

Биология внутренних вод, 2022, № 6, стр. 793-802
Динамика структуры сообщества поденок (Insecta, Ephemeroptera) малой лососевой реки (Южное Приморье, Россия)
Е. А. Горовая *
Федеральный научный центр биоразнообразия наземной биоты Восточной Азии
Дальневосточного отделения Российской академии наук
Владивосток, Россия
* E-mail: brouny@mail.ru
Поступила в редакцию 17.05.2022
После доработки 30.05.2022
Принята к публикации 17.06.2022
- EDN: EPLLVQ
- DOI: 10.31857/S0320965222060079
Аннотация
Приведены сведения об изменении структуры сообщества поденок малой лососевой р. Тигровая в условиях различных гидрологических режимов и слабого антропогенного воздействия. В водотоке отмечено обитание ~40 видовых таксонов отр. Ephemeroptera. В бентосном сообществе на грунтах с фракцией <22 см личинки поденок занимали лидирующую позицию, их численность достигала 176–8601 экз./м2, биомасса 0.38–19.67 г/м2. Минимальные значения отмечены во время летнего паводка, максимум по численности – в ледовый период, по биомассе – в весенне-летнюю межень. На исследованном участке р. Тигровая зарегистрированы виды поденок, относящиеся к пяти экологическим группам. Ведущую роль в формировании сообщества поденок на протяжении года играли представители сем. Heptageniidae и Ephemerellidae, преимущественно, родов Cinygmula и Drunella. В ряде случаев высокие показатели количественных характеристик достигнуты за счет нимф и личинок ранних возрастов.
ВВЕДЕНИЕ
Начиная с середины ХХ в., одной из приоритетных задач гидробиологических исследований был сбор, накопление и анализ данных по видовому составу и количественным характеристикам зообентоса. Изучение состояния придонных сообществ, обладающих определенной экологической целостностью, привело к разработке и широкому применению биоиндикационных методов комплексной оценки антропогенного загрязнения и определило особое положение амфибиотических насекомых, в частности, поденок, в системе экологического мониторинга. Установлено, что рекам различной типологической принадлежности свойственна определенная структура бентосного сообщества, меняющаяся в зависимости от типа и зоны реки, сезона и широтности (Тиунова, 2007, Чертопруд, 2021). В связи с этим, стало очевидным исключительное положение малых рек, находящихся в тесной связи с окружающим их ландшафтом, быстро реагирующих на процессы, происходящие на малом водосборе (Ткачев, Булатов, 2002). На территории Дальнего Востока России данная категория водотоков преимущественно является местом нереста и нагула молоди лососевых рыб, что определяет их высокую значимость (Леванидова и др., 1989). Интерес к проведению исследовательских работ, в первую очередь, вызывают реки, не подверженные антропогенной нагрузке или испытывающие очень слабое ее воздействие, что позволяет создавать паспорта “чистых” рек и использовать данные в качестве эталонных. В связи с этим, основное внимание уделяется водотокам, расположенным на охраняемых природных территориях. Однако большинство малых рек густой гидрологической сети региона испытывает значительное воздействие хозяйственной деятельности человека, сохраняя при этом статус лососевых, а данные об их состоянии разрозненны, носят случайный характер, отсутствуют фоновые сведения, в том числе, о циклических изменениях сообществ. Учитывая перманентность трансформаций, происходящих в биоценозах под действием антропогенных и естественных природных факторов, изучение процессов формирования, а также, сезонных и географических изменений сообществ водотоков различной типовой принадлежности, по-прежнему актуально. В работе впервые приводятся сведения о годовой динамике количественных характеристик сообщества поденок в условиях малой лососевой р. Тигровая (Партизанский городской округ, Приморский край), отражающие фоновое состояние водотока при незначительном антропогенном воздействии.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
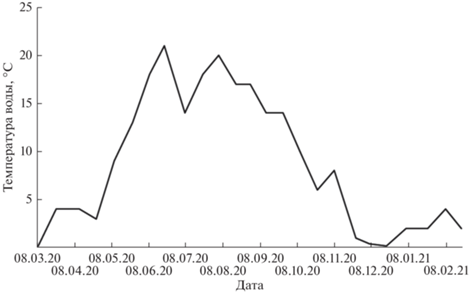
Река Тигровая, протяженностью 53 км, берет свой исток на северо-восточном склоне хребта Ливадийский. Протекая по горно-сопочной местности, почти сплошь покрытой смешанным лесом, она впадает в р. Партизанская справа. Площадь водосбора р. Тигровая – 698 км2, его залесенность 98%. Русло извилистое, неустойчивое, разделяется на множество проток и рукавов. Плесы и перекаты чередуются через 100–200 м. Глубина на перекатах ≤0.6–0.7 м, на плесах – ≤1.0–1.5 м. Наблюдается чередование двух–семи резких подъемов в 2–3 сут и спадов в 5–10 сут, с мая по сентябрь. Уровень воды снижается в конце октября. Первый лед появляется во второй декаде ноября, что сопровождается повышением уровня воды до максимума в январе–феврале. Наибольшая толщина льда 100 см. Весенний подъем уровня воды происходит в конце марта–начале апреля. Переход температуры воды через 0.2°C происходил в первой декаде апреля; температурный максимум (до 28°C) – в августе11.
Выбор места отбора проб на р. Тигровая обусловлен наличием незамерзающей полыньи близ впадения одного из ее основных притоков – р. Серебрянка (рис. 1). Выше по течению располагаются села Тигровой, Фридман (на притоке р. Моленная), Бровничи, Серебряное (на притоке р. Серебрянка). Данный участок поймы, шириной до 400 м, доступен для въезда автотранспорта и активно используется как пикниковая зона, посещается рыбаками. Во время паводков поверхность поймы полностью затопляется, смывая кострища, техногенные жидкости и немногочисленный мусор. Русло реки здесь прямолинейное, сравнительно устойчивое, галечное и песчаное на плесах, с мелкими валунами на перекатах и крупными – на сливах. Весной 2020 г. после схода льда, по всей пойме наблюдали интенсивное крошение камней.

За годовой период температура воды на исследуемом участке изменялась в диапазоне 0.2–21°С (рис. 2), скорость течения 0.1–2 м/с (исключение – паводок 30 августа). Значительный подъем уровня воды также отмечен в половодье (23 марта) и после выпадения обильных осадков (7 и 22 июля, 24 октября). Межень наблюдали в мае–июне и октябре. Первые ледовые явления отмечены 25 ноября и к 20 декабря река была полностью покрыта льдом, а к 20 февраля началось ее вскрытие. Таким образом, р. Тигровая – водоток умеренно-холодноводного типа и по классификации Леванидова (1969), исследованный участок можно отнести к III категории (средние участки горных и предгорных рек и предгорные речки).
Материал собирали с периодичностью в 2 нед. Единовременно отбирали по одной качественной и по три количественные пробы (на плесе, сливе и перекате), 23 марта и 30 августа 2020 г. – одна и две пробы соответственно, вне грунтов постоянного русла. В работе использовали гидробиологический сачок и модифицированный бентометр Леванидова с площадью захвата 0.0484 м2, применявшийся на глубине до 50 см на грунтах, состоявших из песка, гальки и мелких валунов (≤22 см). Грунт взмульчивали на глубину до 15 см. Всего отобрано 73 количественных и 19 качественных проб, зафиксированных 96%-ным этанолом.
В результате первичной сортировки получены сведения о количественных характеристиках зообентоса. Для последующего изучения таксономического состава сообщества поденок использовали только пригодный для определения личинок. Это привело к появлению расхождений в значениях, полученных при первичной сортировке и после суммирования результатов по отдельным таксонам сообщества поденок. Таким образом, использованные в работе процентные показатели долевого вклада таксономических групп отр. Ephemeroptera, отображают численность и биомассу только идентифицированных личинок.
Для анализа структуры сообщества поденок и зообентоса в целом использовали классификацию А.М. Чельцова-Бебутова в модификации В.Я. Леванидова (1977): доминанты – ≥15% общей численности и биомассы, субдоминанты – 5.0–14.9%, второстепенные виды – 1–4.9%, третьестепенные – <0.1%.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
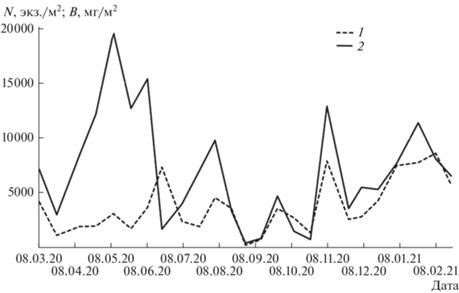
На исследованном участке р. Тигровая на протяжении годового периода численность зообентоса составляла 258–17 596 экз./м2, биомасса – 0.38–26.58 г/м2. Доминантами по долевому вкладу были представители отр. Ephemeroptera (рис. 3) с минимальным показателем численности 176 экз./м2 во время паводка 30 августа и максимальным – 8601 экз./м2 в ледовый период 7 февраля (рис. 4). Биомасса изменялась от 0.38 г/м2 30 августа до 19.67 г/м2 в период межени 10 мая. Начиная с апреля и по первую половину июня включительно, на фоне относительно небольшой численности поденок в водотоке наблюдали увеличение их биомассы. В пробах от 20 июня, 7 и 22 июля отмечены диаметрально противоположные соотношения, с августа по март изменения количественных показателей было однонаправленным. Подобная динамика во многом связана с гидрологическим режимом водотока. Так, резкое снижение количественных показателей происходило во время половодья 23 марта, паводков 30 августа и 24 октября, осенней межени 10 октября. Во время июльского подъема уровня воды численность сообщества уменьшалась, биомасса – увеличивалась. В период весенней межени динамические изменения претерпевали обе характеристики, однако показатели биомассы достигли абсолютного зарегистрированного максимума и были значительными вплоть до 20 июня. С наступлением ледового периода и до 7 февраля отмечен рост численности поденок и их биомассы.
Рис. 3.
Динамика изменения доли численности (1) и биомассы (2) личинок поденок в общих количественных показателях зообентоса р. Тигровая.
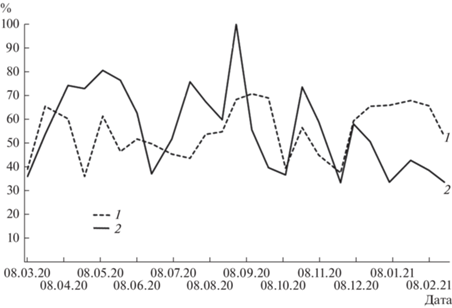
Рис. 4.
Динамика численности (N) и биомассы (B) сообщества поденок в р. Тигровая. 1 – численность; 2 – биомасса.
Фауна поденок была представлена личинками различных возрастов, в том числе зрелыми формами, многочисленными нимфами, а также экземплярами со слабой хитинизацией, невыраженной окраской, поврежденными структурами, выступающими в роли определительных признаков. В ряде случаев это затруднило идентификацию и привело к составлению таблицы, включающей 49 таксонов в ранге вида, рода и на уровне семейства (табл. 1). В каждую из дат отбора в сообществе поденок отмечали стабильно высокое число таксономических групп: от 11 до 23 (исключение – 8 групп в паводок 30 августа).
Таблица 1.
Доли (%) численности (числитель) и биомассы (знаменатель) таксонов в сообществе поденок (март 2020–февраль 2021 г.)
| Таксон, группа |
2020 | 2021 | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 08.03 | 23.03 | 11.04 | 25.04 | 10.05 | 25.05 | 08.06 | 20.06 | 07.07 | 22.07 | 04.08 | 18.08 | 30.08 | 12.09 | 26.09 | 10.10 | 24.10 | 07.11 | 25.11 | 06.12 | 20.12 | 05.01 | 23.01 | 07.02 | 20.02 | |
| Ephemera strigata | – | – | $\frac{{3.79}}{{{\mathbf{27}}{\mathbf{.06}}}}$ | $\frac{{1.23}}{{2.87}}$ | $\frac{{{\text{0}}{\text{.94}}}}{{0.72}}$ | $\frac{{{\text{1}}{\text{.86}}}}{{1.98}}$ | * | $\frac{{{\text{0}}{\text{.2}}}}{{4.22}}$ | $\frac{{{\text{0}}{\text{.86}}}}{{9.58}}$ | – | $\frac{{{\text{0}}{\text{.19}}}}{{6.63}}$ | $\frac{{{\text{3}}{\text{.65}}}}{{4.86}}$ | – | – | $\frac{{{\text{8}}{\text{.51}}}}{{14.24}}$ | – | – | – | – | – | – | – | – | – | – |
| Ephemera spp. | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.91}}}}{{1.21}}$ | – | $\frac{{{\text{1}}{\text{.43}}}}{{0.03}}$ | $\frac{{{\text{7}}{\text{.63}}}}{{0.05}}$ | $\frac{{{\text{8}}{\text{.7}}}}{{1.46}}$ | $\frac{{{\text{3}}{\text{.22}}}}{{1.07}}$ | $\frac{{12.5}}{{3.33}}$ | $\frac{{{\text{1}}{\text{.82}}}}{{4.63}}$ | $\frac{{{\text{0}}{\text{.61}}}}{{0.4}}$ | $\frac{{{\text{1}}{\text{.62}}}}{{4.34}}$ | – | $\frac{{{\text{0}}{\text{.55}}}}{{0.7}}$ | $\frac{{{\text{2}}{\text{.91}}}}{{{\mathbf{17}}{\mathbf{.01}}}}$ | $\frac{{{\text{0}}{\text{.52}}}}{{0.59}}$ | $\frac{{{\text{0}}{\text{.67}}}}{{0.06}}$ | $\frac{{{\text{0}}{\text{.11}}}}{{0.02}}$ | $\frac{{{\text{0}}{\text{.09}}}}{{0.02}}$ | – | – |
| Cinygmula hirasana | $\frac{{{\mathbf{15}}{\mathbf{.34}}}}{{9.89}}$ | $\frac{{{\mathbf{19}}{\mathbf{.06}}}}{{8.85}}$ | $\frac{{{\mathbf{50}}{\mathbf{.38}}}}{{{\mathbf{34}}{\mathbf{.86}}}}$ | $\frac{{{\mathbf{34}}{\mathbf{.43}}}}{{{\mathbf{24}}{\mathbf{.47}}}}$ | * | $\frac{{{\text{0}}{\text{.93}}}}{{1.71}}$ | – | – | – | – | – | – | – | – | – | $\frac{{{\text{1}}{\text{.89}}}}{{7.4}}$ | $\frac{{{\text{1}}{\text{.05}}}}{{{\mathbf{17}}{\mathbf{.51}}}}$ | – | – | – | – | – | – | – | – |
| C. kurenzovi | $\frac{{{\text{0}}{\text{.18}}}}{{0.31}}$ | – | * | – | $\frac{{{\mathbf{32}}{\mathbf{.07}}}}{{{\mathbf{45}}{\mathbf{.46}}}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.39}}}}{{7.17}}$ |
| С. sapporensis | $\frac{{{\text{1}}{\text{.44}}}}{{{\mathbf{40}}{\mathbf{.2}}}}$ | $\frac{{8.49}}{{{\mathbf{74}}{\mathbf{.7}}}}$ | $\frac{{{\text{0}}{\text{.38}}}}{{4.04}}$ | $\frac{{{\mathbf{19}}{\mathbf{.67}}}}{{{\mathbf{24}}{\mathbf{.39}}}}$ | $\frac{{{\text{0}}{\text{.24}}}}{{0.14}}$ | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.52}}}}{{4.82}}$ | $\frac{{\text{1}}}{{8.56}}$ | $\frac{{{\text{0}}{\text{.58}}}}{{5.99}}$ | * | $\frac{{{\text{0}}{\text{.5}}}}{{7.04}}$ | $\frac{{{\text{0}}{\text{.53}}}}{{14.09}}$ | – | $\frac{{{\text{0}}{\text{.57}}}}{{11.04}}$ | $\frac{{{\text{0}}{\text{.13}}}}{{3.94}}$ |
| Cinygmula spp. | – | – | – | – | * | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\mathbf{84}}{\mathbf{.3}}}}{{{\mathbf{60}}{\mathbf{.}}{\mathbf{86}}}}$ | $\frac{{{\mathbf{55}}{\mathbf{.07}}}}{{13.01}}$ | $\frac{{{\mathbf{38}}{\mathbf{.95}}}}{{{\mathbf{22}}{\mathbf{.73}}}}$ | $\frac{{{\mathbf{64}}{\mathbf{.84}}}}{{{\mathbf{18}}{\mathbf{.6}}}}$ | $\frac{{{\mathbf{72}}{\mathbf{.73}}}}{{{\mathbf{45}}}}$ | $\frac{{{\mathbf{69}}{\mathbf{.01}}}}{{{\mathbf{43}}{\mathbf{.98}}}}$ | $\frac{{{\mathbf{66}}{\mathbf{.3}}}}{{{\mathbf{43}}{\mathbf{.52}}}}$ | $\frac{{{\mathbf{82}}{\mathbf{.42}}}}{{{\mathbf{66}}{\mathbf{.92}}}}$ | $\frac{{{\mathbf{71}}{\mathbf{.36}}}}{{{\mathbf{47}}{\mathbf{.45}}}}$ |
| Ecdyonurus asperses | – | – | – | – | – | – | – | – | * | $\frac{{{\text{4}}{\text{.96}}}}{{5.47}}$ | $\frac{{{\text{1}}{\text{.85}}}}{{14.71}}$ | – | * | – | – | – | – | – | – | – | – | – | – | – | – |
| E. bajkovae | – | – | $\frac{{{\text{1}}{\text{.89}}}}{{{\text{0}}{\text{.48}}}}$ | $\frac{{{\text{0}}{\text{.82}}}}{{0.11}}$ | $\frac{{{\text{0}}{\text{.71}}}}{{0.09}}$ | $\frac{{{\text{0}}{\text{.93}}}}{{0.81}}$ | $\frac{{{\text{2}}{\text{.04}}}}{{6.06}}$ | $\frac{{{\text{0}}{\text{.31}}}}{{0.76}}$ | – | – | $\frac{{6.67}}{{0.13}}$ | $\frac{{8.37}}{{1.07}}$ | $\frac{{{\mathbf{18}}{\mathbf{.}}{\mathbf{75}}}}{{2.22}}$ | $\frac{{7.27}}{{2.31}}$ | $\frac{{{\text{0}}{\text{.81}}}}{{0.37}}$ | $\frac{{{\text{3}}{\text{.51}}}}{{8.68}}$ | $\frac{{{\text{1}}{\text{.05}}}}{{2.63}}$ | $\frac{{{\text{1}}{\text{.37}}}}{{1.6}}$ | $\frac{{{\text{1}}{\text{.74}}}}{{1.22}}$ | $\frac{{{\text{0}}{\text{.26}}}}{{0.12}}$ | – | $\frac{{{\text{0}}{\text{.21}}}}{{0.08}}$ | – | $\frac{{0.08}}{{0.01}}$ | $\frac{{0.26}}{{0.01}}$ |
| E. abracadabrus | – | – | – | – | – | – | – | – | – | * | – | $\frac{{6.65}}{{{\mathbf{39}}{\mathbf{.}}{\mathbf{06}}}}$ | – | * | – | – | – | – | – | – | – | – | – | – | – |
| Ecdyonurus spp. | – | – | – | – | – | – | $\frac{{{\text{1}}{\text{.13}}}}{{{\text{0}}{\text{.39}}}}$ | $\frac{{{\text{0}}{\text{.91}}}}{{1.47}}$ | – | – | – | – | $\frac{{6.25}}{{13.33}}$ | $\frac{{{\text{1}}{\text{.82}}}}{{0.77}}$ | – | – | – | – | – | – | – | – | – | – | – |
| Epeorus pellucidus | $\frac{{{\text{0}}{\text{.54}}}}{{0.1}}$ | $\frac{{{\text{0}}{\text{.95}}}}{{1.82}}$ | * | – | – | $\frac{{{\text{0}}{\text{.93}}}}{{0.62}}$ | – | – | – | $\frac{{12.59}}{{12.2}}$ | $\frac{{{\text{4}}{\text{.81}}}}{{{\mathbf{30}}{\mathbf{.}}{\mathbf{51}}}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{9.16}}$ | – | * | $\frac{{{\text{0}}{\text{.4}}}}{{3.25}}$ | * | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.37}}}}{{0.11}}$ | $\frac{{{\text{0}}{\text{.33}}}}{{0.03}}$ | $\frac{{{\text{0}}{\text{.39}}}}{{0.01}}$ |
| Epeorus spp. | * | $\frac{{{\text{0}}{\text{.95}}}}{{0.05}}$ | $\frac{{{\text{0}}{\text{.38}}}}{{0.01}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.02}}$ | – | – | – | $\frac{{{\mathbf{16}}{\mathbf{.}}{\mathbf{09}}}}{{0.56}}$ | $\frac{{{\mathbf{28}}{\mathbf{.}}{\mathbf{08}}}}{{11.91}}$ | $\frac{{{\text{2}}{\text{.67}}}}{{1.26}}$ | $\frac{{{\mathbf{20}}{\mathbf{.}}{\mathbf{37}}}}{{13.52}}$ | $\frac{{13.09}}{{12.16}}$ | – | $\frac{{5.46}}{{2.69}}$ | $\frac{{{\text{2}}{\text{.83}}}}{{5.5}}$ | $\frac{{5.41}}{{13.27}}$ | $\frac{{{\text{1}}{\text{.57}}}}{{0.87}}$ | $\frac{{{\text{1}}{\text{.09}}}}{{1.77}}$ | $\frac{{{\text{0}}{\text{.29}}}}{{0.03}}$ | $\frac{{{\text{0}}{\text{.5}}2}}{{0.01}}$ | $\frac{{{\text{0}}{\text{.5}}}}{{0.06}}$ | $\frac{{{\text{1}}{\text{.06}}}}{{0.05}}$ | $\frac{{{\text{0}}{\text{.37}}}}{{0.02}}$ | $\frac{{{\text{0}}{\text{.08}}}}{{0.01}}$ | – |
| Heptagenia sp. | – | – | – | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.2}}}}{{1.25}}$ | – | – | – | – | – | – | – | – | – | – |
| Rhithrogena sp. | $\frac{{{\text{0}}{\text{.36}}}}{{0.17}}$ | – | – | $\frac{{{\text{4}}{\text{.1}}}}{{0.26}}$ | $\frac{{{\text{4}}{\text{.48}}}}{{0.49}}$ | $\frac{{7.44}}{{2.7}}$ | $\frac{{9.07}}{{8.75}}$ | $\frac{{{\text{3}}{\text{.56}}}}{{10.09}}$ | $\frac{{{\text{2}}{\text{.87}}}}{{6.9}}$ | $\frac{{{\mathbf{17}}{\mathbf{.}}{\mathbf{18}}}}{{11.21}}$ | $\frac{{6.85}}{{8.44}}$ | $\frac{{{\text{0}}{\text{.43}}}}{1}$ | – | $\frac{{{\text{10}}}}{{{\mathbf{19}}{\mathbf{.}}{\mathbf{27}}}}$ | $\frac{{{\text{2}}{\text{.43}}}}{{12.24}}$ | – | – | $\frac{{{\text{1}}{\text{.46}}}}{{1.58}}$ | $\frac{{{\text{1}}{\text{.45}}}}{{0.54}}$ | $\frac{{{\text{3}}{\text{.39}}}}{{3.59}}$ | $\frac{{{\text{2}}{\text{.19}}}}{{2.41}}$ | $\frac{{{\text{0}}{\text{.74}}}}{{1.94}}$ | $\frac{{{\text{0}}{\text{.74}}}}{1}$ | $\frac{{{\text{0}}{\text{.49}}}}{{0.9}}$ | $\frac{{{\text{1}}{\text{.3}}}}{{1.45}}$ |
| Heptageniidae | $\frac{{{\text{0}}{\text{.18}}}}{{0.03}}$ | – | – | – | – | $\frac{{6.51}}{{0.2}}$ | $\frac{{12.7}}{{0.3}}$ | $\frac{{{\mathbf{16}}{\mathbf{.19}}}}{{1.22}}$ | $\frac{{{\mathbf{20}}{\mathbf{.06}}}}{{4.93}}$ | $\frac{{9.54}}{{0.49}}$ | $\frac{5}{{9.47}}$ | – | – | $\frac{{{\text{3}}{\text{.64}}}}{{0.77}}$ | $\frac{{13.97}}{{2.25}}$ | $\frac{{{\mathbf{67}}{\mathbf{.57}}}}{{{\mathbf{29}}{\mathbf{.86}}}}$ | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.16}}}}{{0.01}}$ | – |
| Isonychia sp. | – | – | – | – | – | – | $\frac{{{\text{3}}{\text{.17}}}}{{0.19}}$ | $\frac{{{\text{1}}{\text{.43}}}}{{0.45}}$ | $\frac{{{\text{0}}{\text{.86}}}}{{0.85}}$ | $\frac{{{\text{0}}{\text{.76}}}}{{5.33}}$ | – | * | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Ameletus sp. | $\frac{{{\text{0}}{\text{.18}}}}{{0.03}}$ | $\frac{{{\text{0}}{\text{.95}}}}{{0.05}}$ | $\frac{{{\text{0}}{\text{.38}}}}{{0.03}}$ | – | * | $\frac{{{\text{1}}{\text{.86}}}}{{0.09}}$ | – | – | – | – | $\frac{{{\text{0}}{\text{.37}}}}{{0.01}}$ | $\frac{{{\text{0}}{\text{.21}}}}{{0.03}}$ | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.65}}}}{{0.07}}$ | – | – |
| Baetis (Ni-grobaetis) sp. | $\frac{{{\text{0}}{\text{.54}}}}{{0.07}}$ | $\frac{{{\text{2}}{\text{.86}}}}{{0.23}}$ | – | – | – | – | * | – | – | – | $\frac{{{\text{0}}{\text{.37}}}}{{0.18}}$ | $\frac{{{\text{0}}{\text{.43}}}}{{0.14}}$ | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.26}}}}{{0.02}}$ | – | – | – | – | – |
| B. (N.) bacillus | – | – | – | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.2}}}}{{0.5}}$ | – | $\frac{{{\text{0}}{\text{.52}}}}{{0.44}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{0.24}}$ | $\frac{{{\text{1}}{\text{.45}}}}{{0.34}}$ | – | $\frac{{{\text{0}}{\text{.67}}}}{{0.09}}$ | $\frac{{{\text{0}}{\text{.42}}}}{{0.08}}$ | – | – | – |
| B. fuscatus | – | – | – | – | – | – | $\frac{{8.16}}{{2.5}}$ | $\frac{{{\text{3}}{\text{.87}}}}{{1.22}}$ | – | * | $\frac{{{\text{0}}{\text{.37}}}}{{0.36}}$ | $\frac{{{\text{2}}{\text{.36}}}}{{2.22}}$ | $\frac{{{\mathbf{18}}{\mathbf{.75}}}}{{6.67}}$ | $\frac{{8.18}}{{11.18}}$ | $\frac{{{\text{0}}{\text{.2}}}}{{0.25}}$ | – | – | – | – | – | – | – | – | – | – |
| B. pseudothermicus | $\frac{{{\text{5}}{\text{.06}}}}{{3.35}}$ | $\frac{{{\text{3}}{\text{.81}}}}{{1.36}}$ | $\frac{{{\text{1}}{\text{.14}}}}{{0.48}}$ | – | * | $\frac{{{\text{3}}{\text{.72}}}}{{0.09}}$ | – | * | – | – | – | * | – | – | * | – | – | $\frac{{{\text{0}}{\text{.46}}}}{{0.09}}$ | – | $\frac{{{\text{1}}{\text{.3}}}}{{0.09}}$ | $\frac{{{\text{1}}{\text{.01}}}}{{0.37}}$ | $\frac{{7.32}}{{4.84}}$ | $\frac{{{\text{0}}{\text{.28}}}}{{0.14}}$ | $\frac{{{\text{1}}{\text{.88}}}}{{0.9}}$ | $\frac{{8.16}}{{6.6}}$ |
| Acentrella sibiricum | – | – | – | – | $\frac{{{\text{0}}{\text{.24}}}}{{0.14}}$ | – | – | $\frac{{{\text{0}}{\text{.51}}}}{{0.23}}$ | – | $\frac{{{\text{1}}{\text{.15}}}}{{0.21}}$ | $\frac{{6.11}}{{0.87}}$ | $\frac{{{\text{4}}{\text{.51}}}}{{0.57}}$ | $\frac{{6.25}}{{8.9}}$ | * | $\frac{{{\text{0}}{\text{.4}}}}{{0.08}}$ | – | – | $\frac{{{\text{0}}{\text{.09}}}}{{0.13}}$ | – | – | – | – | – | – | – |
| Acentrella gr. sibirica | – | – | – | – | – | $\frac{{6.05}}{{0.18}}$ | $\frac{{{\text{1}}{\text{.59}}}}{{0.17}}$ | – | $\frac{{9.17}}{{0.85}}$ | – | – | – | – | $\frac{{6.36}}{{1.16}}$ | $\frac{{{\text{0}}{\text{.81}}}}{{0.11}}$ | – | – | – | – | – | – | – | – | – | – |
| Baetiella tuberculatum | – | – | – | – | – | – | $\frac{{5.22}}{{0.17}}$ | $\frac{{{\text{3}}{\text{.77}}}}{{0.54}}$ | * | – | $\frac{{{\text{1}}{\text{.11}}}}{{0.18}}$ | * | – | $\frac{{{\text{0}}{\text{.91}}}}{{0.39}}$ | – | – | – | – | – | – | – | – | – | – | – |
| Cloeon sp. | – | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\mathbf{25}}}}{{{\mathbf{62}}{\mathbf{.22}}}}$ | – | – | – | – | – | – | – | – | – | – | – | – |
| Baetidae | $\frac{{6.14}}{{0.68}}$ | – | $\frac{{{\text{0}}{\text{.76}}}}{{0.05}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.07}}$ | – | – | $\frac{{9.07}}{{0.39}}$ | $\frac{{{\text{0}}{\text{.2}}}}{{0.05}}$ | – | $\frac{{7.25}}{{0.49}}$ | $\frac{{{\text{2}}{\text{.04}}}}{{0.24}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{0.29}}$ | * | $\frac{{{\text{1}}{\text{.82}}}}{{1.31}}$ | $\frac{{{\text{1}}{\text{.21}}}}{{1.31}}$ | $\frac{{{\text{0}}{\text{.81}}}}{{1.53}}$ | $\frac{{{\text{2}}{\text{.62}}}}{{1.49}}$ | $\frac{{{\text{0}}{\text{.18}}}}{{0.03}}$ | $\frac{{{\text{0}}{\text{.58}}}}{{0.07}}$ | – | – | – | $\frac{{{\text{0}}{\text{.09}}}}{{0.01}}$ | – | $\frac{{{\text{0}}{\text{.52}}}}{{0.01}}$ |
| Choroterpes sp. | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.45}}}}{{0.35}}$ | $\frac{{{\text{0}}{\text{.1}}}}{{0.23}}$ | – | * | – | – | – | – | – | $\frac{{{\text{0}}{\text{.54}}}}{{0.25}}$ | $\frac{{{\text{1}}{\text{.05}}}}{{0.87}}$ | $\frac{{{\text{0}}{\text{.18}}}}{{0.01}}$ | – | – | – | – | – | – | – |
| Neoleptophlebia japonica | * | $\frac{{7.63}}{{9.3}}$ | $\frac{{{\text{0}}{\text{.76}}}}{{0.7}}$ | $\frac{{{\text{0}}{\text{.82}}}}{{0.5}}$ | * | – | $\frac{{{\text{1}}{\text{.36}}}}{{0.46}}$ | $\frac{{{\text{4}}{\text{.79}}}}{{2.16}}$ | * | $\frac{{{\text{1}}{\text{.91}}}}{{1.61}}$ | * | – | $\frac{{6.25}}{{1.11}}$ | $\frac{{10.91}}{{8.87}}$ | $\frac{{5.67}}{2}$ | $\frac{{{\text{4}}{\text{.87}}}}{{6.89}}$ | $\frac{{{\text{1}}{\text{.57}}}}{{1.75}}$ | $\frac{{{\text{3}}{\text{.83}}}}{{7.82}}$ | $\frac{{{\text{2}}{\text{.33}}}}{{2.72}}$ | $\frac{{{\text{2}}{\text{.86}}}}{{4.85}}$ | $\frac{{{\text{1}}{\text{.85}}}}{{3.01}}$ | $\frac{{{\text{0}}{\text{.85}}}}{{1.79}}$ | $\frac{{{\text{1}}{\text{.57}}}}{{2.46}}$ | $\frac{{{\text{0}}{\text{.16}}}}{{0.1}}$ | $\frac{{{\text{0}}{\text{.26}}}}{{0.19}}$ |
| N. vladivostokica | – | – | * | – | – | – | $\frac{{{\text{3}}{\text{.63}}}}{{1.52}}$ | $\frac{{5.09}}{{2.36}}$ | $\frac{{{\text{0}}{\text{.86}}}}{{1.13}}$ | * | – | – | – | – | $\frac{{{\text{2}}{\text{.43}}}}{{0.75}}$ | $\frac{{{\text{1}}{\text{.89}}}}{{4.6}}$ | $\frac{{{\text{0}}{\text{.52}}}}{{1.31}}$ | $\frac{{{\text{1}}{\text{.64}}}}{{3.39}}$ | – | – | – | – | – | – | – |
| Neoleptophlebia spp. | – | – | – | – | $\frac{{{\text{0}}{\text{.24}}}}{{0.01}}$ | $\frac{{{\text{0}}{\text{.93}}}}{{0.04}}$ | – | – | – | $\frac{{{\text{3}}{\text{.82}}}}{{0.04}}$ | $\frac{{{\text{0}}{\text{.56}}}}{{0.02}}$ | $\frac{{{\text{4}}{\text{.51}}}}{{0.57}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Drunella aculea | $\frac{{{\text{0}}{\text{.72}}}}{{{\mathbf{20}}{\mathbf{.8}}}}$ | – | * | $\frac{{{\text{3}}{\text{.28}}}}{{{\mathbf{42}}{\mathbf{.94}}}}$ | $\frac{{{\text{1}}{\text{.89}}}}{{{\mathbf{36}}{\mathbf{.97}}}}$ | $\frac{{{\text{2}}{\text{.33}}}}{{{\mathbf{63}}{\mathbf{.2}}}}$ | * | – | – | $\frac{{{\text{0}}{\text{.38}}}}{{1.96}}$ | – | $\frac{{{\text{2}}{\text{.36}}}}{{0.57}}$ | – | $\frac{{6.36}}{{14.65}}$ | $\frac{{{\text{7}}{\text{.89}}}}{{{\mathbf{19}}{\mathbf{.49}}}}$ | $\frac{{{\text{0}}{\text{.81}}}}{{7.66}}$ | – | $\frac{{\text{1}}}{{{\mathbf{19}}{\mathbf{.34}}}}$ | $\frac{{{\text{0}}{\text{.29}}}}{{4.02}}$ | $\frac{{{\text{2}}{\text{.6}}}}{{{\mathbf{44}}{\mathbf{.24}}}}$ | $\frac{{{\text{0}}{\text{.84}}}}{{{\mathbf{26}}{\mathbf{.16}}}}$ | $\frac{{{\text{0}}{\text{.32}}}}{{13}}$ | $\frac{{{\text{1}}{\text{.1}}}}{{{\mathbf{41}}{\mathbf{.74}}}}$ | $\frac{{{\text{0}}{\text{.33}}}}{{13.71}}$ | $\frac{{{\text{0}}{\text{.78}}}}{{{\mathbf{21}}{\mathbf{.73}}}}$ |
| D. cryptomeria | – | – | – | – | $\frac{{{\mathbf{16}}{\mathbf{.04}}}}{{1.72}}$ | $\frac{{{\mathbf{43}}{\mathbf{.72}}}}{{6.56}}$ | $\frac{{8.16}}{{11.82}}$ | $\frac{{11.61}}{{{\mathbf{20}}{\mathbf{.22}}}}$ | $\frac{{6.59}}{{{\mathbf{38}}{\mathbf{.18}}}}$ | $\frac{{11.07}}{{{\mathbf{56}}{\mathbf{.14}}}}$ | $\frac{{{\text{1}}{\text{.48}}}}{{6.54}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{7.73}}$ | * | – | – | – | – | – | – | – | – | – | – | – | – |
| D. lepnevae | – | $\frac{{{\text{1}}{\text{.91}}}}{{0.11}}$ | – | $\frac{{{\text{1}}{\text{.64}}}}{{0.12}}$ | $\frac{{{\text{0}}{\text{.94}}}}{{0.23}}$ | $\frac{{{\text{0}}{\text{.47}}}}{{0.99}}$ | $\frac{{{\text{0}}{\text{.23}}}}{{2.94}}$ | $\frac{{{\text{0}}{\text{.2}}}}{{2.49}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| D. solida | $\frac{{14.27}}{{10.7}}$ | – | $\frac{{{\text{3}}{\text{.4}}}}{{0.59}}$ | $\frac{{{\text{2}}{\text{.46}}}}{{1.03}}$ | $\frac{{{\text{4}}{\text{.72}}}}{{3.46}}$ | $\frac{{{\text{2}}{\text{.79}}}}{{7.37}}$ | $\frac{{{\text{4}}{\text{.08}}}}{{{\mathbf{51}}{\mathbf{.62}}}}$ | $\frac{{{\text{1}}{\text{.73}}}}{{{\mathbf{38}}{\mathbf{.81}}}}$ | – | – | – | – | – | – | – | – | – | – | – | $\frac{{8.6}}{{1.44}}$ | $\frac{{{\text{1}}{\text{.52}}}}{{0.65}}$ | $\frac{{{\text{2}}{\text{.65}}}}{{1.2}}$ | $\frac{{{\text{2}}{\text{.85}}}}{{1.05}}$ | $\frac{{{\text{1}}{\text{.4}}}}{{1.73}}$ | $\frac{{{\text{4}}{\text{.92}}}}{{1.55}}$ |
| D. triacantha | – | – | – | $\frac{{{\text{2}}{\text{.46}}}}{{0.12}}$ | $\frac{{{\text{0}}{\text{.24}}}}{{0.14}}$ | – | – | – | $\frac{{{\text{0}}{\text{.29}}}}{{9.3}}$ | – | – | – | – | – | * | – | – | – | – | – | $\frac{{{\text{0}}{\text{.17}}}}{{2.13}}$ | – | – | – | – |
| Druella spp. | – | $\frac{{{\text{1}}{\text{.91}}}}{{0.11}}$ | – | $\frac{{{\text{0}}{\text{.82}}}}{{0.04}}$ | – | – | $\frac{{5.22}}{{0.2}}$ | – | – | – | – | $\frac{{{\mathbf{20}}{\mathbf{.82}}}}{{7.58}}$ | $\frac{{6.25}}{{2.22}}$ | – | – | – | – | – | $\frac{{9.3}}{{2.31}}$ | – | $\frac{{10.44}}{{3.15}}$ | $\frac{{{\text{0}}{\text{.11}}}}{{1.79}}$ | $\frac{{{\text{1}}{\text{.2}}}}{{0.02}}$ | $\frac{{{\text{2}}{\text{.21}}}}{{1.03}}$ | – |
| Ephemerella aurivillii | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.11}}}}{{2.26}}$ | – | – | – |
| E. atagosana | $\frac{{{\text{2}}{\text{.53}}}}{{1.3}}$ | – | $\frac{{{\text{4}}{\text{.17}}}}{{0.97}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.18}}$ | $\frac{{{\text{2}}{\text{.12}}}}{{3.62}}$ | $\frac{{{\text{0}}{\text{.47}}}}{{1.17}}$ | * | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{0}}{\text{.36}}}}{{0.17}}$ | $\frac{{{\text{0}}{\text{.58}}}}{{0.48}}$ | – | $\frac{{{\text{0}}{\text{.51}}}}{{0.46}}$ | $\frac{{{\text{0}}{\text{.96}}}}{{2.26}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{1.18}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.48}}$ | $\frac{{{\text{1}}{\text{.17}}}}{{1.07}}$ |
| E. kozhovi | – | – | – | – | $\frac{{{\text{1}}{\text{.41}}}}{{0.32}}$ | $\frac{{{\text{2}}{\text{.33}}}}{{1.98}}$ | $\frac{{{\text{1}}{\text{.59}}}}{{0.3}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Cincticostella levanidovae | $\frac{{{\text{0}}{\text{.72}}}}{{6.63}}$ | – | $\frac{{6.44}}{{{\mathbf{27}}{\mathbf{.15}}}}$ | – | $\frac{{{\text{1}}{\text{.18}}}}{{5.12}}$ | $\frac{{{\text{0}}{\text{.93}}}}{{4.72}}$ | $\frac{{{\text{0}}{\text{.23}}}}{{5.02}}$ | – | – | – | – | – | – | $\frac{{{\mathbf{15}}{\mathbf{.45}}}}{{9.64}}$ | $\frac{{13.97}}{{10.68}}$ | $\frac{{{\text{2}}{\text{.7}}}}{{11.49}}$ | $\frac{{{\text{0}}{\text{.52}}}}{{5.25}}$ | $\frac{{{\text{3}}{\text{.65}}}}{{{\mathbf{24}}{\mathbf{.53}}}}$ | $\frac{{{\text{3}}{\text{.5}}}}{{{\mathbf{19}}{\mathbf{.2}}}}$ | * | – | $\frac{{{\text{0}}{\text{.53}}}}{{5.61}}$ | $\frac{{{\text{0}}{\text{.37}}}}{3}$ | – | – |
| C. tshernovae | $\frac{{{\text{0}}{\text{.36}}}}{{2.19}}$ | – | $\frac{{{\text{0}}{\text{.38}}}}{{2.53}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.86}}$ | – | $\frac{{{\text{0}}{\text{.47}}}}{{4.5}}$ | * | $\frac{{{\text{0}}{\text{.1}}}}{{1.97}}$ | – | – | – | * | – | $\frac{{7.27}}{{{\mathbf{16}}{\mathbf{.58}}}}$ | $\frac{{12.75}}{{{\mathbf{18}}{\mathbf{.18}}}}$ | $\frac{{{\text{0}}{\text{.27}}}}{{0.1}}$ | – | $\frac{{{\text{1}}{\text{.73}}}}{{13.27}}$ | $\frac{{{\text{1}}{\text{.16}}}}{{14.77}}$ | $\frac{{{\text{2}}{\text{.08}}}}{{{\mathbf{26}}{\mathbf{.26}}}}$ | $\frac{{{\text{0}}{\text{.34}}}}{{9.26}}$ | $\frac{{{\text{0}}{\text{.42}}}}{{5.22}}$ | $\frac{{{\text{0}}{\text{.18}}}}{{3.82}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{2.63}}$ | $\frac{{{\text{0}}{\text{.26}}}}{{5.37}}$ |
| Cincticostella spp. | – | – | – | – | – | – | – | – | – | – | $\frac{{{\text{4}}{\text{.07}}}}{{0.24}}$ | $\frac{{{\text{1}}{\text{.72}}}}{{1.43}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Serratella ignita | – | – | – | – | – | $\frac{{{\text{0}}{\text{.47}}}}{{0.54}}$ | $\frac{{{\text{0}}{\text{.23}}}}{{0.74}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| S. setigera | – | – | – | – | * | – | – | $\frac{{{\text{1}}{\text{.43}}}}{{1.27}}$ | – | $\frac{{{\text{1}}{\text{.53}}}}{{0.42}}$ | $\frac{{{\text{0}}{\text{.37}}}}{{0.6}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| S. zapekinae | – | – | – | – | – | – | – | – | $\frac{{12.88}}{{2.54}}$ | $\frac{{{\mathbf{15}}{\mathbf{.27}}}}{{3.01}}$ | $\frac{{{\text{3}}{\text{.15}}}}{{3.32}}$ | $\frac{{{\text{0}}{\text{.64}}}}{{2.86}}$ | * | * | – | – | – | – | – | – | – | – | – | – | – |
| Torleya sp. | – | – | – | – | – | – | – | $\frac{{6.21}}{{1.27}}$ | $\frac{{{\text{1}}{\text{.72}}}}{{2.11}}$ | – | $\frac{{{\text{0}}{\text{.56}}}}{{0.57}}$ | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Teloganopsis punctisetae | $\frac{{{\text{4}}{\text{.51}}}}{1}$ | – | $\frac{{{\text{3}}{\text{.03}}}}{{0.22}}$ | $\frac{{{\text{1}}{\text{.23}}}}{{0.12}}$ | $\frac{{{\text{0}}{\text{.24}}}}{{0.14}}$ | $\frac{{{\text{0}}{\text{.47}}}}{{0.13}}$ | $\frac{{5.9}}{{4.16}}$ | $\frac{{{\text{2}}{\text{.85}}}}{{5.73}}$ | * | – | $\frac{{{\text{0}}{\text{.19}}}}{{0.6}}$ | $\frac{{{\text{0}}{\text{.43}}}}{{1.15}}$ | – | – | $\frac{{{\mathbf{23}}{\mathbf{.9}}}}{7}$ | $\frac{{{\text{1}}{\text{.35}}}}{{2.04}}$ | – | $\frac{{{\text{3}}{\text{.19}}}}{{1.43}}$ | $\frac{{{\text{1}}{\text{.75}}}}{{1.02}}$ | $\frac{{{\text{1}}{\text{.3}}}}{{0.08}}$ | $\frac{{{\text{0}}{\text{.17}}}}{{0.02}}$ | $\frac{{{\text{1}}{\text{.06}}}}{{0.64}}$ | $\frac{{{\text{0}}{\text{.46}}}}{{0.23}}$ | $\frac{{{\text{1}}{\text{.06}}}}{{0.38}}$ | $\frac{{{\text{0}}{\text{.26}}}}{{0.13}}$ |
| Ephemerellidae | $\frac{{{\mathbf{46}}{\mathbf{.21}}}}{{2.6}}$ | $\frac{{{\mathbf{51}}{\mathbf{.48}}}}{{3.45}}$ | $\frac{{{\mathbf{22}}{\mathbf{.34}}}}{{0.82}}$ | $\frac{{{\mathbf{24}}{\mathbf{.99}}}}{{1.89}}$ | $\frac{{{\mathbf{27}}{\mathbf{.11}}}}{{1.13}}$ | $\frac{{5.58}}{{0.22}}$ | $\frac{{8.16}}{{0.43}}$ | $\frac{{14.05}}{{1.5}}$ | $\frac{{{\text{1}}{\text{.72}}}}{{0.35}}$ | $\frac{{{\text{1}}{\text{.53}}}}{{0.01}}$ | $\frac{{{\mathbf{24}}{\mathbf{.81}}}}{{1.4}}$ | $\frac{{{\mathbf{23}}{\mathbf{.82}}}}{{6.45}}$ | – | $\frac{{12.73}}{{5.78}}$ | $\frac{{{\text{0}}{\text{.81}}}}{{0.15}}$ | $\frac{{{\text{1}}{\text{.89}}}}{{1.02}}$ | $\frac{{{\text{4}}{\text{.71}}}}{{2.2}}$ | $\frac{{{\mathbf{20}}{\mathbf{.78}}}}{{2.25}}$ | $\frac{{{\mathbf{23}}{\mathbf{.84}}}}{{7.28}}$ | $\frac{{10.95}}{{0.09}}$ | $\frac{{5.39}}{{0.11}}$ | $\frac{{13.06}}{{1.13}}$ | $\frac{{{\mathbf{21}}{\mathbf{.27}}}}{{1.59}}$ | $\frac{{7.6}}{{0.11}}$ | $\frac{{9.58}}{{1.03}}$ |
| Caenis rivulorum | $\frac{{{\text{0}}{\text{.72}}}}{{0.07}}$ | – | $\frac{{{\text{0}}{\text{.38}}}}{{0.01}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.01}}$ | $\frac{{5.19}}{{0.1}}$ | $\frac{{8.84}}{{0.2}}$ | $\frac{{7.7}}{{0.56}}$ | $\frac{{{\text{4}}{\text{.8}}}}{{1.18}}$ | $\frac{{12.61}}{{11.34}}$ | $\frac{{{\text{0}}{\text{.76}}}}{{0.1}}$ | – | $\frac{{{\text{0}}{\text{.86}}}}{{0.03}}$ | – | – | – | $\frac{{{\text{4}}{\text{.87}}}}{{0.87}}$ | – | $\frac{{{\text{1}}{\text{.73}}}}{{0.08}}$ | $\frac{{9.3}}{{0.27}}$ | $\frac{{{\text{0}}{\text{.52}}}}{{0.02}}$ | $\frac{{{\text{0}}{\text{.5}}}}{{0.02}}$ | $\frac{{{\text{0}}{\text{.53}}}}{{0.02}}$ | $\frac{{{\text{1}}{\text{.47}}}}{{0.02}}$ | $\frac{{{\text{0}}{\text{.41}}}}{{0.01}}$ | – |
По экологической классификации (по: Тиунова, 2005, с изменениями в систематике) в сообществе преобладали гемиритрофилы (Ephemera strigata Eaton, Epeorus (Belovius) pellucidus (Brodsky), Acentrella sibiricum (Kazl.), Neoleptophlebia japonica (Mats.), Drunella cryptomeria (Iman.), D. triacantha Tshern., Ephemerella aurivillii Bengts., E. kozhovi Bajk., Cincticostella levanidovae (Tshern.), Serratella setigera (Bajk.), Caenis rivulorum Eaton) и эвриритробионты (Cinygmula kurenzovi (Bajk.), C. sapporensis (Mats.), Ecdyonurus (Afghanurus) aspersus Kluge, E. (A.) bajkovae Kluge, Baetis (Baetis) pseudothermicus Kluge, Baetiella tuberculatum (Kazl.), Neoleptophlebia vladivostokica (Kluge), Drunella solida Bajk., Cincticostella tshernovae (Bajk.)). Также присутствовали психроритрофилы (Cinygmula hirasana Iman., Drunella aculea Allen, D. lepnevae Tshern., Ephemerella (Hosoba) atagosana Iman., Serratella zapekinae Bajk.), гемипотамофилы (Ecdyonurus (Atopopus) abracadabrus Kluge, Baetis (Baetis) fuscatus L., Serratella ignita (Poda), Teloganopsis punctisetae (Mats.)) и мезопотамобионты (Baetis (Nigrobaetis) bacillus Kluge).
В видовом отношении наиболее широко представлены семейства Heptageniidae (девять видов) и Ephemerellidae (15 видов), преимущественно, составившие категорию доминант. Так, представители рода Cinygmula (Heptageniidae), относящиеся к группе плоских личинок, приспособленных к жизни между камнями на сильном течении, присутствовали в водотоке с октября по май включительно. В осенние и зимние месяцы наиболее многочисленной была группа Cinygmula spp., представленная неидентифицируемыми до вида нимфами и личинками ранних возрастов – доминант по численности и субдоминант по биомассе. В марте–апреле значимую долю в численность сообщества вносили личинки C. hirasana и C. sapporensis, в первой половине мая – C. kurenzovi. Высокие показатели биомассы, отмеченные для этих видов в весенний период, закономерно связаны с ростовыми изменениями, а динамика количественных характеристик внутри рода – с перераспределением личинок по микробиотопам в зависимости от видовой принадлежности и особенностей жизненного цикла (Тиунова, 2008).
Нимфы из группы Ephemerellidae формировали категории доминант по численности с марта по первую половину мая, в августе, ноябре и конце января. Однако их присутствие в сообществе не оказало значительного влияния на показатели биомассы. Так, в пробе от 18 августа отмечено численное доминирование неидентифицируемых до вида нимф рода Drunella, 10 и 25 мая – молоди D. cryptomeria с очень низкой массой. В то же время средне- и поздневозрастные представители этого рода, обитатели верхней стороны камней (Чернова, 1952), формировали основную биомассу сообщества поденок в следующие временные периоды: с конца июня по конец июля – D. cryptomeria, в июне – D. solida, в сентябре–мае (с несколькими исключениями) – D. aculea, что связано с фенологическими особенностями созревания этих крупных видов (Тиунова, 2014).
Среди поденок из сем. Ephemerellidae значительную массу тела также имеют личинки рода Cincticostella, представленные в р. Тигровая двумя видами. Отмеченный для C. levanidovae доминантный показатель (12 сентября) и переход через категорию субдоминант (26 сентября) во второстепенные виды (с 10 октября), отражают процессы массового отрождения нимф, роста и последующего перемещения в зоны скопления растительных остатков (Тиунова, 2014). Эти же процессы, вкупе с активным ростом, объясняют положение вида в категории субдоминант по биомассе в сентябре–октябре, доминант – в ноябре. Для C. levanidovae также следует отметить пробу от 11 апреля, содержавшую немногочисленных крупных личинок поздних возрастов.
Ситуация, при которой личинки, приближающиеся к стадии зрелости, обеспечивают доминирование группы по биомассе, не закономерна. Так, в пробе от 12 сентября при показателе биомассы в 16.58% личинки C. tshernovae, также предпочитающие места скопления листового опада, были ранневозрастными и имели среднюю массу тела всего 1.34 мг, 7 ноября, занимая категорию субдоминант, уже 13.47 мг, а 25 мая, при массе в 50 мг, формировали категорию второстепенных видов.
В периоды резкого подъема воды, увеличения скоростей течения и невозможностью взятия проб на грунтах постоянного русла проанализированы сообщества, формирующиеся на ранее осушенных, незаселенных участках поймы. В период весеннего половодья по численности доминировали нимфы из группы Ephemerellidae и средневозрастные Cinygmula hirasana, присутствовали субдоминанты С. sapporensis и Neoleptophlebia japonica. Эти три вида обеспечили и основную долю биомассы сообщества поденок.
К началу летних паводков количество таксономических групп сообщества увеличилось, и с начала июля освоение новых грунтов происходило значительно бóльшим числом таксонов, достигших показателей до уровней категории доминант и субдоминант. В пробах 7 июля в количественном отношении преобладали личинки ранних возрастов из групп Heptageniidae и Epeorus spp., 22 июля – Rhithrogena sp. и Serratella zapekinae. Доминантами по массе в июле были средневозрастные личинки Drunella cryptomeria.
Проба от 30 августа, собранная непосредственно после ливневых осадков, содержала случайный набор видов, принесенных током воды.
В осенний паводок отмечено резкое сокращение числа таксономических групп в категории доминант и субдоминант. Так, в пробах 24 октября 84.3% численности и 60.86% биомассы сообщества поденок приходилось на ранневозрастных личинок Cinygmula spp., еще 17.51% биомассы привнесли C. hirasana и всего 5.25% – Cincticostella levanidovae.
В период весенней межени, наблюдавшейся на р. Тигровая в мае–июне, только личинки Cinygmula kurenzovi, имевшие среднюю массу ~7.5 мг, доминировали и по численности, и по биомассе. В остальных случаях, доминантами по численности были нимфы и личинки ранних возрастов, имеющие незначительную массу: в мае – это Drunella cryptomeria и Ephemerellidae, 20 июня – Epeorus spp. и Heptageniidae. Cубдоминант 10 мая представлял лишь один вид – Caenis rivulorum, 25 мая – уже четыре группы, но с низким долевым вкладом: Rhithrogena sp., Heptageniidae, Aсentrella gr. sibirica, Ephemerellidae. Особенностью пробы 8 июня было полное отсутствие доминант по численности, а категорию субдоминант с показателями 5.22–12.7%, представляли 10 из 23 таксономических групп сообщества, зарегистрированных в день отбора. Среди них следует отметить быстринных, активно плавающих личинок Baetis fuscatus, Baetiella tuberculatum, группу Baetidae, обитателей быстротекущих вод Teloganopsis punctisetae и приспособленных к жизни в заиленных стациях Caenis rivulorum. К 20 июня число таксономических групп в категории субдоминант снизилось до двух: Ephemerellidae и Drunella cryptomeria.
Основной вклад в биомассу сообщества в мае вносили D. aculea, в июне D. cryptomeria и D. solida. Категория субдоминант отсутствовала в пробе 10 мая, а 25 мая включала только D. solida и D. cryptomeria. К июню видовой состав категории значительно изменился: к D. cryptomeria добавились Ecdyonurus bajkovae и Rhithrogena sp., а 20 июня сохранились личинки Rhithrogena sp. и появились Teloganopsis punctisetae.
В период осенней межени (10 октября) основу сообщества поденок р. Тигровая слагали мелкие личинки из группы Heptageniidae (67.57% по численности и 29.86% по биомассе).
Важно отметить, что подобная картина монодоминирования (>60%) отмечена только в осенний и зимний периоды и исключительно у сем. Heptageniidae. В частности, группа видов Cinygmula spp. на 82.42% сформировала зимний (7 февраля), он же годовой максимум численности поденок сообщества. В целом, пиковые значения на протяжении года формировали одна–две доминанты из семейств Heptageniidae и (или) Ephemerellidae: осенью (10 ноября) – личинки групп Cinygmula spp. и Ephemerellidae, весной (8 марта) – Ephemerellidae и C. hirasana. Однако календарно-летний максимум, пришедшийся на последний день затяжной весенней межени (20 июня), сформировался при участии таксонов из категории доминант (Epeorus spp., Heptageniidae) и субдоминант (Drunella cryptomeria, Ephemerellidae). Аналогично этому, годовой максимум биомассы сообщества поденок р. Тигровая, отмеченный 10 мая, был достигнут благодаря вкладу личинок Cinygmula kurenzovi и Drunella aculea, пик 8 июня – D. solida и D. cryptomeria, 4 августа – Epeorus pellucidus, Ecdyonurus aspersus и Epeorus spp., 7 ноября – Drunella aculea, Cincticostella levanidovae, C. tshernovae и Cinygmula spp.
Следует отметить, что поденки рода Ephemera – представители эколого-морфологической группы грунтовых закапывающихся личинок – лишь дважды занимали категорию доминант по биомассе. Из наблюдений за процессом вылета субимаго 8 июня и по наличию крупных скоплений линных шкурок на затишных участках мелководья, можно предположить массовость этих поденок, а небольшое количество в пробах объяснить приуроченностью к определенным типам биотопов в течение жизненного цикла и перемещением к урезу реки во время перехода в крылатую стадию.
Способностью быстро перемещаться в условиях потока, и следовательно избегать ловчей конструкции, обладают Isonychia sp., Ameletus sp., Baetis (Nigrobaetis) sp., B. (N.) bacillus, занимающие в данном водотоке категории второстепенных и третьестепенных. Однако для них, а также для представителей рода Neoleptophlebia, ряда видов сем. Ephemerellidae и Caenis rivulorum можно предполагать и истинную малочисленность в р. Тигровая, и несоответствие видовой специфики и гидрологических условий на точках отбора. Следует отметить наличие расхождений в таксономическом и, зачастую, возрастном составе количественных и качественных проб, основная причина которых видится в площади бентометра, определившего гранулометрический состав исследованных грунтов. Соответственно, в исследование не попали сообщества валунов, размеры которых превышали 22 см в диаметре. Это отразилось и на составе, и на количественных характеристиках всего бентосного сообщества. Так, например, не были учтены личинки ручейника Stenopsyche marmorata, Navas, 1920, прикрепляющие свои сети к крупным валунам, и зрелые личинки ряда видов поденок.
Выводы. Сообщество поденок р. Тигровая – динамичное образование, объединяющее ~40 видовых таксонов и изменяющееся под воздействием факторов среды и индивидуальных фенологических процессов. В соответствии с ранее выявленными закономерностями (Тиунова, 2007), в бентосном сообществе исследованного участка метаритрали этого умеренно-холодноводного водотока протяженностью >50 км, на грунтах с фракциями <22 см в диаметре, поденки занимали категорию доминант с показателями численности 176–8601 экз./м2, биомассы – 0.38–19.67 г/м2. Минимальные значения отмечены во время летнего паводка, максимум по численности – в ледовый период, по биомассе – в весенне-летнюю межень.
По экологической классификации, гидрологические условия места отбора проб были привлекательны для (по убывающей): гемиритрофилов, эвриритробионтов, психроритрофилов, гемипотамофилов и мезопотамобионтов.
Ведущую роль в формировании сообщества поденок на протяжении года играли представители семейств Heptageniidae и Ephemerellidae, преимущественно, родов Cinygmula и Drunella. В ряде случаев высокие показатели количественных характеристик достигнуты за счет нимф и личинок ранних возрастов, например, осеннее-зимнее монодоминирование по численности (84.3%) группы видов Cinygmula spp. Но, если процессы массового отрождения приводили к росту численности, то ситуация, при которой ростовые процессы обеспечивают доминирование группы по биомассе, не была закономерной. Это объясняется видовой спецификой поденок, выражающейся, в приуроченности личинок разных размерно-возрастных групп к определенным местообитаниям в зависимости от особенностей жизненного цикла. Процессы перераспределения личинок между микробиотопами отразились на долевом вкладе таксонов, что, в частности, отмечено для представителей родов Cincticostella и Cinygmula.
При заселении новых субстратов в весенний период массовыми и пионерными, в первую очередь, были нимфы и личинки ранних возрастов с очень низкой массой тела (преимущественно сем. Ephemerellidae). В период летних паводков наблюдали аналогичную ситуацию, однако преобладающую численность обеспечивали представители сем. Heptageniidae. При этом, между таксонами сообщества поденок имело место более равномерное распределение количественных показателей, что привело к увеличению числа групп, относящихся к категориям доминант и субдоминант. В осенний паводок оно резко снизилось.
Качественный и количественный состав сообщества поденок на протяжении всего периода исследований определялся их фенологическими особенностями и изменялся с течением времени в соответствии с гидрологическими процессами и особенностями онтогенеза.
Список литературы
Леванидов В.Я. 1969. Воспроизводство амурских лососей и кормовая база их молоди в притоках Амура // Изв. ТИНРО. Т. 67.
Леванидов В.Я. 1977. Биомасса и структура донных биоценозов реки Кедровой // Пресноводная фауна заповедника “Кедровая падь”. Владивосток: Дальневосточ. науч. центр АН СССР. Т. 45 (148). С. 126.
Леванидова И.М., Лукьянченко Т.И., Тесленко В.А. и др. 1989. Экологические исследования лососевых рек Дальнего Востока СССР // Систематика и экология речных организмов. Владивосток: ДВО АН СССР. С. 74.
Тиунова Т.М. 2005. Экологическая классификация реофильных личинок поденок (Ephemeroptera) юга Российского Дальнего Востока // Чтения памяти В.Я. Леванидова. Вып. 3. С. 113.
Тиунова Т.М. 2007. Динамика биомассы бентоса в экосистемах лососевых рек юга Дальнего Востока // Биологические ресурсы Дальнего Востока России: комплексный региональный проект ДВО РАН. Москва: Тов-во науч. изданий КМК. С. 196.
Тиунова Т.М. 2008. Состав и структура сообществ зообентоса микробиотопов малой предгорной реки умеренно холодноводного типа // Чтения памяти В.Я. Леванидова. Вып. 4. С. 31.
Тиунова Т.М. 2014. Жизненные циклы дальневосточных видов поденок (Insecta, Ephemeroptera) // Чтения памяти В.Я. Леванидова. Вып. 6. С. 682.
Ткачев Б.П., Булатов В.И. 2002. Малые реки: современное состояние и экологические проблемы: аналитический обзор. Новосибирск: Гос. публ. науч.-тех. библиотека СО РАН.
Чернова О.А. 1952. Поденки (Ephemeroptera) бассейна р. Амура и прилежащих вод, и их роль в питании амурских рыб // Материал к познанию флоры и фауны СССР. М.: Мос. общ-во испытателей природы. Т. 32(47). С. 229.
Чертопруд М.В. 2021. Основные классы реофильных сообществ макробеспозвоночных и их региональная изменчивость в Евразии // Биология внутр. вод. № 5. С. 481.https://doi.org/10.31857/S032096522105003X
Дополнительные материалы отсутствуют.
Инструменты
Биология внутренних вод