | $\frac{{5.82 \pm 0.09}}{{3 - 9}}$ | $\frac{{1.92 \pm 0.06}}{{0 - 4}}$ | 58.06 |
| 14 | Оз. Юхор, | 2004–2005 | 107 | $\frac{{8.15 \pm 0.07}}{{8 - 9}}$ | $\frac{{10.92 \pm 0.11}}{{10 - 13}}$ | $\frac{{41.28 \pm 0.31}}{{39 - 44}}$ | $\frac{{19.84 \pm 0.11}}{{18 - 21}}$ | $\frac{{2.47 \pm 0.11}}{{1 - 4}}$ | $\frac{{17.21 \pm 0.11}}{{16 - 18}}$ | $\frac{{39.57 \pm 0.13}}{{38 - 41}}$ | $\frac{{8.32 \pm 0.16}}{{6 - 11}}$ | $\frac{{4.38 \pm 0.14}}{{2 - 6}}$ | $\frac{{5.93 \pm 0.17}}{{4 - 8}}$ | $\frac{{1.75 \pm 0.12}}{{0 - 3}}$ | 56.38 |
| 15 | Оз. Санхар | 2004 | 50 | $\frac{{8.16 \pm 0.05}}{{8 - 9}}$ | $\frac{{11.28 \pm 0.09}}{{10 - 13}}$ | $\frac{{41.41 \pm 0.09}}{{40 - 42}}$ | $\frac{{19.96 \pm 0.05}}{{19 - 21}}$ | $\frac{{2.38 \pm 0.07}}{{2 - 3}}$ | $\frac{{17.10 \pm 0.07}}{{16 - 18}}$ | $\frac{{39.44 \pm 0.08}}{{38 - 41}}$ | $\frac{{7.83 \pm 0.10}}{{6 - 10}}$ | $\frac{{4.48 \pm 0.11}}{{3 - 7}}$ | $\frac{{5.72 \pm 0.10}}{{4 - 7}}$ | $\frac{{1.91 \pm 0.09}}{{0 - 3}}$ | 56.4 |
| 16 | Оз. Чистое, | 2009 | 36 | $\frac{{8.33 \pm 0.05}}{{8 - 9}}$ | $\frac{{11.22 \pm 0.11}}{{9 - 12}}$ | $\frac{{42.03 \pm 0.13}}{{41 - 44}}$ | $\frac{{19.97 \pm 0.06}}{{19 - 21}}$ | $\frac{{2.22 \pm 0.07}}{{2 - 3}}$ | $\frac{{17.61 \pm 0.11}}{{16 - 19}}$ | $\frac{{39.81 \pm 0.10}}{{39 - 41}}$ | $\frac{{8.43 \pm 0.14}}{{7 - 11}}$ | $\frac{{4.38 \pm 0.13}}{{3 - 6}}$ | $\frac{{6.13 \pm 0.13}}{{4 - 8}}$ | $\frac{{2.08 \pm 0.09}}{{1 - 4}}$ | 57.71 |
| 17 | Чебоксарское вдхр | 1990 | 22 | $\frac{{8.00}}{8}$ | $\frac{{11.36 \pm 0.10}}{{11 - 12}}$ | $\frac{{41.50 \pm 0.18}}{{40 - 43}}$ | $\frac{{19.73 \pm 0.10}}{{19 - 20}}$ | $\frac{{2.41 \pm 0.11}}{{2 - 3}}$ | $\frac{{17.32 \pm 0.12}}{{16 - 18}}$ | $\frac{{39.46 \pm 0.13}}{{38 - 40}}$ | $\frac{{8.66 \pm 0.15}}{{7 - 10}}$ | $\frac{{4.18 \pm 0.17}}{{3 - 6}}$ | $\frac{{6.10 \pm 0.18}}{{5 - 8}}$ | $\frac{{1.94 \pm 0.13}}{{1 - 3}}$ | 56.33 |
| 18 | Саратовское вдхр | 1990 | 22 | $\frac{{8.18 \pm 0.08}}{{8 - 9}}$ | $\frac{{10.84 \pm 0.11}}{{10 - 12}}$ | $\frac{{41.67 \pm 0.16}}{{40 - 43}}$ | $\frac{{19.38 \pm 0.13}}{{19 - 21}}$ | $\frac{{2.76 \pm 0.10}}{{2 - 3}}$ | $\frac{{17.29 \pm 0.10}}{{17 - 18}}$ | $\frac{{39.43 \pm 0.13}}{{39 - 41}}$ | $\frac{{8.43 \pm 0.21}}{{7 - 10}}$ | $\frac{{4.65 \pm 0.18}}{{3 - 6}}$ | $\frac{{6.10 \pm 0.19}}{{4 - 9}}$ | $\frac{{1.88 \pm 0.11}}{{1 - 3}}$ | 52.77 |
| 19 | Волгоградское вдхр | 1985 | 62 | $\frac{{8.10 \pm 0.04}}{{8 - 9}}$ | $\frac{{10.69 \pm 0.06}}{{10 - 12}}$ | $\frac{{39.37 \pm 0.12}}{{37 - 42}}$ | $\frac{{19.02 \pm 0.06}}{{18 - 20}}$ | $\frac{{2.76 \pm 0.05}}{{2 - 3}}$ | $\frac{{16.66 \pm 0.07}}{{15 - 18}}$ | $\frac{{38.44 \pm 0.09}}{{37 - 41}}$ | $\frac{{8.19 \pm 0.12}}{{5 - 10}}$ | $\frac{{4.19 \pm 0.11}}{{2 - 6}}$ | $\frac{{5.70 \pm 0.09}}{{4 - 8}}$ | $\frac{{1.93 \pm 0.09}}{{0 - 4}}$ | 51.56 |
| 20 | Волго-Каспийский р-н, Белинский банк | 1983 | 56 | $\frac{{8.00 \pm 0.04}}{{7 - 9}}$ | $\frac{{10.70 \pm 0.08}}{{9 - 12}}$ | $\frac{{40.07 \pm 0.13}}{{39 - 42}}$ | $\frac{{18.93 \pm 0.07}}{{18 - 20}}$ | $\frac{{2.77 \pm 0.07}}{{2 - 4}}$ | $\frac{{16.59 \pm 0.08}}{{15 - 18}}$ | $\frac{{38.29 \pm 0.09}}{{36 - 40}}$ | – | $\frac{{4.41 \pm 0.10}}{{2 - 6}}$ | $\frac{{5.76 \pm 0.22}}{{4 - 8}}$ | $\frac{{2.21 \pm 0.09}}{{1 - 4}}$ | 46.14 |
| 21 | Гандуринский банк | 1983 | 59 | $\frac{{8.05 \pm 0.05}}{{7 - 9}}$ | $\frac{{10.67 \pm 0.07}}{{9 - 11}}$ | $\frac{{39.81 \pm 0.13}}{{37 - 42}}$ | $\frac{{18.86 \pm 0.06}}{{18 - 20}}$ | $\frac{{2.90 \pm 0.07}}{{2 - 4}}$ | $\frac{{16.49 \pm 0.07}}{{16 - 18}}$ | $\frac{{38.25 \pm 0.07}}{{37 - 39}}$ | – | $\frac{{4.05 \pm 0.11}}{{2 - 6}}$ | $\frac{{5.66 \pm 0.11}}{{4 - 8}}$ | $\frac{{2.09 \pm 0.11}}{{0 - 4}}$ | 45.73 |
| 22 | Волго-Каспийский канал | 1983 | 52 | $\frac{{8.19 \pm 0.06}}{{8 - 9}}$ | $\frac{{10.73 \pm 0.07}}{{10 - 12}}$ | $\frac{{40.22 \pm 0.13}}{{39 - 42}}$ | $\frac{{18.96 \pm 0.07}}{{18 - 20}}$ | $\frac{{2.98 \pm 0.06}}{{2 - 4}}$ | $\frac{{16.35 \pm 0.09}}{{15 - 18}}$ | $\frac{{38.29 \pm 0.10}}{{37 - 40}}$ | – | $\frac{{4.58 \pm 0.07}}{{3 - 7}}$ | $\frac{{5.74 \pm 0.09}}{{4 - 8}}$ | $\frac{{2.15 \pm 0.02}}{{0 - 4}}$ | 45.3 |
| 23 | Низовье р. Кума | 1990 | 24 | – | – | – | $\frac{{18.75 \pm 0.11}}{{18 - 20}}$ | $\frac{{2.96 \pm 0.14}}{{2 - 4}}$ | $\frac{{16.33 \pm 0.10}}{{16 - 17}}$ | $\frac{{38.08 \pm 0.13}}{{37 - 39}}$ | $\frac{{8.33 \pm 0.10}}{{7 - 10}}$ | $\frac{{4.40 \pm 0.10}}{{3 - 6}}$ | $\frac{{5.70 \pm 0.11}}{{4 - 7}}$ | $\frac{{1.90 \pm 0.13}}{{0 - 3}}$ | 44.9 |
| 24 | Р. Уил, Казахстан | 1989 | 8 | $\frac{{8.00}}{8}$ | $\frac{{10.25 \pm 0.25}}{{9 - 11}}$ | $\frac{{40.00 \pm 0.62}}{{38 - 42}}$ | $\frac{{18.50 \pm 0.19}}{{18 - 19}}$ | $\frac{{2.75 \pm 0.16}}{{2 - 3}}$ | $\frac{{16.00 \pm 0.19}}{{15 - 17}}$ | $\frac{{37.25 \pm 0.25}}{{36 - 38}}$ | $\frac{{8.47 \pm 0.19}}{{7 - 10}}$ | $\frac{{4.25 \pm 0.25}}{{3 - 6}}$ | $\frac{{5.44 \pm 0.27}}{{4 - 7}}$ | $\frac{{1.69 \pm 0.12}}{{1 - 2}}$ | 48.91 |
| 25 | Р. Эмба, Казахстан | 1991 | 39 | $\frac{{8.24 \pm 0.08}}{{7 - 9}}$ | $\frac{{10.54 \pm 0.09}}{{10 - 12}}$ | $\frac{{39.63 \pm 0.18}}{{38 - 43}}$ | $\frac{{18.82 \pm 0.08}}{{18 - 20}}$ | $\frac{{2.82 \pm 0.06}}{{2 - 3}}$ | $\frac{{16.44 \pm 0.10}}{{15 - 18}}$ | $\frac{{38.10 \pm 0.12}}{{36 - 40}}$ | $\frac{{8.06 \pm 0.09}}{{6 - 10}}$ | $\frac{{4.50 \pm 0.07}}{{3 - 6}}$ | $\frac{{5.77 \pm 0.09}}{{4 - 8}}$ | $\frac{{1.82 \pm 0.08}}{{0 - 3}}$ | 49.14 |
| 26 | Р. Сагиз, Казахстан | 1989 | 48 | $\frac{{8.02 \pm 0.02}}{{8 - 9}}$ | $\frac{{10.43 \pm 0.07}}{{10 - 11}}$ | $\frac{{39.87 \pm 0.23}}{{36 - 42}}$ | $\frac{{18.44 \pm 0.08}}{{13 - 15}}$ | $\frac{{2.48 \pm 0.08}}{{2 - 4}}$ | $\frac{{16.60 \pm 0.09}}{{15 - 18}}$ | $\frac{{37.50 \pm 0.09}}{{36 - 39}}$ | $\frac{{8.08 \pm 0.09}}{{6 - 10}}$ | $\frac{{4.64 \pm 0.08}}{{3 - 7}}$ | $\frac{{5.74 \pm 0.08}}{{4 - 8}}$ | $\frac{{1.60 \pm 0.06}}{{0 - 3}}$ | 48.2 |
| 27 | Дивичинский лиман, Азербайджан | 1991 | 29 | $\frac{{8.50 \pm 0.09}}{{8 - 9}}$ | $\frac{{10.78 \pm 0.10}}{{10 - 12}}$ | $\frac{{39.45 \pm 0.15}}{{38 - 41}}$ | $\frac{{18.67 \pm 0.09}}{{18 - 19}}$ | $\frac{{2.74 \pm 0.11}}{{2 - 4}}$ | $\frac{{16.33 \pm 0.11}}{{15 - 17}}$ | $\frac{{37.74 \pm 0.11}}{{37 - 39}}$ | $\frac{{8.58 \pm 0.16}}{{6 - 11}}$ | $\frac{{4.39 \pm 0.10}}{{2 - 6}}$ | $\frac{{5.39 \pm 0.09}}{{4 - 7}}$ | $\frac{{1.96 \pm 0.10}}{{0 - 4}}$ | 41.06 |
| 28 | Оз. Камыслыбаш, Казахстан | 1983 | 5 | – | – | – | $\frac{{18.60 \pm 0.24}}{{18 - 19}}$ | $\frac{{2.60 \pm 0.24}}{{2 - 3}}$ | $\frac{{16.60 \pm 0.40}}{{16 - 18}}$ | $\frac{{37.80 \pm 0.58}}{{37 - 40}}$ | $\frac{{8.60 \pm 0.34}}{{7 - 11}}$ | $\frac{{4.20 \pm 0.28}}{{4 - 6}}$ | $\frac{{5.40 \pm 0.22}}{{5 - 6}}$ | $\frac{{1.88 \pm 0.23}}{{1 - 3}}$ | 46.15 |
Примечание. D – число ветвистых лучей в спинном плавнике, A – число ветвистых лучей в анальном плавнике, CPMd – число отверстий преоперкуло-мандибулярного канала на зубной кости, CPMp – число отверстий преоперкуло-мандибулярного канала на предкрышечной кости, CSOf – число отверстий надглазничного канала на лобной кости, CSTp – число отверстий надвисочного канала на теменной кости, ll – число перфорированных чешуй в боковой линии, Va – число грудных позвонков, Vi – число переходных позвонков, Vc – число хвостовых позвонков, Vt – общее число позвонков, L – географическая широта. Над чертой – среднее значение и его ошибка, под чертой – min–max, “–” – данные отсутствуют.
Два последних ветвистых луча в плавниках, опирающиеся на один птеригиофор, считали за один луч. Грудными позвонками мы называем туловищные позвонки, несущие ребра и короткие парапофизы, обычно сочлененные с телом позвонка, в отличие от переходных позвонков, расположенных в задней части туловищного отдела и несущих удлиненные парапофизы, срощенные с телами позвонков. Всего обработано 1083 экз. рыб из 28 локальностей, расположенных в широтном диапазоне от 41.06° до 58.06° с.ш.
Координаты мест лова для рыб, собранных до 2000 г. включительно, определяли при помощи картографического сервиса Google. Проведена статистическая обработка данных, включая расчет средних и ошибок выборочных распределений, корреляционный и регрессионный анализ. Для визуализации СФР использованы двумерные графики.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Статистические характеристики исследованных признаков красноперки представлены в табл. 1. Для признаков, традиционно используемых в биометрических исследованиях карповых рыб (D, A, ll), выборочные средние в целом укладываются в диапазон величин, содержащихся в работах по изменчивости данного вида, если не принимать во внимание вероятные опечатки или явные расхождения в методике подсчета. Например, в публикациях Климчик-Яниковской (Klimczyk-Janikowska, 1970; 1975) число ветвистых лучей в непарных плавниках увеличено на единицу по сравнению с работами других авторов, исследовавших красноперок из того же региона, что, вероятнее всего, указывает на учет двух последних лучей, опирающихся на один птеригиофор, как самостоятельных счетных единиц.
По нашим данным, выборочные средние D колеблются от 8.0 до 8.5, в подавляющем большинстве случаев не отклоняясь вовсе или лишь незначительно отклоняясь от ближайшего целого (8.0). Как отмечено ранее, за небольшими исключениями, это совпадает с данными других авторов по красноперке водоемов Европы. Вместе с тем для красноперок Аральского бассейна ряд авторов (Митрофанов и др., 1987) приводят значения D, близкие к 9.0, что требует проверки. Разброс значений А больше, чем D, и составляет от 10.25 (р. Уил, Казахстан) до 11.36 (Чебоксарское водохранилище), причем тяготения к ближайшему целому в этом случае не наблюдается – выборочные средние равномерно распределяются между целыми значениями 10.0 и 11.0. Отмеченный нами нижний предел А почти совпадает с литературными данными – 10.1 для р. Уил (Митрофанов и др., 1987), однако верхний предел, как и в случае с D, ниже – 11.66 для р. Западная Двина, Белоруссия (Жуков, 1965), 11.7 для оз. Лукнайно, Польша (Szlachciak, Strachowska, 2010), 12.4 для р. Прут, Молдавия (Попа, 1976). При этом, если никаких географических закономерностей в распределении значений D по нашим данным не прослеживается, то для А заметна тенденция к снижению с севера на юг, и особенно на юго-восток. У красноперок бассейна Верхней и Средней Волги средние значения А находятся в диапазоне 10.92–11.36, в бассейне Нижней Волги и в водных объектах Казахстана – 10.25–10.84.
В исследованных нами выборках значения ll колеблются от 39.37 (Волгоградское водохранилище) до 42.03 (оз. Чистое, Ярославская обл.). По литературным данным, этот диапазон несколько шире – от 38.7 для р. Эльба (Chitravadivelu, 1972) до 42.63 для Днепро-Бугского лимана (Гейна, 2016). Важная особенность ll, как и других полимерных признаков, связанных с сегментацией тела рыб, – зависимость от географической широты, обусловленная, по мнению ряда авторов, влиянием температуры (Бабкина и др., 2013; Smirnov et al., 2006; Татарко, 1968). В этой связи необходимо рассмотреть различия исследованных выборок красноперки с учетом широты мест лова (L). Зависимость ll от L показана на рис. 2а. Видно, что ll связано с L тесной положительной корреляцией при линейном характере связи (r = 0.702, b = = 0.115, p < 0.001). Однако в вытянутом облаке точек отчетливо выделяются группы выборок, которые принадлежат к отдельным речным бассейнам, последовательно сменяющим друг друга с севера на юг. Наибольшее ll наблюдается у красноперок бассейнов Верхней и Средней Волги и Балтики, несколько ниже значения этого признака – в выборках из водохранилищ р. Днепр, еще ниже – в бассейне Азовского моря и, наконец, самые низкие значения – в выборках из низовьев р. Волги, водоемов и водотоков Прикаспия и Казахстана. Из более западных локальностей к этой группе крупночешуйных красноперок примыкают только рыбы из оз. Палеостоми и Азовского моря. Значения ll в выборках, взятых почти на одной широте, могут значительно различаться. Так, между красноперками дельты р. Волги и приманычских речек Джурак-Сал и Чекалда различия средних приближаются к одной чешуе. Наиболее резкий перепад наблюдается между выборками из Нижней Волги: на протяжении немногим >100 км значение ll падает с 41.67 в Саратовском водохранилище до 39.36 в верхней части Волгоградского водохранилища, т.е. сразу более, чем на две чешуи.
Рис. 2.
ll (а) и Vt (б) в выборках красноперки в зависимости от географической широты (° с.ш.). 1 – бассейн Балтийского моря, 2 – бассейн Черного моря, 3 – бассейн Азовского моря, 4 – бассейн Верхней и Средней Волги, 5 – бассейн Нижней Волги и Прикаспий, 6 – бассейн р. Сырдарья. Пунктирной линией показана прямая регрессии.
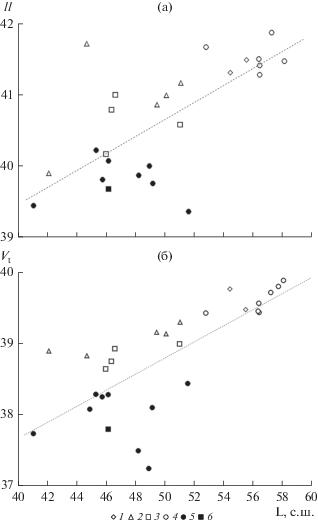
В целом исследованный материал можно разделить по этому признаку на две региональные группы (рис. 1). В одну из них входят выборки из бассейнов Балтийского, Черного и Азовского морей, а также Верхней и Средней Волги, включая выборку из Саратовского водохранилища. Другая группа представлена остальными выборками из Нижней Волги, включая Волго-Каспийский район, Южного Каспия (Дивичинский лиман) и бессточных водоемов Казахстана. Следует отметить, что снижение ll с севера на юг, столь очевидное при рассмотрении всей совокупности выборок, наблюдается только в первой из этих двух групп (выраженное значительно слабее) и совершенно не прослеживается во второй.
Сходная картина наблюдается и при анализе Vt (рис. 2б). При наличии тесной связи Vt и L при рассмотрении всего материала (r = 0.734, b = = 0.110, p < 0.001), у красноперки по этому признаку выделяются те же две четко географически очерченные группы выборок. Как и в случае ll, внутри первой группы связь Vt с L проявляется как результат межгрупповых различий по этому признаку, а у красноперок из низовьев р. Волги, водоемов и водотоков Прикаспия и Казахстана (для краткости будем называть их юго-восточными выборками) такой связи не отмечается. Скачок среднего значения Vt при переходе от Саратовского водохранилища к Волгоградскому также резко выражен и составляет один позвонок. Весьма показательно, что точно такой же паттерн наблюдается при совместном рассмотрении красноперок по признакам ll и Vt (рис. 3а): в группе юго-восточных выборок корреляция между переменными отсутствует, на остальной части региона исследований она опосредована межбассейновыми различиями популяций. При этом юго-восточная и западная группы разделяются в пространстве вышеуказанных признаков без перекрывания.
Рис. 3.
Распределение выборок красноперки по сочетанию ll и Vt (а), Va и Vc (б). Обозначения как на рис. 2.
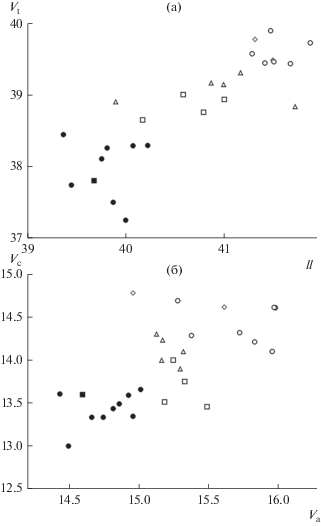
Как показал анализ распределения Vt по отделам позвоночника, хвостовой и туловищный (грудной) отделы вносят сопоставимый вклад в дифференциацию выборок красноперки (рис. 3б). В пространстве признаков Va + i – Vc и Va – Vc (на рисунке дан график для второй из этих пар признаков) отчетливо выделяются те же две совокупности разделяемых по географическому признаку выборок, которые выявлены при последовательном анализе связи Vt и ll с L. Внутри каждой из двух совокупностей не наблюдается значимой связи между двумя признаками, однако при их объединении сильная положительная связь очевидна.
Каких-либо географических или экологических закономерностей в распределении значений числа отверстий сейсмосенсорной системы на костях черепа красноперки выявить не удалось. Размах межпопуляционных различий средних по признакам CPMp, CPMd, CSOf и CSTp составляет около одного отверстия, что сопоставимо с изменчивостью такого полиморфного вида как плотва Rutilus rutilus (L.) (Касьянов, Изюмов, 1990). Во многих случаях различия между выборками красноперки (независимо от их географической близости) по признакам этой группы статистически значимы. Однако высокие и низкие значения числа отверстий чередуются в ареале хаотически и не связаны с какими-либо определенными типами водных объектов (табл. 1).
У красноперки нами выявлено большое разнообразие вариантов формулы глоточных зубов (D. ph1 + D. ph2), значительно выходящее за пределы, описанныe в литературе: 3.55.3 (“норма”), 2.55.3, 3.55.2, 3.54.3, 2.54.2, 2.54.3, 3.45.3, 2.45.3, 2.44.2, 4.55.3, 4.54.3, 1.55.2, 3.65.3 и др. Большое число билатеральных комбинаций означает редкость большинства из них и, следовательно, невозможность сравнительного изучения частот при умеренных объемах выборок. D. ph1, напротив, демонстрирует весьма ограниченную внутри- и межпопуляционную изменчивость, однако в картине варьирования этого признака прослеживается система. Возможные варианты D. ph1 на левой глоточной кости – 4, 5 и 6, на правой – 4 и 5. При этом, как и у других видов подсемейства ельцовых, имеет место направленная асимметрия признака: в случае его асимметричного проявления значение слева, как правило, больше, чем справа. Следует отметить, что частота асимметричных вариантов неодинакова в западной и юго-восточной группах популяций красноперки. В первой из них таковые либо отсутствуют вовсе (в 9 из 18 выборок), либо попадаются единично, причем преобладание левосторонних вариантов (5–4) над правосторонними (4–5) выражено слабо. Из 18 выборок западной группы оказалось всего шесть с преимущественно левосторонней асимметрией по числу глоточных зубов, тогда как в юго-восточной группе таких выборок было восемь из 10 (относительные частоты для сравнения по критерию “фи”, соответственно, f1 = 0.333 и f2 = 0.8, φ = 2.493, p < 0.01). В некоторых выборках юго-востока сочетание “5–4” уже нельзя назвать редким, например, в выборке из р. Сагиз частота этого варианта достигала 17%, в р. Эмба – 11%.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные данные указывают на то, что основной вклад в географическую подразделенность вида S. erythrophthalmus вносят меристические признаки, связанные с сегментацией тела – Vt, распределение позвонков по отделам позвоночника, ll и А. Следует отметить, что наши первоначальные представления об относительном вкладе генетического своеобразия популяций и температурного фактора (как функции географической широты) в клинальную изменчивость пресноводных рыб умеренной зоны требуют в случае красноперки существенной ревизии. До сих пор влияние температуры, при которой происходит развитие рыб, на формирование широтных клин мы считали главенствующим (Кожара и др., 1996). Предполагалось, что на базовую температурную зависимость числа позвонков могут накладываться генетически и эволюционно обусловленные особенности отдельных популяций и внутривидовых форм (статус которых в дальнейшем может быть пересмотрен). Об этой специфике мы судили по отклонениям значений признака в таких выборках от общевидовой регрессионной зависимости “признак–широта”. Ситуация с ll и Vt у красноперки демонстрирует принципиальную возможность широтных клин, имеющих преимущественно ступенчатую структуру. Корреляция меристических признаков с широтой в этом случае указывает не столько на средовую детерминацию признака, сколько на значительную популяционно-генетическую гетерогенность исследуемого материала, вплоть до отнесения географически удаленных популяций к разным таксонам.
Другой существенный вывод, к которому приводят полученные результаты, – чисто географический, викариантный характер формообразования у красноперок в рассматриваемой части ареала, не связанный с четко выраженными экологическими факторами. В каждой из двух популяционных систем, выделенных по географическому принципу, представлены выборки из водных объектов, различных в экологическом отношении. Это озера, различающиеся по размеру, генезису, гидрологическим и гидрохимическим характеристикам, водохранилища, дельтово-эстуарные системы и бессточные водоемы аридной зоны. Тем не менее, в пространстве признаков близко соседствуют выборки из разнотипных водоемов, объединенные принадлежностью к тому или иному речному бассейну. Таковы, например, выборки из Рыбинского и Чебоксарского водохранилищ, ледникового оз. Селигер, карстовых озер Юхор и Санхар и пойменного оз. Чистое в бассейне Верхней и Средней Волги. Аналогичным образом, единый фенон образуют красноперки из Волгоградского водохранилища, дельты р. Волги, низовьев р. Кума, Дивичинского лимана Каспийского моря и бессточных осолоняющихся речек Казахстана, ранее принадлежавших бассейну Каспия.
Особенный интерес представляет отсутствие видимых закономерностей в распределении значений признаков сейсмосенсорной системы в исследованных нами выборках. Данная группа признаков подвержена существенной изменчивости у тех видов ельцовых, которые многочисленны в дельтово-эстуарных системах внутренних морей и не избегают солоноватых вод: плотвы Rutilus rutilus (L.), леща Abramis brama (L.), синца Ballerus ballerus (L.), густеры Blicca bjoerkna (L.) (Мироновский, 1988, 1991; Kozhara, 1997, 2002). Направленность изменений при таком формообразовании таксоноспецифична: у видов трибы Abramidini в условиях повышенной минерализации происходит закономерное снижение числа отверстий сейсмосенсорной системы, тогда как у полупроходной плотвы Понто-Каспия оно, напротив, увеличено по сравнению с чисто пресноводными формами. Беспорядочный характер распределения значений этих признаков в ареале Scardinius erythrophthalmus указывает, что наряду с генетическими особенностями популяций их изменчивость детерминирована какими-то пока неизвестными (неконтролируемыми) факторами локального значения. Хотя этот вид в изобилии встречается в дельтах рек и лиманах Понто-Каспия, он придерживается их пресноводных участков и в солоноватой воде редок (Танасийчук, 1957). По-видимому, значение фактора минерализации для формообразования в роде Scardinius относительно невелико. Возможно, закономерности или тенденции такого рода могут быть выявлены при более детальном исследовании, учитывающем гидрологические и гидрохимические характеристики мест обитания красноперки.
Полученные данные о структуре фенетического разнообразия красноперки в исследованной части ареала не согласуются с представлением о ней как о мономорфном виде. По ll, Vt, соотношению Va и Vc, а также отчасти по А на исследованном материале выделяются два обособленных фенона, соответствующие географически четко разграниченным популяционным системам. В зависимости от используемой терминологии и критериев выделения, такие системы могут считаться географическими расами, подвидами или даже видами, причем последний выбор в настоящее время делается исследователями все чаще.
В частности, у карповых рыб в последние два десятилетия видовой ранг присвоен значительному числу ранее описанных внутривидовых форм. Так, в самостоятельные виды выделены азово-черноморская тарань Rutilus heckelii (Nordmann, 1840) и каспийская вобла R. caspicus (Yakovlev, 1870) (Kottelat, Freyhof, 2007), которые до недавнего времени рассматривались как подвиды обыкновенной плотвы; видовой статус присвоен сразу нескольким подвидам быстрянки Alburnoides bipunctatus (Bloch, 1792) (Bogutskaya, Coad, 2009). Таких примеров довольно много, они известны и для рода Scardinius. Упоминавшаяся выше анталийская красноперка S. elmaliensis (Bogutskaya, 1997) первоначально описана как подвид S. erythrophthalmus elmaliensis на основании, главным образом, более высокого числа жаберных тычинок: 15–20 против 9–12 у обыкновенной красноперки (Bogutskaya, 1997; Fricke et al., 2007). Подвид S. erythrophthalmus scardafa Karaman, 1928 из оз. Скадар и Охрид на Балканском полуострове, отличающийся от типичной формы меньшими A и ll, недавно переописан как новый вид S. knezevici (Bianco, Kottelat, 2005). Аналогичные ревизии выполнены и в отношении ряда других форм рода Scardinius из южной Европы.
Из меристических признаков, используемых в систематике красноперок, наибольшее значение имеют sp.br, А и ll (Kottelat, Freyhof, 2007). Так, сестринские викариирующие виды S. erythrophthalmus и S. elmaliensis в Турции различаются по ll приблизительно на одну чешую и по A на один луч (Ilhan, 2019); такие же различия отмечены в другой паре сестринских видов из озер Греции, S. acarnanicus (Stephanidis) и S. graecus (Stephanidis) (Iliadou et al., 1996). Без сомнения, к таксономически важным признакам в данном случае следует отнести и число позвонков, которое в публикациях приводится значительно реже, чем наружные счетные признаки, и зачастую вызывает вопросы, связанные с методикой подсчета. Различия по Vt у двух вышеупомянутых видов из Греции достигают почти двух позвонков (Iliadou et al., 1996). На этом фоне различия в две чешуи и более, чем в два позвонка, отмечаемые нами между красноперками западной и юго-восточной групп популяций, вполне соответствуют видовому уровню. Наконец, в свете приведенных выше данных по изменчивости формулы глоточных зубов у красноперок юго-восточной группы, представляет интерес указание Митрофанова и др. (1987), что у 23% особей из р. Чу отмечены наборы глоточных зубов “3.5–4.3”.
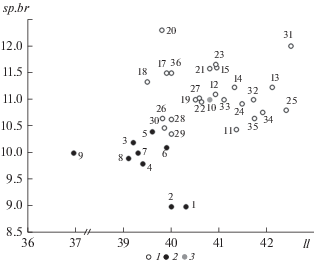
У нас имеются только литературные данные по sp.br у красноперок. Для наглядности мы прибегли к двумерному представлению. На рис. 4 показано распределение выборок S. erythrophthalmus из водоемов центральной и восточной Европы, Закавказья, Казахстана, Малой и Средней Азии по средневыборочным значениям ll и sp.br, взятых из доступных публикаций. Условные обозначения на этом рисунке отражают отнесение выборок на основе предыдущего анализа к западной или же к юго-восточной группе. Отдельно показана выборка из р. Урал, которую по имеющимся данным пока нельзя уверенно отнести к одной из совокупностей, поскольку эта река занимает пограничное положение между их ареалами и при этом не представлена в наших сборах.
Рис. 4.
Распределение выборок красноперки по сочетанию ll и sp.br согласно литературным данным: 1 – западная группа, 2 – юго-восточная группа, 3 – р. Урал. Нумерация выборок: 1 – р. Амударья (Никольский, 1940); 2 – р. Чу, 1933 г. (Дрягин, 1936); 3 – то же, 1958 г. (Митрофанов и др., 1987); 4 – то же, 1959 г. (Пивнев, 1963); 5 – то же, 1985 г. (Митрофанов и др., 1987); 6 – р. Сарысу (Ерещенко, 1956); 7 – Фархадское водохранилище, р. Сырдарья (Максунов, 1956); 8 – р. Уил (Митрофанов и др., 1987); 9 – каспийское побережье Ирана (http://www.briancoad.com/Species%20Accounts/Cyprinidae%20Garra%20to%20Vimba.htm#Scardinius); 10 – р. Урал (Шапошникова, 1964); 11 – Воткинское водохранилище, 1965–1971 (Устюгова, 1978); 12 – то же, 2018 (Ложкина, 2019); 13 – Камское водохранилище (Устюгова и др., 2008); 14 – бассейн р. Вычегда (Бознак, 2008); 15 – бассейн р. Луза (Бознак, 2008); 16 – бассейн р. Вага (Соловкина, 1969); 17 – оз. Палеостоми (Эланидзе, 1983); 18 – водохранилище Бююкчекмедже, Турция (Özuluğ, 1999); 19 – водоемы СЗ Турции (Ilhan, 2019); 20 – оз. Вольви, Греция (Economidis, Wheeler, 1989); 21 – оз. Корония, Греция (Iliadou et al., 1996); 22 – р. Камчия, Болгария (Манолов, Сивков, 1975); 23 – оз. Шабла, Болгария (Манолов, Сивков, 1975); 24 – р. Днестр, верхнее течение (Опалатенко, 1970); 25 – бассейн р. Прут (Попа, 1976); 26 – дельта р. Дунай (Мовчан, Смiрнов, 1981); 27 – р. Днепр, верхнее течение (Жуков, 1965); 28 – р. Днепр, среднее течение (Мовчан, Смiрнов, 1981); 29 – р. Южный Буг (Мовчан, Смiрнов, 1981); 30 – р. Северский Донец (Мовчан, Смiрнов, 1981); 31 – Онежское оз. (Покровский, 1935); 32 – р. Западная Двина (Жуков, 1965); 33 – р. Неман (Жуков, 1965); 34 – водохранилище Пшечице, бассейн верхнего течения р. Висла, Польша (Klimczyk-Janikowska, 1970); 35 – водохранилище Хехло, бассейн верхнего течения р. Висла, Польша (Klimczyk-Janikowska, 1970); 36 – бассейн р. Дунай, Венгрия (Berinkey, 1960). Пояснения в тексте.
В тех немногих случаях, когда средние арифметические не были приведены в публикациях, вместо них использовали полусумму пределов колебаний признака. Из массива доступных данных исключены значения sp.br, резко отклоняющиеся от региональной и общевидовой нормы. Такие величины указываются для трех озер Эстонии (средние 17.49–17.94) (Хаберман и др., 1974), водосбросного канала теплоэлектростанции “Поможаны” в Щецине, Польша (10–17, среднее 14.56 ± 0.14) (Krzykawski et al., 1997), оз. Шильян в Азербайджане (12–19, среднее 15) (Абдурахманов, 1962) и оз. Челкар в Казахстане (13–14) (Серов, 1956). Столь резкие отличия от данных большинства авторов могут объясняться использованием иной методики подсчета, ошибками при записи и обработке данных, подготовке публикаций и, наконец, опечатками. Например, в случае с озерами Эстонии завышенные значения sp.br, по-видимому, связаны с подсчетом тычинок не на внешней, а на внутренней стороне первой жаберной дуги (Мовчан, Смiрнов, 1981).
Остальные данные (рис. 4) свидетельствуют о выраженных различиях между западной и юго-восточной группами по данному признаку и о высокой гомогенности его значений в ареалах каждой из групп, где представлены разного типа и генезиса водные объекты, находящиеся в различных природных зонах – от европейской тайги до средиземноморских субтропиков. У красноперок Европы, западного Закавказья и Малой Азии пределы варьирования sp.br составляют менее двух единиц (10.36–12.3), в выборках из Казахстана и Средней Азии размах колебаний еще меньше (9–10.4) при очень незначительном перекрывании с предыдущей группой. При этом у многотычинковых красноперок наблюдаются и более высокие значения ll, что создает искусственную корреляцию между двумя признаками (r = 0.418, p = 0.011), хотя по отдельности ни в одной из групп такой корреляции не наблюдается.
Эти результаты подтверждают мнение Л.С. Берга (1949), что красноперки Аральского бассейна заслуживают выделения в особое племя (natio), основанное, прежде всего, на меньшем sp.br и ll (Дрягин, 1936; Никольский, 1940). Взгляд Л.С. Берга поддержали Ф.А. Турдаков и К.В. Пискарев (1955), отметившие выраженное отличие красноперок из бассейна Арала, включая ныне бессточную р. Чу, (8–10 тычинок), от типичной формы (11–12 тычинок). В противоположность этому, В.И. Ерещенко (1956), ссылаясь (без указания источников) на некие новые литературные данные по морфометрии красноперок Аральского бассейна, счел выявленные различия недостаточными для выделения этих популяций в особую внутривидовую форму.
Действительно, накопление литературных данных по изменчивости конкретного вида неизбежно сопровождается аккумуляцией разного рода ошибок и неточностей, обусловленных “человеческим фактором”, в том числе, индивидуальностью операторов и погрешностями при подготовке публикаций. По сути, речь идет о дополнительном источнике вариации, привносящем шум в исходные данные и размывающем “истинную” картину СФР. Рис. 4 также дает основания для таких предположений. Так, обращает на себя внимание весьма компактная и при этом обособленная от остальных группа выборок из дельты р. Дунай, среднего участка р. Днепр, рек Южный Буг и Северский Донец (№ 26, 28–30 на рис. 4) (Мовчан, Смiрнов, 1981). В этой группе значения sp.br заметно ниже, чем в других выборках из водоемов черноморского бассейна. То же касается и ll в этих выборках, которое также одно из самых низких в пуле литературных данных по западной группе и почти на единицу ниже значений, полученных нами для днепровских водохранилищ (табл. 1). Тем не менее, средневыборочные значения sp.br для красноперок бассейнов Арала и ближнего Каспия11 явно ниже, чем в выборках, взятых к северу и западу. Этот результат интересен и сам по себе, но с учетом представленных выше наших данных по ll и признакам осевого скелета, можно заключить, что между юго-восточной и западной группами популяций красноперки существуют выраженные различия, создающие основу для систематической ревизии вида S. erythrophthalmus в нынешнем объеме.
По-видимому, популяции красноперки Нижней Волги и всех остальных рек, озер и лиманов бассейна Каспия, а также бессточных рек и озер Казахстана и Средней Азии (рис. 1) следует относить к самостоятельному виду, отличному от S. erythrophthalmus (статус популяций Аральского бассейна требует дополнительного анализа). Этот вид характеризуется наиболее частыми значениями ll 39–40 (против 41–42 у обыкновенной красноперки), Vt 37–38 (против 39–40) и sp.br 9–10 (против 10–12). Более подробное описание этого вида и оценка его места в системе рода Scardinius – предмет отдельного исследования.
Выводы. Анализ СФР обыкновенной красноперки выявил две географически обособленные группы популяций, заслуживающие видового ранга: ареал западной группы включает бассейн р. Волги (кроме низовьев) и бассейны, расположенные западнее, ареал юго-восточной группы – всю остальную часть бассейна Каспия, бессточные бассейны западного Казахстана и бассейн Арала. Наибольший вклад в дифференциацию этих групп вносят признаки осевого скелета, а также ll, sp.br и A, играющие важную роль в систематике рода Scardinius. Морфологическая специфика популяций красноперок почти целиком определяется географическим положением мест обитания, включая L; связь с типом водных объектов и их гидрологическими и гидрохимическими характеристиками не выявлена.
Список литературы
Абдурахманов Ю.А. 1962. Рыбы пресных вод Азербайджана. Баку: Изд-во АН АзССР.
Атлас пресноводных рыб России. 2003. Москва: Наука. Т. 1.
Бабкина И.Б., Петлина А.П., Шестакова А.С. 2013. Морфо-экологические особенности уклейки (Alburnus alburnus (L.)) нижней Томи // Вестник Томского государственного педагогического университета. № 8(136). С. 61.
Берг Л.С. 1949. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР. Т. 2. С. 469.
Бознак Э.И. 2008. Красноперка Scardinius erythrophthalmus (L.) притоков реки Северная Двина // Вопр. ихтиол. Т. 48. № 3. С. 427.
Гейна К.М. 2016. Морфологiчнi особливостi вiкова структура та темп росту краснопiрки (Scardinius erythrophthalmus L., 1758) Днiпровсько-Бузькоï гирловоï системи // Рибогосподарська наука Украïни № 4(38). С. 64. https://doi.org/10.15407/fsu2016.04.064
Дворянкин Г.А. 2014. Первая находка красноперки Scardinius erythrophthalmus (Cyprinidae, Cypriniformes) в бассейне реки Онега // Вопр. ихтиол. Т. 54. № 1. С. 118. https://doi.org/10.7868/S0042875213060027
Ерещенко В.И. 1956. Ихтиофауна бассейна реки Сары-Су // Cборник работ по ихтиологии и гидробиологии. Алма-Ата: АН КазССР. № 1. С. 94.
Дрягин П.А. 1936. Рыбы реки Чу и рыбохозяйственное использование этой реки // Рыбное хозяйство КиргССР. Труды Киргизской комплексной экспедиции 1932–1933 гг. Москва: Изд-во АН СССР. Т. 3. № 1. С. 49.
Жуков П.И. 1965. Рыбы Белоруссии. Минск: Наука и техника.
Касьянов А.Н., Изюмов Ю.Г. 1990. Изменчивость числа отверстий в сейсмосенсорных каналах черепа у плотвы Rutilus rutilus // Вопр. ихтиол. Т. 30. № 1. С. 13.
Кожара А.В., Изюмов Ю.Г., Касьянов А.Н. 1996. Общая и географическая изменчивость числа позвонков у некоторых пресноводных рыб // Вопр. ихтиол. Т. 36. № 2. С. 179.
Ложкина Е.Н. 2019. Морфобиологическая характеристика красноперки Sсardinius еrythrophthalmus Воткинского водохранилища // Материалы XI Всероссийского конгресса молодых ученых-биологов с международным участием. Пермь: Пермский государственный национальный исследовательский университет. С. 185.
Максунов В.А. 1956. К морфологии и биологии красноперки Фархадского водохранилища // Доклады АН ТаджССР. № 15. С. 63.
Манолов Ж., Сивков Я. 1975. Биометрични изследования върху червеноперката Sсardinius еrythrophthalmus (L.) от р. Камчия и Шабленското езеро // Известия на Народния музей – Варна кн. ХI/ХХVI. С. 180.
Митрофанов В.П., Дукравец Г.М., Сидорова А.Ф. и др. 1987. Рыбы Казахстана. Карповые. Т. 2. Алма-Ата: Наука Казахской ССР.
Мироновский А.Н. 1988. О географической изменчивости карповых рыб: особенности микроэволюции популяций дельтово-эстуарных участков // Зоол. журн. Т. 67. № 7. С. 1013.
Мироновский А.Н. 1991. Особенности изменчивости популяционной структуры некоторых карповых рыб Волго-Каспийского и сопредельных районов. 2. Анализ изменчивости признаков // Вопр. ихтиол. Т. 31. № 5. С. 734.
Мовчан Ю.В., Смiрнов А.I. 1981. Короповi // Фауна Украïни. Т. 8. Риби. Вип. 2. Ч. 1. Київ: Наукова думка.
Никольский Г.В. 1940. Рыбы Аральского моря. Москва: Красный пролетарий.
Опалатенко Л.К. 1970. Рыбы-лимнофилы бассейна Верхнего Днестра // Вестник зоологии № 1. С. 34.
Пивнев И.А. 1963. Материалы по морфологии и биологии чуйской красноперки // Ихтиологический сборник. Фрунзе: Киргизский гос. ун-т. № 1. С. 61.
Покровский В.В. 1935. О красноперке (Scardinius erythrophthalmus L.) из Онежского озера // Труды Карельской научно-исследовательской рыбохозяйственной станции. Ленинград. Т. 1. С. 391.
Серов Н.П. 1956. Рыбы озера Челкар // Сборник работ по ихтиологии и гидробиологии. Алма-Ата: АН КазССР. № 1. С. 278.
Попа Л.Л. 1976. Рыбы бассейна р. Прут. Кишинев: Штиинца.
Соловкина Л.Н. 1969. О находках серебряного карася Саrassius auratus gibelio (Bloch) и красноперки (Scardinius erythrophthalmus L.) на европейском северо-востоке СССР // Вопр. ихтиол. Т. 9. № 5(58). С. 945.
Танасийчук В.С. 1957. Закономерности формирования численности некоторых каспийских рыб//Труды КаспНИРО. Т. 13. С. 3.
Татарко К.И. 1968. Влияние температуры на меристические признаки рыб // Вопр. ихтиол. Т. 8. № 3(50). С. 425.
Турдаков Ф.А., Пискарев К.В. 1955. Материалы по систематике и биологии чуйской красноперки // Труды Института зоологии и паразитологии АН КиргССР. Фрунзе: АН КиргССР. № 3. С. 79.
Шапошникова Г.Х. 1964. Биология и распределение рыб в реках уральского типа. Москва: Наука.
Устюгова Т.В. 1978. Красноперка Воткинского водохранилища // Основы рационального использования рыбных ресурсов камских водохранилищ. Пермь: Пермский государственный университет. С. 52.
Устюгова Т.В., Красулина О.С., Зиновьев Е.А. 2008. Красноперка Камского водохранилища // Биология и экология рыб Прикамья: межвузовский сборник научных трудов. Пермь: Пермский государственный университет. № 2. С. 129.
Хаберман Х., Нооритс Т., Паавер Т. 1974. О морфологии красноперки в некоторых озерах Эстонии // Гидробиол. исслед. Т. 6. С. 134.
Эланидзе Р.Ф. 1983. Ихтиофауна рек и озер Грузии. Тбилиси: Мецниереба.
Яковлев В.Н., Изюмов Ю.Г. Касьянов А.Н. 1981. Фенетический метод исследования популяций карповых рыб // Биол. науки. № 2. С. 98.
Berinkey L. 1960. Further Morphological and Osteological Investigations on the Hybrids of Hungarian Cyprinids // Annales historico-naturales Musei nationalis hungarici. V. 52. Part Zoologica. P. 447.
Bianco P.G., Kottelat M. 2005. Scardinius knezevici, a new species of rudd from Lake Skadar, Montenegro (Teleostei: Cyprinidae) // Ichthyol. Explor. Freshwater. № 16(3). P. 231.
Bogutskaya N.G., Coad B.W. 2009. A review of vertebral and fin-ray counts in the genus Alburnoides (Teleostei: Cyprinidae) with a description of six new species // Zoosystematica Rossica. № 18(1). P. 126.
Bogutskaya N.G. 1997. Contribution to the knowledge of leuciscine fishes of Asia Minor. 2. An annotated check-list of leuciscine fishes (Leuciscinae, Cyprinidae) of Turkey with descriptions of a new species and two new subspecies // Mitt. Hamb. Zool. Mus. Inst. № 94. P. 161.
Chitravadivelu K. 1972. On the systematics of the rudd, Scardinius erythrophthalmus (Linnaeus, 1758) (Osteichthyes: Cyprinidae) from Bohemia // Věstník Československé společnosti zoologické. V. 36. № 3. P. 169.
Economidis P.S., Wheeler A. 1989. Hybrids of Abramis brama with Scardinius erythrophthalmus and Rutilus rutilus from Lake Volvi, Macedonia, Greece // J. Fish Biol. V. 35. № 2. P. 295.
Fricke R., Bilecenoglu M., Sari H.M. 2007. Annotated checklist of fish and lamprey species (Gnathostomata and Petromyzontomorphi) of Turkey, including a Red List of threatened and declining species // Stuttgarter Beitr. Naturk. Ser. A. № 706. P. 1.
Ilhan A. 2019. Morphometric comparison between Scardinius erythrophthalmus (Linnaeus, 1758) and Scardinius elmaliensis (Bogutskaya, 1997) // Fresenius Environ. Bull. V. 28. № 5. P. 3824.
Iliadou K., Sourdis J., Economidis P.S., Wheeler A. 1996. Morphological differentiation among species genus Scardinius (Pisces: Cyprinidae) in Greece // J. Natural History. V. 30. P. 459.
Klimczyk-Janikowska M. 1970. Die Rotfeder (Scardinius erythrophthalmus L.) aus den Staubecken Przeczyce und Chechło // Acta Hydrobiol. V. 12. P. 263.
Klimczyk-Janikowska M. 1975. Biometric characteristic and food of the rood (Scardinius erythrophthalmus L.) from the reservoir at Goczałkowice // Acta Hydrobiol. V. 17. № 1. P. 71.
Kottelat M., Freyhof J. 2007. Handbook of European freshwater fishes. Berlin: Kottelat, Cornol and Freyhof.
Kozhara A.V. 1997. Regular phenotypic changes accompanying osmotic adaptations in some cyprinids: micro vs macroevolution // Zhurn. Obshch. Biol. V. 58. № 3. P. 17.
Kozhara A.V. 2002. Patterns of intraspecific variability in carp fishes of the dace subfamily: ecological factors and pathways of phenotypic transformation // Zhurn. Obshch. Biol. V. 63. № 5. P. 393.
Krzykawski S., Więcaszek B., Modrzewski P. 1997. Charac-terystyka cech biometrycznych wzdręgi Scardinius erythrophthalmus (L., 1758) z kanału zrzutowego wód pochłodniczych Electrowni “Pomorzany” w Szczecinie // Systematyka i filogeneza świata zwierzęcego. Systematyka ryb karpiowatych Cyprinidae. III Sesja Sympozjum Olsztyn-Kortowo 8 kwietnia 1997. P. 117.
Mavrin A.S., Kasyanov A.N., Goroshkova T.V. 2017. Morphological characteristics of the rudd Scardinius erythrophthalmus (Linnaeus) from Small Waterbodies of the Upper Volga and Oka Basins // Inl. Wat. Biol. № 10. P. 436. https://doi.org/10.1134/S1995082917040083
Özuluğ M. 1999. A Taxonomic Study on the Fish in the Basin of Büyükçekmece Dam Lake // Turk. J. Zoology. V. 23. № 4. P. 439.
Smirnov S.V., Dzerzhinskii K.F., Levin B.A. 2006. On the relationship between scale number in the lateral line in the african barbel Barbus intermedius and the rate of ontogeny (by experimental data) // J. Ichthyol. V. 46. P. 129. https://doi.org/10.1134/S0032945206010176
Szlachciak J., Strakowska E. 2010. Morphological characteristics and variation of rudd Scardinius erythrophthalmus (L.) from the Łuknajno Lake, Poland // AACL Bioflux. V. 3. № 2. P. 91.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод