

Биология внутренних вод, 2022, № 2, стр. 136-146
Динамика высшей водной растительности защищенного мелководья в связи с уровнем воды в Волжском плесе Рыбинского водохранилища
С. А. Поддубный a, Е. В. Чемерис a, *, А. В. Кутузов a, А. И. Цветков a, А. А. Бобров a
a Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: lechem@ibiw.ru
Поступила в редакцию 29.06.2021
После доработки 30.09.2021
Принята к публикации 07.11.2021
- EDN: EWMCRQ
- DOI: 10.31857/S0320965222020085
Аннотация
Проанализированы изменения высшей водной растительности заостровного защищенного мелководья “Красный ручей” с 1989 по 2019 гг. по данным аэрофотоснимков, разновременных спутниковых снимков и прямых полевых исследований. Для изучения зарастания использованы снимки за период со второй половины июля до конца сентября. Показана обратная зависимость степени зарастания и уровня воды. При низких значениях уровня воды (99.6–99.7 м) степень зарастания мелководья наибольшая (82–86%), при более высоких (100.3–100.7 м) – ниже (63–78.8%). Полученные данные не совпадают с таковыми, опубликованными по Рыбинскому водохранилищу до 2013 г. и требуют дальнейшего изучения на более обширном материале. Специфика регулировки уровня воды Рыбинского водохранилища, преобладающая обводненная фаза мелководья в период ледостава и на протяжении большей части вегетационного периода и высокие темпы седиментации создают благоприятные условия для распространения высшей водной растительности. К 2019 г. растительность на мелководье распространилась до глубин 1.8 м, площадь зарастания достигла 73% от нормального подпорного уровня. Гелофиты заняли ~70% оптимальной площади мелководья, гидрофиты − лишь 20%. С учетом высокого темпа седиментации и обмеления, процессы зарастания исследованного мелководья в ближайшее время будут происходить более быстро, их замедлению может способствовать поддержание уровня, близкого к нормальному подпорному уровню 101.81 м.
ВВЕДЕНИЕ
Изучение формирования и функционирования ВВР в водоемах имеет важное практическое значение, поскольку зарастающие мелководные зоны определяют процессы самоочищения и воспроизводства рыбных запасов в условиях естественных водоемов (Midwood, Chow-Fraser, 2010, 2012; Tan et al., 2020) и водохранилищ (Мордухай-Болтовской, Экзерцев, 1971; Довбня, 1983; Буторин, Успенский, 1984; Эдельштейн, 1998; Тимченко, 2006; Кочеткова и др., 2018; Sharip, 2021). Рыбинское водохранилище, второе по площади из водохранилищ Волжского каскада, представляет собой важный ресурс в экономике региона, в том числе, как рыбопромысловый водоем. Из 38 видов рыб, обитающих в Рыбинском водохранилище, большинство – фитофилы, откладывающие икру на растения. К ним относятся все карповые и щука. Молодь этих видов нагуливается среди зарослей водных растений все лето и уходит с мест нагула только при резком ухудшении условий, к которым можно отнести резкое падение уровня и осеннее охлаждение воды (Столбунов, 2007). ВВР вносит основной вклад (74.4%) в первичную продукцию водохранилища (Копылов и др., 2019). Таким образом, понимание закономерностей процессов зарастания защищенных мелководий и их направленности представляет собой важную фундаментальную и прикладную задачу.
Характер растительного покрова водохранилищ и его динамика определяются многими факторами: географическим положением, режимом эксплуатации и связанных с ним изменений уровня воды, осушения и обводнения мелководий, присутствием зачатков водных растений, степенью защищенности от воздействия ветрового волнения, трофическими и физико-химическими качествами воды и грунтов и др. (Экзерцев, Лисицына, 1984; Кочеткова и др., 2018). В последние десятилетия в свете происходящих масштабных изменений климата изучение воздействия колебаний уровня воды на биоту пресных водоемов приобретает особую актуальность (Wantzen et al., 2008). Известно, что пространственное распределение ВВР и многолетняя динамика зарастания мелководий зависят от гидрологического режима, и особенно от режима уровня. Это показано для озер (Keddy, Reznicek, 1986; Gasith, Gafny, 1990; Geest et al., 2005; Bresciani et al., 2012; Midwood, Chow-Fraser, 2012; Tan et al., 2020) и водохранилищ (Hill et al., 1998; Krolová et al., 2012, 2013; Папченков, 2013; Кочеткова и др., 2018; Поддубный и др., 2018). Зависимости “уровень воды–зарастание” для разных водоемов и разных климатических зон могут существенно различаться, но отмечена важность обводненной фазы, поскольку обводнение в начале вегетационного сезона определяет развитие растительности в текущем году, а в конце сезона – в последующие годы (Hill et al.,1998; Krolová et al., 2012, 2013; Поддубный и др., 2018). Зная это, можно оценивать тенденции развития ВВР и путeм регулирования уровня влиять на зарастание мелководий.
Первые попытки изучения воздействия гидрологических факторов на распределение ВВР в водоемах бассейна Верхней Волги проведены на примере озер Ярославской обл. и отдельных участков открытых мелководий Рыбинского вдхр. В результате исследований выявлены зависимости динамического воздействия ветровых волн и течений на отдельные виды макрофитов (Поддубный, Сухова, 2002). Количественная оценка влияния внешнего водообмена на ВВР получена на пойменных участках водохранилищ Днепровского каскада. Установлено, что по мере увеличения периода внешнего водообмена интенсивность продукции ВВР уменьшается (Тимченко, 2006). Достаточно хорошо изучена динамика зарастания Цимлянского вдхр., где общая тенденция увеличения площади зарастания в значительной степени связана с изменениями уровня воды и благоприятными условиями формирования мелководий (Кочеткова и др., 2018). В работе Папченкова (2013) показано изменение зарастания и продукции ВВР Рыбинского вдхр. за период с 2003 по 2009 гг., отмечен пульсирующий характер этих процессов, связанный с чередованием маловодных и многоводных фаз, что подтверждено нашими исследованиями (Поддубный и др., 2018).
Применение методов ДЗЗ позволяет осуществлять мониторинг окружающей природной среды на качественно ином уровне, дополняя и даже частично заменяя непосредственные полевые исследования. Спутниковые данные, наряду с использованием малых беспилотных летательных аппаратов, могут успешно применяться для изучения особенностей распределения отдельных видов гидробионтов и, в частности, позволяют идентифицировать пятна и пояса доминирующих растительных сообществ мелководий, их размеры и плотность (Midwood, Chow-Fraser, 2010, 2012; Novikova et al., 2012; Кочеткова и др., 2018; Кутузов, 2018). Снимки водоемов на разных временных отрезках – ценный материал, изучение и сравнение которого позволяет проводить анализ современного состояния растительности, сезонных и многолетних динамических процессов, и в итоге – прогнозировать и управлять ими.
Цель работы – сопоставить сезонную и многолетнюю динамику растительности и уровня воды, как определяющего фактора, на примере защищенного мелководья Рыбинского вдхр. “Красный ручей” на основе спутниковых снимков за 31-летний период (1989–2019 гг.), данных полевых исследований ВВР в 2009, 2015, 2018 гг., архивных материалов (аэрофотосъемка 1989 г.) и сведений о сезонном и многолетнем ходе уровня воды.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Многолетние среднемесячные данные с 1989 по 2019 гг. изменения уровня воды Рыбинского вдхр., объема притока в водоем и по атмосферным осадкам на его зеркало взяты из баз данных Рыбинской гидрометеообсерватории и Всероссийского научного исследовательского института гидрометеорологической информации – Мировой центр данных (ВНИИГМИ-МЦД) (Булыгина и др., 2019).
Для определения границ мелководья и крупномасштабного картографирования использовали: материалы ДЗЗ (данные спутников Landsat-5–8, Sentinel-2) с пространственным разрешением 10–15 м/пиксель; значения уровня воды на дату снимков за период 2009–2019 гг.; аэрофотоснимки мелководья, выполненные в 1989 г. (архивные материалы). Контуры побережья Рыбинского водохранилища для разных уровней сработки воды получены автоматизированной обработкой сцен спутниковых съемок среднего разрешения (до 10 м/пиксель) в ближнем инфракрасном оптическом диапазоне. Для определения на снимках открытой воды и зарослей макрофитов применяли индекс вегетации NDVI, который в простейшем виде представляет собой отношение:
NDVI = (NIR – RED) / (NIR + RED),
где NIR – отражение в ближней инфракрасной и RED – отражение в красной области спектра.
Границы суши выделяли по ранее полученным контурам (по материалам инфракрасной съемки ДЗЗ для ряда уровней стояния воды), что хорошо согласуется с использованными сезонными данными ДЗЗ. Диапазон полученных значений NDVI пикселей за пределами этой суши разбивали на три экспоненциально распределенных класса – открытая вода, разреженные заросли, плотные заросли. Для анализа сезонной динамики ВВР использовали серию безоблачных снимков за 2016 и 2019 гг., где наиболее полно отражен вегетационный период. Основную обработку данных проводили в стандартных пакетах модулей географических информационных систем (QGIS 3, ArcGIS 10). Поскольку на продолжительных временных отрезках видовой состав сообществ, формирующих зарастание, может меняться коренным образом (показано для мелководий Рыбинского вдхр. (Ляшенко, 1999)), а также невозможно идентифицировать сообщества конкретных видов на спутниковых снимках без привязки координат к описаниям, видовой состав сообществ не рассматривали.
Степень зарастания защищенного мелководья определяли за 1989 г. по архивным фотоснимкам путeм прямого подсчета, за 2009 г. – по результатам полевых исследований и одновременно с расчетом NDVI по спутниковым снимкам. Степень зарастания всего участка и обводненной части мелководья (протоки между островом и берегом) определяли как отношение площади зарастания к площади рассматриваемого мелководья в период максимального развития растительности при Zmaх (Распопов, 1985; Папченков, 2001, 2013). Следует отметить, что в 2009 и 2016 гг. Zmaх соответствовал или был очень близок к НПУ (101.81 м). Для уточнения доминирующих сообществ и пространственного распределения их зарослей в октябре 2015 и 2018 гг. проводили полевые исследования на отдельных участках мелководья.
Коэффициенты множественной детерминации и уравнения множественной регрессии для зависимости уровня воды и площадей зарастания ВВР вычисляли в MS Excel 2010 и STATISTICA 10.
Характеристика мелководья. Мелководье “Красный ручей” по классификации (Поддубный, 2013) относится к заостровным. Согласно архивным материалам, до создания Рыбинского вдхр. будущее мелководье располагалось на первой надпойменной террасе р. Волги. Урез воды проходил у подножья второй надпойменной террасы среди лесного массива. Площадь затапливаемого участка достигала ~9.4 км2 при протяженности ~3.8 км и средней ширине ~2.5 км. Заостровное пространство включало три типа ландшафтов: лесные массивы, луга и кустарники, проходимые болота с отдельными озерками. Ландшафты занимали площади 3.8, 3.9 и 1.7 км2 соответственно (рис. 1а). Перед затоплением ложе очищали от древесной растительности только до отметки 100 м, кустарники сохранялись и на отметке 98 м (Буянова, 1954). После затопления ~80% площади рассматриваемого заостровного мелководья занимали леса и закустаренные пространства.
Рис. 1.
Карта-схема защищенного мелководья “Красный ручей”: а ‒ до создания водохранилища (по: Буяновa, 1954), 98, 100 ‒ отметки изолиний высот, м; б ‒ в современных условиях (2000-е гг.). 1, 2, 3 ‒ отметки изолиний глубин, м; стрелкой указан судоходный канал. 1 ‒ границы участка, 2 ‒ затопленные русла рек, 3 ‒ “острова”, 4 ‒ зона кустарников, 5 ‒ зона леса, 6 ‒ проходимые болота, 7 ‒ обвалованный водоем.
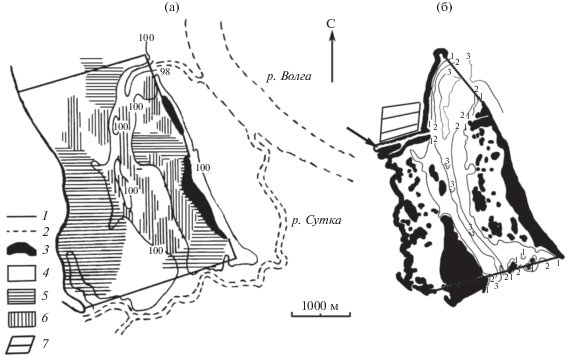
В начале 1960-х гг. в результате рекультивации северо-западной части мелководья (строительства судоходного канала и обвалованного водоема для рыборазведения) (Поддубный, 2006) его площадь уменьшилась до 6.8 км2 и сохранилась в таком виде до настоящего времени (рис. 1б). Современная протяженность по продольной оси не изменилась, а средняя ширина сократилась до 1.8 км. В результате образования большого количества мелких островков, площадь водного зеркала при НПУ (без учета ВВР) стала еще меньше – 5.21 км2 (рис. 1б). Глубины от 0 до 2 м занимают 69%, а >3 м – лишь 3.6% общей площади акватории.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Режим уровня. За 74 года эксплуатации Рыбинского вдхр. средняя многолетняя величина НПУ была 101.47 м (при проектной отметке 101.81 м) без проявления тренда к его увеличению или снижению. Минимальный уровень предполоводной сработки наблюдается в марте, в отдельные годы – в феврале, в среднем опускается до 98.80 м. За весь период наблюдений минимальный уровень предполоводной сработки постепенно повышался и после 2004 г. достиг ~99.00 м (рис. 2).
Рис. 2.
Многолетняя динамика уровня воды в Рыбинском водохранилище (1947‒2020 гг.): 1 ‒ Zmaх, 2 ‒ уровень предполоводной сработки, 3 ‒ принятая отметка дна исследуемого мелководья, 4 ‒ тренд Zmaх (R2 = 0.1), 5 ‒ тренд уровня предполоводной сработки (R2 = 0.4).

На исследованном мелководье минимальная отметка дна его ложа находится несколько ниже отметки 98.81 м или по глубине >3 м относительно НПУ. Среднее многолетнее сезонное изменение уровня воды в водохранилище таково, что в марте мелководье полностью обсыхает с вероятностью сохранения в пределах ложбины дна незначительных по площади изолированных обводненных участков. В зимние месяцы глубина мелководья изменяется от 0.37 до 1.1 м, осенью – от 1.2 до 1.6 м и летом – от 2.0 до 2.6 м. За весь период наблюдений мелководье в 50% случаев полностью обсыхало, причем 78% из них приходилось на период с 1947 по 1983 гг. Начиная с 2004 г., полное осушение мелководья наблюдалось только в 2015 г. Основными факторами осушения и обводнения заостровного мелководья выступают годовые изменения объема притока в водохранилище, зависящие в свою очередь от количества осадков на зеркало водоема и его водосбор. Влияние этих природных факторов регулируется режимом работы гидроузлов.
Растительный покров и его динамика. Основа растительного покрова мелководья “Красный ручей” и других защищенных мелководий Рыбинского вдхр. представлена гелофитами – наиболее приспособленной к переменному уровню воды группой видов (Папченков, 2013; Поддубный и др., 2017, 2018). В табл. 1 приведены обобщенные данные по результатам картирования мелководья в конце августа 2009 г. Гелофиты занимали 27% площади мелководья (Phragmites australis (Cav.) Trin. ex Steud, Schoenoplectus lacustris (L.) Palla, Glyceria maxima (C. Hartm.) Holmb. и др.), гидрофиты ‒ 11% (с плавающими листьями Nuphar lutea (L.) Smith, Persicaria amphibia (L.) S.F. Gray и др. и погруженные Potamogeton perfoliatus L., P. lucens L., Stuckenia pectinatа (L.) Börner и др.), гигрофиты и гигрогелофиты (Carex spp., Phalaroides arundinacea (L.) Rauschert и др.) ‒ лишь 3%. При этом вклад экогрупп в зарастание был 65.9, 27.3 и 6.8%, что хорошо соотносится с закономерностями, выявленными для растительного покрова защищенных мелководий Верхневолжских водохранилищ (Поддубный и др., 2017). Согласно этим расчетам, площадь зарослей макрофитов (Sovg1) достигала 2.13 км2 (41% общей площади мелководья при Zmaх, равном НПУ).
Таблица 1.
Основные экогруппы ВВР и занимаемые площади (S) на мелководье в 2009 г.
| Основные экогруппы | S, м2 | Вклад в зарастание, % |
|---|---|---|
| Гидрофиты: | ||
| c плавающими листьями | 73 000 | 3.4 |
| погружeнные | 508 280 | 23.9 |
| Всего | 581 280 | 27.3 |
| Гелофиты: | ||
| высокотравные | 121 1680 | 56.9 |
| низкотравные | 190 960 | 9 |
| Всего | 1 402 640 | 65.9 |
| Гигрофиты и гигрогелофиты: | ||
| осоки и травы | 14 000 | 6.8 |
Сезонная динамика. Пик развития ВВР в умеренных широтах приходится на вторую половину июля–август, именно в этот период рекомендуется проводить изучение продукции и зарастания (Распопов, 1985). Для оценки зарастания методами ДЗЗ важно понимать, какой период будет наиболее показательным в случае конкретного мелководья. Нами отобраны серии снимков мелководья “Красный ручей”, наиболее полно отражающие вегетационный сезон: серия из пяти сцен (май–октябрь) за 2016 г. (рис. 3) и серия из трех (июнь–август) за 2019 г. Заостровная протока – участок площадью 2.21 км2, на котором наиболее ярко прослеживается пространственно-временная динамика растительного покрова, поскольку оставшаяся часть мелководья полностью заросла. Этот участок и был выбран для оценки динамики сезонного и многолетнего зарастания.
Рис. 3.
Сезонная динамика зарастания защищенного мелководья в 2016 г.: a – 02.05, б – 29.06, в – 24.07, г – 01.09, д – 09.10. Масштаб 500 м. 1 – открытая вода, 2 – разреженные заросли, 3 – плотные заросли.
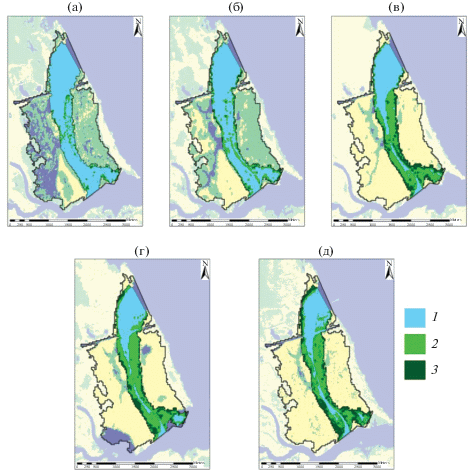
При компьютерной обработке космоснимков диапазон полученных значений NDVI разбивали на три экспоненциально распределенных класса: W – открытая вода; L – разреженные заросли с общим проективным покрытием <50%; D – плотные заросли с общим проективным покрытием >50%. На исследованном мелководье класс L в целом хорошо соотносится с сообществами гидрофитов (Nuphar lutea, Persicaria amphibia, Potamogeton perfoliatus, P. lucens и др.), класс D – с сообществами гелофитов (Phragmites australis, Schoenoplectus lacustris, Sparganium emersum Rehm., Typha latifolia L. и др.). Результаты анализа сезонных снимков мелководья за 2016 и 2019 гг. приведены в табл. 2.
Таблица 2.
Изменение площадей открытой воды и макрофитов в протоке защищенного мелководья за вегетационные периоды 2016 и 2019 гг.
| Показатель | 2016 г. | 2019 г. | ||||||
|---|---|---|---|---|---|---|---|---|
| V | VI | VII | IX | X | VI | VII | VIII | |
| SW | 1.95 | 1.71 | 1.02 | 0.91 | 0.9 | 1.92 | 1.13 | 1.06 |
| SL | 0.19 | 0.35 | 0.79 | 0.96 | 0.83 | 0.23 | 0.66 | 0.78 |
| SD | 0.04 | 0.14 | 0.39 | 0.33 | 0.45 | 0.05 | 0.4 | 0.35 |
| Sovg2 | 0.23 | 0.49 | 1.18 | 1.29 | 1.28 | 0.28 | 1.06 | 1.13 |
| SL/SD | 4.75 | 2.5 | 2 | 2.9 | 1.8 | 4.6 | 1.65 | 2.23 |
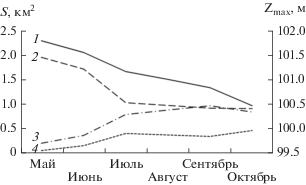
Активный рост макрофитов на мелководье приходится на вторую половину июня–начало июля, что приводит к значительному и резкому росту зарастания (рис. 3). С июля до конца августа площадь, занимаемая ВВР, прирастает заметно медленнее и стабилизируется с незначительным увеличением к началу осени (табл. 2). Основной вклад в зарастание протоки принадлежит сообществам гидрофитов, занимаемые ими площади в 2–4.5 раза превышают таковые гелофитов (рис. 4, табл. 2), но основной вклад в зарастание всего мелководья остается за гелофитами (см. выше).
Рис. 4.
Сезонная динамика площадей открытой воды (S) и классов макрофитов в протоке в 2016 г.: 1 – уровень воды; 2 – открытая вода; 3 – разреженные заросли; 4 – плотные заросли.
Зависимость площади открытой воды (SW) в протоке, а также площади зарастания протоки макрофитами (Sovg2) от Zmaх и τ (времени (сут) с начала вегетации, отсчeт проведeн с 1 мая) выражается следующими уравнениями множественной регрессии:
SW = –78.0823 – 0.0008τ + 0.7851Zmaх (r = 0.91);
Sovg2 = 72.2471 + 0.00148τ – 0.7073Zmaх (r = 0.91).
Многолетняя динамика зарастания мелководья. Зарастание протоки в многолетнем аспекте изучали по отобранным спутниковым снимкам во время пика развития ВВР (вторая половина июля–первая половина сентября) в безоблачную погоду за 31 год (с 1989 по 2019 гг.).
За расчетную площадь мелководья принимали его площадь при НПУ 101.81 м за вычетом площадей поднятий “островов” (рис. 1б), на которых развивается древесно-кустарниковая растительность, – 5.21 км2 для всего мелководья и 2.21 км2 для протоки. Вычисление степени зарастания всего мелководья и протоки при Zmaх позволили оценить изменения ВВР (рис. 5) за это время.
Рис. 5.
Динамика степени зарастания (Covg) заостровной протоки и мелководья “Красный ручей” в разные годы. 1 ‒ Zmaх, м, 2 ‒ степень зарастания всего мелководья, 3 – степень зарастания протоки, 4 – линия тренда.
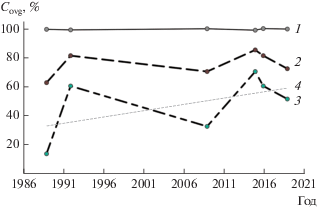
При низких (99.6–99.7 м) уровнях воды в конце лета и начале осени (август 1992 г. и сентябрь 2015 г.) в результате значительной осушки мелководья и образования песчаных пляжей степень зарастания протоки и всего мелководья наибольшая (рис. 5). В случае более высоких уровней воды (100.3–100.7 м) степень зарастания несколько ниже (рис. 5). Выявлен импульсный характер зарастания, что типично для Рыбинского вдхр. в целом (Папченков, 2013). На протяжении 31-летнего периода прослеживается тенденция к увеличению зарастания мелководья, при этом зарастание его обводненной части имеет более ярко выраженные максимумы и минимумы (рис. 5).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
До затопления водами Рыбинского вдхр. на участке будущего мелководья в заболоченных и обводненных понижениях, по прибрежным участкам рек были распространены сообщества Carex sp., Calamagrostis sp. и Equisetum sp. Также отмечены ценозы Schoenoplectus lacustris, Caltha palustris L., Alisma plantago-aquatica L., Galium sp., Comarum palustre L., Sparganium sp., Typha sp., Phragmites australis, Eleocharis sp., Oenanthe aquatica (L.) Poir. и Lemna sp. (Буянова, 1954). Эти сообщества стали источником диаспор будущей ВВР водохранилища. Первые исследования по изучению формирования ВВР на отдельных участках мелководий Рыбинского вдхр. проводили с 1947 г. (окончание заполнения водоeма водой до НПУ) по 1949 г. В этот период уровень наполнения водохранилища изменялся в пределах 101.41‒101.81 м. Мелководье “Красный ручей” занимало ~1/3 части исследуемого ранее в Волжском плесе прибрежного участка. Согласно данным Богачева (1950), здесь на начальном этапе растительный покров формировали исключительно свободноплавающие растения, занимающие до 66% общей площади зарастания мелководья. К 1949 г. произошла постепенная смена доминантов на воздушно-водные виды (>50% площади ВВР).
По приблизительной оценке, расчетная площадь зарастания увеличилась с 0.1 в 1947 г. до 0.5 в 1949 г. Проведенные Ляшенко (1995) исследования в 1963–1987 гг. выявили увеличение площадей зарастания водохранилища, абсолютное доминирование в растительном покрове гелофитов (94.8%) и изменение в их структуре ‒ низкотравные гелофиты сменились на высокотравные. Последующие наблюдения на стационарных полигонах Волжского плеса в 1987–1997 гг. показали непрерывное нарастание площадей Phragmites australis, Phalaroides arundinaceae, Glyceria maxima и увеличение их доли в растительном покрове (Ляшенко, 1999). Подробное описание и картирование ВВР заостровного мелководья “Красный ручей” проведено В.Г. Папченковым в 2009 г. К этому времени гелофиты занимали 27%, гидрофиты ‒ 11%, гигрогелофиты ‒ 3% площади мелководья. Общая площадь зарослей макрофитов достигала 2.13 км2 (т.е. степень зарастания всего мелководья 41%). Рассчитанная по спутниковым снимкам степень зарастания изучаемого мелководья в 2009 г. была выше ‒ 71%, что, вероятно, связано с использованием разных методик.
Ранее показано (Поддубный и др., 2018), что для Рыбинского вдхр. существует нелинейная зависимость уровня воды и площади зарастания. Аналогичная зависимость Sovg3 = f(Z) получена и для Волжского плеса, где расположено мелководье:
Sovg3 = 30‒41e0.95Z (R2 = 0.92).
Изменение площадей зарастания, вычисленных по приведенной выше зависимости, представлено на рис. 6.
Рис. 6.
Изменение площадей зарастания (Sovg3) Волжского плеса за 31-летний период: 1 – линия тренда.
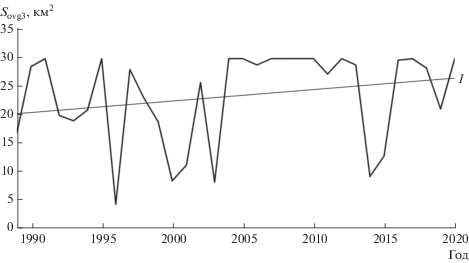
На мелководье “Красный ручей” наблюдается следующая тенденция: при уменьшении значений уровня заполнения наблюдается увеличение зарастания (рис. 5). Таким образом, пики максимальных значений зарастания этого участка находятся в противофазе с таковыми, рассчитанными для Волжского плеса (рис. 6) и всего водохранилища (Поддубный и др., 2018). При этом тренд увеличения зарастания со временем одинаково статистически мало значим для рассматриваемых объектов (рис. 5, 6), что связано с недостаточной выборкой данных. Расчеты по изменению площадей зарастания Рыбинского водохранилища с момента заполнения по 2015 г. (Поддубный и др., 2018), продленные с 2016 по 2020 гг., также показали тенденцию к увеличению зарастания, что может быть связано с повышением минимального уровня предполоводной сработки в течение последних пяти лет (рис. 2), преобладанием обводненной фазы до начала вегетации, а также с естественным старением водоема.
На основании анализа спутниковых снимков для летнего периода 2019 г. показано увеличение площади макрофитов до 3.8 км2, т.е. степень зарастания возросла до 73% НПУ. Допуская сохранение соотношения распространения экогрупп ВВР, можно заключить, что на мелководье, в отсутствии заиления и сохранении глубин, только для гидрофитов сохраняется достаточно большой процент резервной площади их распространения (80%). Гелофиты заняли полностью свою оптимальную площадь, а в резерве осталось ≤30% максимально возможной площади их распространения.
Полученные данные по степени зарастания мелководья и тенденции еe изменения отличаются от результатов исследования ВВР Рыбинского водохранилища с 2003 по 2009 гг., показавших зависимость зарастания от многоводных (как правило, с высокими значениями НПУ) и маловодных (с низким НПУ) циклов (Папченков, 2013). Отмечено, что в маловодные периоды (1963–1976 и 1996–2007 гг.) происходило снижение степени зарастания, в многоводные (1977–1995 гг. и с 2008 г.), за счет освоения гидро- и гелофитами новых площадей наблюдалось увеличение зарастания. Для мелководья “Красный ручей” получена противоположная зависимость ‒ снижение зарастания наблюдалось после завершения маловодных циклов (рис. 5). Возможная причина этого – разные методики расчета зарастания, Папченков (2013) использовал глазомерное картирование и вычислял степень зарастания относительно площади всего мелководья, а не от Zmax. Этот вопрос требует изучения на более обширном материале. Аналогичные закономерности получены и для мелководий Волгоградского вдхр. (Кочеткова и др., 2018). В этой работе выделены две фазы зарастания: медленного постепенного зарастания (1987–1998 гг.) и быстрого импульсивного (1998–2017 гг.), когда общая площадь заросших мелководий увеличилась до ≥4% акватории водохранилища. Авторами было также отмечено, что в последние годы (2014–2017 гг.) процессы зарастания ВВР заметно активизировались. Возможно, подобные процессы происходят и в Рыбинском вдхр., однако для таких выводов необходим более репрезентативный материал.
Особенности регулировки режима уровня Рыбинского вдхр., а именно: среднее многолетнее значение НПУ поддерживается на 34 см ниже проектной отметки, средний многолетний уровень в течение периода вегетации (июнь–сентябрь) на ~80 см ниже НПУ и преобладающая обводненная фаза мелководья в период ледостава, создают благоприятные условия для постепенного распространения ВВР на более глубокие участки мелководья. В настоящее время открытая вода при Zmax занимает площадь 2.21 км2 и представляет собой узкую протоку шириной 350–750 м (рис. 2а). В течение вегетационного периода на фоне сезонного снижения уровня воды и развития водных растений площадь открытой воды постепенно сокращается с одновременным увеличением площадей гидро- и гелофитов (табл. 2, рис. 3, 4).
Заиление водохранилищ – неотъемлемый процесс существования водоема. Согласно современным представлениям, средняя многолетняя скорость осадконакопления в Рыбинском вдхр. к 2010 г. достигала 2.2 мм/г (Законнов, 2018). Защищенные мелководья, в силу их изолированности от ветро-волнового воздействия и влияния стоковых течений, должны характеризоваться бóльшими, по сравнению с открытой литоралью, скоростями осадконакопления. Этому, кроме пониженной гидродинамики, способствует и исходное (до затопления) состояние подстилающей поверхности (болотная растительность, остатки кустарников и леса). Такие процессы, как слабая гидродинамическая активность, размыв в начальной стадии существования водохранилища и вторичное переотложение донных осадков в пределах мелководья, интенсивное зарастание и отмирание макрофитов служат источниками поступления органики и осадочного материала на дно мелководья. Допуская достаточно точное отображение изолиний рельефа местности до затопления и современного рельефа дна мелководья (рис. 1), можно оценить накопление осадков за 74-летний период, которое составляет ~80–100 см (по изолинии рельефа 100 м, где 2 м глубины до затопления соответствует современной глубине в 1–1.2 м). В этом случае средняя скорость осадконакопления может быть 11–13 мм/год, что в ~2 раза превышает таковую в глубоководной зоне Рыбинского вдхр. – 6 мм/год (Законнов, 2018). Акватория исследуемого мелководья с глубинами <1 м занимает >2.7 км2 его площади. К 2009 г. эти глубины фактически полностью освоены макрофитами. К 2019 г. растительность уже распространилась до глубины 1.8 м. С учeтом происходящего обмеления, процессы зарастания будут идти в более быстром темпе. Как отмечено ранее (Поддубный и др., 2018), экогруппы ВВР имеют достаточно большой резерв площади их распространения на защищенных мелководьях водохранилища. На данный момент сложно определить временной период полного зарастания мелководий. Темпы зарастания будут зависеть от ряда факторов: размеров мелководья, степени изолированности его от глубоководной зоны, глубины участка, в речных плесах расположением относительно направления стоковых течений, интенсивности проявления ветро-волнового воздействия, характера сезонных изменений уровня воды.
Выводы. Использование спутниковых снимков позволяет проследить за сезонной и многолетней динамикой зарастания ВВР отдельных типов мелководий и всей мелководной зоны Рыбинского вдхр. Существующая методика расчетов не дает возможности оценить площади, занимаемые отдельными доминантами ВВР, однако при глубинах ≤2 м позволяет оценить общие площади зарастания основными экогруппами. Предложенные методы определения степени зарастания мелководий ВВР могут быть использованы при проведении аналогичных исследований водохранилищ, имеющих явно выраженные сезонные изменения уровня воды. На примере серии снимков растительного покрова мелководья в 2016 г. показано, что для изучения зарастания в Верхне-Волжском регионе следует использовать период со второй половины июля до конца сентября. Для мелководья “Красный ручей” выявлена обратная зависимость площади зарастания от уровня воды – увеличение зарастания при уровнях ниже НПУ, и его снижение при уровнях превышающих НПУ. Это противоречит данным, полученным ранее для Волжского плеса и Рыбинского вдхр., и требует дальнейшего изучения. Рассмотренное заостровное мелководье в настоящее время близко к стадии полного зарастания ВВР, несмотря на имеющийся резерв по площади для еe распространения. Последующие годы с малой водностью и уровнем воды на ~1 м ниже проектного НПУ (101.81 м) с большой вероятностью могут привести к резкому увеличению зарастания гелофитной растительностью всех типов защищенных мелководий – заостровных, карманных и заливов со средней глубиной ~1.5 м. За время эксплуатации Рыбинского вдхр. таких периодов было семь (1963–1964, 1971−1973, 1980–1981, 1988–1989, 1995–1996, 2002–2003, 2014–2015 гг.) с цикличностью 6–12 лет. При поддержании уровня, близкого к НПУ, зарастание будет протекать медленнее.
Список литературы
Богачев В.К. 1950. О развитии водной растительности в Рыбинском водохранилище // Тр. биол. ст. “Борок”. Москва; Ленинград: Наука. Вып. 1. С. 302.
Буторин Н.В., Успенский С.М. 1984. Значение мелководий в биологической продуктивности водохранилищ // Биологические ресурсы водохранилищ. Москва: Наука. С. 23.
Булыгина О.Н., Разуваев В.Н., Александрова Т.М. 2019. Описание массива данных суточной температуры воздуха и количества осадкой на метеорологических станциях России и бывшего СССР (TTTR). Свидетельство о государственной регистрации базы данных № 2014620942/ВНИИГМИ-МЦД. Обнинск. URL: http://meteo.ru/data/162-temperatureprecipitation# описание-массива-данных (дата обращения: 17.10.2019).
Буянова О.Ф. 1954. Зарастание Рыбинского водохранилища и его анофелогенность в первые годы существования. Строительство водохранилищ. Москва: Медгиз. С. 121.
Довбня И.В. 1983. Продукция высшей растительности волжских водохранилищ // Пресноводные гидробионты и их биология: Тр. ИБВВ АН СССР. Ленинград: Наука. Вып. 48(51). С. 71.
Законнов В.В. 2018. Характеристика донных отложений // Структура и функционирование экосистемы Рыбинского водохранилища в начале XXI века. Москва: РАН. С. 59.
Копылов А.И., Масленникова Т.С., Рыбакова И.В., Минеева Н.М. 2019. Вклад автотрофных сообществ в формирование первичной продукции в экосистеме Рыбинского водохранилища // Биология внутр. вод. № 4. С. 94.https://doi.org/10.1134/S0320965219040260
Кочеткова А.И., Брызгалина Е.С., Калюжная И.Ю. и др. 2018. Динамика зарастания Цимлянского водохранилища // Принципы экологии. № 1. С. 60. https://doi.org/10.15393/j1.art.2018.7202
Кутузов А.В. 2018. Использование современных и архивных данных ДЗЗ для ГИС мониторинга околоводных экосистем // Трансформация экосистем. № 1. С. 86. https://doi.org/10.23859/estr-180209
Ляшенко Г.Ф. 1995. Высшая водная растительность Рыбинского водохранилища: Автореф. дис. … канд. биол. наук. Санкт-Петербург.
Ляшенко Г.Ф. 1999. Динамика высшей водной растительности Рыбинского водохранилища // Ботан. журн. Т. 84. № 12. С. 106.
Мордухай-Болтовской Ф.Д., Экзерцев В.А. 1971. Гидробиологический режим мелководий и их значение для продуктивности волжских водохранилищ // Вопросы комплексного использования водохранилищ. Киев: Наукова думка. С. 57.
Папченков В.Г. 2001. Растительный покров водоемов и водотоков Среднего Поволжья. Ярославль: ЦМП МУБиНТ.
Папченков В.Г. 2013. Степень зарастания Рыбинского водохранилища и продуктивность его растительного покрова // Биология внутр. вод. № 1. С. 24. https://doi.org/10.7868/S0320965212030102
Поддубный С.А. 2006. Научно-исследовательский флот Института биологии внутренних вод им. И.Д. Папанина Российской академии наук. Страницы истории. Рыбинск: Изд-во ОАО “Рыбинский Дом печати”.
Поддубный С.А. 2013. Защищенные мелководья верхневолжских водохранилищ и их экологическое значение // Вода: химия и экология. № 11. С. 35.
Поддубный С.А., Сухова Э.В. 2002. Моделирование влияния гидродинамических и антропогенных факторов на распределение гидробионтов в водохранилищах: руководство для пользователей. Рыбинск: Рыбинский Дом печати.
Поддубный С.А., Папченков В.Г., Чемерис Е.В., Бобров А.А. 2017. Зарастание защищенных мелководий верхневолжских водохранилищ в связи с их морфометрией // Биология внутр. вод. № 1. С. 65. https://doi.org/10.7868/S0320965217010144
Поддубный С.А., Чемерис Е.В., Бобров А.А. 2018. Влияние режима уровня воды на зарастание мелководий Рыбинского водохранилища (обзор) // Биология внутр. вод. № 4. С. 38. https://doi.org/134/S0320965218030142
Распопов И.М. 1985. Высшая водная растительность больших озер Северо-Запада СССР. Ленинград: Наука.
Столбунов И.А. 2007. Особенности распределения молоди рыб в прибрежной зоне Рыбинского водохранилища // Биология внутр. вод. № 4. С. 55.
Тимченко В.М. 2006. Экологическая гидрология водоемов Украины. Киев: Наукова думка. 384 с.
Эдельштейн К.К. 1998. Водохранилища России: экологические проблемы, пути их решения. Москва: ГЕОС.
Экзерцев В.А., Лисицына Л.И. 1984. Изучение растительных ресурсов водохранилищ Волжского каскада // Биол. ресурсы водохранилищ. Москва: Наука. С. 89.
Bresciani M., Bolpagni R., Braga F. et al. 2012. Retrospective assessment of macrophytic communities in southern Lake Garda (Italy) from in situand MIVIS (Multispectral Infrared and Visible Imaging Spectrometer) data // J. Limnol. V. 71(1). P. 180. https://doi.org/10.4081/jlimnol.2012.e19
Geest G.V., Wolters H., Roozen F. et al. 2005. Water-level fluctuations affect macrophyte richness in floodplain lakes // Hydrobiologia. V. 539. P. 239.
Gasith A., Gafny S. 1990. Effects of Water Level Fluctuation on the Structure and Function of the Littoral Zone // Large Lakes. Brock/Springer Series in Contemporary Bioscience. Berlin; Heidelberg: Springer. P. 156. https://doi.org/10.1007/978-3-642-84077-7_8
Hill N.M., Keddy P.A., Wiesheu I.C. 1998. A hydrological model for predicting the effects of dams on the shoreline vegetation of lakes and reservoirs // Environ. Manage. V. 22. P. 723. https://doi.org/10.1007/s002679900142
Keddy P.A., Reznicek A.A. 1986. Great Lakes vegetation dynamics: the role of fluctuating water levels and buried seeds // J. Great Lakes Res. V. 12. P. 25.
Krolová M., Čižkovab H., Hejzlarc J. 2012. Depth limit of littoral vegetation in a storage reservoir: A case study of Lipno Reservoir (Czech Republic) // Limnologica. V. 42. P. 165.
Krolová M., Čižkovab H., Hejzlarc J. 2013. Response of littoral macrophytes to water level fluctuations in a storage reservoir // Knowl. Managt. Aquatic Ecosyst. V. 408(07). P. 2. https://doi.org/10.1051/kmae/2013042
Midwood J.D., Chow-Fraser P. 2010. Mapping floating and emergent aquatic vegetation in coastal wetlands of Eastern Georgian Bay, Lake Huron, Canada // Wetlands. V. 30. P. 1. https://doi.org/10.1007/s13157-010-0105-z
Midwood J.D., Chow-Fraser P. 2012. Changes in aquatic vegetation and fish communities following 5 years of sustained low water levels in coastal marshes of eastern Georgian Bay, Lake Huron // Global Change Biology. V. 18. P. 93. https://doi.org/10.1111/j.1365-2486.2011.02558.x
Novikova N., Kalioujnaia I., Kalioujnaia N. et al. 2012. Identification and mapping of environmental conflicts for the Tsimlyansk water reservoir // Arid Ecosystems. V. 2. № 3. P. 156. https://doi.org/10.1134/S2079096112030110
Sharip Z. 2021. Changes in phytoplankton and zooplankton abundance and diversity in macrophyte-dominated and open pelagic ecosystem of shallow reservoirs // Inland Water Biology. V. 14. № 4. P. 427. https://doi.org/10.1134/S1995082921040088
Tan W., Xing J., Yang S. et al. 2020. Long-term aquatic vegetation dynamics in Longgan lake using Landsat Time Series and their responses to water level fluctuation // Water. V. 12. P. 21. https://doi.org/10.3390/w12082178
Wantzen K.M., Rothhaupt K.-O., Mörtl M. et al. 2008. Ecological effects of water-level fluctuations in lakes: an urgent issue // Hydrobiologia. V. 613. P. 1. https://doi.org/10.1007/s10750-008-9466-1
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод