

Биология внутренних вод, 2021, № 5, стр. 481-500
Основные классы реофильных сообществ макробеспозвоночных и их региональная изменчивость в Евразии
М. В. Чертопруд *
Биологический факультет Московского государственного университета им. М.В. Ломоносова
Москва, Россия
* E-mail: lymnaea@yandex.ru
Поступила в редакцию 09.11.2020
После доработки 16.01.2021
Принята к публикации 26.02.2021
Аннотация
Для десяти регионов Евразии, представляющих все климатические пояса и многие природные зоны континента, описаны особенности пяти выделенных ранее классов сообществ реофильного макрозообентоса: сообщества плотных грунтов (ритрали), мягких грунтов (пелали), зарослей макрофитов (фитали), ручьев и родников (кренали) и береговой кромки (рипали). Показано, что основные трофические группы, жизненные формы и таксоны в каждом классе сообществ повторяются в различных регионах. В зависимости от климата и рельефа региона меняется характерное соотношение различных жизненных форм, а в зависимости от биогеографического региона – состав сообществ на уровне видов и, отчасти, родов. Каждый класс сообществ обладает большой внутренней неоднородностью и может быть подразделен на значительное число вариантов. Все изученные сообщества по структуре делятся на сообщества Petersen-типа (с преобладанием комплекса личинок насекомых, низким и неустойчивым во времени обилием и относительно четкой привязкой к биотопу) и сообщества Mebius-типа (со сверхдоминированием одного–двух видов ракообразных или моллюсков, высоким обилием и слабой привязкой к конкретному биотопу). Предлагаемая классификация может считаться географически универсальной для Евразии, и потенциально может быть расширена на все типы континентальных водных объектов.
ВВЕДЕНИЕ
Сообщества водотоков, как и другие биотические сообщества, формируются в условиях сложной мозаики экологических факторов, на которую накладывается большое разнообразие составляющей их видов, различных в каждом регионе (Hynes, 1970). Картина многообразия самих сообществ также весьма сложна и требует, как минимум, адекватной классификации для любых практических действий (Pennak, 1971).
Первая типология пресноводных (как и наземных) сообществ животных принадлежит Шелфорду (Shelford, 1913) и основывается на натуралистических наблюдениях, без привлечения количественных данных. Типология Шелфорда использует как доминирующие таксоны, так и факторы среды: автор описывает наиболее выделяющиеся формации (формация Hydropsyche, формация Planorbis и др.) и группирует их по типам водоемов.
Известна система донных биоценозов Жадина (Жадин, 1940), созданная на основе особенностей грунта и течения: все биоценозы делятся на рео- и лимнофильные, и далее – по основным типам субстратов. Автор рассматривает только средние и крупные равнинные реки, игнорируя малые водотоки, а также влияние рельефа, климата и др.
Интересная типология предложена Иллиесом (Illies, 1961) для продольной зональности рек, где он выделяет креналь (зону ручьев и родников, холодных, чистых, маломощных), ритраль (зону быстрых прохладных каменистых малых рек) и потамаль (зону больших теплых равнинных рек с илисто-песчаным дном). Ключевой критерий предложенного разделения на зоны – летняя (максимальная) температура воды; при этом имеется в виду, что другие факторы (размер водотока, скорость течения, субстрат) более или менее сопряжены с ней. Эта схема в общем адекватна при умеренном климате в местности, где ручьи стекают с гор или с возвышенностей (для которой и была предложена). В других условиях определения кренали, ритрали и потамали по Иллиесу не соблюдаются (Hynes, 1970). Принципиальные структурные различия сообществ малых, средних и крупных водотоков существуют, но причины их определяются сложнее. Частично они описаны в концепции речного континуума (Vannote et al., 1980), определяющей трофические особенности разных продольных зон водотока. В ней, в частности, постулировано, что в ручьях основной источник питания – аллохтонный листовой опад и среди животных должны преобладать детритофаги-разгрызатели; в малых реках ведущую роль приобретают альгообрастания и макрофиты, а среди животных – соскребатели; в крупных реках питание обеспечивается приносимым течением микроскопическим детритом, доминируют детритофаги-собиратели и фильтраторы. Эта закономерность также искажается в разных регионах и ландшафтах (Statzner, Higler, 1985). Позднее было выдвинуто еще несколько концептуальных обобщений о закономерностях формирования и структуры реофильных сообществ (Townsend, 1989; Богатов, 1995; Poff, 1997; Thorp et al., 2006, Suren, Riis, 2010), но они не связаны прямо с проблемой типологии сообществ. В последнее время стала известна так называемая “концепция градиента речного биома” (Dodds et al., 2015), постулирующая влияние на речные сообщества регионального климата и растительности через гидрологический режим водотоков и через специфичные для каждого ландшафта нарушающие факторы (например, влияние выпаса скота или пожаров). В основном, в вышеназванных работах рассмотрено изменение различных абиотических факторов в градиенте влажности и преобладающей растительности (пустыня–степь–лес). Подобный подход представляется весьма перспективным, но пока не углубляется в описание структуры сообществ соответствующих биомов.
Все эти построения не противоречат друг другу, но освещают лишь отдельные аспекты разнообразия реофильных сообществ. Текущая задача – построение более или менее единой и цельной схемы классификации, причем основанной на достаточном массиве данных, чтобы быть устойчивой в разных регионах и ландшафтах.
Еще несколько закономерностей, неоднократно описаных (Chertoprud, 2011; Chertoprud et al., 2018), являются ключевыми для дальнейших классификационных построений. Наиболее важный результат: как правило, отдельные типы сообществ более или менее соответствуют определенным комбинациям факторов абиотической среды, т.е. биотопам в широком понимании этого слова. Из этого правила есть исключения, но они могут оговариваться отдельно. Соответствие (хотя бы и грубое) типов сообществ и биотопам позволяет дополнять биотическую классификацию биотопической, что существенно упрощает поставленную задачу.
Важный результат касается региональной изменчивости сообществ. Сопоставление региональных классификаций показывает, что многие (возможно, почти все) типы сообществ повторяются в различных регионах вместе с биотопами, к которым они приурочены, (хотя встречаются в одних регионах чаще, в других – реже). Структура сообществ одного типа в разных регионах имеет значительное (но не полное) сходство – так называемый эффект параллельных сообществ, впервые описанный Шелфордом (Shelford, 1913). Как правило, при смене региона меняется видовой состав сообщества, но сохраняется набор жизненных форм, семейств и, отчасти, родов (происходит замещение близкородственных видов). Поэтому географически универсальные типы следует выделять по составу и обилию родов и семейств, а не видов.
Наблюдаемая структура сообщества, помимо прочих факторов, может быть связана с размером рассматриваемого биотопа. Мы предпочитаем работать с масштабом первых метров – мезобиотоп (mesohabitat) в понимании Парасевича (Parasiewicz, 2001). Этот масштаб примерно соответствует индивидуальной подвижности макрофауны, т.е. ее масштабу восприятия биотопа. В большем масштабе (макробиотоп – десятки, сотни метров, километры) объектом изучения становится целый участок водотока, в этом масштабе устанавливается зональность по Иллиесу (Illies, 1961), проводится классификация рек по рыбному населению (Hawkes, 1975) и работает так называемая “Европейская рамочная директива” по контролю качества вод (European…, 2000). Масштаб макробиотопа представляется оптимальным для типизации самих водотоков, описания рыбного населения или оценки распределения качества воды, но слишком крупный для изучения сообществ бентоса. В меньшем масштабе (сантиметры-дециметры, т.е. микробиотоп) структура сообществ макрофауны теряет устойчивость в связи со случайным наличием/отсутствием особей отдельных видов (особенно крупных) – с этим, вероятно, связаны многие неудачные попытки описания этих сообществ по одиночным пробам площадью, как правило, от 0.01 до 0.1 м2.
Цель работы – на основе обзора накопленных региональных данных по типологии сообществ макрофауны выделить специфику классов сообществ, характерных для всех регионов (в пределах Евразии) и заложить основы для дальнейшей, более подробной, классификации сообществ континентальных вод. Под типом сообщества понималось повторяющееся в ряду проб соотношение обилий таксонов, не затрагивая гораздо более сложный вопрос о реальной связи видов в сообществе между собой.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
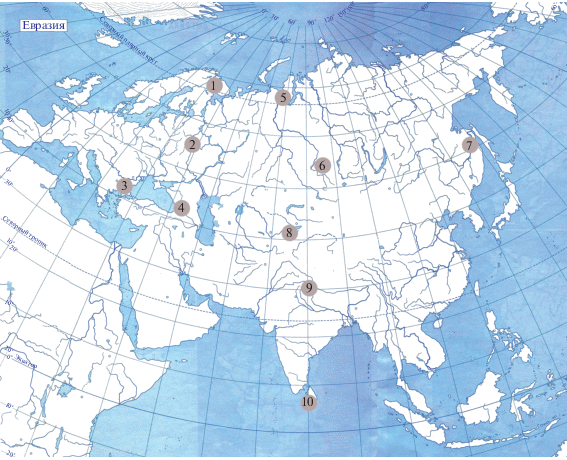
Обзор охватывает ~2950 количественных проб макробентоса, собранных для изучения разнообразия донных и зарослевых реофильных сообществ в десяти различных регионах Евразии (рис. 1): Кольский п-ов (субарктический климатический пояс, климат в горах до арктического, природные зоны северной тайги и тундры) (Чертопруд, Палатов, 2013); Московский регион (Центр Европейской России) (умеренный климатический пояс, зоны южной тайги и смешанных лесов) (Chertoprud, 2011); Восточные Балканы в Болгарии и Турции (субтропический пояс, климат в горах до умеренного, природные зоны средиземноморских кустарников, широколиственных и таежных лесов) (Чертопруд, Палатов, 2017); Кавказ и Закавказье (умеренный и субтропический пояса, климат в горах до альпийского, природные зоны средиземноморских кустарников, широколиственных и таежных лесов, альпийских лугов) (Палатов и др., 2016, Palatov, Chertoprud, 2018, 2020); Южный Ямал (субарктический пояс, зона тундры) (Палатов, Чертопруд, 2012); Новосибирский регион (умеренный климатический пояс, природные зоны степей и тайги) (Chertoprud, Palatov, 2013); Нижнее Приамурье (умеренный климатический пояс, зоны смешанных муссонных лесов и тайги (Чертопруд и др., 2020а); Западный Тянь-Шань (умеренный пояс, климат в горах до альпийского, природные зоны степей, тайги, альпийских лугов (Чертопруд и др., 2020б); Гималаи и Северная Индия (тропический пояс, климат в горах до умеренного, природные зоны переменно-влажных тропических, субтропических, широколиственных и хвойных лесов) (Chertoprud et al., 2018); о. Шри-Ланка (экваториальный пояс, климат в горах до субтропического, природные зоны влажных тропических и субтропических лесов) (Chertoprud, 2019).
Рис. 1.
Карта расположения обследованных регионов в Евразии. Цифрами обозначены регионы: 1 – Кольский п-ов, 2 – Московский регион, 3 – Восточные Балканы, 4 – Кавказ и Закавказье, 5 – Южный Ямал, 6 – Новосибирское Приобье, 7 – Нижнее Приамурье, 8 – Западный Тянь-Шань, 9 – Южные Гималаи, 10 – Шри-Ланка.
В целом, приведенные данные охватывают регионы всех климатических поясов и большинства природных зон Евразии. Большая часть материала обработана и опубликована ранее, ниже представлен обзор ранее полученных результатов, некоторые данные дополнены и проанализированы заново. Пробы отбирали во всех характерных биотопах участка водотока, за исключением глубоководных. Как правило, материал собирали с берега.
Каждую пробу отбирали на участке протяженностью от трех до десяти метров, что соответствует термину мезобиотоп в работе (Parasiewicz, 2001). На протяжении этого участка, при условии сохранения более или менее однородного биотопа, собирали несколько (обычно – 5 или 10) суммарной пробы – 0.1 или 0.2 м2 соответственно. В случае линейного биотопа (например, при изучении береговой кромки водотока подпроб, усредняемых в одну пробу, которая и служила для описания сообщества. Площадь пробоотборника была 0.02 м2) подпробы располагались вдоль образуемой субстратом линии, но так же в пределах нескольких метров. Когда биотоп был неоднороден даже в пределах пробоотборника (обычно в ручьях и родниках), его интерпретировали как смешанный. В некоторых случаях раздельно изучали биотопы, находящиеся друг под другом на разной глубине (например, зарослей макрофитов и илистого дна под ними).
В качестве основной меры обилия таксонов использовали величину метаболизма, рассчитываемую на основе численности и биомассы каждого таксона по формуле: D = kN0.25B0.75, где N – численность вида, В – биомасса, k – специфичный для группы коэффициент. Считается, что этот показатель прямо отражает потребление животными пищи, а также их потребности в дыхании и, соответственно, более объективен для оценки роли организмов в сообществах (Алимов, 1979; Жирков, 2010). В сравнении с ним, оценка обилия по биомассе приводит к завышению роли крупных организмов (например, двустворчатых моллюсков сем. Unionidae), а оценка по численности – наоборот, к завышению роли мелких форм (чаще всего хирономид). Для изучения структуры сообществ вычисляли относительный метаболизм – доля каждого вида по метаболизму в процентах. При оценке абсолютного метаболизма вносили поправки на локальную температуру воды в соответствии с кривой Крога (Алимов, 1979). Следует отметить, что при работе с относительной биомассой результаты по типизации сообществ были сходны с таковыми по метаболизму.
Для выделения типов сообществ применяли популярный в геоботанике метод Браун-Бланке (Braun-Blanquet, 1964), модифицированный для количественных данных (Chertoprud, 2011). Он включает выделение комплексов совместно встречающихся массовых видов, с последующей привязкой, насколько это возможно, к типу биотопа и описанию действия других факторов, существенных для данной серии станций. Классификационные построения проводили раздельно в каждом из изученных регионов, затем их сопоставляли для выявления региональной специфики, общих закономерностей и построения классификации, не привязанной к конкретному региону.
При описании структуры сообществ использовали, помимо названий таксонов, их жизненные формы, включающие, в применяемой нами трактовке, характеристики подвижности организма (плавающий, ползающий, роющий, малоподвижный), основного способа защиты от хищников (панцирный, роющий, щелевой и др.) и характера питания (альгофаг-соскребатель, эврифаг, хищник и др.). Тип питания в основном определен по классификации Камминса (Cummins, Klug, 1979) с уточнениями по другим авторам (Монаков, 1998; Паньков, 2004); тип подвижности и защиты – главным образом, по собственным наблюдениям.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Сравнение региональных классификаций реофильных сообществ показало, что сообщества всех регионов относятся к пяти крупным классам, в основном имеющим биотопическую привязку. Эти классы получили названия по соответствующим биотопам: сообщества ритрали (плотных грунтов), пелали (илисто-песчаных грунтов), фитали (зарослей макрофитов), кренали (ручьев и родников) и рипали (субстратов береговой кромки). Все эти классы характеризуются специфической структурой на уровне жизненных форм и семейств. Сообщества пелали, как правило, занимают основную площадь дна крупных равнинных рек длиной >100 км, креналь преобладает в ручьях длиной ≤5 км, в малых реках в разных соотношениях сочетаются сообщества ритрали, пелали, фитали и рипали.
Представленность этих классов в разных регионах сильно варьирует, главным образом, в зависимости от преобладающего рельефа местности данного региона. В целом, в горных регионах преобладают сообщества ритрали и кренали, в холмистых равнинах примерно в равной степени представлены все классы сообществ, а на низменностях встречаются в основном сообщества пелали, фитали и рипали. С другой стороны, эти данные зависят и от выбора исследователем конкретных водотоков и биотопов, а также от уровня антропогенной деградации водотоков в каждом регионе. Например, в тропических и субтропических регионах сильно подвержены деградации равнинные водотоки и их сообщества, в силу чего данные по ним более скудны, и большинство сборов относится к горным районам.
Ниже приведено подробное описание выделенных нами классов сообществ: особенности таксономической структуры и условий местообитания, показатели суммарного обилия и географическая изменчивость.
Сообщества ритрали (плотных субстратов)
Класс сообществ, обычно связанный значительной скоростью течения. Наиболее типичный биотоп ритрали – каменистые перекаты быстрых рек, хотя близкие условия (и сообщества) могут наблюдаться также на затопленных древесных объектах, некоторых водных растениях, скальных монолитах, галечном грунте и даже на каменистых прибойных берегах крупных озер. Предложивший этот термин для предгорных малых рек Иллиес (Illies, 1961) вводил для ритрали также температурные ограничения, но они теряют смысл при рассмотрении регионов с различным климатом и рельефом. Ритраль, в нашем понимании, в основном, сходна с расширенным понятием ритрали Леванидова (1981) и с литореофильными сообществами в понимании Жадина (1940). Как правило, в сообществах ритрали преобладают личинки амфибиотических насекомых (поденок, ручейников или двукрылых). Центральное, несущее наиболее полный набор жизненных форм сообщество ритрали, – сообщество каменистых перекатов некрупных быстрых, чистых и прохладных рек (как и описана типичная ритраль у Иллиеса). Сообщества ритрали встречаются во всех регионах и ландшафтах Евразии, в горах являются основным классом сообществ и наиболее разнообразны по таксономической структуре, на возвышенностях встречаются наравне с другими классами сообществ, в низменностях относительно редки и часто обеднены таксонами.
Местообитание ритрали имеет характерный комплекс взаимосвязанных условий среды. Постоянное течение воды требует специальных адаптаций макробеспозвоночных к нему: надежного прикрепления к субстрату, обтекаемой формы тела или укрытия в щелях. С другой стороны, течение обеспечивает постоянное перемешивание воды и в целом стабильно благоприятный кислородный режим. Кроме того, вода постоянно приносит мелкодисперсные взвеси, что позволяет организмам зообентоса реализовать энергетически очень эффективную стратегию пассивной фильтрации. Плотные субстраты обычно снабжены надежной поверхностью для прикрепления и щелевыми микробиотопами. Сочетание небольшой глубины и высокой освещенности благоприятно для развития водорослей на верхней поверхности субстратов, которые служат пищей для альгофагов-соскребателей. С другой стороны, при их сильном развитии (обычно в крупных водотоках) обрастания становятся недоступны для соскребателей. По-видимому, зообентос ритрали слабо выедается рыбами, хотя литературные данные на эту тему противоречивы – от констатации отсутствия значимого влияния рыб (Culp, 1986; Flecker, Allan, 1984) до вполне существенного их воздействия (Forrester, 1994; Power, 1992). Предположительно, роль выедания рыбами возрастает в более крупных реках и в тропиках с увеличением разнообразия реофильных рыб (Dudgeon, 1999), но снижается или даже полностью исчезает в горах. Кроме того, рыбы эффективно выедают бентос (в том числе ритральный) во время дрифта (Forrester, 1994).
Состав основных жизненных форм: малоподвижный альгофаг-соскребатель (брюхоногие моллюски сем. Ancylidae; ручейники сем. Glossosomatidae, Goeridae, Apataniidae, Thremmatidae, Drusinae, Neophylacinae; поденки сем. Ephemerellidae; двукрылые сем. Blephariceridae). Главным образом панцирные формы. Таксономически наиболее разнообразная группа, разные таксоны которой сменяют друг друга в разных регионах. Наиболее массовы в небольших водотоках без заиления, на умеренном течении воды; ползающий альгофаг-соскребатель (поденки сем. Heptageniidae, частично Leptophlebiidae). Наиболее массовы в горах. В разных регионах и температурных зонах замещают друг друга разные рода и виды сем. Heptageniidae (Epeorus, Iron, Rhithrogena, Ecdyonurus, Electrogena, Heptagenia), в Ориентали их отчасти (на о. Шри-Ланка – полностью) сменяют Leptophlebiidae; плавающий альгофаг-соскребатель (мелкие поденки сем. Baetidae). Массовы почти во всех регионах, в горах и на равнинах. При умеренном течении воды преобладают виды рода Baetis (включая Rhodobaetis), на быстром течении, обычно в горах, – более уплощенные Acentrella, Baetiella, Gratia; малоподвижный червеобразный микрофаг, обычно в толще обрастаний (ручейники сем. Psychomyidae, двукрылые сем. Chironomidae). В холодноводных биотопах при отсутствии эффективных хищников хирономиды могут жить и на пленочных обрастаниях (Diamesa), в крупных водотоках с корковыми обрастаниями эта группа обитает в норках в толще корки (ручейники рода Psychomyia, хирономиды родов Orthocladius, Cricotopus, Eukiefferiella и др.); прикрепленный фильтратор-микрофаг (мошки сем. Simuliidae). Встречаются спорадично, подвержены резкой сезонной динамике, тяготеют к участкам с максимальными скоростями течения и нередко доминируют при скоростях течения ≥1 м/с; малоподвижный фильтратор-эврифаг, факультативно – хищник-засадчик (ручейники сем. Hydropsychidae, Polycentropodidae, Philopotamidae, Brachycentridae). Массовый элемент почти всех сообществ ритрали (кроме некоторых арктических), обитает в широком диапазоне температуры, сапробности, размера водотока и скорости течения. В Палеарктике наиболее распространен род Hydropsyche, в Ориентали широко сочетаются многие роды, особенно из сем. Hydropsychidae и Philopotamidae; ползающий щелевой детритофаг-разгрызатель (веснянки сем. Nemouridae, Leuctridae; поденки сем. Leptophlebiidae; бокоплавы сем. Gammaridae). Второстепенная для ритрали группа, населяющая щели под камнями, где скапливается приносимый детрит. Характерна для малых лесных водотоков; ползающий щелевой хищник-поисковик (ручейники сем. Rhyacophilidae; веснянки сем. Perlidae, Perlodidae; планарии сем. Dugesiidae). Более типичны для холодноводных и горных регионов, в водотоках без существенного заиления. Охотятся преимущественно на поденок и хирономид; при этом для веснянок сем. Perlodidae показана широкая эврифагия (Паньков, 2004).
Структура сообщества ритрали довольно сложна (табл. 1). При всех региональных различиях, в нем выделяется группа основных семейств и жизненных форм, доминирующих в большинстве случаев. Это малоподвижные пассивные фильтраторы (ручейники сем. Hydropsychidae, мошки Simuliidae), подвижные и малоподвижные альгофаги-соскребатели (поденки сем. Heptageniidae, Baetidae, Ephemerellidae; ручейники сем. Glossosomatidae), подвижные хищники (ручейники сем. Rhyacophilidae), а также Chironomidae (разные роды – соскребатели, детритофаги и хищники). Их роль, тем не менее, изменяется в зависимости от региона: относительное обилие сем. Hydropsychidae возрастает от Арктики к тропикам, Rhyacophilidae, напротив, более обильны в холодноводных регионах. Поденки сем. Heptageniidae наиболее массовы в горных регионах субтропического пояса, Ephemerellidae – в восточной Азии, роль Chironomidae и Simuliidae возрастает в высоких широтах и так далее.
Таблица 1.
Усредненная структура сообществ ритрали в различных регионах Евразии (доли семейств в общем метаболизме сообщества, %)
| Количественный показатель и таксон | Кольский п-ов | Московский регион | Восточные Балканы | Кавказ и Закавказье | п-ов Южный Ямал | Новосибирское Приобье | Нижнее Приамурье | Западный Тянь-Шань | Южные Гималаи | о. Шри-Ланка | Среднее |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Количество проб | 47 | 150 | 46 | 315 | 17 | 26 | 34 | 180 | 205 | 54 | 1074 |
| биомасса, г/м2 | 8.5 | 26.5 | 27.1 | 31.1 | 38.7 | 18.4 | 28.7 | 12.8 | 21.7 | 10.3 | 26.5 |
| метаболизм, мл O2/(м2 ч) | 0.7 | 3.3 | 4.6 | 4.8 | 5.7 | 2.4 | 4.0 | 1.3 | 3.5 | 2.4 | 4.4 |
| Ephemeroptera | |||||||||||
| Baetidae | 18.0 | 22.4 | 14.4 | 15.8 | 0.2 | 14.4 | 4.2 | 12.8 | 13.9 | 5.8 | 16.8 |
| Heptageniidae | 1.2 | 1.3 | 13.9 | 25.4 | 0.1 | 11.8 | 15.4 | 12.0 | 15.5 | – | 16.0 |
| Ephemerellidae | 2.3 | 1.6 | 3.0 | 0.4 | 0.1 | 12.1 | 5.1 | 0.4 | 2.0 | – | 1.7 |
| Leptophlebiidae | 0.1 | 0.4 | 0.7 | 1.2 | 0.7 | 0.8 | 0.4 | – | 2.5 | 28.4 | 2.7 |
| Odonata | |||||||||||
| Euphaeidae | – | – | 0.1 | – | – | – | – | – | 0.4 | 7.4 | 0.5 |
| Plecoptera | |||||||||||
| Perlidae | – | – | 2.7 | 12.9 | – | 0.2 | – | 0.3 | 1.6 | 1.7 | 5.0 |
| Perlodidae | 15.6 | 3.1 | 0.1 | 0.6 | 6.3 | 1.7 | 2.7 | 3.7 | – | – | 2.4 |
| Nemouridae | 1.4 | 1.6 | 2.4 | 1.1 | 2.4 | 4.9 | 0.1 | 0.9 | 0.5 | – | 1.3 |
| Trichoptera | |||||||||||
| Hydropsychidae | 3.2 | 13.1 | 12.1 | 7.5 | – | 3.2 | 3.8 | 1.6 | 18.6 | 14.7 | 10.8 |
| Rhyacophilidae | 6.6 | 7.3 | 2.9 | 1.8 | 25.7 | 10.5 | 1.3 | 4.9 | 1.2 | – | 4.2 |
| Glossosomatidae | 0.3 | 2.3 | 4.9 | 3.0 | 5.1 | – | 11.9 | 7.3 | 5.5 | 3.4 | 4.9 |
| Limnephilidae | 1.9 | 4.1 | 2.7 | 1.4 | 5.7 | 2.9 | 2.0 | 0.3 | 0.1 | – | 1.7 |
| Uenoidae | – | – | – | – | – | – | 5.3 | – | 1.3 | – | 0.5 |
| Apataniidae | 2.4 | – | – | 0.1 | – | – | 4.3 | 0.3 | 0.3 | – | 0.4 |
| Coleoptera | |||||||||||
| Psephenidae | – | – | – | – | – | – | – | – | 0.9 | 4.4 | 0.4 |
| Diptera | |||||||||||
| Chironomidae | 11.7 | 5.9 | 5.2 | 9.1 | 3.3 | 4.3 | 12.1 | 25.9 | 4.3 | 1.5 | 11.4 |
| Simuliidae | 20.1 | 6.6 | 8.2 | 8.2 | 41.4 | 17.0 | 13.5 | 8.7 | 11.5 | 9.2 | 11.6 |
| Blephariceridae | – | – | 2.5 | 1.6 | – | – | – | 3.5 | 1.0 | – | 1.5 |
| Malacostraca | |||||||||||
| Gammaridae | – | 0.1 | 7.7 | 2.3 | – | 2.9 | 4.7 | 1.2 | – | – | 1.6 |
| Gecarcinulidae | – | – | – | – | – | – | – | – | – | 5.9 | 0.3 |
| Gastropoda | |||||||||||
| Ancylidae | – | 8.8 | 4.1 | 1.2 | – | – | – | – | – | – | 2.0 |
Примечание. Здесь и в табл. 2–5 приведены семейства, достигающие ≥3% суммарного метаболизма в одном из регионов, “–” – отсутствие семейства.
Распространение еще ряда семейств в ритрали спорадично и зависит от регионального климата и географического положения. Так, сем. Blephariceridae и Perlidae отсутствуют в северных постгляциальных регионах, сем. Ancylidae характерно только для Западной Палеарктики, сем. Gammaridae типично для гор субтропического пояса; целый ряд семейств появляется только в тропиках. Поденки сем. Leptophlebiidae встречаются по всей Евразии, но только в Южной Азии (особенно на о. Шри-Ланка) появляются среди доминантов ритрали, замещая целый ряд других таксонов (в первую очередь Heptageniidae).
К характерным второстепенным семействам, обычно не входящим в комплекс доминантов, относятся: веснянки Leuctridae, Nemouridae, Perlodidae, ручейники Polycentropodidae, Psychomyidae, Philopotamidae, Goeridae, Apataniidae, Limnephilidae, жуки Elmidae, мухи Athericidae, планарии Dugesiidae.
В целом региональная изменчивость таксономической структуры сообществ ритрали весьма высока, и в северных, и в южных аберрантных регионах (например, п-ов Ямал и о. Шри-Ланка) на уровне семейств имеет немного общего, сохраняя сходство на уровне жизненных форм. С другой стороны, горные регионы умеренного и субтропического поясов континентальной Евразии (Кавказ, Балканы, Тянь-Шань, Гималаи) характеризуются большим структурным сходством при наиболее высоком таксономическом разнообразии и могут считаться эталонными для ритральных сообществ.
Суммарное обилие (биомасса и метаболизм) макробентоса в ритрали невелики (табл. 1), подвержено умеренным региональным вариациям и не образует заметного широтного тренда. Максимальные региональные значения (биомасса 38.7 г/м2) наблюдаются на арктическом п-ове Ямал, несмотря на явно обедненную структуру сообществ, и связаны с огромной плотностью личинок Simuliidae. В большинстве регионов биомасса варьирует от 20 до 30 г/м2, в некоторых регионах – несколько меньше (вероятно, в связи с меньшей продуктивностью наземных экосистем). Метаболизм и биомасса четко связаны между собой, что говорит об устойчивой размерной структуре ритральных сообществ (средняя масса тела особи находится в пределах 0.01–0.02 г). Кроме того, существует высокая изменчивость обилия внутри каждого региона.
Сообщества пелали (мягких грунтов)
Комплекс сообществ мягких грунтов (ил различного состава, в разной степени заиленные песок, гравий, детрит, глина) с преобладанием в макрофауне роющих организмов. Объединяет пелофильные, псаммофильные и аргиллофильные биоценозы в понимании Жадина (1940). Формально класс сообществ пелали близок к потамали в понимании Иллиеса (Illies, 1961), но на равнинах встречается не только в крупных реках, но и в водотоках любых размеров и термического режима. Тем не менее, в целом пелаль наиболее развита в крупных равнинных реках и крупных стоячих водоемах, в малых водотоках формирует таксономически обедненные и, видимо, сезонные варианты, перемежающиеся с сообществами других классов. В горах редка и спорадична.
Мягкие грунты в водотоках встречаются при различных скоростях течения, вплоть до его полного отсутствия. Как правило, беспозвоночные пелали находятся под сильным прессом выедания рыбами (Gilliam et al., 1989), основной защитой от рыб служит закапывание в грунт (Persson, Swensson, 2006). В сообществах пелали преобладает роющая инфауна. В толще илисто-песчаного грунта остро встает проблема обеспечения кислородом, которая заставляет бентосные организмы развивать специальные приспособления (промываемые водой трубчатые домики, гемоглобин в гемолимфе и т.п.), но в целом ограничивает глубину проникновения бентоса в грунт (Persson, Swensson, 2006). Поскольку более заиленные грунты богаче пригодными для питания макробеспозвоночных органическими частицами, но бедны кислородом, бентосные организмы концентрируются у поверхности грунта во избежание замора. Промываемые песчаные грунты, напротив, способствуют более равномерному вертикальному распределению макробеспозвоночных в толще грунта при небольшой общей плотности.
Состав основных жизненных форм: роющий панцирный фильтратор (двустворчатые моллюски сем. Sphaeriidae, Pisidiidae). Шаровки и горошинки; избегая выедания рыбами, полностью зарываются в грунт. Кроме типичной фильтрации воды, способны собирать органические частицы с поверхности грунта. Населяют практически все водотоки, кроме горных, характерны для регионов Северной Палеарктики, в тропиках и субтропиках редки; малоподвижный панцирный фильтратор (крупные двустворчатые моллюски Unionidae, Margaritiferidae, Cyrenidae). Имея мощную раковину, мало подвержены выеданию хищниками и слабо зарываются в грунт. Чувствительны к недостатку кислорода в воде, медленно расселяются и распространены спорадично, в основном в более крупных и чистых реках. Известны для большинства регионов, но довольно редки, особенно в тропиках. При благоприятных условиях накапливают огромную биомассу и становятся сверхдоминантами сообществ; червеобразный роющий детритофаг (олигохеты сем. Tubificidae, Lumbriculidae, двукрылые сем. Chironomidae). Наиболее распространенная и таксономически разнообразная группа, особенно за счет многочисленных родов и видов хирономид. Массовы в илисто-песчаных грунтах равнинных водотоков, в гравийно-галечных биотопах занимают подчиненное положение; роющий детритофаг (поденки сем. Ephemeridae, ручейники сем. Sericostomatidae, Odontoceridae). Относительно крупные формы, способные раскапывать песок и гравий. Более типичны для биотопов с существенным течением и грубозернистыми грунтами, в горах иногда доминируют; полностью отсутствуют в некоторых регионах; ползающий панцирный эврифаг (брюхоногие моллюски сем. Viviparidae, Bithyniidae, Lithoglyphidae, Valvatidae, Thiaridae; ручейники сем. Molannidae, поденки сем. Caenidae). По-видимому, в основном, защищены от выедания рыбами и обычно не зарываются в грунт, формируя эпифауну пелали. Распространены спорадично, главным образом, в крупных равнинных водотоках с илистым дном, где нередко достигают огромного обилия (в Палеарктике – за счет крупных гастропод Viviparus и Lithoglyphus); роющий хищник (двукрылые сем. Tabanidae, Limoniidae, Pediciidae; вислокрылки сем. Sialidae). Охотятся (в основном на олигохет и личинок хирономид) в толще грунта (вислокрылки – в поверхностном наилке). Типичны для всех регионов, везде немногочисленны; малоподвижный хищник (стрекозы сем. Gomphidae, Cordulegasteridae). Как правило, подстерегают более мелких и подвижных животных, зарываясь в грунт. Редки и малочисленны в северных регионах, характерны для тропиков.
Наиболее типичные доминирующие таксоны (табл. 2): комары сем. Chironomidae, олигохеты сем. Tubificidae и двустворки сем. Pisidiidae (включая Euglesinae). В равнинных регионах, где преобладают илисто-песчаные биотопы, высоки доля и разнообразие двустворчатых моллюсков (в том числе, появляются крупноразмерные Unionidae). В горах, где преобладают галечно-гравийные биотопы, возрастает роль роющих ручейников и стрекоз. Семейство Leptophlebiidae, как и в ритрали, входит в комплекс доминантов лишь на о. Шри-Ланка.
Таблица 2.
Усредненная структура сообществ пелали в различных регионах Евразии (доли семейств в общем метаболизме сообщества, %)
| Количественный показатель и таксон | Кольский п-ов | Московский регион | Восточные Балканы | Кавказ и Закавказье | п-ов Южный Ямал | Новосибирское Приобье | Нижнее Приамурье | Западный Тянь-Шань | Южные Гималаи | о. Шри-Ланка | Среднее |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Количество проб | 17 | 122 | 16 | 132 | 14 | 12 | 10 | 51 | 76 | 32 | 482 |
| Биомасса, г/м2 | 5.2 | 76.9 | 10.5 | 24.1 | 9.1 | 8.2 | 17.9 | 9.3 | 7.7 | 13.8 | 37.9 |
| Метаболизм, мл O2/(м2 ч) |
0.4 | 5.5 | 1.8 | 2.7 | 0.7 | 1.7 | 2.8 | 1.2 | 1.3 | 3.1 | 3.6 |
| Ephemeroptera | |||||||||||
| Ephemeridae | 7.8 | 6.6 | 12.8 | 7.1 | – | 12.0 | – | – | 6.9 | 7.5 | 7.7 |
| Baetidae | 3.7 | 2.9 | 2.2 | 0.5 | – | 2.6 | – | 1.0 | 1.7 | 1.6 | 2.0 |
| Leptophlebiidae | 0.2 | 0.3 | 0.1 | 1.3 | – | – | – | – | 0.7 | 18.8 | 2.2 |
| Caenidae | 0.1 | 0.6 | 3.9 | 0.6 | – | 1.3 | 0.1 | – | 3.6 | 0.4 | 1.3 |
| Odonata | |||||||||||
| Gomphidae | – | 1.2 | 6.2 | 5.1 | – | – | – | – | 11.1 | 4.5 | 4.9 |
| Cordulegasteridae | – | – | 0.1 | 4.0 | – | – | – | – | – | – | 1.4 |
| Trichoptera | |||||||||||
| Sericostomatidae | 13.8 | 0.4 | 8.1 | 26.6 | – | – | – | – | – | – | 10.1 |
| Odontoceridae | – | – | 9.0 | – | – | – | – | – | 3.6 | 30.1 | 3.6 |
| Megaloptera | |||||||||||
| Sialidae | 4.0 | 0.7 | – | 0.1 | 3.0 | 0.6 | 5.1 | – | – | – | 0.7 |
| Diptera | |||||||||||
| Chironomidae | 23.1 | 24.1 | 26.4 | 12.2 | 16.6 | 30.2 | 69.2 | 64.6 | 26.6 | 2.7 | 31.0 |
| Ceratopogonidae | 0.1 | 1.2 | 0.1 | 0.1 | 0.8 | 0.6 | – | 0.1 | 3.9 | 0.5 | 1.3 |
| Limoniidae | 6.9 | 4.7 | 1.8 | 2.1 | 2.9 | 6.1 | – | 3.0 | 7.6 | 4.1 | 5.1 |
| Pediciidae | 1.4 | 1.3 | 3.1 | 0.3 | – | 9.8 | 1.8 | 0.2 | – | – | 1.1 |
| Tabanidae | – | 2.3 | 0.7 | 2.6 | – | – | – | 0.6 | 9.5 | – | 3.6 |
| Bivalvia | |||||||||||
| Unionidae | – | 4.4 | – | 0.7 | – | – | – | – | – | – | 1.6 |
| Sphaeriidae | – | 3.0 | 0.5 | – | 24.0 | 6.6 | – | – | – | – | 2.0 |
| Pisidiidae | 10.8 | 17.5 | 3.5 | 5.9 | 31.0 | 17.1 | 5.4 | 1.9 | 0.1 | 1.4 | 10.3 |
| Oligochaeta | |||||||||||
| Tubificidae | 11.8 | 6.9 | 6.5 | 9.3 | 10.3 | 3.5 | 10.2 | 11.3 | 1.4 | 5.0 | 9.0 |
Суммарный метаболизм сообществ пелали (табл. 2) немного ниже, чем ритрали (в среднем 3.6 против 4.4 мл O2/м2 ч), биомасса – напротив, выше, что свидетельствует о преобладании более крупных организмов. Этот эффект достигается за счет равнинного Московского региона с огромным обилием крупных двустворчатых моллюсков в некоторых пелальных сообществах при средних биомассе 76.9 г/м2 и массе тела 0.995 г. Высокая биомасса сообществ Кавказа формируется за счет большой доли крупных роющих ручейников сем. Sericostomatidae. В большинстве других регионов в пелали преобладают мелкие формы, биомасса сообществ обычно 10–20 г/м2 при средней массе тела макробентосных организмов 0.005–0.02 г.
Сообщества фитали (зарослей макрофитов)
Класс сообществ, развивающихся в густых зарослях водных растений (макрофитов), главным образом цветковых с погруженными и плавающими листьями (рдестовые, кувшинковые, водокрасовые, рясковые и т.п.), на замедленном течении или без течения. Сходен с фитофильным биоценозом в понимании Жадина (1940). Беспозвоночные этого класса сообществ не имеют явных адаптаций к течению и часто встречаются в стоячих водоемах. На быстром течении рек и ручьев макрофиты также иногда растут, но там их населяют ритральные или кренальные сообщества.
Фиталь в водотоках встречается спорадично, как и сами заросли макрофитов. В умеренной и субарктической зонах фитальные сообщества, как правило, сезонные (летние), на зиму деградируют вместе с зарослями макрофитов. В тропиках зарослевым сообществам присущи более плавные циклы, связанные с муссонами и засухами, но и там они обычно сезонные. В арктической зоне и в горах фиталь почти не развиваета, в основном из-за недостаточной летней температуры воды. По нашим данным, все сообщества фитали встречены в водотоках с летней температурой воды ≥18°С. Как результат – из десяти обследованных нами регионов только в трех удалось собрать достаточное число описаний фитальных сообществ.
Состав основных жизненных форм: малоподвижный панцирный фито/альго/детритофаг (брюхоногие моллюски сем. Lymnaeidae, Planorbidae, Physidae, Bithyniidae, Valvatidae и др., а также жуки-листоеды Chrysomelidae). Это доминирующая группа во всех полноценных сообществах фитали. В Палеарктике обычно преобладают прудовики родов Lymnaea и Radix, в Ориентали усиливается роль жаберных гастропод. В биотопах плавающих листьев макрофитов (например, кубышек Numphar) могут преобладать личинки листоедов Galerucella; плавающий альгофаг (поденки сем. Baetidae, жуки сем. Haliplidae, олигохеты сем. Naididae, хирономиды). В целом мелкие формы, встречаются в большинстве регионов и проб, обычно с невысоким обилием; малоподвижный хищник-засадчик (стрекозы сем. Coenagrionidae, Calopterygidae, Libellulidae; клопы сем. Nepidae). Обычно лазают по растениям, охотятся на поденок, хирономид и планктонных ракообразных. При большом количестве зоопланктона могут достигать высокого обилия и изредка доминируют по метаболизму в сообществе макрофауны; плавающий хищник-поисковик (жуки сем. Dytiscidae, Noteridae; клопы сем. Naucoridae, Notonectidae, Pleidae). Также охотятся на мелких плавающих животных. Обычно малочисленные в реках, тяготеют к биотопам без заметного течения.
Доминирующая группа сообществ фитали – брюхоногие моллюски, представленные здесь большим числом семейств и родов. В палеарктических регионах по биомассе и метаболизму преобладают прудовики (Lymnaeidae), битинии (Bithyniidae) и катушки (Planorbidae), в Ориентали к ним добавляются Thiaridae. В тропической зоне резко возрастает роль креветок (Palaemonidae, Atyidae) и разнокрылых стрекоз (Libellulidae), роль моллюсков несколько снижается.
Суммарное обилие (табл. 3) заметно выше, чем в ритрали и пелали, но сильно варьирует по регионам. Максимальные показатели (биомасса 77.7 г/м2) выявлены в Московском регионе с большим количеством летних проб из крупных равнинных водотоков. Заметим, что пелаль в этом регионе также развита максимально и показатели биомассы близкие. В регионах Приамурья и Гималаев (возможно, из-за более частых и мощных летних паводков) обилие сообществ фитали гораздо меньше и не превышает средних значений для других классов сообществ. Во всех регионах сообщество составляют довольно крупные организмы, средняя масса тела 0.07–0.15 г.
Таблица 3.
Усредненная структура сообществ фитали в различных регионах Евразии (доли семейств в общем метаболизме сообщества, %)
| Количественный показатель и таксон | Московский регион | Нижнее Приамурье | Южные Гималаи | Среднее |
|---|---|---|---|---|
| Количество проб | 44 | 22 | 10 | 76 |
| Биомасса, г/м2 | 77.7 | 9.9 | 32.3 | 58.0 |
| Метаболизм, мл O2/(м2 ч) | 12.5 | 2.2 | 6.1 | 9.7 |
| Ephemeroptera | ||||
| Baetidae | 9.9 | 8.9 | 1.8 | 9.5 |
| Odonata | ||||
| Libellulidae | – | – | 14.2 | 2.1 |
| Coenagrionidae | 1.4 | 15.0 | 8.3 | 7.0 |
| Heteroptera | ||||
| Nepidae | 2.1 | 6.5 | 0.2 | 3.5 |
| Corixidae | 0.4 | 4.3 | 1.9 | 1.9 |
| Coleoptera | ||||
| Chrysomelidae | 3.2 | 3.6 | – | 3.2 |
| Diptera | ||||
| Chironomidae | 0.7 | 3.1 | 0.8 | 1.6 |
| Malacostraca | ||||
| Asellidae | 1.1 | 3.7 | – | 1.9 |
| Atyidae | – | – | 15.2 | 2.2 |
| Palaemonidae | – | – | 3.2 | 0.5 |
| Gastropoda | ||||
| Lymnaeidae | 43.4 | 24.4 | 21.1 | 39.0 |
| Planorbidae | 3.9 | 9.7 | 16.9 | 8.1 |
| Physidae | 4.1 | – | 0.7 | 2.7 |
| Bithyniidae | 8.1 | 9.1 | 2.5 | 8.5 |
| Valvatidae | 4.6 | 1.0 | – | 3.3 |
| Thiaridae | – | – | 5.9 | 0.9 |
Сообщества кренали (ручьев и родников)
Класс сообществ, связанных со спецификой малых водотоков – родников и ручьев с меженным водорасходом ≤0.01 м3/с и шириной русла ≤1 м, с преобладанием мелкомозаичных, в большой степени органических субстратов. Это близко к кренали в понимании Иллиеса (Illies, 1961), но без ограничений по температурным условиям. Малый размер водотока с высокой степенью вероятности определяет целый комплекс абиотических факторов, в совокупности создающих специфику местообитаний кренали. Это малая глубина водотока (обычно 0.01–0.1 м) и малая скорость течения (≤0.1 м/с, даже при значительном уклоне русла); сложный состав донных субстратов с большой долей грубодисперсного детрита наземного происхождения, главным образом, листового опада, служащего основным источником питания беспозвоночных; затенение русла окружающей наземной растительностью; слабое развитие собственных продуцентов (макрофитов и микроводорослей в бентосе и планктоне); пониженная и относительно стабильная температура воды; почти полное отсутствие рыб (Hynes, 1970; Illies, 1961). Гидрологический режим водотока варьирует в зависимости от основного источника питания – от почти стабильного (в родниках) до полного сезонного высыхания водотока (в ручьях со снеговым питанием).
Точной границы размера водотока, при которой кренальные сообщества сменяются другими классами, установить нельзя. Она сдвигается вверх по ручью при наличии на нем запруд или озер, при отсутствии растительности по берегам, протекании по незадернованному аллювиальному грунту и ряду других причин. С другой стороны, кренальные сообщества проникают в более крупные водотоки вдоль береговой кромки и по скоплениям листового опада и детрита в заводях. Соответственно, существует переходная зона водотоков, где на одних субстратах развиты сообщества кренали, на других, рядом с ними, – сообщества других классов (обычно ритрали или пелали).
Полный комплекс кренальной специфики абиотических условий (и, соответственно, биоты) характерен для равнинной лесной умеренной зоны, где ручьи резко отличаются от рек по летней температуре, освещенности и преобладающим донным субстратам. В тропиках специфика кренали отчасти сглаживается за счет выравнивания температуры в разных водотоках, а в безлесных зонах – за счет уменьшения значимости листового опада в питании беспозвоночных. В арктическом тундровом Ямале типичные сообщества кренали не развиты.
Состав основных жизненных форм: ползающий щелевой детритофаг-разгрызатель или эврифаг (веснянки сем. Nemouridae, Leuctridae, поденки сем. Leptophlebiidae, жуки сем. Scirtidae, бокоплавы сем. Gammaridae, ряд Chironomidae). Наиболее распространенная группа, характерная для всех регионов. Основные таксоны в ее пределах сменяются: в Северной Палеарктике (до умеренной зоны) обычно преобладают Nemouridae и другие веснянки, в Южной Палеарктике (от Средиземноморья до Приамурья) – бокоплавы, в тропиках Ориентали – поденки сем. Leptophlebiidae; малоподвижный панцирный детритофаг-разгрызатель (ручейники сем. Limnephilidae, Lepidostomatidae, Calamoceratidae). Вторая массовая жизненная форма кренали, в пределах которой также наблюдается географическое замещение: в Северной Палеарктике преобладают Limnephilidae (роды Stenophylax, Potamophylax, Chaetopteryx и др.), в Южной Палеарктике представители Lepidostomatidae и Calamoceratidae дополняют их, а в Ориентальной области – полностью сменяют; плавающий альгофаг (поденки сем. Baetidae, Siphlonuridae). Обычно второстепенная группа, включает реофильные таксоны (роды Baetis, Nigrobaetis), усиливающие позиции при ускорении течения, и почти лимнофильные, населяющие микрозаводи без течения (Centroptilum, Siphlonurus, Metreletus); ползающий щелевой хищник-поисковик (веснянки сем. Perlodidae; стрекозы сем. Gomphidae, Cordulegasteridae, Corduliidae; крабы сем. Potamidae, Gecarcinulidae; некоторые Limoniidae и Tipulidae (Diptera)). Второстепенная группа, существенно увеличивающая присутствие в Ориентальной области за счет крабов и стрекоз. Вероятно, многие кренальные крабы – эврифаги, активно потребляющие детрит.
В сообществах кренали (табл. 4) доминируют, как правило, детритофаги-разгрызатели или заменяющие их эврифаги. Таксономическая структура сообществ существенно различается в разных регионах Евразии. Обследованные нами регионы образуют три контрастные группы. В постгляциальных регионах северной части Палеарктики (Кольский п-ов, Московский регион) доминируют по обилию веснянки сем. Nemouridae (роды Nemoura, Nemurella, Amphinemura), ручейники сем. Limnephilidae, а также Chironomidae. В южной части Палеарктики их роль уменьшается, преобладают, главным образом, бокоплавы сем. Gammaridae (Gammarus), местами (в родниках и подземных карстовых водотоках) также сем. Niphargidae (Niphargus). В тропических регионах Южной Азии все эти группы исчезают, их замещают представители семейств: поденки сем. Leptophlebiidae, ручейники сем. Lepidostomatidae (Goerodes) и Calamoceratidae (Anisocentropus), стрекозы сем. Gomphidae, крабы сем. Potamidae и Gecarcinulidae, причем состав доминантов в Гималаях и на о. Шри-Ланка существенно различается. Кроме того, ряд семейств, играя в сообществах кренали второстепенную роль, встречается в большинстве регионов. Это поденки сем. Baetidae (Baetis, Centroptilum) и Heptageniidae (Ecdyonurus, Electrogena), ручейники сем. Polycentropodidae (Plectrocnemia), двустворки сем. Pisidiidae; в пределах Палеарктики – веснянки сем. Perlodidae (Isoperla), жуки сем. Scirtidae и Hydraenidae, двукрылые сем. Tipulidae (Tipula), Simuliidae (Cnetha) и другие.
Таблица 4.
Усредненная структура сообществ кренали в различных регионах Евразии (доли семейств в общем метаболизме сообщества, %)
| Количественный показатель и таксон | Кольский п-ов | Московский регион | Восточные Балканы | Кавказ и Закавказье | Новосибирское Приобье | Нижнее Приамурье | Западный Тянь-Шань | Южные Гималаи | о. Шри-Ланка | Среднее |
|---|---|---|---|---|---|---|---|---|---|---|
| Количество проб | 15 | 79 | 84 | 406 | 37 | 31 | 73 | 95 | 39 | 859 |
| Биомасса, г/м2 | 2.6 | 17.5 | 23.6 | 28.0 | 17.2 | 10.0 | 19.0 | 8.9 | 11.8 | 26.4 |
| Метаболизм, мл O2/(м2 ч) | 0.1 | 1.8 | 2.1 | 3.8 | 2.2 | 1.2 | 2.4 | 0.9 | 2.1 | 2.7 |
| Ephemeroptera | ||||||||||
| Leptophlebiidae | 2.2 | 3.3 | 2.1 | 3.9 | 0.6 | 0.6 | – | 3.9 | 16.5 | 4.5 |
| Baetidae | 0.8 | 6.7 | 1.3 | 1.9 | 7.5 | 0.5 | 4.5 | 5.6 | 4.3 | 3.9 |
| Heptageniidae | – | 0.2 | 1.9 | 1.9 | 0.9 | 2.2 | 1.2 | 8.6 | – | 2.8 |
| Odonata | ||||||||||
| Gomphidae | – | – | – | – | – | – | – | 0.6 | 8.1 | 0.5 |
| Plecoptera | ||||||||||
| Perlodidae | 5.1 | 1.0 | 0.6 | 0.9 | 0.1 | 0.4 | 0.8 | – | – | 0.9 |
| Nemouridae | 19.3 | 13.1 | 9.1 | 3.3 | 6.4 | 1.7 | 4.6 | 4.6 | – | 6.4 |
| Trichoptera | ||||||||||
| Polycentropodidae | 2.4 | 1.6 | 2.8 | 1.0 | – | – | 0.2 | 3.0 | 5.4 | 1.9 |
| Limnephilidae | 8.7 | 36.1 | 7.5 | 8.1 | 6.2 | 1.6 | 1.8 | 2.9 | – | 10.9 |
| Lepidostomatidae | – | 0.5 | – | 0.3 | – | 0.2 | 1.5 | 14.9 | 12.1 | 3.1 |
| Apataniidae | 0.7 | – | – | 0.3 | – | 0.5 | 4.8 | 0.6 | – | 0.8 |
| Calamoceratidae | – | – | – | – | – | – | – | 2.2 | 4.3 | 0.5 |
| Diptera | ||||||||||
| Chironomidae | 42.2 | 5.1 | 1.0 | 1.6 | 2.3 | 3.1 | 9.7 | 3.1 | 2.5 | 4.4 |
| Simuliidae | 2.8 | 2.1 | 0.5 | 0.7 | 4.9 | 0.7 | 0.4 | 0.8 | 0.4 | 1.2 |
| Tipulidae | 0.2 | 1.2 | 2.7 | 0.2 | 1.4 | – | 4.3 | 4.3 | – | 1.7 |
| Malacostraca | ||||||||||
| Gammaridae | – | 0.1 | 44.7 | 45.2 | 52.6 | 80.9 | 45.1 | – | – | 42.8 |
| Niphargidae | – | – | 7.1 | 7.9 | – | – | – | – | – | 5.4 |
| Gecarcinulidae | – | – | – | – | – | – | – | – | 12.8 | 0.7 |
| Gastropoda | ||||||||||
| Lymnaeidae | – | 3.7 | 0.1 | 1.2 | – | – | 0.1 | 0.1 | – | 1.2 |
| Bivalvia | ||||||||||
| Pisidiidae | 0.1 | 1.9 | 1.3 | 3.5 | 1.3 | 0.1 | 0.1 | 0.2 | 1.6 | 2.6 |
| Turbellaria | ||||||||||
| Planariidae | 0.3 | – | – | – | – | 1.3 | 7.6 | 0.9 | – | 1.0 |
Суммарное обилие макробентоса (табл. 4) невелико, примерно на уровне ритрали (биомасса 15–30 г/м2, метаболизм 1–3 мл O2/м2 ч), и несколько повышено в горных регионах Южной Палеарктики за счет сообществ с большой плотностью бокоплавов рода Gammarus. Очень низко обилие в арктическом Кольском регионе. Средняя масса тела макробентосных организмов обычно 0.015–0.030 г, в регионах с доминированием бокоплавов несколько выше, чем в прочих.
Сообщества рипали (субстратов береговой кромки)
Класс сообществ, связанных с мозаичными, главным образом, органическими субстратами наземного происхождения вдоль береговой кромки водотоков, обычно на расстоянии ≤0.5 м от берега. Термин “рипаль” введен Семеновым-Тянь-Шанским (1937) для обозначения соответствующей гидрологической зоны рек, в наших работах мы рассматриваем его как биотоп отдельного класса сообществ. Обитание в рипали не требует специальных адаптаций и доступно для наиболее эврибионтных таксонов всех других классов сообществ.
Класс рипальных сообществ имеет некоторые черты сходства с сообществами кренали: он также формируется на неоднородных субстратах с существенной органической составляющей и также включает наиболее эврибионтные элементы многих других классов сообществ. Однако, в рипали нет преобладания детритофагов-разгрызателей и нет сдвига в сторону преобладания холодноводных форм. Доминанты сообществ рипали – главным образом, смесь эврибионтов, присущих также сообществам ритрали, фитали и пелали.
Класс сообществ рипали можно было бы считать “сорным” и не рассматривать вообще, но во многих бедных субстратами водотоках именно эти сообщества определяют большую часть таксономического разнообразия макробентоса (хотя и занимает ничтожно малую часть площади дна). Так, сообщества многих равнинных рек (особенно северных, в которых плохо развиваются макрофиты) представлены только классами пелали и рипали; во многих горных реках развиты только сообщества ритрали и рипали. В первом случае рипаль – единственное местообитание ритральных и фитальных форм, во втором – фитальных и пелальных. Поэтому при оценке общей продуктивности водотока игнорировать сообщества рипали можно, но при изучении биоразнообразия они очень важны. Оценку индексов качества воды (например, индекса сапробности) в реках также, по нашему мнению, лучше проводить по сообществам рипали – они близки к оценкам, получаемым при изучении участка водотока в целом. Сообщества рипали встречаются во всех регионах и типах ландшафта, хотя на равнинах чаще, чем в горах.
Состав основных жизненных форм: плавающий альгофаг (поденки сем. Baetidae, Ameletidae, жуки сем. Haliplidae). Повсеместно распространенная и весьма разнообразная группа, таксоны которой отчасти сменяют друг друга в разных регионах. Так, в рипали встречается большинство евразийских родов и подродов сем. Baetidae (роды Baetis, Nigrobaetis, Labiobaetis, Procloeon, Centroptilum, Similicloeon) и других плавающих поденок (роды Ameletus, Metretopus, Siphlonurus, Isonychia); плавающий эврифаг (клопы-гребляки сем. Corixidae, наиболее часто – из рода Micronecta). Встречаются почти во всех регионах, часто бывают очень многочисленны, особенно на заиленных отмелях; малоподвижный панцирный детритофаг-разгрызатель (ручейники сем. Limnephilidae, Phryganeidae, Leptoceridae). Группа, общая с сообществами кренали, но обычно включает более теплолюбивые таксоны (в Палеарктике – роды Limnephilus, Anabolia, Halesus, Semblis, Leptocerus и др.); ползающий детритофаг-разгрызатель или эврифаг (веснянки сем. Nemouridae, поденки сем. Leptophlebiidae, бокоплавы сем. Gammaridae). Как и в кренали, распространяется, главным образом, по грубодетритным субстратам и иногда доминирует за счет массового развития бокоплавов; малоподвижный панцирный эврифаг (брюхоногие моллюски сем. Viviparidae, Bithyniidae, Thiaridae, Pachychilidae, Paludomidae). Спорадично распространенные таксоны, местами достигающие огромного обилия и сверхдоминирования в сообществе. Считаются детритофагами-собирателями, но у некоторых форм отмечена также высокая эффективность фильтрации с помощью наружных жабр (Hӧckelmann, Pusch, 2000). Разные семейства замещают друг друга в разных регионах: живородки и битинии больше характерны для Палеарктики, Thiaridae и Pachychilidae – для Ориентальной области, Paludomidae – в основном для о. Шри-Ланка; все они избегают высокоширотных регионов; прикрепленный фильтратор-альгофаг (мошки Simuliidae). В основном ритральная группа, в рипали встречается на участках с усиленным течением; малоподвижный хищник-засадчик (стрекозы сем. Calopterygidae, Plathycnemididae, Libellulidae, Corduliidae). Группа, общая с сообществами фитали, но достигающая максимального обилия и роли в сообществе именно в рипали. Большинство стрекоз в реках – преимущественно рипальная группа; в пределах Евразии их разнообразие возрастает к югу; плавающий хищник-поисковик (жуки сем. Dytiscidae). Второстепенная группа, общая с классом фитали. Для Палеарктики наиболее типичны роды Laccophilus, Platambus, Oreodytes; для Ориентальной области – Hydrovatus, Neptosternus и Laccophilus; роющий панцирный фильтратор (двустворчатые моллюски Unionidae, Sphaeriidae, Pisidiidae). Характерны для пелали, но нередко достигают максимума обилия именно в рипали – на береговых склонах и прибрежных желобах, где усиливается осаждение взвеси и оптимизированы условия для фильтрации.
Потенциальный спектр жизненных форм рипали обширен и в основном пересекается с жизненными формами других сообществ. Лишь несколько семейств (в частности, поденки Ameletidae, ручейники Leptoceridae, стрекозы Corduliidae, Calopterygidae и Plathycnemididae) могут считаться характерными именно для рипали, но они обычно играют второстепенную роль.
Сообщества рипали подвержены большой изменчивости различной природы. Во-первых, четко выражена географическая изменчивость, связанная с климатом и рельефом различных регионов и выделяющая регионы северные и холодноводные (с большой долей семейств Ameletidae, Perlodidae, Limnephilidae, Simuliidae), южнопалеарктические (с высокой долей бокоплавов Gammaridae) и ориентальные (с возрастанием доли поденок Leptophlebiidae, стрекоз Libellulidae, креветок и тропических гастропод). Также сообщества рипали многих регионов подвержены спорадическому вселению крупных эврибионтных и всеядных форм высших ракообразных (бокоплавов, крабов, креветок) и жаберных моллюсков (Viviparidae, Thiaridae, Pachychilidae, Paludomidae), часто достигающих очень высокого обилия и сверхдоминирования в сообществах, с возрастанием его суммарной биомассы в 5–10 раз. Наконец, сообществам рипали, как и фитали, присуща сильная сезонная изменчивость, связанная с колебаниями водности рек и вынужденными миграциями всего сообщества вслед за урезом воды. В водотоках с особенно быстрыми колебаниями уровня воды сообщество рипали редуцируется до нескольких мелких плавающих форм (роды Micronecta, Nigrobaetis и др.).
Суммарные биомасса и метаболизм рипальных сообществ (табл. 5) в среднем несколько выше, чем во всех других классах, но подвержены очень сильной региональной изменчивости. Большие значения обилия наблюдаются в Московском регионе за счет проб с огромной плотностью крупных моллюсков сем. Viviparidae, с биомассой 100–500, иногда до 2500 г/м2. Сходный эффект дают некоторые пробы в Амурском регионе (за счет Viviparidae и Pachychilidae) и на Балканах (за счет Gammaridae). В пробах и регионах, не подверженных оккупации жаберных моллюсков и высших ракообразных, обилие находится на среднем уровне (биомасса 10–20 г/м2, метаболизм 3–6 мл O2/м2 ч).
Таблица 5.
Усредненная структура сообществ рипали в различных регионах Евразии (доли семейств в общем метаболизме сообщества, %)
| Количественный показатель и таксон | Кольский п-ов | Московский регион | Восточные Балканы | Кавказ и Закавказье | п-ов Южный Ямал | Новосибирское Приобье | Нижнее Приамурье | Западный Тянь-Шань | Южные Гималаи | о. Шри-Ланка | Сред-нее |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Число проб | 15 | 134 | 12 | 76 | 20 | 10 | 26 | 52 | 67 | 31 | 443 |
| Биомасса, г/м2 | 4.7 | 124.8 | 31.6 | 21.0 | 20.5 | 12.6 | 31.8 | 10.8 | 21.7 | 21.8 | 66.4 |
| Метаболизм, мл O2/(м2 ч) | 0.3 | 9.5 | 5.2 | 3.2 | 1.8 | 2.1 | 2.5 | 1.1 | 2.3 | 4.1 | 6.1 |
| Ephemeroptera | |||||||||||
| Ameletidae | 31.1 | – | – | – | 0.1 | – | – | 33.5 | 2.3 | – | 6.9 |
| Baetidae | 10.0 | 13.5 | 7.2 | 15.2 | 2.0 | 22.1 | 17.5 | 13.6 | 12.2 | 8.9 | 16.7 |
| Heptageniidae | – | 1.8 | 5.6 | 2.8 | – | 2.1 | 2.7 | 1.5 | 2.3 | – | 2.5 |
| Ephemerellidae | 7.5 | 0.5 | 4.6 | 7.1 | – | 0.9 | 4.2 | 0.2 | – | – | 2.6 |
| Leptophlebiidae | 0.3 | 3.6 | 0.3 | 1.5 | 2.4 | 0.2 | – | – | 3.2 | 11.3 | 3.5 |
| Odonata | |||||||||||
| Calopterygidae | – | 4.2 | 3.7 | 4.8 | – | – | – | – | 1.1 | 2.6 | 3.3 |
| Cordulegasteridae | – | – | 6.4 | 0.4 | – | – | – | 2.5 | – | – | 0.7 |
| Libellulidae | – | – | – | – | – | – | – | – | 12.1 | 3.9 | 2.7 |
| Plecoptera | |||||||||||
| Perlodidae | 26.8 | 0.6 | 0.1 | 0.6 | – | 1.2 | 1.4 | 3.2 | – | – | 2.2 |
| Nemouridae | 3.0 | 1.2 | 0.8 | 4.0 | 4.1 | 5.1 | 2.9 | 1.8 | 0.7 | – | 2.5 |
| Heteroptera | |||||||||||
| Corixidae | 0.7 | 2.2 | 1.9 | 0.3 | 0.6 | 3.8 | 1.3 | – | 5.9 | – | 2.4 |
| Coleoptera | |||||||||||
| Dytiscidae | 0.3 | 1.0 | 0.7 | 0.4 | 9.7 | 1.7 | 1.4 | 0.7 | 0.3 | – | 1.3 |
| Trichoptera | |||||||||||
| Limnephilidae | 5.9 | 10.5 | 0.5 | 16.5 | 32.2 | 21.1 | 8.0 | 3.8 | 0.3 | – | 11.7 |
| Diptera | |||||||||||
| Chironomidae | 2.5 | 4.6 | 2.5 | 6.7 | 4.1 | 0.6 | 4.7 | 7.9 | 2.6 | 0.4 | 5.8 |
| Simuliidae | 0.1 | 0.3 | 0.1 | 0.6 | 28.3 | 0.3 | 0.1 | 2.1 | 0.2 | 0.7 | 2.3 |
| Tipulidae | 1.3 | 0.4 | 0.6 | – | 0.5 | – | – | 5.7 | 0.3 | – | 1.2 |
| Malacostraca | |||||||||||
| Gammaridae | – | 2.8 | 29.9 | 10.8 | 1.2 | 14.7 | 7.8 | 10.8 | – | – | 7.3 |
| Asellidae | – | 2.0 | 8.0 | – | – | – | – | – | – | – | 1.1 |
| Atyidae | – | – | – | – | – | – | – | – | 11.5 | 1.7 | 2.4 |
| Gecarcinulidae | – | – | – | – | – | – | – | – | – | 8.1 | 0.7 |
| Gastropoda | |||||||||||
| Lymnaeidae | – | 4.8 | 0.5 | 0.4 | – | 3.6 | 5.5 | 1.5 | 0.1 | – | 2.8 |
| Viviparidae | – | 10.8 | – | – | – | – | 21.8 | – | 0.8 | – | 5.5 |
| Bithyniidae | – | 3.0 | – | – | – | – | 1.1 | – | 0.2 | – | 1.3 |
| Pachychilidae | – | – | – | – | – | – | 15.9 | – | – | – | 1.2 |
| Thiaridae | – | – | – | – | – | – | – | – | 18.0 | – | 3.5 |
| Paludomidae | – | – | – | – | – | – | – | – | – | 22.9 | 2.1 |
Сравнение трофической структуры сообществ разных классов
Для сопоставления трофики выделяемых классов сообществ мы объединили основные семейства макрофауны в группы по преимущественному типу питания (рис. 2). При этом могут возникать ошибки, связанные со смешанным питанием и слабой изученностью питания многих семейств, но они не должны существенно искажать общую картину. Видно, что все классы сообществ существенно различаются уже на уровне трофических групп. Наиболее контрастно выделяются классы кренали (с преобладанием детритофагов-разгрызателей) и пелали (преобладают детритофаги-собиратели, включая грунтоедов). Классы ритрали и фитали трофически сближены – в них наибольшую роль играют альгофаги-соскребатели, хотя и разных таксонов (в сообществах ритрали – главным образом, поденок и ручейников, в фитали – легочных брюхоногих моллюсков). Структура сообществ рипали отличается высокой выравненностью соотношений обилия большинства трофических групп и очень близка к средней трофической структуре реофильных сообществ (по ~25% соскребателей, разгрызателей и собирателей, ~15% хищников и ~10% фильтраторов). Этот результат согласуется с постулатами концепции речного континуума: в малых водотоках (креналь) должны преобладать детритофаги-разгрызатели, в средних – соседствуют все трофические группы, в крупных (пелаль) – доминируют детритофаги-собиратели.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Перспективы распространения предложенной классификации. Выглядит логичным расширение предлагаемой системы на другие типы водоемов, но пока для этого нет достаточных данных. Предварительные наблюдения показывают, что описанные выше классы сообществ характерны не только для текучих, но и для стоячих континентальных водоемов. Биотопы и классы сообществ пелали, фитали и рипали должны быть типичны для озер, прудов и водохранилищ. Для малых временных водоемов (луж), видимо, нужен дополнительный класс эфемерали, несколько описаний которого мы сделали ранее (Чертопруд и др., 2020б). Классы ритрали и кренали не типичны для стоячих вод, хотя могут развиваться в некоторых редких случаях. Возможно, дополнительная типизация потребуется для сообществ морского происхождения (например, каспийских) и сообществ древних озер с эндемичной фауной (как в оз. Байкал). Этот вопрос пока не решен. Еще более спорна возможность расширения нашей системы на морские сообщества, хотя и здесь прослеживаются некоторые параллели.
Внутренняя неоднородность классов сообществ и дальнейшая их типизация. Разумеется, многообразие реофильных сообществ не ограничивается наличием описанных выше классов в силу их высокой внутренней неоднородности. Так, суммарный метаболизм ритральных сообществ только в Московском регионе варьирует в 18 раз. При этом ни один вид или даже семейство не имеет 100%-ной встречаемости, но отмечены ритральные сообщества с доминированием шести различных семейств (Baetidae, Rhyacophilidae, Hydropsychidae, Chironomidae, Simuliidae и Ancylidae). Таким образом, в пределах каждого из выделенных классов существует значительная изменчивость, требующая изучения и типизации. Эта типизация проведена нами для большинства обследованных регионов (Палатов, Чертопруд, 2012, 2018; Чертопруд, Палатов, 2013, 2017; Чертопруд и др., 2020а, б; Chertoprud, 2011, 2019; Chertoprud, Palatov, 2013; Chertoprud et al., 2018) и позволила установить в пределах каждого из основных классов до 10–15 вариантов, обычно связанных с действием одного–двух специфических факторов среды. Они в основном повторяются в разных регионах, их общее число, по нашим данным, достигает 60–70. Детальный обзор разнообразия и изменчивости этих вариантов мы планируем в дальнейшем.
Альтернативные стратегии формирования реофильных сообществ. При построении классификации реофильных сообществ беспозвоночных обнаружена следующая закономерность (Чертопруд, 2011): одни типы сообществ четко соответствуют определенным донным биотопам (их биотическая классификация соответствует биотопической), другие типы не подчиняются этому правилу, проявляя высокую степень эврибионтности. Для обозначения этих двух классов использована концепция PM-биоценотического градиента, предложенная Протасовым (1989). В ее рамках выделяются два крайних варианта структуры сообществ. Сообщества P (Petersen)-типа, отвечающие определению сообществ Петерсена (Протасов, 1989, 2011), управляются преимущественно абиотическими факторами и обычно соответствуют определенным биотопам – именно в таком ключе рассматривает сообщества концепция континуализма (Раменский, 1938; Gleason, 1926). Сообщества М (Mebius)-типа, отвечающие определению К. Мебиуса (Mӧbius, 1877), обычно имеют в составе ярко выраженные виды-эдификаторы, управляются в основном биотическими связями и соответствуют концепции структурализма (Разумовский, 1981; Жирков, 2010). В реофильных сообществах Р-типа, как правило, преобладают личинки насекомых – чаще всего хирономид, поденок или ручейников; причем степень и постоянство доминирования отдельных видов невысоки. Это большинство встреченных нами сообществ во всех классах. Сообщества М-типа связаны с резким и устойчивым доминированием высших ракообразных (бокоплавов сем. Gammaridae, иногда креветок и крабов) и крупных жаберных моллюсков (обычно Unionidae, Viviparidae и др.) и встречаются в водотоках существенно реже (обычно в классах рипали и кренали).
Видовое разнообразие в сообществах Р-типа явно больше. Так, сообщества Р-типа ритрали крупных рек Московского региона включают в среднем 18 видов на пробу, а формирующиеся в сходных условиях сообщества М-типа – 12 видов (Чертопруд, 2011). По-видимому, доминанты сообществ М-типа прямо или косвенно вытесняют часть других видов. Суммарная биомасса в сообществах Р-типа обычно варьирует от 2 до 280, в среднем 23.7 г/м2; в сообществах М-типа – примерно от 100 до 2500, в среднем 845 г/м2, то есть в 35 раз больше. Средняя индивидуальная масса организмов в сообществах P-типа – 0.031 г, в сообществах М-типа – 0.590 г (почти в 20 раз больше). Таким образом, сообщества М-типа укомплектованы гораздо более крупными организмами. Доля первого доминанта по биомассе для сообществ Р-типа составляет в среднем 47%, в сообществах М-типа – 87% (так называемое сверхдоминирование). Сообщества М-типа менее чувствительны к типу донного биотопа, но они более однородны в пределах одного биотопа и менее подвержены паводковой и сезонной динамике.
Сообщества Р- и М-типов нередко могут формироваться в сходных условиях. Определяющим фактором формирования сообщества здесь служит вселение отдельных видов – доминантов сообществ М-типа (обычно каких-либо бокоплавов или моллюсков). При их наличии формируется сообщество М-типа; при их отсутствии – сообщество Р-типа. Таким образом, основную стратегию формирования сообществ Р-типа можно охарактеризовать так: быстрое заселение водоемов – узкая специализация по биотопам – высокая динамичность распределения – низкое обилие – регуляция абиотическими факторами. Сообщества М-типа имеют обратные черты: медленное расселение по водоемам – спорадичность распространения – эврибионтность внутри водоемов – большая устойчивость во времени – высокое обилие – высокая конкурентоспособность – регуляция структуры собственными доминантами.
Описанная нами закономерность, возможно, позволяет отчасти разрешить вопросы о дискретности и континуальности сообществ и о предпочтительности биотических или абиотических данных при классифицировании сообществ, широко обсуждаемые в экологии (Жирков, 2010). По-видимому, для сообществ М-типа более применимы представления о дискретности и биотическая классификация, а для сообществ Р-типа – наоборот.
В целом можно говорить о двух разных стратегиях основных макротаксонов, формирующих изучаемые нами сообщества. Насекомые, с большими способностями к расселению, регулируются в первую очередь абиотической средой, формируемые ими сообщества имеют небольшое обилие и высокую изменчивость в пространстве и времени, но довольно четкую привязанность к определенным биотопам. Высшие ракообразные и жаберные моллюски, напротив, медленно расселяются между водотоками и в большом масштабе распределены спорадично, зато более эврибионтны и способны накапливать огромную биомассу на широком диапазоне биотопов, независимо от обитающих там насекомых. Такие же свойства имеют и сообщества, где они доминируют. В более однородных и стабильных морских экосистемах, видимо, преобладают именно сообщества M-типа (Жирков, 2010), в относительно эфемерных речных, по нашим данным, сообщества Р-типа.
Выводы. Представленная нами классификация сообществ донных макробеспозвоночных объединяет более ранние схемы Жадина (Жадин, 1940) и Иллиеса (Illies, 1961), но существенно расширяет трактовку многих классов сообществ и вводит дополнительный класс сообществ рипали. В предлагаемом виде она позволяет включить почти все наши описания реофильных сообществ (хотя периодически встречаются пробы, занимающие пограничное положение между разными классами, или очень обедненные пробы, не пригодные для классификации). Для многих практических целей (оценки биоразнообразия, продуктивности, качества воды, кормовой базы рыб и т.п.) предлагаемой схемы может быть достаточно, хотя очевидна значительная неоднородность сообществ внутри каждого класса и возможность более дробной классификации (Chertoprud, 2011). Кроме того, высокая изменчивость сообществ связана с их динамикой и различными нарушениями (Tounsend, 1989; Thorp et al., 2006) и не может быть учтена предлагаемой схемой.
Хотя представленная схема объединяет данные только по одному материку, разнообразие обследованных регионов позволяет предположить, что реофильные сообщества других континентов также могут быть корректно распределены по пяти предлагаемым классам сообществ (со своими структурными особенностями). Чтобы убедиться, что данная схема имеет глобальный характер, требуются дальнейшие исследования.
Список литературы
Алимов А.Ф. 1979. Интенсивность обмена у водных пойкилотермных животных // Общие основы изучения водных экосистем. Ленинград: Наука. С. 5.
Богатов В.В. 1995. Комбинированная концепция функционирования речных экосистем // Вестник Дальновосточного отделения РАН. № 3. С. 51.
Жирков И.А. 2010. Жизнь на дне. Био-география и био-экология бентоса. Москва: Товарищество науч. изд. КМК.
Леванидов В.Я. 1981. Экосистемы лососевых рек Дальнего Востока // Беспозвоночные животные в экосистемах лососевых рек Дальнего Востока. Владивосток: Дальневосточного Научного Центра АН СССР. С. 3.
Монаков А.В. 1998. Питание пресноводных беспозвоночных. Москва: Институт проблем экологии и эволюции.
Палатов Д.М., Чертопруд М.В. 2012. Реофильная фауна и сообщества беспозвоночных тундровой зоны на примере Южного Ямала // Биология внут. вод. № 1. С. 23.
Палатов Д.М., Чертопруд М.В., Фролов А.А. 2016. Фауна и типы сообществ макрозообентоса мягких грунтов водотоков горных районов Восточного Причерноморья // Биология внут. вод. № 2. С. 45. https://doi.org/10.7868/S0320965216020145
Паньков Н.Н. 2004. Структурные и функциональные характеристики зообентоценозов р. Сылвы (бассейн Камы). Пермь: Изд-во Пермского ун-та.
Протасов А.А. 1989. Классификация сообществ пресноводного перифитона // Гидробиол. журн. Т. 25. № 6. С. 3.
Протасов А.А. 2011. Жизнь в гидросфере. Очерки по общей гидробиологии. Киев: Академпериодика.
Разумовский С.М. 1981. Закономерности динамики биоценозов. Москва: Наука.
Раменский Л.Г. 1938. Введение в комплексное почвенно-геоботаническое исследование земель. Москва: Сельхозгиз.
Семенов-Tян-Шанский A.П. 1937. Основные черты истории развития альпийских фаун // Докл. Академии наук СССР. С. 1211.
Чертопруд М.В. 2011. Альтернативные стратегии формирования реофильных сообществ макробентоса. Тезисы Всероссийской конференции “Экология малых рек в XXI веке: биоразнообразие, глобальные изменения и восстановление экосистем” (Тольятти, 5–8 сентября 2011 г.). С. 84.
Чертопруд М.В., Палатов Д.М. 2013. Реофильные сообщества макробентоса юго-западной части Кольского полуострова // Биология внутр. вод. № 4. С. 34. https://doi.org/10.7868/S0320965213040050
Чертопруд М.В., Палатов Д.М. 2017. Сообщества макрозообентоса малых водотоков Восточных Балкан // Биология внутр. вод. № 3. С. 45. https://doi.org/10.7868/S0320965217030068
Чертопруд М.В., Чертопруд Е.С., Воробьева Л.В. и др. 2020а. Сообщества макрозообентоса предгорных и равнинных водотоков Нижнего Приамурья // Биология внутр. вод. № 1. С. 49. https://doi.org/10.31857/S0320965220010040
Чертопруд М.В., Палатов Д.М., Чертопруд Е.С. 2020б. Реофильные сообщества макрозообентоса гор Центральной Азии на примере Западного Тянь-Шаня // Биология внутр. вод. № 3. С. 276. https://doi.org/10.31857/S0320965220020035
Braun-Blanquet J. 1964. Pflanzensociologie. Wien: Springer.
Chertoprud M. 2011. Diversity and classification of rheophilic communities of macrozoobenthos in middle latitudes of European Russia // Biol. Bull. Rev. V. 1. № 3. P. 165. https://doi.org/10.1134/S2079086411030017
Chertoprud M. 2019. Macrofauna communities in the mountain streams of Sri Lanka (Ceylon) // Inl. Wat. Biol. V. 12. № 2. P. 199. https://doi.org/10.1134/S1995082919020044
Chertoprud M., Palatov D. 2013. Macroscale ecotone effects for the stream macrobenthic fauna and communities of the Eastern Novosibirsk Region // Moscow Univ. Biol. Sci. Bull. V. 68. № 4. P. 200. https://doi.org/10.3103/S0096392513040020
Chertoprud M., Palatov D., Chertoprud E. 2018. Rheophilic macrozoobenthos communities of the Southern Himalayas // Inl. Water Biol. V. 11. № 4. P. 435. https://doi.org/10.1134/S1995082918040041
Culp J. 1986. Experimental evidence that stream macroinvertebrate community structure is unaffected by different densities of coho salmon fry // J. N. Am. Benthol. Soc. V. 5. № 2. P. 140.
Cummins K., Klug M. 1979. Feeding ecology of stream invertebrates // Ann. Rev. Ecol. Sys. V. 10. P. 147.
Dodds W., Gido K., Whiles M. et al. 2015. The Stream Biome Gradient Concept: factors controlling lotic systems across broad biogeographic scales // Freshwater Science. V. 34. № 1. P. 1.
Dudgeon D. 1999. Tropical Asian streams: zoobenthos, ecology and conservation. Hong Kong: Hong Kong Univ. Press.
European Commission. 2000. Water Framework Directive 2000/60/EC // Eur. Parliament and Eur. Community Official J. V. 327. P. 1.
Flecker A., Allan J. 1984. The importance of predation, substrate and spatial refugia in determining lotic insect distributions // Oecologia. V. 64. P. 306.
Forrester G. 1994. Influences of predatory fish on the drift dispersal and local density of stream insects // Ecology. V. 75. P. 1208.
Gilliam J., Sabat A., Fraser D. 1989. Strong effects of foraging minnows on a stream benthic invertebrate community // Ecology. V. 70. № 2. P. 445.
Gleason H. 1926. The individualistic concept of the plant association // Bull. Torrey Bot. Club. V. 53. № 1. P. 7.
Hawkes H. 1975. River zonation and classification // River Ecology. Oxford: Blackwell Scientific Publications. P. 312.
Hӧckelmann C., Pusch M. 2000. The respiration and filter-feeding rates of the snail Viviparus viviparus (Gastropoda) under simulated stream conditions // Arch. Hydrobiol. V. 149. № 4. P. 553.
Hynes H. 1970. The Ecology of running waters. Univ. Toronto Press: Toronto.
Illies J. 1961. Versuch einer allgemeinen biozonotischen Gliederung der Fliessgewasser // Int. Revue Ges. Hydrobiol. V. 46. P. 205.
Palatov D., Chertopud M. 2018. Macrozoobenthic communities of compact grounds in streams of eastern Black Sea region // Russ. J. Ecology. V. 49. № 1. P. 80. https://doi.org/10.1134/S1067413618010125
Palatov D., Chertoprud M. 2020. Macrozoobenthos communities of springs and streams of the Eastern Circum-Pontic region // Inl. Wat. Biol. V. 13. № 4. P. 592. https://doi.org/10.1134/S1995082920040070
Parasiewicz P. 2001. MesoHABSIM – a concept for application of instream flow models in river restoration planning. Fisheries. V. 26. № 9. P. 6.
Pennak R. 1971. Toward a classification of lotic habitats // Hydrobiologia. V. 38. № 2. P. 321.
Persson A., Swensson J. 2006. Vertical distribution of benthic community responses to fish predators, and effects on algae and suspended material // Aquat. Ecol. V. 40. P. 85.
Poff N. 1997. Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology // J. North American Bent. Soc. V. 16. P. 391.
Power M. 1992. Habitat heterogeneity and the functional significance of fish in river food webs // Ecology. V. 73. № 5. P. 1675.
Shelford V. 1913. Animal communities in temperate America. Geogr. Soc. Chicago, Chicago. Bull. V. 5. P. 368.
Statzner B., Higler B. 1985. Questions and comments on the River Continuum Concept // Can. J. Fish. Aquat. Sci. V. 42. P. 1038.
Suren A., Riis T. 2010. The effects of plant growth on stream invertebrate communities during low flow: a conceptual model // J. N. Am. Benthol. Soc. V. 29. № 2. P. 711.
Thorp J., Thoms M., Delong M. 2006. The riverine ecosystem synthesis: biocomplexity in river networks across space and time // River Res. Applic. V. 22. P. 123.
Townsend C. 1989. The Patch dynamics concept of stream community ecology // J. the North American Benthological Society. V. 8. № 1. P. 36.
Vannote R., Minshall G., Cummins K., Sedell I., Cushing C. 1980. The river continuum concept // Can. J. Fish. Aquat. Sci. V. 37. № 1. P. 130.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод