

Биология внутренних вод, 2022, № 1, стр. 96-100
Подтверждение суждения о повышенной устойчивости палеолимнических беспозвоночных к антропогенному загрязнению воды
А. В. Макрушин a, *, Е. В. Кузьмин a, Е. А. Соколова a, А. С. Васильев a, Э. В. Гарин a
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: makru@ibiw.ru
Поступила в редакцию 29.06.2020
После доработки 24.08.2021
Принята к публикации 03.09.2021
- EDN: DNQUBW
- DOI: 10.31857/S0320965222010077
Аннотация
Проведено сравнение чувствительности к антропогенному загрязнению видов Cladocera (Crustacea) и сделана попытка объяснить различия их чувствительности. Исследовались пробы планктона, собранные в июне–августе 2010–2013 и 2019 гг. на шести станциях Рыбинского водохранилища. При сравнении доли пораженных токсикозом особей у самых ранних вселенцев из моря в континентальные воды палеолимнических видов рода Daphnia с таковой у позже вселившихся в континентальные воды мезолимнических видов Leptodora kindti и Limnosida frontosa (Cladocera (Crustacea)) было выявлено, что у мезолимнических видов она выше, чем у палеолимнических. Это подтвердило взгляды Я.И. Старобогатова о меньшей устойчивости мезолимнических беспозвоночных к антропогенному загрязнению воды, чем палеолимнических.
Полученные авторами результаты можно объяснить, если принять точку зрения Я.И. Старобогатова (1970) – советского и российского зоолога, малаколога, профессора, доктора биологических наук, главного научного сотрудника Зоологического института РАН. Первичноводных беспозвоночных континентальных вод он делил по давности вселения их предков из моря в континентальные воды на нео-, мезо- и палеолимнические. Неолимнические беспозвоночные – недавние вселенцы и поэтому имеют близких морских родичей. Они еще мало приспособлены ко всему разнообразию условий жизни в континентальных водах, требовательны к содержанию в воде кислорода, не переносят резких колебаний температуры и поэтому населяют лишь крупные водоемы, режим которых относительно стабилен. Число видов в неолимнических родах мало, а ареалы их узкие. В мезолимнических родах число видов тоже невелико. Их предки вселились в континентальные воды раньше неолимнических, и поэтому у них приспособления к жизни в континентальных водах совершеннее, чем у неолимнических видов, а ареалы шире. Но и они населяют только крупные водоемы. Наиболее приспособлены ко всему разнообразию условий жизни в континентальных водах палеолимнические беспозвоночные, самые древние вселенцы. Их роды, в отличие от нео- и мезолимнических родов, часто содержат много видов, а семейства – много родов. Видовое их разнообразие − результат адаптивной радиации, произошедшей за долгое время жизни в континентальных водах. Близких морских родичей у них нет. Населяют они не только крупные водоемы, но и пруды, и лужи, а также осолоненные водоемы. Их ареалы широкие. Вселяясь из моря в континентальные воды, предки каждой группы становились сначала неолимническими, затем мезолимническими и потом палеолимническими. Палеолимнические беспозвоночные задолго до появления на Земле человека приобретали устойчивость к естественному загрязнению воды, т.е. к сапробности, к изменениям степени минерализованности воды, к насыщению ее гуминовыми и фульвокислотами, к выделениям макрофитов и водорослей и к другим изменениям ее химического состава. Поэтому, по мнению Я.И. Старобогатова, которое он устно сообщил в беседе с первым автором этой статьи, они менее чувствительны к антропогенному загрязнению, чем нео- и мезолимнические беспозвоночные. Прав ли он был? Авторы попытались ответить на этот вопрос.
Проверка утверждения Я.И. Старобогатова проводилась на представителях трех родов Cladocera – Leptodora, Daphnia и Limnosida. Среди Cladocera, по Я.И. Старобогатову (1970), неолимнических видов нет. Haplopoda, Onychopoda и с некоторым сомнением Holopedidae он относил к мезолимническим видам, а Anomopoda и Sididae – к палеолимническим. Таким образом, по Я.И. Старобогатову, Leptodora kindti (Focke, 1844) (Haplopoda) – мезолимнический вид, род Daphnia (Anomopoda) и вид Limnosida frontosa Sars, 1862 (Sididae) – палеолимнические. Отнесение Я.И. Старобогатовым всех Sididae к палеолимническим группам встречает возражение. В этом семействе палеолимнический род лишь Diaphanosoma. В нем 30 видов, которые встречаются во всех типах континентальных водоемов (Коровчинский, 2004). Род Penilia (Sididae) не палеолимнический, а вторично-морской. Род Limnosida (Sididae) тоже не палеолимнический, а мезолимнический. Мезолимническим его следует считать в соответствии с критерием давности заселения предками континентальных вод (Старобогатов, 1970). Этот род включает только один вид – Limnosida frontosa, который обитает лишь в крупных водоемах. Проблема неодновременности вселения предков Cladocera из моря в континентальные воды вскользь обсуждается Я.И. Старобогатовым в монографии, посвященной моллюскам (Старобогатов, 1970). Деление этих рачков на мезо- и палеолимнические он рассматривал лишь как возможное, из-за неполноты своих знаний. По нашему мнению, объяснить наличие в сем. Sididae столь различающихся по давности вселения в пресные воды родов Diaphanosoma, Penilia и Limnosida можно либо тем, что это семейство произошло от разных предков, либо, что входящие в него виды эволюционировали в разных направлениях и с разной скоростью. В настоящей работе чувствительность к загрязнению двух мезолимнических видов Lеptodora kindti и Limnosida frontosa сравнивали с таковой палеолимнических видов Daphnia (всех оказавшихся в пробах представителей этого рода).
Пробы собирали на двух плесах Рыбинского водохранилища̶: Волжском (ст. 1) и Главном (ст. 5) с судна в 2010–2013 гг. дважды в месяц с июня по август. Использовали планктонную сеть с газом № 76 размером ячеи 0.082 мм. В такую сеть L. kindti попадалaсь редко. В 2019 г. для увеличения количества обследованных рачков этого вида также применяли сеть с газом № 7 размером ячеи 1.093 и с бóльшим диаметром входного отверстия (~1 м). Пробы собирали 18 июня, 9, 25 июля и 19 августа на тех же станциях, что и в 2010–2013 гг. Рачков фиксировали в 4%-ном формалине. Просмотрены 220 проб (сборы 2010–2013 гг.) и 34 пробы (сборы 2019 г.). В пробах, собранных в 2010–2013 гг., сравнивали чувствительность к загрязнению Limnosida frontosa с таковой видов рода Daphnia (Leptodora kindti была очень малочисленна). В пробах 2019 г. сравнивали чувствительность к загрязнению L. kindti с таковой видов рода Daphnia. Пробы просматривали в камере Богорова под микроскопом МБС-9. Фотографии сделаны на цифровом микроскопе “VHX-1000E” (Япония). Перед фотографированием пробы окрашивали жидкостью Буэна. Содержащаяся в ней пикриновая кислота иногда окрашивала содержащуюся в отеках жидкость в желтый цвет и делала отеки выводковой сумки рачков более заметными.
За меру чувствительности вида к загрязнению воды принимали соотношение в пробах особей, имеющих отек выводковой сумки, и особей без видимых признаков этого токсикоза. Оценку достоверности различия проводили при помощи критерия χ2-квадрат Пирсона (Лакин, 1980; Животовский, 1991). Полученные значения этого критерия сравнивали с критическими для 5%-ного уровня значимости и соответствующего числа степеней свободы. Эти соотношения в 2010–2013 гг., за редким исключением, достоверно различались между собой, поэтому сравнивали долю особей Limnosida frontosa и видов рода Daphnia за каждый из 2010–2013 гг. по отдельности.
Отек – избыточное накопление жидкости во внеклеточном пространстве организма. У Cladocera отекает наружная стенка выводковой сумки (раковина) и внутренняя ее стенка (дно выводковой сумки, т.е. дорсальная сторона туловища) (Макрушин и др., 2014). Отек дна выводковой сумки наблюдали только у видов рода Daphnia. У Limnosida frontosa и Lеptodora kindti отекала только раковина. У обследованных Cladocera раковина состоит из двух слоев гиподермы. Оба они выстланы хитином и соединены протоплазматическими мостиками. Отек на раковине возникает вследствие разрушения мостиков, расслоения раковины и заполнения пространства между разошедшимися слоями ее гиподермы жидкостью. На дне выводковой сумки отек образуется вследствие отслоения от туловища гиподермы и накопления под ней жидкости. Отек имеет вид пузыря, наполненного прозрачной жидкостью.
У видов рода Daphnia наружный листок гиподермы раковины гораздо менее гибкий, чем внутренний. Внутренний ее листок у видов этого рода очень тонкий и очень прозрачный. Рассмотреть его в случае его отслоения от наружного листка и образования отека удавалось редко. У находящихся в зафиксированных пробах особей видов рода Daphnia он обычно был разорван, а содержащаяся под ним жидкость отека разлита. Но о том, что отек прежде присутствовал и что он давил на туловище живого рачка, свидетельствовало отогнутое положение его туловища (рис. 1а). На рис. 1а видно, что отек раковины у Daphnia sp. еще не достиг больших размеров. Вероятно, поэтому внутренний листок гиподермы, ограждающий отек изнутри, не поврежден. Он выпячивается внутрь выводковой сумки и немного похож на эфиппий. Наружной стенкой ему служит наружный листок гиподермы раковины, внутренней – внутренний ее листок. Из-за отека раковины и отека дна выводковой сумки пространство выводковой сумки у рачка уменьшилось, и его туловище выступает из створок раковины наружу (рис. 1а), а не находится, как у здоровых особей, между створками раковины.
Рис. 1.
Ветвистоусые рачки с отечной выводковой сумкой: а – Daphnia sp., б – Limnosida frontosa, в – Leptodora kindti. Т – туловище; K – прежние края раковины, место соединения наружного (H) и внутреннего (B) листков гиподермы. O1 – отек раковины, O2 – отек дна выводковой сумки. Масштаб 500 мкм.
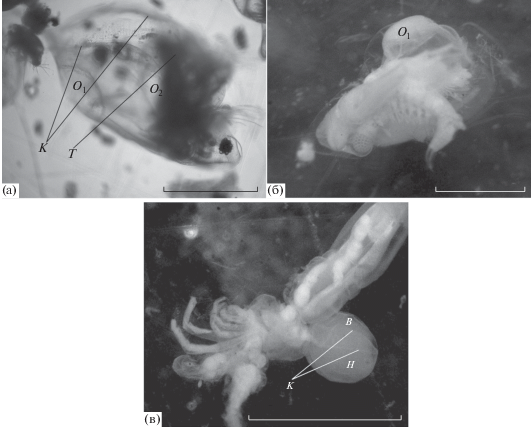
У Lеptodora kindti и у Limnosida frontosa, в отличие от Daphnia, оба листка гиподермы раковины одинаково гибкие. На рис. 1б у L. frontosa часть раковины расслоена. При отеке раковины этого вида наружный слой гиподермы выгибается наружу, внутренний – внутрь выводковой сумки. Жидкость, находящаяся между слоями гиподермы, окрашена пикриновой кислотой и поэтому не прозрачная, а мутная.
У Lеptodora kindti раковина имеет форму ложки. Она не охватывает туловище рачка с боков, как у Limnosida frontosa и у видов рода Daphnia, а прижимается к дорсальной стороне туловища. Между раковиной и туловищем у Leptodora kindti расположено пространство выводковой сумки, в ней самка вынашивает партеногенетических зародышей. При небольшом отеке раковины, т.е. при отслоении части внутреннего листка гиподермы от наружного, на раковине L. kindti, как и у Limnosida frontosa, возникает пузырь. Он уменьшает пространство выводковой сумки и снижает плодовитость самки. При обширном отеке у Lеptodora kindti расслаивается не часть, как у Limnosida frontosa, а вся раковина, и раковина приобретает форму вытянутого шара (рис. 1в). Поскольку пространство выводковой сумки у такой самки перестает существовать, при овуляции она вынуждена сбрасывать яйца в воду, где они, вероятно, погибают.
Вызывает вопрос низкая доля с отеком выводковой сумки особей рода Daphnia в 2019 г. – лишь 2% (табл. 1). Возможно, это связано с тем, что лето 2019 г. было холодным (табл. 2). C понижением температуры негативное действие загрязнения на Cladocera, по-видимому, снижается. Вероятно, среди Lеptodora kindti доля пораженных токсикозом особей летом 2019 г. тоже была ниже, чем в 2010–2013 гг. Доля представителей рода Daphnia с отечной выводковой сумкой за период 2010–2013 гг. была значительно ниже, чем доля Limnosida frontosa, пораженных этим токсикозом. Статистическая проверка показала достоверность различий: в 2010 г. $\chi _{{\text{ф}}}^{2}$ = 18.40 ⪢ $\chi _{{{\text{ст}}}}^{2}$ = 3.84, в 2011 г. $\chi _{{\text{ф}}}^{2}$ = = 63.88 ⪢ $\chi _{{{\text{ст}}}}^{2}$ = 3.84, в 2012 г. $\chi _{{\text{ф}}}^{2}$ = 54.67 ⪢ $\chi _{{{\text{ст}}}}^{2}$ = = 3.84, в 2013 г. $\chi _{{\text{ф}}}^{2}$ = 198.37 ⪢ $\chi _{{{\text{ст}}}}^{2}$ = 3.84. Достоверные различия в доле заболевших особей зарегистрированы и в 2019 г. между видами рода Daphnia и Lеptodora kindti: $\chi _{{\text{ф}}}^{2}$ = 6.17 > $\chi _{{{\text{ст}}}}^{2}$ = 3.84. Следовательно, L. kindti и Limnosida frontosa более чувствительны к антропогенному загрязнению Рыбинского водохранилища, чем представители рода Daphnia.
Таблица 1.
Доля особей с отечной выводковой сумкой
| Год | N | Рачки с отечной выводковой сумкой | |
|---|---|---|---|
| число | доля, % | ||
| Виды рода Daphnia | |||
| 2010 | 882 | 268 | 30 |
| 2011 | 993 | 259 | 26 |
| 2012 | 560 | 102 | 18 |
| 2013 | 489 | 131 | 27 |
| 2019 | 1158 | 19 | 2 |
| Limnosida frontosa | |||
| 2010 | 45 | 27 | 60 |
| 2011 | 96 | 62 | 65 |
| 2012 | 146 | 69 | 47 |
| 2013 | 91 | 76 | 86 |
| Leptodora kindti | |||
| 2019 | 126 | 7 | 6 |
Таблица 2.
Температура (°С) поверхностного слоя воды Рыбинского водохранилища в летний период
| Год | Июнь | Июль | Август | Среднелетняя |
|---|---|---|---|---|
| 2010 | 18.4 ± 2.1 | 26.2 ± 0.8 | 22.5 ± 2.4 | 22.3 ± 1.4 |
| 2011 | 17.0 ± 0.6 | 23.1 ± 0.6 | 21.2 ± 0.2 | 20.5 ± 0.9 |
| 2012 | 17.8 ± 0.9 | 21.3 ± 0.5 | 17.6 ± 0.5 | 19.9 ± 0.4 |
| 2013 | 20.2 ± 1.0 | 21.7 ± 1.1 | 19.8 ± 0.3 | 20.6 ± 0.5 |
| 2019 | 18.1 ± 0.9 | 18.5 ± 0.6 | 16.8 ± 0.8 | 18.0 ± 0.5 |
Выводы. Проведенное исследование подтверждает мнение Я.И. Старобогатова о большей чувствительности к антропогенному загрязнению мезолимнических беспозвоночных по сравнению с палеолимническими.
Список литературы
Животовский Л.А. 1991. Популяционная биометрия. Москва: Наука.
Коровчинский Н.М. 2004. Ветвистоусые ракообразные отряда Ctenopoda мировой фауны (морфология, систематика, экология, зоогеография). Москва: Товарищество науч. изданий КМК.
Котов А.А. 2013. Морфология и филогения Anomopoda (Cladocera, Crustacea). Москва: Тов-во науч. изд. КМК.
Лакин Г.Ф. 1980. Биометрия. Москва: Высш. шк.
Макрушин А.В., Семенова А.С., Дубовская О.П. и др. 2014. Географическое распространение отека раковинки у Cladocera (Crustacea) // Вода, химия и экология. № 10. С. 69.
Старобогатов Я.И. 1970. Фауна моллюсков и зоогеографическое районирование континентальных водоемов. Ленинград: Наука.
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод