

Биология внутренних вод, 2022, № 3, стр. 278-289
Рост муксуна Coregonus muksun в различных условиях водности р. Обь
А. К. Матковский a, *, Т. А. Красноперова a
a Тюменский филиал Всероссийского научно-исследовательского института рыбного хозяйства и океанографии (“Госрыбцентр”)
Тюмень, Россия
* E-mail: Matkovskiy@gosrc.ru
Поступила в редакцию 14.05.2021
После доработки 23.09.2021
Принята к публикации 29.09.2021
- EDN: EKDZGY
- DOI: 10.31857/S0320965222030147
Аннотация
Положительное влияние на рост муксуна Coregonus muksun (Раllas) оказывают температура воды в границах избираемого оптимума, снижение концентрации рыбы, высокая степень развития кормовой базы и продолжительный период затопления поймы; из неблагоприятных факторов выделяются низкая водность и сильный летний прогрев воды в пойменно-соровой системе. В холодные многоводные годы скорость роста у половозрелого муксуна снижается. Особи поколений, рожденных в благоприятных условиях нагула и воспроизводства, в первые годы жизни отличаются более высоким темпом роста. Увеличение частоты жарких лет неблагоприятно сказывается на росте и воспроизводстве муксуна. Наиболее благоприятными для роста особей являются теплые многоводные и средней водности годы.
ВВЕДЕНИЕ
Для Обь-Иртышского бассейна характерны значительные годовые и сезонные колебания уровня воды, что оказывает существенное влияние на все стороны жизни рыб и через развитие и использование кормовой базы определяет темп роста особей, а также циклические изменения в численности популяций (Никольский, 1965). Рыбы разных широт, в зависимости от избираемых температур, особенностей энергетического обмена и метаболизма, по-разному реагируют на изменения в окружающей среде (Карамушко, 2007; Голованов, 2013). Происходящие климатические изменения сказываются на гидрологическом режиме водных объектов и отрицательно влияют на холодолюбивую ихтиофауну (Богданов, 2010; Герасимов, 2019; Матковский, 2019).
Хотя муксун в р. Обь до недавнего времени считался важнейшим промысловым объектом, особенности его роста в условиях изменения водности и потепления климата недостаточно исследованы (Москаленко, 1956; Замятин, 1977; Князев, Набоков, 1988; Князев, Крохалевский, 1995). Изучение этого вопроса актуально не только с точки зрения анализа происходящих изменений в условиях обитания, но и с позиции необходимости восстановления запасов муксуна, отдельные популяции вида внесены в Красную книгу РФ, а их численность продолжает снижаться.
Цель исследования – изучить особенности влияния гидрологического режима на рост муксуна р. Обь. В ходе исследования планировали изучить половые и возрастные различия в росте муксуна, а также выяснить насколько происходящие изменения могут сказываться на воспроизводстве этого вида.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для изучения роста использовали данные биологического анализа муксуна за период 2007–2020 гг. Индивидуальный рост изучали посредством измерения радиусов чешуи (Чугунова, 1959). Измеряли краниальный (передний) и латеральный (боковой) радиусы чешуи (рис. 1). Материал, в основном, собирали в весенний период (май–июнь) во время анадромной миграции муксуна в р. Обь, проанализировано 1058 экз. рыб (табл. 1). Поскольку в период анадромной нагульно-нерестовой миграции в стаде присутствовали, главным образом, половозрелые особи, дополнительно анализировали чешую и у неполовозрелых рыб (71 экз.), пойманных в зимний период (декабрь) в Обской губе в районе пос. Яптик-Сале.
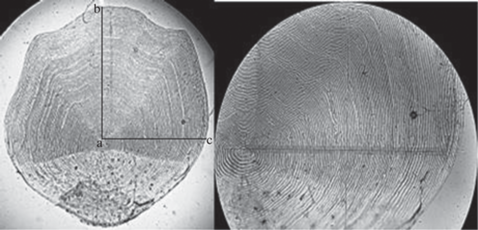
Таблица 1.
Места сбора и объем обработанного материала для обратных расчетов роста муксуна
| Год | Место | Водный объект | Число особей, экз. |
|---|---|---|---|
| 2007 | У пос. Ямбура | p. Обь | 60 |
| 2010 | То же | То же | 151 |
| 2012 | » | » | 131 |
| 2013 | » | » | 87 |
| 2014 | » | » | 120 |
| 2015 | » | » | 161 |
| 2017 | » | » | 180 |
| 2018 | » | » | 49 |
| 2019 | » | » | 69 |
| 2020 | » | » | 50 |
| 2001 | У поc. Яптик-Сале | Обская губа | 23 |
| 2007 | То же | То же | 48 |
| Всего | 1129 | ||
На основе полученных измерений строили зависимости промысловой длины рыбы от радиуса чешуи. Выбирали зависимость с наибольшей коррелятивной связью и использовали для расчетов. При вычислении коэффициентов уравнений размерный ряд представляли особи 15.1–55.7 см промысловой длины (от начала рыла до основания лучей хвостового плавника). Наиболее тесная связь зарегистрирована с латеральным радиусом чешуи (рис. 2).
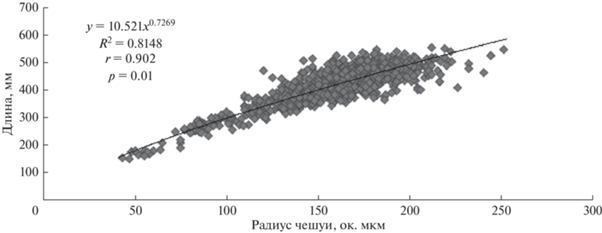
Из полученного уравнения было выведено следующее соотношение для проведения обратных вычислений:
Расчетные значения длины сопоставляли по критерию Стьюдента с фактическими, на основании чего был сделан вывод о допустимости использования полученного соотношения (табл. 2).
Таблица 2.
Сравнение результатов расчета промысловой длины муксуна (поколения 2003 г.р.) с фактическими значениями
| Возраст, лет | Фактическая длина, см |
n, экз. | Расчетная длина, см | t-критерий выборки |
t-критерий при р = 0.01 |
n, экз. |
|---|---|---|---|---|---|---|
| 9 | 45.14 ± 0.630 | 24 | 43.18 ± 0.750 | 2.01 | 2.62 | 40 |
| 10 | 45.8 ± 0.810 | 17 | 45.46 ± 0.722 | 0.38 | 2.70 | 23 |
| 11 | 47.3 ± 0.930 | 7 | 47.21 ± 0.717 | 0.09 | 2.83 | 16 |
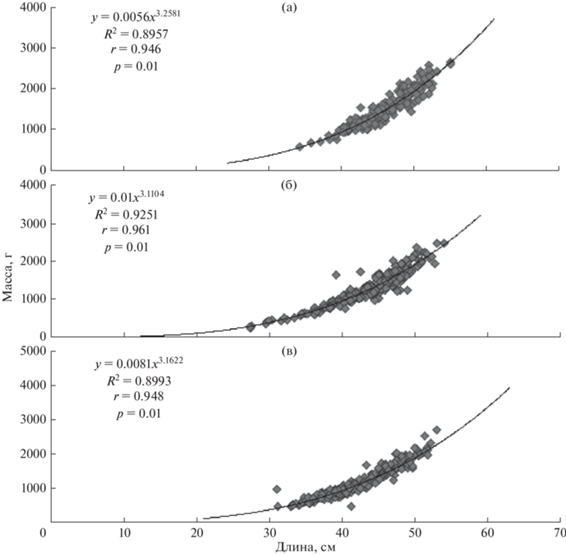
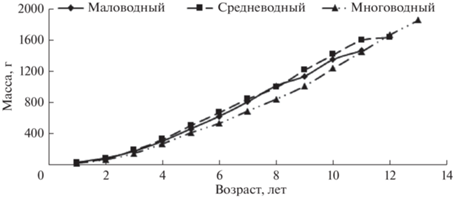
Для изучения массового роста использовали фактические и расчетные данные. Для этого вычисляли зависимости между массой и длиной рыбы. При этом учитывали различия в условиях обитания в разные годы водности (рис. 3) – отдельно для наиболее типичных маловодных (данные 2011 и 2012 гг.), средневодных (2010, 2014, 2020 гг.) и многоводных лет (2015, 2016 гг.) (рис. 4).
Рис. 4.
Зависимости между промысловой длиной и массой тела муксуна в маловодные 2011, 2012 гг. (а), средневодные 2010, 2014 гг. (б) и многоводные 2015, 2016 гг. (в).
Поскольку обратные расчисления проводят для изучения различных линейных изменений в росте (Зубова и др., 2015), для выяснения возможности последующего получения объективного представления о массе рыбы оценивали достоверность по критерию Стьюдента. С этой целью анализировали только данные, которые не использовали для построения зависимостей. В частности, сравнивали расчетные данные с фактическими по маловодному 2006 г., средневодному 2008 г. и многоводному 2007 г. Результаты свидетельствуют об отсутствии достоверных различий между расчетными и фактическими значениями (табл. 3).
Таблица 3.
Сравнение фактической массы муксуна с расчетными значениями в годы различной водности
| Возраст, лет | Масса фактическая | Масса расчетная, г | n | t-критерий выборки | t-критерий при p = 0.01 |
|---|---|---|---|---|---|
| 2006 г. (маловодный) | |||||
| 8+ | 1358 ± 114.70 | 1308 ± 52.60 | 4 | 0.40 | 4.6041 |
| 9+ | 1404 ± 69.49 | 1340 ± 58.20 | 10 | 0.71 | 3.2498 |
| 10+ | 1419 ± 75.62 | 1399 ± 65.70 | 11 | 0.20 | 3.1693 |
| 2007 г. (многоводный) | |||||
| 8+ | 1264 ± 49.66 | 1241 ± 57.90 | 13 | 0.30 | 3.0545 |
| 9+ | 1354 ± 34.31 | 1279 ± 38.40 | 25 | 1.46 | 2.7969 |
| 10+ | 1445 ± 30.69 | 1417 ± 38.11 | 33 | 0.57 | 2.7500 |
| 11+ | 1504 ± 53.41 | 1445 ± 64.30 | 14 | 0.71 | 3.0123 |
| 12+ | 1618 ± 29.46 | 1537 ± 108.90 | 6 | 0.72 | 4.0321 |
| 2008 г. (средневодный) | |||||
| 8+ | 1245 ± 71.62 | 1236 ± 52.90 | 13 | 0.10 | 3.0545 |
| 9+ | 1430 ± 32.55 | 1426 ± 35.80 | 20 | 0.08 | 2.8609 |
| 10+ | 1508 ± 31.77 | 1494 ± 29.30 | 55 | 0.32 | 2.7045 |
| 11+ | 1757 ± 46.43 | 1590 ± 44.80 | 25 | 2.59 | 2.7969 |
| 12+ | 1796 ± 121.86 | 1659 ± 77.00 | 10 | 0.95 | 3.2498 |
Для выяснения влияния гидрологического режима на рост рыб анализировали данные за маловодные 2004, 2005, 2011, 2012 гг., средневодные 2000, 2008, 2010, 2013, 2017 гг. и многоводные 2002, 2007, 2015, 2016 гг. Исходя из средней температуры воздуха в июне–июле в районе г. Салехард, из данного перечня к холодным годам можно отнести 2002, 2008, 2010 гг., к теплым – 2000, 2004, 2005, 2007, 2012, 2013, 2015–2017 гг., из них к сравнительно жарким – 2012 и 2016 гг. В целом, после 2011 г. количество теплых лет существенно возросло (табл. 4).
Таблица 4.
Температура воздуха в районе г. Салехард, °С
| Год | Июнь | Июль | Средняя |
|---|---|---|---|
| 2000 | 11.7 | 14.7 | 13.2 |
| 2001 | 9.9 | 13.0 | 11.5 |
| 2002 | 8.9 | 14.0 | 11.5 |
| 2003 | 11.2 | 14.4 | 12.8 |
| 2004 | 10.8 | 16.7 | 13.8 |
| 2005 | 10.4 | 16.0 | 13.2 |
| 2006 | 11.4 | 14.4 | 12.9 |
| 2007 | 8.6 | 18.7 | 13.7 |
| 2008 | 8.6 | 15.3 | 12.0 |
| 2009 | 8.6 | 13.7 | 11.2 |
| 2010 | 8.7 | 13.3 | 11.0 |
| 2011 | 13.2 | 11.7 | 12.5 |
| 2012 | 16.2 | 16.0 | 16.1 |
| 2013 | 11.7 | 18.5 | 15.1 |
| 2014 | 11.2 | 11.2 | 11.2 |
| 2015 | 13.5 | 13.5 | 13.5 |
| 2016 | 14.1 | 19.4 | 16.8 |
| 2017 | 9.5 | 16.8 | 13.2 |
| 2018 | 9.8 | 16.8 | 13.3 |
| 2019 | 8.1 | 16.9 | 12.5 |
| 2020 | 9.9 | 15.5 | 12.7 |
Изменения в массовом и линейном росте изучали с учетом существующих рекомендаций и посредством анализа ряда показателей: относительных приростов, средних значений массы и удельной скорости роста (Шмальгаузен, 1935; Мина, Клевезаль, 1976; Дгебуадзе, 2001).
Статистическую обработку проводили с использованием пакета Statistica 6.0. Корреляции считали по формуле Пирсона.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Полученные данные по линейному росту муксуна свидетельствуют, что относительные линейные приросты муксуна резко снижаются после 5–6-годовалого возраста и становятся минимальными с 8-годовалого возраста (рис. 5). Существенного влияния на линейный рост различных фаз водности не выявлено. Поскольку в отличие от линейного роста, массовый рост, связанный степенной зависимостью с линейным, быстрее реагирует на изменения условий обитания, проведен аналогичный анализ по этому показателю. Кроме того, из-за непостоянства соотношения полов в популяции проведена возможность различий удельной скорости роста самцов и самок. Данные, представленные на рис. 6, не дали оснований говорить о половом диморфизме, и в дальнейших расчетах использованы объединенные выборки. Анализ массового роста муксуна в годы разной водности свидетельствует о тенденции к его снижению в многоводные годы, наиболее быстрый соматический рост отмечен в средневодные годы (рис. 7). Таким образом, для разных по водности лет по массовому и линейному росту прослеживаются свои особенности.
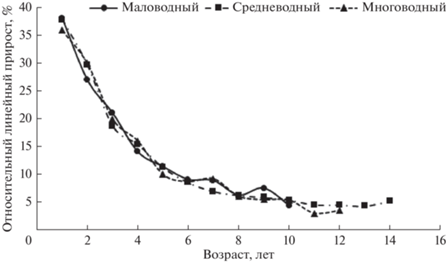
Рис. 6.
Удельная скорость линейного (а) и массового (б) роста самок и самцов муксуна. 1 – самки, 2 – самцы.
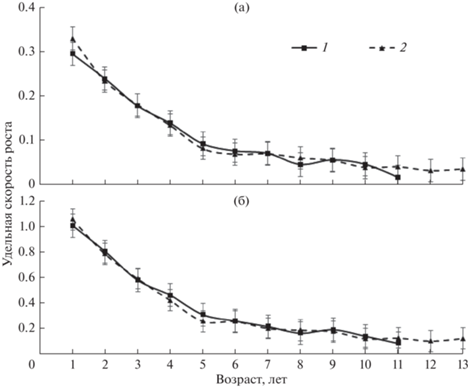
Рис. 7.
Массовый рост муксуна в годы разной водности: маловодные – 2004, 2005, 2011, 2012 гг.; средневодные – 2000, 2008, 2010, 2013, 2017 гг.; многоводные – 2002, 2007, 2015, 2016 гг.
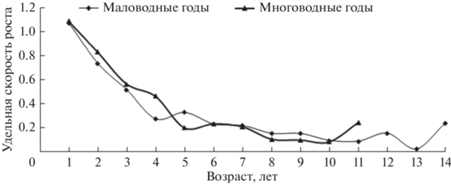
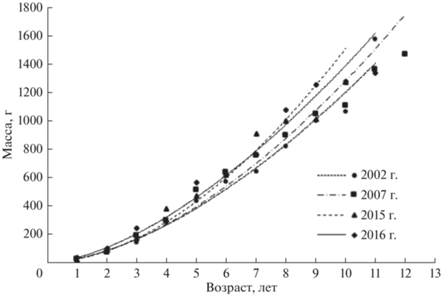
Поскольку гидрологический режим определяет условия нагула рыб и оказывает влияние на плотность рыбного населения, это сказывается и на темпе роста рыб. У большинства младших возрастных групп муксуна удельная скорость массового роста в многоводные годы заметно выше, чем в маловодные (рис. 8). В этом отношении младшевозрастные группы наиболее показательны, поскольку у их особей темп роста высокий, и энергетические вещества в основном тратятся на соматический рост. В итоге, одним из важнейших факторов, оказывающим влияние на рост, служит плотность населения. Результаты предшествующих исследований свидетельствуют, что прирост биомассы сиговых находится в прямой зависимости от периода затопления поймы, продолжительности нагула и температуры воды (Князев, Огурцова, 1988; Князев, Брусынина, 1990). Все это можно проследить на примере различных по температурному режиму многоводных лет – холодного 2002 г. (средняя температура воздуха в районе г. Салехард в июне–июле была 11.5°С) и теплых 2007 г. (13.3°С), 2015 г. (13.5°С), 2016 г. (18.1°С). В теплые годы темп роста выше, превышал таковой в холодный 2002 г. (рис. 9).
Рис. 8.
Удельная скорость весового роста муксуна в годы разной водности, маловодные – 2004, 2005, 2011, 2012 гг., многоводные – 2002, 2007, 2015, 2016 гг.
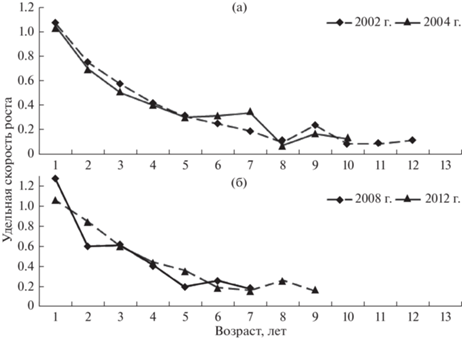
Отмечено, что многочисленные поколения, родившиеся в период или после высокой водности, в первые годы жизни отличаются более высоким темпом массового роста, который в последующем снижается. В качестве примера рассмотрим различия в массовом росте сравнительно многочисленных поколений 2002 и 2008 гг. рождения (высокой водности) и менее многочисленных 2004 и 2012 гг. рождения (низкой водности). Только после пятого года жизни малочисленные поколения начинают опережать по темпу роста многочисленные генерации (рис. 10).
Рис. 10.
Удельная скорость весового роста особей поколений муксуна 2002 и 2004 г.р. (а), 2008 и 2012 г.р. (б).
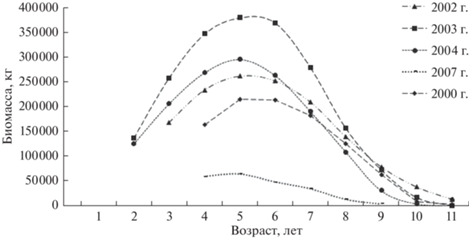
Также выявлено, что у муксуна наступление возраста максимальной продуктивности не зависит от гидрологического режима р. Обь и в среднем приходится на пятый, иногда на шестой год жизни, т.е. на период до наступления половой зрелости (рис. 11).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Существенное снижение темпа линейного роста у муксуна, наблюдаемое с 6–8-годовалого возраста (рис. 5), связано, очевидно, с началом полового созревания и достижением половой зрелости, а максимальные линейные приросты, как и у других сигов (Зубова и др., 2015; Савчук и др., 2017; Шестаков, 2021), происходят в первые годы жизни (Чаликов, 1931).
Некоторое опережение темпа роста у самок в отдельные годы перед возрастом наступления половой зрелости (рис. 6), по-видимому, связано с их более интенсивным питанием, что обусловлено необходимостью накопления большего количества энергетических веществ для созревания гонад (Крохалевский, 1983). У самок, в отличие от самцов, формирование фонда половых клеток происходит в летний период (Исаков, Селюков, 2010), стимулируя их активный нагул. Кроме того, известно, что при созревании семенников расходуется гораздо меньше жира, чем при созревании яйцеклеток (Белянина, Макарова, 1965).
Снижение с определенного возраста темпа роста муксуна в многоводные годы (рис. 7, 8) обусловливается несколькими причинами. Во-первых, многоводные годы, как правило, более холодные (Лещинская, 1962; Кузикова, 1984; Семенова и др., 1989), вода медленнее прогревается. Во-вторых, они наиболее благоприятные для созревания рыб, следовательно, в этот период у половозрелых рыб происходит перераспределение части энергетических веществ на генеративный процесс, нерестовую миграцию и нерест (Белянина, Макарова, 1965; Решетников, 1966; Решетников и др., 1971; Князев, 1989; Зубова и др., 2015; Rijnsdorp et al., 2005). Подтверждается известный факт, что в многоводные годы создаются благоприятные условия для нагула и созревания муксуна (Москаленко, 1956; 1958; Замятин, 1977). Именно многоводные годы дают наиболее существенный импульс в созревании большинства особей популяций сиговых (Михайличенко, 1989, 1992; Богданов, Агафонов, 2001; Матковский, 2006; Matkovskiy, 2014; Госькова, 2016). Снижение темпа роста у половозрелых рыб в многоводье происходит, несмотря на более продолжительный период нагула в пойменной системе. Поэтому, такой нагул лишь положительно сказывается на росте молоди муксуна.
Массовое созревание сигов в многоводные годы происходит в силу благоприятных условий нагула. Для холоднолюбивой оксифильной ихтиофауны свойственно более интенсивное накопление энергетических веществ в холодные годы. Так, установлено увеличение коэффициента общей жирности сигов Чунозера (Кольский п-ов, бассейн р. Чуна) в холодные годы (Решетников и др., 1971). Аналогично в холодные годы происходит и более интенсивное накопление жира в печени наваги (Шатуновский, 1980). Поэтому неслучайно основная часть особей муксуна созревает в периоды высокой водности (Матковский, 2006), когда термический режим более низкий. Однако в настоящее время в связи с потеплением климата ситуация меняется, и многоводные годы все чаще становятся теплыми (рис. 9, табл. 4), что, по-видимому, негативно сказывается на воспроизводстве сигов и на ритме функционирования их популяций.
Таким образом, в многоводье соматический рост у половозрелого муксуна снижается из-за возрастающих энергетических трат на созревание гонад, миграцию и нерест. В период нерестовой миграции и нереста, когда муксун не питается, существенно увеличиваются затраты на активный обмен. В это время в составе крови увеличивается количество эритроцитов и повышается уровень гемоглобина (Петкевич, 1981). Особенно это выражено у самцов (Лугаськова, 1990), которые дольше задерживаются в районе нерестилищ и в большей степени истощаются (Шатуновский, 1980). Кроме того, существенно возрастают траты энергетических веществ на саму миграцию (Мельниченко, 1988). Доля таких затрат на активный обмен может достигать 80–90% (Яржомбек и др., 1977).
В годы средней и низкой водности (рис. 7) положительное влияние на рост муксуна оказывают температура воды, более высокая степень развития кормовой базы (Карзинкин, 1952; Салазкин, 1976; Добринская и др., 1983; Семенова и др., 1989; Лелеко, 1989), а также снижение энергетических трат на созревание гонад. Поскольку в годы средней водности условия для нагула сигов лучше, чем в маловодье, то это отражается и на темпе их роста (Крохалевский, 1983). Гидрологический режим, продолжительность нагула и обеспеченность пищей оказывают влияние на рост сигов (Замятин, 1977; Шестаков, 2021). В целом по совокупности рассматриваемых факторов годы средней водности наиболее благоприятны для роста сигов, по-видимому, они оптимальны и по температурному режиму.
Полученные нами данные свидетельствуют, что наименьшую массу тела имел муксун в холодный 2002 г. (рис. 9), подтверждая вывод об увеличении темпа его роста в теплые годы. Тем не менее, более высокий прогрев воды в 2015 и 2016 гг. ухудшил условия для роста половозрелого муксуна в относительно мелководной пойменной системе и в целом неблагоприятно сказался на холоднолюбивой ихтиофауне (Матковский и др., 2017). В жаркие годы период нагула у сигов в пойменных водоемах почти прекращается при достижении температуры воды >20°С (Князев, Набоков, 1988). Молодь муксуна в районе салм Обской губы перестает питаться при температуре воды 18–20°С (Дрягин, 1948), поэтому в жаркие многоводные годы аналогично маловодным годам количество созревающих рыб сокращается. Известно, что температура определяет скорость энергетического обмена, соответственно, и темп роста (Васнецов, 1953; Винберг, Печень, 1968; Анохина, 1971; Никольский, 1974; Мина, Клевезаль, 1976; Рыжков, 1976; Карамушко, 2007; Князев, 2017), особенно если температура находится в границах видового избираемого оптимума (Ивлев, 1958; Шатуновский, 1980; Голованов, 2013; Голованов, Некрутов, 2018; Смирнов и др., 2020). У муксуна данная прямая связь нарушается лишь при температуре воды >21°С (Князев, Огурцова, 1988).
С увеличением температуры воды выше видового избираемого оптимума, что часто отмечается в маловодные годы, муксун прекращает активный нагул. В то же время, скорость обменных процессов продолжает сохраняться на достаточно высоком уровне. В итоге возрастающие энергетические траты не могут компенсироваться энергетическими веществами, поступающими с пищей, и темп роста снижается (Бретт, 1983). Данный эффект может прослеживаться не только в маловодные, но и в многоводные годы. Поэтому в сравнительно жаркие многоводные годы (2015 и 2016 гг.) темп роста у муксуна оказался ниже, чем в теплый 2007 г. (рис. 9). Можно сделать вывод, что годы с высоким прогревом воды неблагоприятны для созревания и формирования многочисленных нерестовых стад муксуна. По-видимому, неслучайно в современный период роль северных нерестовых притоков р. Обь в воспроизводстве сиговых возрастает (Богданов, 2010). Предположительно, сиги заходят в эти реки, чтобы нагуливаться в более холодной воде. Для восстановления запасов муксуна актуально создание новых северных центров его воспроизводства (Никонов, 1963).
Наряду с температурой воды на рост муксуна оказывает влияние плотность популяции (Мина, Клевезаль, 1976; Силин, 1990; Дгебуадзе, 2001), а также то, в каких условиях появилось на свет поколение (рис. 8, 10). Особенно это заметно по урожайным поколениям многоводных лет и поколениям, появившимся после благоприятных условий нагула. Особи этих генераций обладают высоким стартовым потенциалом роста, темп которого снижается только со вступлением в фазу пониженной водности. В условиях низкой водности происходит не только ухудшение условий нагула рыб, но и обостряются различные конкурентные взаимоотношения. При этом темп роста у особей многочисленных генераций снижается не сразу, а с неким временным интервалом. У поколений, рожденных в неблагоприятных условиях, мы наблюдаем обратную картину – темп роста возрастает в более позднем возрасте, т.е. со вступлением генерации в фазу повышенной водности, когда концентрация рыб снижается. Схожие особенности роста регистрируют в рыбоводстве при использовании крупного посадочного материала, полученного за счет разряженной плотности посадки (Легкодимова и др., 2019). Рассмотренная особенность роста, по-видимому, свойственна не только для муксуна, но и для большинства других видов рыб, так как для крупной молоди спектр доступного размерного состава пищи расширяется, а при увеличении концентрации рыб возрастает внутривидовая конкуренция.
Известно, что возраст максимальной продуктивности может существенно варьировать по годам (Кудерский, 1986) и зависит от скорости убыли генерации и темпа массового роста особей. У муксуна в возрасте пяти–шести лет эти процессы приводят к некому равновесному состоянию биомассы (рис. 11). Во-первых, в этот период абсолютные приросты массы заметно возрастают, во-вторых, естественная смертность до начала наступления половой зрелости постоянно снижается (Тюрин, 1972). Кульминация биомассы в более позднем возрасте не происходит из-за того, что смертность во время миграций и нереста может возрастать (Шатуновский, 1980). Ранее возраст кульминации биомассы муксуна не изучали. Возраст массового достижения половой зрелости у этого вида оценен Л.А. Зыковым (1996) как равный 7.5 годам, который, исходя из теоретических положений метода, определения естественной смертности (Зыков, Слепокуров, 1982) также должен быть близким к возрасту максимальной продуктивности (Матковский, 2014; Шибаев, 2014).
Выводы. У обского муксуна наиболее высокий темп линейного роста прослеживается до пятигодовалого возраста, половой диморфизм не выражен. Гидрологический режим оказывает существенное влияние на темп массового роста муксуна. Данное воздействие носит многогранный характер – через развитие кормовой базы, продолжительность нагула рыб, плотность населения, температурный режим, изменение скорости обменных процессов и способности особей к накоплению необходимых резервных веществ происходят те или иные изменения в росте. Существенное влияние оказывают температура воды, обеспеченность пищей и плотность населения. К наиболее значимым отрицательным факторам относятся маловодье и сильный прогрев воды в пойменно-соровой системе, приводящие к сокращению периода нагула и повышенному расходу энергетических веществ в организме рыбы. Многоводные годы благоприятны для роста, главным образом, неполовозрелого и, возможно, пропускающего нерест муксуна. Эти годы, как правило, более холодные и процесс накопления резервных веществ, необходимых для созревания половых продуктов, идет более интенсивно. Значительные затраты энергетических веществ на генеративный процесс, нерестовую миграцию и нерест снижают темп массового роста. Многоводные годы положительно сказываются на воспроизводстве муксуна. Рожденные в этот период особи на протяжении ряда лет сохраняют высокий потенциал роста. Годы средней водности наиболее благоприятны для роста муксуна – в основном, это теплые годы, с высоким развитием кормовой базы и относительно длительным периодом нагула. Дальнейшее потепление климата будет отрицательно сказываться на росте и воспроизводстве муксуна.
Список литературы
Анохина Л.Е. 1971. Об изменчивости роста онежской сельди Белого моря // Закономерности роста и созревания рыб. Москва: Наука. С. 21.
Белянина Т.Н., Макарова Н.П. 1965. Некоторые закономерности в распределении жира в организме рыб в связи с созреванием гонад // Теоретические основы рыбоводства. Москва: Изд-во “Наука”. С. 42.
Богданов В.Д. 2010. Состояние воспроизводства и динамика генераций сиговых Нижней Оби // Биология, биотехника разведения и состояние запасов сиговых: Матер. VII междунар. научно-произв. совещ. (Тюмень, 16–18 февраля 2010 г.). Тюмень: Госрыбцентр. С. 83.
Богданов В.Д., Агафонов Л.И. 2001. Влияние гидрологических условий поймы Нижней Оби на воспроизводство сиговых рыб // Экология. № 1. С. 50.
Бретт Дж.Р. 1983. Факторы среды и рост // Биоэнергетика и рост рыб. Москва: Легк. и пищ. пром-сть. С. 275.
Васнецов В.В. 1953. О закономерностях роста рыб // Очерки по общим вопросам ихтиологии. Москва: Изд-во АН СССР. С. 218.
Винберг Г.Г., Печень Г.А. 1968. Рост, скорость развития и плодовитость в зависимости от условий среды // Методы определения продукции водных животных. Минск: “Вышэйшая школа”. С. 45.
Герасимов Ю.В. 2019. Изменение популяционных показателей массовых видов рыб водоемов умеренной зоны при потеплении климата // Тез. докл. XII съезда гидробиологического общества при Российской академии наук (г. Петрозаводск, 16–20 сентября 2019 г.). Петрозаводск. С. 101.
Голованов В.К. 2013. Температурные критерии жизнедеятельности пресноводных рыб. Москва: Полиграф-Плюс.
Голованов В.К., Некрутов Н.С. 2018. К вопросу классификации пресноводных и морских рыб по отношению к температурному фактору среды // Рыбохозяйственные водоемы России: Фундаментальные и прикладные исследования: Матер. II Всерос. науч. конф. с междунар. участием (Санкт-Петербург, 2–4 апреля 2018 г.). Санкт-Петербург: ФГБНУ “Государственный научно-исследовательский институт озерного и речного хозяйства им. Л.С. Берга”. С. 414.
Госькова О.А. 2016. Итоги многолетнего мониторинга воспроизводства сиговых рыб в реке Сыня // Биология и биотехника разведения и состояние запасов сиговых рыб: IX Междyнар. научн.-произв. совещ. (Россия, Тюмень, 1–2 декабря 2016 г.). Тюмень: ФГБНУ “Госрыбцентр”. С. 22.
Добринская Л.А., Богданов В.Д., Богданова Е.Н. и др. 1983. Влияние экологических условий на воспроизводство сиговых рыб в уральских притоках Нижней Оби // Биологические основы рыбного хозяйства Западной Сибири. Новосибирск: Изд-во “Наука”. С. 155.
Дрягин П.А. 1948. Промысловые рыбы Обь-Иртышского бассейна // Изв. ГосНИИ озер. и реч. рыб. хоз-ва. Т. 25. Вып. 2. С. 3.
Дгебуадзе Ю.Ю. 2001. Экологические закономерности изменчивости роста рыб. Москва: Наука.
Замятин В.А. 1977. Влияние гидрологического режима на рыбные запасы р. Оби // Рыбное хозяйство Обь-Иртышского бассейна // Труды Обь-Тазовского отд. Сибирского научно-исследовательского и проектно-конструкторского института рыбного хозяйства. Нов. серия. Свердловск: Средне-Уральское книжное изд-во. Т. 4. С. 76.
Зыков Л.А. 1996. Рост и естественная смертность основных промысловых рыб Обь-Иртышского бассейна // Биологические ресурсы и проблемы развития аквакультуры на водоемах Урала и Западной Сибири: Тез. докл. Всерос. конф. (Тюмень, 17–18 сентября 1996 г.). Тюмень. С. 56.
Зыков Л.А., Слепокуров В.А. 1982. Уравнение для оценки естественной смертности рыб (на примере пеляди оз. Ендырь) // Рыбное хоз-во. № 3. С. 36.
Зубова Е.М., Кашулин Н.А., Терентьев П.М. 2015. Линейный рост малотычинкового сига Coregonus lavaretus lavaretus (L.) (Coregonidae) Бабинской Имандры (оз. Имандра) // Биология внутр. вод. № 1. С. 81.
https://doi.org/10.7868/S0320965215010155
Ивлев В.С. 1958. Эколого-физиологический анализ распределения рыб в градиентных условиях среды // Труды совещания по физиологии рыб. Москва. Вып. 8. С. 288.
Исаков П.В., Селюков А.Г. 2010. Сиговые рыбы в экосистеме Обской губы. Тюмень: Изд-во Тюменского государственного университета.
Карамушко Л.И. 2007. Биоэнергетика рыб северных морей. Москва: Наука.
Карзинкин Г.С. 1952. Основы биологической продуктивности водоемов. Москва: Пищепромиздат.
Князев И.В. 1989. Оценка прироста пеляди в пойменном водоеме Нижней Оби // Сб. научн. тр. ГосНИИ озер. и реч. рыб. хоз-ва. Вып. 305. С. 106.
Князев И.В. 2017. О моделировании скорости роста рыб при различной температуре и физиологическом температурном оптимуме // Вестник рыбохозяйственной науки. Т. 4. № 2(14). С. 4.
Князев И.В., Брусынина И.Н. 1990. О мониторинге сиговых рыб в пойме Нижней Оби // Тез. докл. IV Всесоюз. совещ. по биологии и биотехнике разведения сиговых рыб (Вологда, ноябрь 1990 г.). Ленинград. С. 88.
Князев И.В., Крохалевский В.Р. 1995. Ретроспективный анализ изменения темпа роста промысловых рыб Обь-Иртышского бассейна // Сб. научн. тр. ГосНИИ озер. и реч. рыб. хоз-ва. Ленинград. Вып. 327. С. 79.
Князев И.В., Набоков Н.А. 1988. О продукции сиговых рыб в пойменной системе Нижней Оби // Пути повышения продуктивности и рационального использования рыбных ресурсов внутренних водоемов. Тез. докл. обл. науч.-практ. конф. (Тюмень, 19–20 декабря 1988 г.). Тюмень. С. 11.
Князев И.В., Огурцова Н.Н. 1988. Некоторые особенности роста пеляди в пойме Нижней Оби // Сб. научн. тр. ГосНИИ озер. и реч. рыб. хоз-ва. Ленинград. Вып. 284. С. 73.
Крохалевский В.Р. 1983. Половое созревание и периодичность нереста обской пеляди // Биология и экология гидробионтов экосистемы Нижней Оби. Свердловск: УНЦ АН СССР. С. 93.
Кудерский Л.А. 1986. Типы популяций промысловых рыб // Динамика численности промысловых рыб. Москва: Наука. С. 231.
Кузикова В.Б. 1984. Бентофауна Ханты-Питлярского сора Нижней Оби // Вид и его продуктивность в ареале: Матер. IV Всесоюз. совещ. (Свердловск, 3–7 апреля 1984 г.). Ч. IV. Беспозвоночные. Свердловск. С. 59.
Легкодимова З.И., Сильникова Г.В., Кияшко В.В. и др. 2019. Опыт выращивания рыбопосадочного материала укрупненной массы в прудах IV рыбоводной зоны // Вестн. рыбохоз. науки. Т. 6. № 1(21). С. 74.
Лещинская А.С. 1962. Зоопланктон и бентос Обской губы // Труды Салехардского стационара УФ АН СССР. Свердловск. Вып. 2. С. 245.
Лугаськова Н.В. 1990. Половые различия показателей крови сиговых рыб Нижней оби в период размножения // Тезисы докл. IV Всесоюз. совещ. по биологии и биотехнике разведения сиговых рыб (Вологда, ноябрь 1990 г.). Ленинград. С. 51.
Лелеко Т.И. 1989. Организация и энергетические характеристики зоопланктонных сообществ в условиях замыкающего створа реки Оби // Сб. научн. тр. ГосНИИ озер. и реч. рыб. хоз-ва. Вып. 305. С. 74.
Матковский А.К. 2006. Основные закономерности динамики численности муксуна Coregonus muksun р. Обь // Вопр. рыболовства. Т. 7. № 3(27). С. 505.
Матковский А.К. 2014. Определение смертности и численности рыб с использованием стандартизированного улова, данных по селективности и интенсивности промысла // Вестник рыбохозяйственной науки. Т. 1. № 4(4). С. 35.
Матковский А.К. 2019. Происходящие изменения в ихтиоценозах Обь-Иртышского рыбохозяйственного района под влиянием антропогенных факторов и глобального потепления климата // Проблемы обеспечения экологической безопасности и устойчивое развитие арктических территорий: Матер. Всерос. конф. с междунар. участием “II Юдахинские чтения” (Архангельск, 24–28 июня 2019 г.). Архангельск. С. 488.
Матковский А.К., Ширшов В.Я., Кривенко И.А. и др. 2017. Причины гибели сибирской ряпушки в нерестовой реке Щучья бассейна Оби // Вестн. рыбохоз. науки. Т. 4. № 1(13). С. 48.
Мельниченко С.М. 1988. Содержание жира у пеляди при миграции на нерест в реке Северной Сосьве // Экологическая энергетика животных. Свердловск: УрО АН СССР. С. 118.
Мина М.В., Клевезаль Г.А. 1976. Рост животных. Москва: Наука.
Михайличенко Л.В. 1989. Сравнительный анализ динамики роста ооцитов пеляди и чира р. Маньи во время зимовки и нагульной миграции // Экологическая обусловленность фенотипа рыб: Сб. науч. тр. УрО АН СССР. Свердловск. С. 93.
Михайличенко Л.В. 1992. Анализ роста и развития ооцитов сига-пыжьяна р. Маньи во время зимовки и миграции к местам нагула // Изучение экологии водных организмов восточного Урала: Сб. науч. тр. УрО АН СССР. Свердловск. С. 58.
Москаленко Б.К. 1956. Влияние многолетних колебаний уровня реки Оби на рост, плодовитость и размножение некоторых рыб // Зоол. журн. Т. 35. Вып. 5. С. 219.
Москаленко Б.К. 1958. Биологические основы эксплуатации и воспроизводства сиговых рыб Обского бассейна // Труды Обь-Тазовского отделения ВНИОРХ. Нов. серия. Т. 1. Тюмень: Тюмен. книж. изд-во.
Никольский Г.В. 1965. Теория динамики стада рыб. Москва: Изд-во “Наука”.
Никольский Г.В. 1974. Экология рыб. Изд. 3-е, доп. Москва: Высшая школа.
Никонов Г.И. 1963. Опыт перевозки муксуна в р. Сев. Сосьву // Труды Обь-Тазовского отделения ГосНИОРХ. Нов. серия. Тюмень. Т. 3. С. 205.
Петкевич Н.В. 1981. Гематологические особенности производителей пеляди и сига-пыжьяна р. Маньи // Второе Всесоюз. совещ. по биологии и биотехнике разведения сиговых рыб (Петрозаводск, октябрь 1981 г.). Петрозаводск. С. 77.
Решетников Ю.С. 1966. Особенности роста и созревания сигов в водоемах Севера // Закономерности динамики численности рыб Белого моря и его бассейна. Москва: Изд-во “Наука”. С. 93.
Решетников Ю.С., Белянина Т.Н., Паранюшкина Л.П. 1971. Характер жиронакопления и созревание сигов // Закономерности роста и созревания рыб. Москва: Изд-во “Наука”. С. 60.
Рыжков Л.П. 1976. Морфофизиологические закономерности и трансформация вещества и энергии в раннем онтогенезе пресноводных лососевых рыб. Петрозаводск: Карелия.
Савчук П.Ю., Матковский А.К., Перепелин Ю.В. и др. 2017. Сравнительный анализ линейного роста арктического омуля Coregonus autumnalis autumnalis (Coregonidae) из различных районов Баренцева и Карского морей // Вопр. рыболовства. Т. 18. № 3. С. 336.
Салазкин А.А. 1976. Основные типы озер гумидной зоны СССР и их биолого-продукционная характеристика // Изв. ГосНИИ озер. и реч. рыб. хоз-ва. Т. 108. Ленинград.
Семенова Л.А., Лелеко Т.И., Алексюк В.А. 1989. Сток планктона Нижней Оби // Сб. научн. тр. ГосНИИ озер. и реч. рыб. хоз-ва. Вып. 305. С. 56.
Силин В.В. 1990. Об изменении популяционной структуры муксуна р. Колымы // Тезисы докл. четвертого Всесоюз. совещ. по биологии и биотехнике разведения сиговых рыб (Вологда, ноябрь 1990 г.). Ленинград. С. 98.
Смирнов А.К., Голованов В.К., Голованова И.Л., Артаев О.Н. 2020. Избираемая температура и температурная устойчивость озерного гольяна Rhynchocypris percnurus // Биология внутр. вод. № 2. С. 156.
https://doi.org/10.31857/S0320965220020138
Тюрин П.В. 1972. “Нормальные” кривые переживания и темпов естественной смертности рыб как теоретическая основа регулирования рыболовства // Тр. ГосНИИ озер. и реч. рыб. хоз-ва. Т. 71. Ленинград. С. 71.
Чаликов Б.Г. 1931. Из биологии муксуна бассейна р. Оби в связи с вопросом его охраны // Тр. Томского университета. Томск: Биологический факультет Томского гос. ун-та. Т. 3. С. 316.
Чугунова Н.И. 1959. Руководство по изучению возраста и роста рыб. Москва: Изд-во АН СССР.
Шатуновский М.И. 1980. Экологические закономерности обмена веществ морских рыб. Москва: Изд-во “Наука”.
Шестаков А.В. 2021. Размерно-возрастная структура и рост сиговых рыб (Coregonidae) арктического оз. Иони (Восточная Чукотка) // Биология внутр. вод. № 2. С. 171. https://doi.org/10.31857/S0320965221010125
Шибаев С.В. 2014. Промысловая ихтиология. Калининград: ООО “Аксиос”.
Шмальгаузен И.И. 1935. Определение основных понятий и методика исследования роста // Рост животных. Москва: Биомедгиз. С. 8.
Яржомбек А.А., Кляшторин Л.Б., Подражанская С.Г. 1977. Активный обмен у рыб // Экология. № 1. С. 65.
Matkovskiy A.K. 2014. The influence of the hydrological regime on populations of whitefish in the Ob basin //12-th International Symposium on the Biology and Management of Coregonid fishes (25–30 August, 2014, Irkutsk, Listvyancka, Russia). P. 50.
Rijnsdorp A.D., Grift R.E., Kraak S.B.M. 2005. Fisheries-induced adaptive change in reproductive investment in North Sea plaice (Pleuronectes platessa)? // Can. J. Fish. Aquat. Sci. V. 62. P. 833. https://doi.org/10.1139/F05-039
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод