

Биология внутренних вод, 2021, № 4, стр. 367-377
Эффективность переноса биологически ценных веществ от фитопланктона к планктонным ракообразным в мезотрофном оз. Обстерно (Беларусь)
Ж. Ф. Бусева a, *, М. И. Гладышев b, c, Н. Н. Сущик b, c, Е. А. Сысова a, К. В. Мягкова a, Ш. Б. Газерани Фарахани a, И. Ю. Фенева d, Е. Г. Сахарова e, В. П. Семенченко a
a Государственное научно-производственное объединение “Научно-практический центр
Национальной академии наук Беларуси по биоресурсам”
Минск, Республика Беларусь
b Федеральный исследовательский центр Красноярский научный центр Сибирского отделения
Российской академии наук
Красноярск, Россия
c Сибирский федеральный университет
Красноярск, Россия
d Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
e Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
Ярославская обл., Некоузский р-н, пос. Борок, Россия
* E-mail: buseva_j@mail.ru
Поступила в редакцию 27.01.2020
После доработки 26.03.2020
Принята к публикации 15.05.2020
Аннотация
Получена оценка эффективности переноса биологически ценных веществ (углерода, фосфора, азота и жирных кислот, включая полиненасыщенные жирные кислоты) от фитопланктона к зоопланктону в трех биотопах (пелагиали, заросшей литорали и литорали без зарослей) мезотрофного оз. Обстерно (Беларусь). Эффективность переноса веществ выражена как отношение их вторичной продукции к первичной в единице объема и в единице биомассы. Эффективность, выраженная на единицу объема воды, характеризует эффективность переноса веществ в водоеме, эффективность, выраженная на единицу биомассы, – способность зоопланктона аккумулировать биологически ценные вещества, поступающие из пищевых ресурсов в биомассе, т.е. характеризует качество зоопланктона как ресурса для вышестоящих трофических уровней. Эффективность передачи полиненасыщенных жирных кислот была ниже, чем эффективность передачи углерода. Напротив, аккумуляция биогенных элементов, особенно фосфора была более эффективной по сравнению с углеродом. Это свидетельствует о зоопланктоне как резервуаре, повышающем содержание биогенных элементов, и важном поставщике биогенных элементов в трофической цепи. Планктонные сообщества пелагиали мезотрофного озера оказались более эффективными в переносе веществ от первичных продуцентов к консументам, чем литоральные сообщества.
ВВЕДЕНИЕ
Функционирование высших трофических уровней и перенос вещества и энергии в водных экосистемах зависит от эффективности переноса биологически ценных веществ от фитопланктона к зоопланктону. Один из важнейших компонетов в этом потоке – углерод, выступающий главным строительным материалом консументов всех уровней. Как правило, в природе, в условиях умеренного климата, углерод в пищевых ресурсах зоопланктона находится в избытке и не выступает лимитирующим фактором для планктонных ракообразных в безледный период (Sterner, Schulz, 1998), однако ЖК, в том числе ПНЖК, азот и фосфор, содержатся в фитопланктоне в недостаточном количестве (White, 1993). Поэтому взаимодействия в системе “фитопланктон‒зоопланктон” могут зависеть от химических составляющих фитопланктона, а обилие видов ракообразных – или от содержания семейства омега-3 (Wacker, Von Elert, 2001; Becker, Boersma, 2005; Gladyshev et al., 2006, 2007), или от стехиометрического соотношения C : N : P в биомассе фитопланктона (DeMott, 1998; DeMott et al., 1998; Sterner, Schulz, 1998; Boersma, et al., 2001). Разные химические элементы и биохимические вещества отвечают в живых организмах за разные физиологические функции. Если углерод в основном выполняет строительную функцию и поставляет энергию организму в результате метаболических процессов, то ПНЖК, фосфор и азот требуются для регуляции основных физиологических процессов. ПНЖК в составе фосфолипидов используется для построения клеточных мембран (Feller, 2008; Wassell, Stillwell, 2008), участвуют в регуляции работы сердечно-сосудистой системы, свертывания крови, иммунных реакций, работы репродуктивных органов (Schmitz, Ecker, 2008). При недостатке жизненно важных ЖК на уровне популяций отмечается снижение скорости роста и увеличение смертности у планктонных животных и личинок рыб (Glencross, 2009). Недостаток ключевых питательных элементов (углерода, азота и фосфора) влияет на синтез основных макромолекул – липидов, протеинов и нуклеиновых кислот (Wagner et al., 2015). При недостатке фосфора происходит замедление образования рибосомных РНК (Loladze, Elser, 2011).
Все указанные биологически ценные вещества растительноядные ракообразные получают от фитопланктона. Разные виды фитопланктона обладают разной ценностью по содержанию в них ПНЖК, азота и фосфора. Так, диатомовые водоросли и динофлагелляты содержат больше ПНЖК, чем зеленые водоросли (Müller-Navarra, 1995). Содержание ПНЖК в цианобактериях очень низкое (Gulati, DeMott, 1997), но они богаты фосфором. Следовательно, от видового состава водорослей будет зависеть количество поступающих биологически ценных веществ по трофической цепи. Кроме того, соотношения C : N : P в фитопланктоне подвержены изменениям в зависимости от солнечной радиации, содержания биогенных веществ в воде и других факторов (Dickman et al., 2008; Elser et al., 2002). Можно предположить, что эффективность передачи биологически ценных веществ зависит от видовой структуры фитопланктона и зоопланктона, а также типа биотопа (литораль или пелагиаль).
Цель работы – исследовать эффективность переноса биологически ценных веществ (углерода, фосфора, азота, ЖК, включая ПНЖК) от фитопланктона к зоопланктону в заросшей литорали, литорали без зарослей и пелагиали в мезотрофном озере в период массового развитии молоди рыб возраста 0+ в литорали.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Полевые исследования проводили в мезо-эвтрофном оз. Обстерно на северо-западе Беларуси. Озеро Обстерно – водоем ледникового происхождения, площадь водной поверхности 9.89 км2, максимальная глубина 12 м, средняя – 5.1 м. Это относительно мелководное озеро с высокой прозрачностью, достигающей 6.5 м в мае–начале лета, и широким (до 50–100 м) поясом макрофитов, опоясывающим озеро по всему периметру (Blue book…, 1994). Видовой состав молоди рыб, обитающих в литорали, представлен в основном сем. Cyprinidae (карповые), молодь в массе развивается на мелководье в июне–июле и питается в это время зоопланктоном (Buseva, Pljuta, 2015). Пробы воды для элементного анализа (углерода, азота, фосфора) в зоопланктоне и фитопланктоне отбирали в конце июня из трех биотопов: в пелагиали на глубине 5 м, в открытой литорали без зарослей и в литорали с зарослями камыша озерного (Scirpus lacustris L.) на глубине 0.7–1.2 м. Пробы для биохимического анализа на содержание общих ЖК и ПНЖК взяты только из двух биотопов литорали, что связано с техническими причинами. Для отбора проб фитопланктона использовали батометр Паталаса объемом 1 л. Воду в литорали отбирали с подповерхностного горизонта с глубины 0.3–0.5 м. В пелагиали формировали интегральную пробу из нескольких проб, отобранных со следующих горизонтов: максимальная глубина, глубина прозрачности, половина глубины прозрачности (на глубине 1 м и 0.3 м). Объем каждой интегрированной пробы был 0.5 л. Пробы фиксировали 2%-ным формалином. Зоопланктон собирали планктонной сетью Джеди (с размером ячеи 100 мкм), протягивая ее от дна до поверхности, и фиксировали 4%-ным формалином. На всех станциях пробы отбирали в трех повторностях.
Фитопланктон концентрировали с помощью осадочного метода. Клетки водорослей подсчитывали под световым стереомикроскопом (Micros MC (CAM500), Австрия). Биомассу фитопланктона (мг/л) определяли счетно-объемным методом, приравнивая клетки водорослей к соответствующим геометрическим фигурам (Vinberg, Lavrenteva, 1982; Mikheeva, 1989). Бphyto (мг С/л), определяли по уравнениям Якоби и Зохари (Yacobi, Zohary, 2010). Размеры клеток измеряли с помощью окуляр-микрометра, размерная структура фитопланктона была представлена тремя фракциями – <30, 30–60 и >60 мкм. Сырую Бzoo в пробах рассчитывали, используя зависимость массы особи от ее длины (Балушкина, Винберг, 1979). Для расчетов Бzoo в единицах углерода применяли коэффициент, предложенный Алимовым (Alimov, 1989).
ПП в озере определяли расчетным методом. Учитывая, что фотосинтетическая активность разных групп планктонных водорослей (цианобактерий, диатомовых водорослей и динофлагеллят) в условиях умеренных широт находится в относительно узких границах и с учетом их эквивалентного соотношения в разных биотопах, а также прозрачности воды в исследуемый период, суточный П/Б коэффициент в озере принимали равным 0.44 (Михеева, 1970, 1999). ВПzoo определяли, используя регрессионные модели Стоквелла и Йоханссона (Stockwell, Johannsson, 1997).
Эффективность переноса углерода от продуцентов к консументам в единице объема воды определяли как отношение ВП к ПП, выраженное в % (Gladyshev et al., 2011). Использование величин продукций на единицу объема воды (литр), характеризует эффективность переноса вещества в условиях конкретного водоема. Продукцию на единицу биомассы оценивали как соотношение ПП/Бphyto и ВП/Бzoo, а эффективность передачи углерода от фитопланктона к зоопланктону на единицу биомассы как ВП/Бzoo : ПП/Бphyto (%). Это последнее отношение характеризует эффективность переноса веществ на единицу биомассы, т.е. оно не зависит от численности планктонных организмов в воде, а скорее является индикаторной величиной “потенциала” данного планктонного сообщества.
Скорость аккумуляции азота (N) и фосфора (P) фитопланктоном (phyto) и зоопланктоном (zoo) на единицу объема воды (А) и на единицу биомассы A' рассчитывали по формулам:
A P/Nphyto = (P/Nphyto : С) × ПП
А' P/Nphyto = (P/Nphyto : С) × ПП/Бphyto
A P/Nzoo = (P/Nzoo : С) × ВП
А' P/Nzoo = (P/Nzoo : С) · ВП/ Бzoo,
где А – скорость аккумуляции на единицу объема воды, A' – скорость аккумуляции на единицу биомассы, P/N – фосфор или азот (единицы измерения: Р/N : C – мг/мг, ВП и ПП – мг С/(л сут), Бphyto и Бzoo – мг С/л).
Эффективность передачи азота и фосфора от фитопланктона к зоопланктону в литре воды и на единицу биомассы рассчитывали по формулам:
(А P/Nzoo/АP/Nphyto) × 100%
(А' P/Nzoo/А'P/Nphyto) × 100%.
Для определения содержания общих ЖК, ПНЖК, углерода, азота и фосфора в фитопланктоне воду профильтровывали через газ с размером ячеи 110 мкм, чтобы удалить зоопланктон и крупные частицы, а далее воду пропускали через стекловолоконные фильтры GF/F (Microbio) до их интенсивного окрашивания. Затем фильтры на ЖК подсушивали, помещали в раствор хлороформа с метанолом (2 : 1 объемных частей) и хранили при температуре –20°С. Фильтры на углерод, азот и фосфор высушивали в течение суток при температуре 60–65°С и хранили в эксикаторе до проведения дальнейшего анализа.
Для определения общего содержания ЖК, ПНЖК, углерода, азота и фосфора в зоопланктоне пробы из озера концентрировали на сите с размером ячеи 110 мкм, оставшийся на сите зоопланктон высушивали с помощью фильтровальной бумаги и взвешивали. Фиксацию и хранение проб зоопланктона для анализа ЖК, углерода, фосфора и азота проводили так же, как для фитопланктона.
Общие ЖК и ПНЖК в фитопланктоне и зоопланктоне измеряли с помощью газового хроматографа с масс-спектрометрическим детектором (модель 6890/5975С, Agilent, США) (Kalachova, 2011). Содержание углерода, азота и фосфора определяли на элементном анализаторе Flash EA 1112 NC Soil/MAS 200 (ThermoQuest, Italy) (Gladyshev et al., 2007). Содержание фосфора измеряли фотоколориметрическим методом (Murphy, Riley, 1962).
Продукцию ПНЖК (ЭПК, ДГК) и общих ЖК в фитопланктоне и зоопланктоне, а также эффективность передачи ПНЖК (ЭПК, ДГК) и общих ЖК от водорослей к зоопланктону на единицу объема воды и на единицу биомассы определяли с использованием таких же отношений, что и для биогенных элементов. Продукция и эффективность передачи каждого вещества и элемента, а также единицы их значений для удобства представлены в табл. 1 и 2.
Таблица 1.
Скорость аккумуляции азота и фосфора в фитопланктоне и зоопланктоне, эффективность передачи азота и фосфора от фитопланктона к зоопланктону (М ± SE) в оз. Обстерно
| Показатель | Формула вычисления, размерность | Пелагиаль | Литораль | |||
|---|---|---|---|---|---|---|
| открытая | заросшая | F | р | |||
| Скорость аккумуляции в фитопланктоне: | ||||||
| азота | $\frac{{{\text{(N:C,}}\,\,{\text{мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ПП,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}})}}{{{\text{(N:C,}}\,\,{\text{мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ПП/Б, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}})}}$ | $\frac{{0.002 \pm 0.001}}{{0.065 \pm 0.005}}$ | $\frac{{0.002 \pm 0.001}}{{0.058 \pm 0.005}}$ | $\frac{{0.003 \pm 0.001}}{{0.041 \pm 0.007}}$ | $\frac{{0.08}}{{4.75}}$ | $\frac{{0.93}}{{0.06}}$ |
| фосфора | $\frac{{{\text{(P:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ПП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}})}}{{{\text{(P:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ПП/Б, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}})}}$ | $\frac{{0.0001 \pm 0.000}}{{0.003 \pm 0.001}}$ | $\frac{{0.0001 \pm 0.000}}{{0.004 \pm 0.001}}$ | $\frac{{0.0004 \pm 0.000}}{{0.004 \pm 0.001}}$ | $\frac{{2.70}}{{0.66}}$ | $\frac{{0.15}}{{0.55}}$ |
| Скорость аккумуляции в зоопланктоне: | ||||||
| азота | $\frac{{{\text{(N:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ВП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}})}}{{{\text{(N:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ВП/Б, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}})}}$ | $\frac{{0.0003 \pm {{{0.000}}^{a}}}}{{0.034 \pm 0.001}}$ | $\frac{{0.0000 \pm {{{0.000}}^{b}}}}{{0.030 \pm 0.004}}$ | $\frac{{0.0000 \pm {{{0.000}}^{b}}}}{{0.026 \pm 0.002}}$ | $\frac{{46.28}}{{2.07}}$ | $\frac{{0.0002}}{{0.21}}$ |
| фосфора | $\frac{{{\text{(P:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ВП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}})}}{{{\text{(P:C, мг}}\,{\text{м}}{{{\text{г}}}^{{ - {\text{1}}}}}) \times ({\text{ВП/Б, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}})}}$ | $\frac{{28.5 \cdot {{{10}}^{{ - 6}}} \pm 7.8 \cdot {{{10}}^{{ - 6\,a}}}}}{{0.003 \pm 0.001}}$ | $\frac{{12.4 \cdot {{{10}}^{{ - 6}}} \pm 4.3 \cdot {{{10}}^{{ - 6\,a{\text{,}}b}}}}}{{0.006 \pm 0.001}}$ | $\frac{{3.3 \cdot {{{10}}^{{ - 6}}} \pm 0.5 \cdot {{{10}}^{{ - 6\,b}}}}}{{0.002 \pm 0.000}}$ | $\frac{{6.11}}{{1.42}}$ | $\frac{{0.04}}{{0.31}}$ |
| Эффективность передачи от фитопланктона к зоопланктону, %: | Скорость аккумуляции в зоопланктоне/скорость аккумуляции в фитопланктоне | |||||
| азота | $\frac{{13.38 \pm {{{2.95}}^{а}}}}{{53.14 \pm 4.71}}$ | $\frac{{1.75 \pm {{{0.48}}^{b}}}}{{50.63 \pm 3.18}}$ | $\frac{{2.93 \pm {{{0.48}}^{b}}}}{{65.70 \pm 6.67}}$ | $\frac{{13.39}}{{2.55}}$ | $\frac{{0.001}}{{0.16}}$ | |
| фосфора | $\frac{{26.33 \pm {{{6.28}}^{а}}}}{{107.99 \pm 24.00}}$ | $\frac{{10.03 \pm {{{5.43}}^{{а,{\text{ }}b}}}}}{{190.77 \pm 111.47}}$ | $\frac{{1.18 \pm {{{0.41}}^{b}}}}{{44.17 \pm 9.47}}$ | $\frac{{7.07}}{{0.03}}$ | $\frac{{1.24}}{{0.35}}$ | |
Таблица 2.
Продукция ПНЖК в фитопланктоне и зоопланктоне, эффективность передачи ПНЖК (ЭПК, ДГК) и ЖК от фитопланктона к зоопланктону (М ± SE) в оз. Обстерно
| Показатель | Формула вычисления, размерность | Литораль | |||
|---|---|---|---|---|---|
| открытая | заросшая | t | р | ||
| Продукция в фитопланктоне: | |||||
| ЭПК | $\frac{{\left( {{\text{ЭПК:C,}}\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ЭПК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{0.033 \pm 0.008}}{{0.797 \pm {{{0.076}}^{a}}}}$ | $\frac{{0.080 \pm 0.017}}{{1.178 \pm {{{0.058}}^{b}}}}$ | $\frac{{2.53}}{{4.00}}$ | $\frac{{0.06}}{{0.02}}$ |
| ДГК | $\frac{{\left( {{\text{ДГК:C,}}\,\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ДГК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}})/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{0.025 \pm 0.005}}{{0.602 \pm {{{0.024}}^{a}}}}$ | $\frac{{0.054 \pm 0.011}}{{0.797 \pm {{{0.026}}^{b}}}}$ | $\frac{{2.36}}{{5.55}}$ | $\frac{{0.08}}{{0.01}}$ |
| ЖК | $\frac{{\left( {{\text{ЖК:C,}}\,\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ЖК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ПП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{1.690 \pm 0.238}}{{42.959 \pm 6.358}}$ | $\frac{{2.989 \pm 0.507}}{{45.254 \pm 3.667}}$ | $\frac{{2.32}}{{0.31}}$ | $\frac{{0.08}}{{0.77}}$ |
| Продукция в зоопланктоне: | |||||
| ЭПК | $\frac{{\left( {{\text{ЭПК:C,}}\,\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ЭПК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{0.0003 \pm 0.0000}}{{0.135 \pm 0.018}}$ | $\frac{{0.0003 \pm 0.0000}}{{0.163 \pm 0.009}}$ | $\frac{{0.87}}{{1.41}}$ | $\frac{{0.43}}{{0.23}}$ |
| ДГК | $\frac{{\left( {{\text{ДГК:C,}}\,\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ДГК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{0.0007 \pm 0.0000}}{{0.101 \pm {{{0.012}}^{a}}}}$ | $\frac{{0.001 \pm 0.000}}{{0.208 \pm {{{0.027}}^{b}}}}$ | $\frac{{1.63}}{{3.59}}$ | $\frac{{0.18}}{{0.02}}$ |
| ЖК | $\frac{{\left( {{\text{ЖК:C,}}\,\,{\text{мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП, мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}} \right)}}{{\left( {{\text{ЖК:C, мг}}\,{{{\text{г}}}^{{ - {\text{1}}}}}} \right) \times \left( {{\text{ВП/Б,}}\,\,{\text{мгС }}{{{\text{л}}}^{{ - {\text{1}}}}}{\text{ су}}{{{\text{т}}}^{{ - {\text{1}}}}}/{\text{мгС }}{{{\text{л}}}^{{ - 1}}}} \right)}}$ | $\frac{{0.006 \pm 0.000}}{{2.888 \pm 0.252}}$ | $\frac{{0.005 \pm 0.001}}{{3.030 \pm 0.190}}$ | $\frac{{2.08}}{{0.45}}$ | $\frac{{0.11}}{{0.68}}$ |
| Эффективность передачи от фитопланктона к зоопланктону: | Продукция в зоопланктоне/продукция в фитопланктоне, % | ||||
| ЭПК | ЭПКzoo/ ЭПКphyto | $\frac{{0.986 \pm {{{0.188}}^{a}}}}{{16.897 \pm 1.335}}$ | $\frac{{0.354 \pm {{{0.048}}^{b}}}}{{13.963 \pm 1.204}}$ | $\frac{{3.25}}{{1.63}}$ | $\frac{{0.03}}{{0.18}}$ |
| ДГК | ДГКzoo/ДГКphyto | $\frac{{0.943 \pm {{{0.039}}^{a}}}}{{16.806 \pm 1.604}}$ | $\frac{{0.646 \pm {{{0.033}}^{b}}}}{{26.359 \pm 3.984}}$ | $\frac{{5.84}}{{2.22}}$ | $\frac{{0.004}}{{0.09}}$ |
| ЖК | ЖКzoo/ЖКphyto | $\frac{{0.395 \pm {{{0.062}}^{a}}}}{{6.840 \pm 0.405}}$ | $\frac{{0.169 \pm {{{0.012}}^{b}}}}{{6.848 \pm 0.954}}$ | $\frac{{3.60}}{{0.01}}$ | $\frac{{0.02}}{{0.99}}$ |
Статистический анализ данных проводили в программе Past 3.20. Для проверки данных на нормальность распределения использовали тест Шапиро–Уилка. Биомассу фитопланктона, размерные группы водорослей, биомассу зоопланктона, продукцию фито- и зоопланктона, П/Б коэффициенты и их соотношения между биотопами сравнивали с помощью однофакторного дисперсионного анализа (Тьюки-тест). Сравнение аккумуляции азота и фосфора, а также эффективности их передачи между разными биотопами проводили с использованием однофакторного дисперсионного анализа. Для сравнения эффективности передачи ПНЖК и суммы ЖК между биотопами использовали t-тест Стьюдента.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
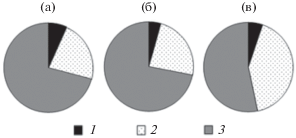
Общая биомасса фитопланктона, биомасса цианобактерий и диатомовых водорослей в оз. Обстерно не различались между исследуемыми биотопами. Биомасса золотистых водорослей была выше в заросшей камышом литорали по сравнению с пелагиалью (F = 6.10, р = 0.04). Во всех трех биотопах по биомассе доминировали крупные диатомовые водоросли Asterionella formosa Hass., Fragilaria crotonensis Kitt. и Aulacoseira granulata (Ehr.) Sim. (рис. 1). Из цианобактерий лидировали в основном два вида – Microcystis aeruginosa (Kütz.) Kütz. и Anabaena spiroides Kleb. Преобладающей размерной фракцией были водоросли >60 мкм, однако достоверных различий в биомассе этой фракции между изучаемыми биотопами не обнаружено (рис. 2). Биомасса водорослей размером 30–60 мкм в заросшей камышом литорали превышала таковую в пелагиали (F = 5.94, р = 0.04). Достоверных различий в биомассе фракции <30 мкм между биотопами не зарегистрировано. Биомасса водорослей этой фракции, предпочитаемой ракообразными-фильтраторами (Sommer, Sommer, 2006), была сравнительно низкой. В составе данной размерной фракции доминировали мелкие виды диатомовых водорослей (Cyclotella sp., Cocconeis placentula Ehr., Achnanthes lanceolata (Bréb. ex Kütz.) Grun., Gomphonema parvulum (Kütz.) Kütz.), мелкие динофитовые Glenodinium pygmaeum (Lindem.) J. Schill., а также монадные зеленые, эвгленовые (Phacus sp.) и криптофитовые (Rhodomonas pusilla (H. Bachm.) Javorn.).
Рис. 1.
Соотношение таксономических групп фитопланктона (мг/л) в оз. Обстерно. 1 – золотистые, 2 – диатомовые, 3 – динофитовые, 4 – цианобактерии, 5 – другие (зеленые, эвгленовые, криптофитовые) водоросли.
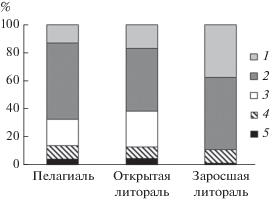
Рис. 2.
Соотношение размерных групп фитопланктона (мг/л) в пелагиали (а), открытой (б) и заросшей (в) литорали оз. Обстерно. 1 – <30 мкм, 2 – 30–60 мкм, 3 – >60 мкм.
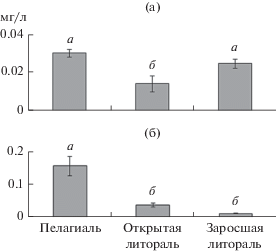
Биомасса веслоногих ракообразных была значимо выше в пелагиали, чем в обоих биотопах литорали (F = 39.66, р = 0.0003) (рис. 3). Биомасса ветвистоусых ракообразных в пелагиали и заросшей камышом литорали достоверно превышала таковую в открытой литорали (F = 13.39, р = 0.01).
Рис. 3.
Средняя биомасса ветвистоусых (а) и веслоногих (б) ракообразных в оз. Обстерно (±SE). а и б над столбцами – достоверные различия при р < 0.05 (однофакторный дисперсионный анализ, Тьюки-тест).
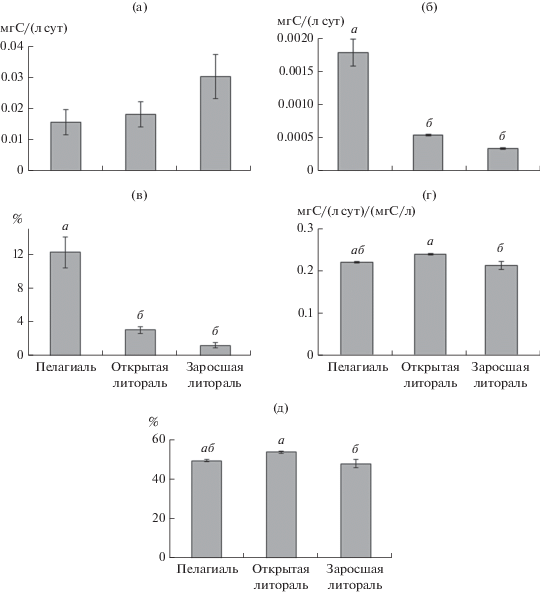
Первичная продукция фитопланктона не различалась между биотопами (рис. 4). Вторичная продукция в пелагиали была выше, чем в обоих биотопах литорали (F = 47.88, р = 0.0002). Эффективность передачи углерода от фитопланктона к зоопланктону в пелагиали на единицу объема воды достигала в среднем 12% и значимо превышала эффективность в открытой (3%) и заросшей литорали (1%) (F = 27.94, р = 0.001). Эффективность передачи углерода от фитопланктона к зоопланктону на единицу биомассы была достоверно выше в открытой, чем в заросшей литорали, 53 и 48% соответственно (F = 5.53, р = 0.04), однако различий в эффективности передачи углерода между биотопами литорали и пелагиалью, где она достигала ~50%, не выявлено.
Рис. 4.
Средние значения первичной продукции фитопланктона (а), вторичной продукции ракообразных (б), отношение вторичной продукции ракообразных и первичной продукции (в), отношение вторичной продукции ракообразных к их биомассе (г) и эффективность передачи углерода от фитопланктона к ракообразным на единицу биомассы (±SE) (д) в оз. Обстерно. Буквами а и б над фигурами обозначены достоверные различия при р < 0.05 (однофакторный дисперсионный анализ, Тьюки-тест).
Скорости аккумуляция N и P в сестоне достоверно не различались между биотопами (табл. 1). Скорость аккумуляции N в зоопланктоне на единицу объема была выше в пелагиали, что связано с более высокой вторичной продукцией в этом биотопе. Фосфор в зоопланктоне аккумулировался с большей скоростью в пелагиали, чем в заросшей литорали. Скорости аккумуляции N и P в зоопланктоне на единицу биомассы не различались между биотопами. Эффективность передачи N на единицу объема была достоверно выше в пелагиали (13%), чем в литорали (2 и 3% в открытой и заросшей литорали соответственно). Эффективность передачи P составляла в пелагиали 26%, тогда как в открытой и заросшей литорали 10% и 1%, соответственно. Таким образом, эффективность передачи азота и фосфора на единицу объема в пелагиали была достоверно выше, чем в биотопах литорали. Однако значимых различий в эффективности передачи N и P на единицу биомассы между биотопами не обнаружено, эффективность передачи N достигала 51–66%, P – 44–191%.
Эффективность переноса ПНЖК (ЭПК и ДГК), а также суммы ЖК на единицу объема в открытой литорали достоверно превышала таковую в заросшей (табл. 2). Эффективность переноса ЭПК, ДГК и суммы ЖК в единице объема составляла <1% и была достоверно выше в литорали без зарослей для всех ЖК. Однако эффективность переноса этих веществ на единицу биомассы составляла 14–17% для ЭПК, 17–26% для ДГК и 7% для суммы ЖК, но не различалась статистически между биотопами. Эффективность переноса ЖК в целом была ниже, чем эффективность переноса углерода, азота и фосфора.
ОБСУЖДЕНИЕ PЕЗУЛЬТАТОВ
Большинство озер умеренной зоны в мире являются малыми и высокопродуктивными, а их литоральная зона представляет собой место нереста и нагула молоди рыб. Синтез и перераспределение органических веществ в трофической сети водоемов – это часть глобального цикла углерода, который может протекать с разной скоростью и эффективностью в водоемах в зависимости от многих факторов. Согласно правилу Раймонда Линдемана (Lindeman, 1942), в любой трофической пирамиде только 10% органического вещества, образованного на одном трофическом уровне, участвует в продукции следующего трофического уровня. Однако эффективность передачи энергии – это изменчивый показатель, зависящий от факторов среды (Slobodkin, 1972; Lacroix, 1999), поэтому важно знать какие факторы влияют на эффективность переноса биологически ценных веществ от продуцентов к консументам. В настоящем исследовании в условиях мезотрофного озера мы сравнивали три биотопа – пелагиаль, литораль без зарослей и заросшую литораль, где в период исследований, происходило питание молоди рыб. Показано, что эффективность переноса биологически ценных веществ от фитопланктона к зоопланктону в оз. Обстерно различалась между биотопами. Эффективность переноса углерода в литорали достигала 1–3%, в пелагиали – 12%. Эффективность передачи фосфора была также наибольшей в пелагиали – 26%, в литорали она варьировала от 1 до 10%. Азот передавался с эффективностью 13% в пелагиали и 2–3% в литорали, однако эффективность передачи ПНЖК и общих ЖК не превышала 1% и была достоверно выше в литорали без зарослей, чем в заросшем мелководье. Таким образом, в озере ЖК, включая ПНЖК, передавались от фитопланктона к зоопланктону с наименьшей эффективностью, чем общие углерод, азот и фосфор. На различия в эффективности передачи разных биологически ценных веществ впервые обращено внимание в работе Гладышева и др. (Gladyshev et al., 2011). В пелагиали эвтрофного водоема эффективность переноса ЖК превышала таковую общего углерода, однако оценку эффективности переноса веществ проводили в масштабе целого сезона. Известно, что в разные сезоны года в планктоне развиваются разные группы водорослей, которые характеризуются разными количественными и качественными потребностями и накоплением тех или иных соединений, например, азота, фосфора, а также способностью синтезировать различные ЖК. Известно, что в большинстве пресноводных водоемов умеренной зоны в процессе сезонной сукцессии в фитопланктоне идет смена доминирующих комплексов (Sommer et al., 1986). Очевидно, сезонная сукцессия видового состава сопряжена с сезонной сукцессией элементного и биохимического состава фитопланктона. Так, летом на смену мелким быстро растущим съедобным диатомовым и криптофитовым, которые богаты азотом и ПНЖК (ДГК), приходят несъедобные цианобактерии, богатые фосфором, но бедные ПНЖК. Осенью возможна их смена на крупные диатомовые с высоким содержанием ПНЖК, но непригодные по размерам в пищу для большинства фильтраторов. В период исследований в озере баланс между отдельными группами фитопланктона (золотистыми, диатомовыми и динофитовыми) во всех биотопах выражался в балансе содержания азота и фосфора и между биотопами концентрация каждого элемента в фитопланктоне достоверно не различалась (табл. 1). Однако содержание в зоопланктоне и азота, и фосфора достоверно превышало таковое в пелагиали по сравнению с литоральными биотопами. Эта разница в содержании отдельных элементнов могла быть обусловлена соотношением веслоногих и ветвистоусых ракообразных в литорали и пелагиали (большей долей и биомассой диаптомид в пелагиали), а также соотношением видов с низким (Bosmina spp.) и высоким (Ceriodaphnia pulchella Sars, 1862) содержанием фосфора (Sterner, Schulz, 1998). Так, общая биомасса Calanoida в пелагиали, заросшей литорали и литорали без зарослей была 0.057 ± 0.009, 0.00012 ± 0.0000 и 0.0045 ± 0.0017 мг/л соответственно. В итоге, в оз. Обстерно на единицу объема воды эффективнее передавались биогенные вещества от фитопланктона к зоопланктону в пелагиали, чем в литорали.
В оз. Обстерно эффективность передачи углерода от фитопланктона к зоопланктону на единицу биомассы достигала 48–53%. ДГК в озере передавалась с эффективностью 16–26%, ЭПК – 14–16%, сумма ЖК – 7%. Фосфор в озере передавался с эффективностью 107–190%, азот – 50–65%. Между биотопами эффективность передачи веществ на единицу биомассы не различалась. Высокая эффективность переноса фосфора от фитопланктона к зоопланктону связана со способностью фосфора накапливаться в тканях зоопланктона, в большей части у ветвистоусых (Karpowicz et al., 2019). ПНЖК и общие ЖК передавались с меньшей эффективностью, чем углерод и биогенные элементы.
Разные вещества могут передаваться с разной эффективностью. Так, Гладышев и др. (Gladyshev et al., 2011) на примере эвтрофного озера показали, что ПНЖК передаются от фитопланктона к зоопланктону в 2 раза эффективнее, чем общий органический углерод. Различия в эффективности передачи разных веществ от фитопланктона к зоопланктону возможны благодаря способности ракообразных регулировать содержание биологически ценных веществ в своем организме, удерживая те, которых недостаточно, и, наоборот, выделяя те, которые в избытке (Sterner, 1993, 1997; Sterner et al., 1998; Sterner, Elser, 2002). Кроме того, ракообразные-фильтраторы преимущественно потребляют пищевые частицы в диапазоне от 1 до 20–30 мкм (Sommer, Sommer, 2006). В оз. Обстерно биомасса фракции водорослей, превышающих по размерам диапазон частиц доступных для ракообразных, составляла более половины общей биомассы, что может быть связано с выеданием мелкого фитопланктона в конце мая–первой половине июня, когда в озере доминируют дафнииды. Кроме того, ракообразные по-разному усваивают пищевые частицы. Таипл и др. (Taipale et al., 2011) показали, что значения δ13C у ветвистоусых ракообразных не коррелируют с δ13C сестона, но коррелируют с δ13C отдельных таксонов фитопланктона, что свидетельствует об избирательном пищевом поведении ракообразных. Избирательность в ассимиляции пищевых частиц также продемонстрирована в экспериментальной работе Гладышева и др. (Gladyshev et al., 2000).
Мы полагаем, что эффективность передачи ПНЖК может меняться в зависимости от состава фитопланктона и зоопланктона и, может быть, как выше (Gladyshev et al., 2011), так и ниже эффективности передачи углерода, в зависимости от состояния планктонных сообществ, времени и условий отбора проб и др. (Slobodkin, 1972). От качества фитопланктона будет зависеть качество зоопланктона как ресурса для вышестоящих трофических уровней. ЖК, включая ПНЖК, передавались менее эффективно, чем углерод. Напротив, аккумуляция биогенных элементов, особенно фосфора была более эффективной по сравнению с углеродом, что свидетельствует о важности зоопланктонных сообществ как поставщиков биогенных элементов вверх по трофической цепи. Планктонные сообщества пелагиали мезотрофного озера оказались более эффективными в передаче веществ от первичных продуцентов к консументам, чем литоральные сообщества.
Выводы. В условиях мезотрофного озера биологически ценные вещества передаются от фитопланктона к зоопланктону с разной эффективностью. Эффективность переноса фосфора от фитопланктона к зоопланктону выше, чем углерода. Жирные кислоты, включая ПНЖК, передаются с меньшей эффективностью, чем углерод. Эффективность переноса ПНЖК в единице объема выше в литорали без зарослей, чем в заросшей литорали. С наибольшей эффективностью передаются ДГК и ЭПК, с наименьшей короткоцепочечные насыщенные ЖК. Эффективность передачи углерода и биогенных веществ в единице объема была выше в пелагиали, чем в литорали, что, предположительно, связано с более высокой долей диаптомид.
Список литературы
Балушкина Е.В., Винберг Г.Г. 1979. Зависимость между длиной и массой тела планктонных ракообразных и коловраток // Эколого-физиологические основы изучения водных экосистем. Ленинград: Наука. С. 169.
Михеева Т.М. 1970. Оценка продукционных возможностей единицы биомассы фитопланктона // Биологическая продуктивность эвтрофного озера. Москва: Наука. С. 50.
Михеева Т.М. 1999. Альгофлора Беларуси // Таксономический каталог. Минск: Белорус. гос. ун-т.
Alimov A.F. 1989. An introduction to production hydrobiology. Leningrad: Gidrometeoizdat.
Becker C., Boersma M. 2005. Differential effects of phosphorus and fatty acids on Daphnia magna growth and reproduction // Limnol., Oceanogr. V. 50. P. 388.
Blue book of Belarus. 1994. Minsk: Bel. En.
Boersma M., Schops C., McCauley E. 2001. Nutritional quality of seston for the freshwater herbivore Daphnia galeatahyalina: biochemical versus mineral limitation // Oecologia. V. 129. P. 342.
Buseva Zh., Pljuta M. 2015. Feeding of YOY fish in littoral zone of shallow lake // Proceedings of the National Academy of Sciences of Belarus. V. 59. № 3. P. 71.
DeMott W.R. 1998. Utilization of a cyanobacterium and a phosphorus-deficient green alga as a complementary resource by daphnids // Ecology. V. 79. P. 2463.
DeMott W.R., Gulati R.D., Siewertsen K. 1998. Effects of phosphorus-deficient diets on the carbon and phosphorus balance of Daphnia magna // Limnol., Oceanogr. V. 43. P. 1147.
Dickman E.M., Newell J.M., González M.J., Vanni M.J. 2008. Light, nutrients, and food chain length constrain planktonic energy transfer efficiency across multiple trophic levels // Proc. Natl. Acad. Sci. USA. V. 105. P. 18408. https://doi.org/10.1073pnas.0805566105
Elser J.J., Kyle M., Frost P. et al. 2002. Effects of light and nutrients on plankton stoichiometry and biomass in a P-limited lake // Hydrobiologia. V. 481. P. 101.
Feller S.E. 2008. Acyl chain conformations in phospholipid bilayers: A comparative study of docosahexaenoic acid and saturated fatty acids // Chem. Phys. Lipids. V. 153. P. 76. https://doi.org/10.1016/j.chemphyslip.2008.02.013
Gladyshev M.I., Sushchik N.N., Dubovskaya O.P. et al. 2006. Influence of sestonic elemental and essential fatty acid contents in a eutrophic reservoir in Siberia on population growth of Daphnia (longispina group) // J. Plankton Res. V. 28. № 10. P. 907. https://doi.org/10.1093/plankt/fbl028
Gladyshev M.I., Sushchik N.N., Kolmakova A.A. et al. 2007. Seasonal correlations of elemental and v-3 PUFA composition of seston and dominant phytoplankton species in a eutrophic Siberian Reservoir // Aquat. Ecol. V. 41. P. 9. https://doi.org/10.1007/s10452-006-9040-8
Gladyshev M.I., Kolmakov V.I., Dubovskaya O.P., Ivanova E.A. 2000. Studying of algae food composition of Daphnia longispina during blue-green bloom of eutrophic pond // Dokl. Akad. Nauk. V. 371. P. 556.
Gladyshev M.I., Sushchik N.N., Anishchenko O.V. et al. 2011. Efficiency of transfer of essential polyunsaturated fatty acids versus organic carbon from producers to consumers in a eutrophic reservoir // Oecologia. V. 165. P. 521. https://doi.org/10.1007/s00442-010-1843-6
Glencross B.E. 2009. Exploring the nutritional demand for essential fatty acids by aquaculture species // Reviews in Aquaculture. V. 1. № 2. P. 71–124. https://doi.org/10.1111/j.1753-5131.2009.01006.x
Gulati R.D., DeMott W.R. 1997. The role of food quality for zooplankton: Remarks on the state -of-the-art, perspectives and priorities // Freshwater Biol. V. 38. № 3. P. 753. https://doi.org/0.1046/j.1365-2427.1997.00275.x
Kalachova G.S., Gladyshev M.I., Sushchik N.N., Makhutova O.N. 2011. Water moss as a food item of the zoobenthos in the Yenisei River // Cent. Eur. J. Biol. V. 6. P. 236. https://doi.org/10.2478/s11535-010-0115-0
Karpowicz M., Feniova I., Gladyshev M.I. et al. 2019. The stoichiometric ratios (C:N:P) in a pelagic food web under experimental conditions // Limnologica. V. 77. 125 690. https://doi.org/10.1016/j.limno.2019.125690
Lacroix G. 1999. Biomass and production of plankton in shallow and deep lakes: are there general patterns? // Ann. Limnol. V. 35. № 2. P. 111.
Lindeman R.L. 1942. The trophic-dynamic aspect of ecology // Ecology. V. 23. P. 399.
Loladze I., Elser J.J. 2011. The origins of the Redfield nitrogen-to-phosphorus ratio are in a homoeostatic proteinto-rRNA ratio // Ecol. Lett. V. 14. P. 244. https://doi.org/10.1111/j.1461-0248.2010.01577.x
Mikheeva T.M. 1989. Methods of quantitative estimates of nanophytoplankton (review) // Hydrobiol. J. V. 25. P. 3.
Müller-Navarra D.C. 1995. Biochemical versus mineral limitation in Daphnia // Limnol., Oceanogr. V. 40. P. 1209.
Murphy J., Riley J.P. 1962. A modified single solution method for the determination of phosphate in natural waters // Anal. Chim. Acta. V. 27. P. 31.
Schmitz G., Ecker. J. 2008. The opposing effects of n-3 and n-6 fatty acids // Prog. Lipid Res. V. 47. P. 147. https://doi.org/10.1016/j.plipres.2007.12.004
Slobodkin L.B. 1972. On the inconstancy of ecological efficiency and the form of ecological theories // Growth by Intussusception: Ecological Essays in Honor of G.E. Hutchinson. New Haven: Trans. Conn. Acad. Arts and Sci. V. 44. P. 291.
Sommer U., Gliwicz Z.M., Lampert W., Duncan A. 1986. The PEG-model of a seasonal succession of planktonic events in fresh waters // Archiv für Hydrobiologie. V. 106. P. 433.
Sommer U., Sommer F. 2006. Cladocerans versus copepods: the cause of contrasting top-down controls on freshwater and marine phytoplankton // Oecologia. V. 147. P. 183. https://doi.org/10.1007/s00442-005-0320-0
Sterner R.W. 1993. Daphnia growth on varying quality of Scenedesmus: Mineral limitation of zooplankton // Ecology. V. 74. P. 2351. https://doi.org/10.2307/1939587
Sterner R.W. 1997. Modelling interactions of food quality and quantity in homeostatic consumers // Freshwater Biol. V. 38. P. 473–481. https://doi.org/10.1046/j.1365-2427.1997.00234.x
Sterner R.W., Elser J.J. 2002. Ecological stoichiometry: the biology of elements from molecules to the biosphere. New York: Princeton University Press.
Sterner R.W., Schulz K.L. 1998. Zooplankton nutrition: recent progress and a reality check // Aquat. Ecol. V. 32. P. 261.
Sterner R.W., Clasen J., Lampert W., Weisse T. 1998. Carbon: phosphorus stoichiometry and food chain production // Ecol. Lett. V. 1. P. 146. https://doi.org/10.1046/j.1461-0248.1998.00030.x
Stockwell J.D., Johannsson O.E. 1997. Temperature-dependent allometric models to estimate zooplankton production in temperate freshwater lakes // Can. J. Fish Aquat. Sci. № 54. P. 2350. https://doi.org/10.1139/f97-141
Taipale S.J., Kainz M.J., Brett M.T. 2011. Diet-switching experiments show rapid accumulation and preferential retention of highly unsaturated fatty acids in Daphnia // Oikos. V. 120. P. 1674. https://doi.org/10.1111/j.1600-0706.2011.19415.x
Vinberg G.G., Lavrenteva G.M. 1982. Guidelines for the collection and processing of materials for hydrobiological studies in freshwater bodies // Phytoplankton and its products. Leningrad: GosNIORKh. USSR. P. 1.
Wacker A., Von Elert E. 2001. Polyunsaturated fatty acids: evidence for non-substitutable biochemical resources in Daphnia galeata // Ecology. V. 82. P. 2507.
Wagner N.D., Lankadurai B.P., Simpson M.J. et al. 2015. Metabolomic differentiation of nutritional stress in an aquatic invertebrate // Physiol. Biochem. Zool. V. 88. P. 43. https://doi.org/10.1086/679637
Wassell S.R., Stillwell W. 2008. Docosahexaenoic acid domains: the ultimate non-raft membrane domain // Chem. Phys. Lipids. V. 153. P. 57. https://doi.org/10.1016/j.chemphyslip.2008.02.010
White T.C.R. 1993. The inadequate environment. Berlin: Springer.
Yacobi Y.Z., Zohary T. 2010. Carbon: chlorophyll a ratio, assimilation numbers and turnover times of Lake Kinneret phytoplankton // Hydrobiologia. V. 639. P. 185. https://doi.org/10.1007/s10750-009-0023-3
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод