

Биология внутренних вод, 2022, № 6, стр. 689-699
Природно-климатические закономерности распространения Sparganium emersum и S. hyperboreum (Typhaceae) на северо-востоке Азии
О. А. Мочалова a, *, Д. Ю. Ефимов b, **
a Институт биологических проблем Севера Дальневосточного отделения Российской академии наук
Магадан, Россия
b Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук
пос. Борок, Некоузский р-н, Ярославская обл., Россия
* E-mail: mochalova@inbox.ru
** E-mail: dnsfmv@ibiw.ru
Поступила в редакцию 26.04.2022
После доработки 11.06.2022
Принята к публикации 16.06.2022
- EDN: UPELIG
- DOI: 10.31857/S032096522206016X
Аннотация
В пределах крупного биогеографического региона – северо-востока Азии – определены географические и биоклиматические факторы распространения широко представленных в регионе ежеголовников – Sparganium hyperboreum и S. emersum. Наиболее широко распространен S. hyperboreum, представлен на всeм пространстве региона, проникает высоко в горы и далеко в Заполярье, занимая наиболее широкий спектр местообитаний. Морфологически вариабельный S. emersum встречается реже, представлен двумя формами: широколистным S. emersum s. str. и узколистным S. emersum “rothertii”. S. emersum s. str. реже всего встречается, он произрастает преимущественно в западной части региона, в основном в Якутии, а S. emersum “rothertii” – в восточной, собственно на Дальнем Востоке. Оба тяготеют к низинам, равнинам, речным долинам. В градиентах биоклиматических переменных S. emersum s. str. и S. hyperboreum наиболее удалены. Промежуточное положение в регионе занимает S. emersum “rothertii”, смещаясь по многим климатическим показателям к S. hyperboreum. Вероятность находок S. hyperboreum возрастает в северо-восточном направлении и с увеличением высоты местности, S. emersum “rothertii”– в восточном, S. emersum s. str. – в западном. S. emersum s. str. наиболее требователен к теплообеспеченности и сухости климата. Распространение ежеголовников в регионе наиболее определяют средний дневной диапазон температуры, температурный годовой диапазон, средняя температура самого засушливого квартала, годовое количество осадков, сезонность осадков, осадки самого влажного квартала и осадки самого засушливого квартала. Вся территория северо-востока Азии по геопространственным и биоклиматическим параметрам пригодна для произрастания Sparganium hyperboreum и S. emersum, их отсутствие в каких-либо частях связано с комбинацией природно-климатических условий или с историческими причинами.
ВВЕДЕНИЕ
Определение закономерностей распространения водных растений – важная экологическая задача и проблема при оценке эффектов климатических изменений и их последствий (Bornette, Puijalon, 2011; Hossain et al., 2017). Особо актуально исследование воздействия изменения климата на редкие, уязвимые виды водных макрофитов, поскольку мелководные и прибрежно-водные экотоны особо уязвимы (Hossain at al., 2017). Данные по влиянию изменений различных природно-климатических факторов на распространение водных сосудистых растений, играющих исключительную роль в разнообразии биоты водоемов, для северных регионов немногочисленны (Lacoul, Fridman, 2006; Alahuhta et al., 2011; Wu et al., 2016; Бобров, Мочалова, 2017; Андриянова и др., 2019). Якутия – единственный северный регион, где японскими и российскими ботаниками проведено количественное сравнение распределения четырех видов Sparganium (S. angustifolium Michx., S. emersum Rehm., S. hyperboreum Laest., S. natans L.) в зависимости от некоторых климатических факторов и широтно-долготного градиента (Takahashi et al., 2001). Авторами показано, что S. hyperboreum и S. emersum демонстрируют явную разницу в особенностях распространения в Якутии и предпочитают местообитания, характеризующиеся различными климатическими и экологическими факторами. Решено было провести анализ связей пространственного распределения видов c климатическими факторами для самых распространенных ежеголовников на большей по площади территории, используя количественные методы анализа (Gillard et al., 2017; Viana, 2017; Banerjee et al., 2020).
Наиболее распространенные виды Sparganium, встречающиеся на северо-востоке Азии (СВА) – S. emersum и S. hyperboreum. Эти циркумголарктические виды произрастают от Заполярья до горных районов на сороковых широтах в арктической, субарктической и бореальной зонах (Юзепчук, 1934; Толмачев, 1960; Cook, Nicholls, 1986; Тимохина, 1988; Цвелев, 1996, Определитель…, 2020). Sparganium emersum характеризуется значительным полиморфизмом, для него приводятся разнообразные формы (Юзепчук, 1934; Cook, Nicholls, 1986; Цвелев, 1996).
Цель работы – проанализировать закономерности распространения Sparganium emersum и S. hyperboreum на СВА в градиенте географических и биоклиматических факторов; выявить, какое влияние оказывают географические и биоклиматические факторы на распространение видов; спрогнозировать возможность смещения границ ареалов в пределах СВА в результате природно-климатических изменений.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Район исследования – СВА, крупный биографический регион, охватывает территорию Якутии (Республика Саха) (3084 тыс. км2), Магаданской обл. (462 тыс. км2), Чукотского Автономного округа (721 тыс. км2) и Камчатского края (464 тыс. км2). Эта обширная территория имеет сложное геологическое строение и разнообразный рельеф: от горных массивов высотой 2000–3000 м, до обширных низин с многочисленными реками и озерами. Территория расположена в четырех климатических зонах: арктической, субарктической, субарктической морской и бореальной. Бо́льшая ее часть находится под влиянием континентального и ультраконтинентального климата, только прибрежные районы – океанического (Алисов, 1956). Значительная часть территории расположена в зоне многолетней мерзлоты. Природно-климатические особенности СВА описаны в (Пармузин, 1964; Клюкин, 1970; Chemeris et al., 2020).
Источник данных о распространении видов Sparganium на СВА – оригинальные сборы, критически пересмотренные коллекции гербариев (MAG, MHA, MW, IBIW, LE, NSK, SASY и др.), цифровые гербарии (https://plant.depo.msu.ru; http://herb.csbg.nsc.ru:8081; http://botsad.ru/herbarium), глобальная информационная система о биоразнообразии GBIF (www.gbif.org) и сведения из литературы (Труфанова, 1972; Вехов, 1993; Takahashi et al., 1994, 2001; Бобров, Мочалова, 2014 и др.). Всего проанализировано 2134 записи о находках всех таксонов Sparganium с территории СВА. Для идентификации образцов использованы морфологические признаки (строение и размер соцветий, форма и размер плодов, жилкование и форма листьев); образцы без генеративных органов включены в анализ только с достаточно выраженными признаками формы и жилкования листьев.
Наиболее представленные виды на СВА – это Sparganium emersum и S. hyperboreum. Вслед за Цвелевым (1984, 1996) и Бобровым с соавт. (2021) сильно морфологически вариабельный в регионе S. emersum разделен в работе на две формы, таксономический статус которых пока неясен: “классический” широколистный с трехгранными листьями S. emersum s. str. и узколистный “нетипичный” S. emersum s. l. уклоняющийся к S. rothertii Tzvel. (далее S. emersum “rothertii”).
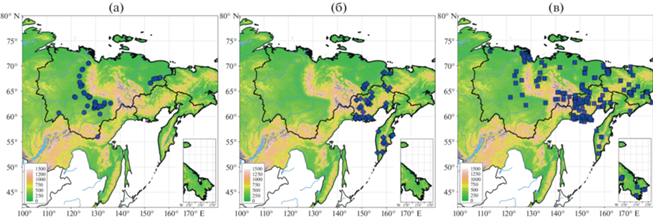
Sparganium emersum s. str. имеет широкие >5 мм листья, в сечении трехгранные с хорошо выраженной центральной жилкой; нижние прицветные листья чуть короче или немного превышают соцветия; пестичных головок 3–5, расставленные, тычиночных головок 4–7, расставленные, отчетливо отделены промежутком от пестичных головок; рыльца 1.8–2.2 мм, пыльники 1.3–1.5 мм; плоды веретеновидные с длинным столбиком. У S. emersum “rothertii” листья длинные, в сечении слегка уплощенные или нечетко трехгранные, с выраженной центральной жилкой, узкие (<5 мм); нижние прицветные листья в 1.5–2 раза превышают соцветие, обычно толще стеблевых листьев; пестичных головок 2–4, расставленные, тычиночных головок 3–5, сближенные, слабо отделены промежутком от пестичных головок; рыльца 1.3–1.5 мм, пыльники 0.8–1.1 мм, плоды широковеретеновидные с длинным столбиком. У S. hyperboreum – листья плоские, без выраженной центральной жилки, узкие (<5 мм); нижние прицветные листья заметно длиннее соцветия; пестичных головок 1–3, более-менее скученные, тычиночных головок 1–2, скученные, сближены с пестичными головками; рыльца 0.3–0.5 мм, пыльники 0.3–0.6 мм; плоды широковеретеновидные с едва заметным столбиком. Для S. emersum (обе формы) и S. hyperboreum после выбраковки сомнительных указаний, плохо сохранившихся экземпляров и дублетов массив данных, включeнных в анализ, насчитывал 397 записей: 158 местонахождений S. emersum (55 – S. emersum s. str., 103 – S. emersum “rothertii”) и 239 местонахождений S. hyperboreum (рис. 1). В базу данных по местонахождениям Sparganium включали только записи, подтвержденные гербарными образцами (Доп. мат. тaбл. S1).
Рис. 1.
Распространение находок ежеголовников Sparganium по элементам рельефа на СВА: a – S. emersum s. str., б – S. emersum “rothertii”, в – S. hyperboreum.
Географические переменные основываются на геопространственных данных (широта, долгота, высота над уровнем моря) цифровой топографической модели (SRTM, www.usgs.gov). Биоклиматические переменные (19 стандартных переменных температуры и осадков) были загружены из базы данных WorldClim (www.worldclim.org; Fick and Hijmans, 2017), с пространственным разрешением 30 угловых секунд.
Дисперсионный анализ (ANOVA) и множественное сравнение средних (TukeyHSD), при уровне значимости 0.05 (McDonald, 2009), применяли для определения различий между видами. Полиноминальную логистическую регрессию (MLR) использовали для определения соотношения шансов (вероятностей) находок видов в градиентах факторов среды (Engel, 2008). В регрессионных моделях данные по S. emersum “rothertii” использовались в качестве “перехвата”. Набор переменных не редуцировали, чтобы оценить связи каждого отдельного фактора с распространением видов ежеголовников на СВА (Доп. мат. txt. S1). Все статистические расчеты и подготовку визуализации проводили в среде R 4.1.2 (R Core Team, 2021)11.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Географические и экологические факторы. Sparganium emersum – плюризональный голарктический вид, на СВА встречающийся в очень разнообразных экотопах, тяготея к долинам крупных рек, обширным межгорным впадинам и равнинам вдоль морского побережья, редок в горных районах. S. hyperboreum – арктобореальный голарктический вид, достаточно равномерно распределенный по всей территории, встречается и в горных, и на равнинных территориях (рис. 1).
S. emersum s. str. (n = 55) в долготном градиенте распространен на СВА прерывисто, в широком диапазоне 115°–157° в.д. Подавляющая часть находок (n = 46; 84%) сосредоточена в “западной” части СВА – в интервале 115°–137° в.д., и лишь небольшая часть (n = 9; 16%) в “восточной” – 152°–157° в.д. (рис. 2). Самые восточные (157° в.д.) местонахождения S. emersum s. str. расположены на северо-востоке Якутии (Среднеколымский р-н, 67.73° с.ш., 156.46° в.д., протока Кружовская на правом берегу р. Колыма, старичное озеро, 24.09.1966, Г.Н. Егорова, опр. Е.Р. Труфанова (SASY)). Находки S. emersum s. str. в широтном градиенте на СВА распределены непрерывно между 56°–76° с.ш., с максимумом в интервале – 60.5°–64.8° с.ш. (n = 35; 64%). Самые северные находки зарегистрированы в Якутии в низовьях р. Лена (Булунский улус, 70.55° с.ш., 127.12° в.д., правобережье р. Лена в 18 км выше пос. Кюсюр, оз. Туччахтах, северный берег, 18.07.2020, Н.П. Тихомиров, Н.А. Конотоп (IBIW)). Подавляющая часть находок сделана на высотах ≤200 м над у.м. (n = 49; 89%), число находок >200 м резко снижается с высотой.
Рис. 2.
Плотности распределения находок ежеголовников Sparganium вдоль географических градиентов на СВА. Широтный и высотный (а, в) градиенты охватывают оба полушария, долготный (б) – только восточное. Высотный градиент представлен в логарифмической шкале, log(m). Таксоны ежеголовников: Se – S. emersum s. str., Sr – S. emersum “rothertii” и Sh – S. hyperboreum. Перекрытие ареалов по широте: Se & Sr* – 43%, Se & Sh – 65%, Sr & Sh* – 56%; по долготе: для Se & Sr* – 13%, Se & Sh* – 23%, Sr & Sh* – 54%; по высоте над у.м.: Se & Sr – 64%, Se & Sh* – 42%, Sr & Sh* – 63% (* значимые различия по результатам ANOVA, TukeyHSD – padj < 0.05).
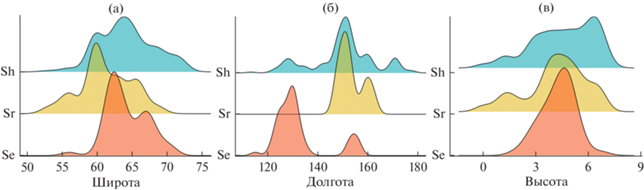
S. emersum “rothertii” (n = 103) в долготном градиенте на СВА сконцентрирован в диапазоне 147°–163° в.д., с максимумом в интервале 147°–154° в.д. (n = 73; 72%) (рис. 2). На востоке известен с Камчатки (Усть-Камчатский р-н, 56.28° с.ш., 162.58° в.д., пос. Усть-Камчатск, 1.5 км к северу от м. Погодный, побережье оз. Нерпичье, старичное слабо подсоленное озерко у дороги, 14.08.2013, А.А. Бобров, О.А. Мочалова, Е.В. Чемерис (MAG)). Западная граница находится в Магаданской обл. (Ольский р-н, 59.66° с.ш., 147.22° в.д., левый берег р. Кава, термокарстовое озеро между оз. Лебяжье и р. Кава, 2.07.2017, О.А. Мочалова (MAG0003360)). Находки S. emersum “rothertii” в широтном градиенте распределены непрерывно между 52°–69° с.ш., с максимумом в узком интервале – 59.5°–60.0° с.ш. (n = 36; 35%). Самые северные местонахождения расположены в низовье р. Колыма (Якутия, Нижнеколымский р-н, 68.51° с.ш., 161.49° в.д., 26 км к ю.-ю.-в. от пос. Черский, правый берег р. Колыма, озеро в восточной части протоки Осетровка, 24.08.2015, А.А. Бобров, О.А. Мочалова (MAG0003330)). Подавляющая часть находок отмечена на высотах ≤200 м над у.м. (n = 49; 89%), с увеличением высоты число находок поступательно снижается. В Магаданской обл. растения собраны на 810 м над у.м. (Сусуманский р-н, 63.31° с.ш., 147.78° в.д., междуречье рек Буркандья и Малык-Сиен, оз. Харюзовое, 24.08.2012, А.А. Бобров, О.А. Мочалова (MAG0003153)). Находки этого таксона известны с СВА, из Магаданской обл., с Чукотки и Камчатки, вне СВА указан для Забайкалья, северо-восточного Китая и Японии (Цвелев, 1984, 1996).
Находки S. hyperboreum (n = 239) в долготном градиенте охватывают всю территорию СВА в диапазоне 110° в.д.–170° з.д. (рис. 1). Основной массив находок сосредоточен на 140°–165° в.д. (n = 156; 65%), с максимумом на 145°–155° в.д. (n = 111; 46%). Самые восточные среди всех ежеголовников находки сделаны в России на ее восточных границах, на 170° з.д. (Чукотка, Чукотский р-н, 66.14° с.ш., 169.97° з.д., окр. лагуны Уэлен, термокарстовое озеро, 07.08.2019, О.А. Мочалова, А.А. Бобров, Е.В. Чемерис (IBIW, MAG)), вид обычен и на п-ове Аляска (https://www.gbif.org/ species/2702302). Находки S. hyperboreum в широтном градиенте на СВА распределены сравнительно более равномерно (рис. 2), с максимумом на 62.5°–65.0° с.ш. (n = 79; 33%). В низовьях р. Колымa вид доходит до 69° с.ш., на р. Лена собран на 72° с.ш., (Якутия, Булунский улус, 72.47° с.ш., 126.21° в.д., 2 км на запад от г. Америка-Хая, озеро в сыром понижении, 30.07.2020, Н.П. Тихомиров (IBIW)). В высотном градиенте основной массив находок (n = 158; 73%) сосредоточен ≤400 м. Максимальная высота находок S. hyperboreum – 1371 м над у.м. отмечена в Магаданской обл. (Тенькинский р-н, 62.14° с.ш., 148.85° в.д., верховья р. Эльгенья, горное озеро, 28.07.2009, Н.В. Синельникова (MAG0003425)).
Ареалы этих ежеголовников в пределах СВА в значительной мере перекрываются между собой (рис. 2). В долготном направлении местонахождения S. emersum s. str. смещены к западу, S. emersum “rothertii”– к востоку. S. hyperboreum более равномерно представлен на всей территории СВА. В градиенте широты эти Sparganium распространены более однородно с максимумом в центральном секторе. В высотном градиенте основная часть находок была на высотах до 200–400 м, при этом, высоко в горы проникает только S. hyperboreum. Соответственно, наиболее значимые различия (ANOVA, df(2, 372), p < 0.05) установлены для долготы (F = 76.8), менее значимые – для широты (F = 20.4) и высоты (F = 9.3). Значимые различия в коэффициентах регрессии (MLR, p < 0.05) получены для S. emersum “rothertii” в паре с S. emersum s. str. – по долготе и с S. hyperboreum –по широте и высоте.
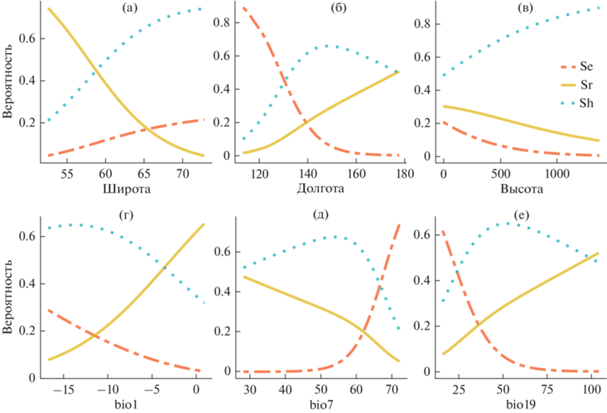
Данные регрессионных моделей по градиенту долготы показывают, что на СВА в направлении с запада на восток вероятность находок S. emersum “rothertii” и S. hyperboreum увеличивается, а S. emersum s. str., наоборот, снижается. Например, на западной границе СВА (113°–114° в.д.) вероятность находок в ряду S. emersum s. str., S. emersum “rothertii” и S. hyperboreum – 89, 1 и 10% соответственно, в восточной части СВА (177°–178° в.д.) вероятности находок этих таксонов – 0, 50 и 50%. В широтном градиенте с юга на север значимо увеличиваются шансы обнаружить только S. hyperboreum, для S. emersum “rothertii”– стремительно уменьшаются (рис. 3а, 3б).
Рис. 3.
Изменчивость оценок вероятности (шансов) находок ежеголовников в градиентах географических (а–в) и биоклиматических (г–е) факторов. Обозначения переменных: Se – S. emersum s. str., Sr – S. emersum “rothertii”, Sh – S. hyperboreum. Биоклиматические параметры: bio1 – среднегодовая температура; bio7 – температурный годовой диапазон; bio19 – осадки самого холодного квартала.
Биоклиматические факторы. Различия между парами таксонов Sparganium по результатам ANOVA (df(2, 372), p < 0.05) получены для всех биоклиматических параметров температуры и количества осадков (табл. 1). Наиболее высокие различия между ежеголовниками по температуре зарегистрированы для среднегодовой температуры (bio 1) (F = 22.6), максимальной температуры самого теплого месяца (bio5) (F = 53.4) и средней температуры самого теплого квартала (bio10) (F = 72.6). По числу осадков наибольшие различия были для годового количества осадков (bio12) (F = 16.2), числа осадков самого засушливого квартала (bio17) (F = 28.4) и осадков самого холодного квартала (bio19) (F = 37.2).
Таблица 1.
Основные статистические характеристики биоклиматических переменных по изученным видам ежеголовника на северо-востоке Азии
| Биоклиматические переменные | S. emersum s. str. | S. emersum “rothertii” | S. hyperboreum |
|---|---|---|---|
| bio1 | $\frac{{ - 10.7 \pm 0.2}}{{ - 13.6\,\, \ldots - 5.4}}$ | $\frac{{ - 7.0 \pm 0.4}}{{ - 13 - 0.9}}$ | $\frac{{ - 9.6 \pm 0.3}}{{ - 16\,\, \ldots 0.0}}$ |
| bio2 | $\frac{{10.8 \pm 0.2}}{{8.5 - 15.0}}$ | $\frac{{10.0 \pm 0.2}}{{5.6 - 13.2}}$ | $\frac{{9.6 \pm 0.2}}{{5.4 - 13.7}}$ |
| bio3 | $\frac{{16.4 \pm 0.2}}{{13.5 - 23.2}}$ | $\frac{{19.9 \pm 0.3}}{{15.1 - 28.2}}$ | $\frac{{18.3 \pm 0.4}}{{13.3 - 26.8}}$ |
| bio4 | $\frac{{2110.8 \pm 19.1}}{{1737.9 - 2337.9}}$ | $\frac{{1539.3 \pm 39.6}}{{836.0 - 2067.3}}$ | $\frac{{1676.4 \pm 26.5}}{{854.0 - 2378.2}}$ |
| bio5 | $\frac{{22.9 \pm 0.3}}{{17.8 - 26.1}}$ | $\frac{{18.8 \pm 0.3}}{{14.6 - 23.3}}$ | $\frac{{18.4 \pm 0.2}}{{10.5 - 24.5}}$ |
| bio6 | $\frac{{ - 42.4 \pm 0.3}}{{ - 46.9\,\, \ldots - {\kern 1pt} 33.4}}$ | $\frac{{ - 30.1 \pm 0.9}}{{ - 42.7\,\, \ldots - {\kern 1pt} 13.5}}$ | $\frac{{ - 34.1 \pm 0.6}}{{ - 49.7\,\, \ldots - 14.3}}$ |
| bio7 | $\frac{{65.3 \pm 0.6}}{{57.6 - 71.9}}$ | $\frac{{48.9 \pm 1.4}}{{28.9 - 64.1}}$ | $\frac{{52.6 \pm 0.8}}{{28.3 - 72.1}}$ |
| bio8 | $\frac{{13.8 \pm 0.4}}{{8.0 - 16.6}}$ | $\frac{{7.7 \pm 0.5}}{{ - 9.7 - 12.1}}$ | $\frac{{9.2 \pm 0.2}}{{0.0 - 15.6}}$ |
| bio9 | $\frac{{ - 26.7 \pm 1.0}}{{ - 34.6\,\, \ldots - 12.9}}$ | $\frac{{ - 14.2 \pm 0.7}}{{ - 25.7 - 2.7}}$ | $\frac{{ - 18.9 \pm 0.5}}{{ - 40.1 - 0.0}}$ |
| bio10 | $\frac{{14.4 \pm 0.3}}{{10.2 - 16.6}}$ | $\frac{{11.9 \pm 0.1}}{{9.3 - 14.5}}$ | $\frac{{11.3 \pm 0.2}}{{6.5 - 15.9}}$ |
| bio11 | $\frac{{ - 36.8 \pm 0.3}}{{ - 40.8\,\, \ldots - 27.2}}$ | $\frac{{ - 25.1 \pm 0.9}}{{ - 36.7\,\, \ldots - 9.6}}$ | $\frac{{ - 29.0 \pm 0.6}}{{ - 44.4\,\, \ldots 0.0}}$ |
| bio12 | $\frac{{277.2 \pm 6.2}}{{201.0 - 512.0}}$ | $\frac{{436.4 \pm 18.0}}{{170.0 - 1025.0}}$ | $\frac{{335.8 \pm 7.8}}{{154.0 - 613.0}}$ |
| bio13 | $\frac{{46.9 \pm 1.3}}{{37.0 - 101.0}}$ | $\frac{{67.3 \pm 2.2}}{{32.0 - 130.0}}$ | $\frac{{56.8 \pm 1.1}}{{28.0 - 97.0}}$ |
| bio14 | $\frac{{7.0 \pm 0.3}}{{4.0 - 13.0}}$ | $\frac{{14.3 \pm 1.1}}{{5.0 - 68.0}}$ | $\frac{{10.0 \pm 0.3}}{{3.0 - 46.0}}$ |
| bio15 | $\frac{{61.1 \pm 1.0}}{{49.3 - 81.0}}$ | $\frac{{50.9 \pm 1.1}}{{15.0 - 85.4}}$ | $\frac{{56.4 \pm 0.9}}{{30.7 - 89.2}}$ |
| bio16 | $\frac{{130.3 \pm 3.5}}{{93.0 - 273.0}}$ | $\frac{{180.7 \pm 5.7}}{{82.0 - 347.0}}$ | $\frac{{150.6 \pm 2.9}}{{72.0 - 258.0}}$ |
| bio17 | $\frac{{24.9 \pm 0.8}}{{15.0 - 43.0}}$ | $\frac{{52.3 \pm 3.2}}{{20.0 - 213.0}}$ | $\frac{{37.8 \pm 1.2}}{{15.0 - 154.0}}$ |
| bio18 | $\frac{{129.7 \pm 3.5}}{{90.0 - 273.0}}$ | $\frac{{156.3 \pm 3.9}}{{77.0 - 307.0}}$ | $\frac{{141.7 \pm 2.2}}{{29.0 - 291.0}}$ |
| bio19 | $\frac{{32.0 \pm 0.9}}{{19.0 - 55.0}}$ | $\frac{{75.4 \pm 4.9}}{{25.0 - 252.0}}$ | $\frac{{52.4 \pm 1.6}}{{16.0 - 216.0}}$ |
Примечание. Биоклиматические переменные: bio1 – среднегодовая температура; bio2 – средний дневной диапазон; bio3 – изотермичность; bio4 – сезонность температуры (стандартное отклонение ×100); bio5 – максимальная температура самого теплого месяца; bio6 – минимальная температура самого холодного месяца; bio7 – температурный годовой диапазон; bio8 – средняя температура самого влажного квартала; bio9 – средняя температура самого засушливого квартала; bio10 – средняя температура самого теплого квартала; bio11 – средняя температура самого холодного квартала; bio12 – годовое количество осадков; bio13 – осадки самого влажного месяца; bio14 – осадки самого засушливого месяца; bio15 – сезонность осадков (коэффициент вариации); bio16 – осадки самого влажного квартала; bio17 – осадки самого засушливого квартала; bio18 – осадки самого теплого квартала; bio19 – осадки самого холодного квартала. Над чертой – среднее значение и стандартная ошибка среднего (M ± SE), под чертой – min–max.
Значимые различия коэффициентов в регрессионных моделях (MLR, p < 0.05) получены для S. emersum “rothertii” в паре с S. emersum s. str. почти по всем биоклиматическим переменным температуры и осадков (Доп. мат. табл. S2 ), за редким исключением (изотермичность (bio3), максимальная температура самого теплого месяца (bio5), средняя температура самого влажного квартала (bio8), осадки самого засушливого месяца (bio14)). В свою очередь, значимые различия S. emersum “rothertii” с S. hyperboreum установлены только для некоторых переменных по температуре (средний дневной диапазон (bio2), максимальная температура самого теплого месяца (bio5), температурный годовой диапазон (bio7), средняя температура самого засушливого квартала (bio9)) и по осадкам (годовое количество осадков (bio12), сезонность осадков (bio15), осадки самого влажного квартала (bio16), осадки самого засушливого квартала (bio17)), т.е. S. emersum “rothertii” по ряду показателей сопоставим с S. hyperboreum, по-видимому, этим можно объяснить высокую степень перекрытия находок двух таксонов в пространственных градиентах (Доп. мат. табл. S3 ).
Регрессионные модели по градиенту среднегодовой температуры демонстрируют, что на СВА по мере роста значений увеличивается вероятность находок S. emersum “rothertii”, в свою очередь, шансы S. emersum s. str. снижаются. На участках СВА со среднегодовой температурой на нижнем пределе (–16.7°С) вероятность находок в ряду S. emersum s. str., S. emersum “rothertii” и S. hyperboreum – 29, 8 и 63%. На верхнем пределе среднегодовой температуры (0.9°С) вероятность находок этих таксонов – 3, 65 и 32% соответственно. В градиенте температурного годового диапазона шансы обнаружить S. emersum “rothertii” снижаются поступательно. При этом температура 50°–60° по данному показателю представляет собой некий “критичный” участок градиента, поскольку существенно меняются направленность и динамика значений вероятностей находок S. hyperboreum и S. emersum s. str. (рис. 3г, 3д). Рост количества осадков в самый холодный квартал сопровождается снижением шансов обнаружить S. emersum s. str. на СВА, и увеличением вероятности произрастания S. emersum “rothertii” и S. hyperboreum.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Географические и экологические факторы. Показано, что пресноводные макрофиты в Европе и Северной Америке, распределены по числу видов, биомассе и другим параметрам в географических градиентах довольно неравномерно, что объясняется неоднородностью окружающей среды (Alahuhta et al., 2017; García-Girón et al., 2021). Полученные по широко распространенным Sparganium на СВА данные свидетельствуют о справедливости этих наблюдений.
Большинство местонахождений ежеголовников на СВА сосредоточено в долинах крупных рек (Лена, Вилюй, Колыма, Омолон) или недалеко от морских побережий. Эти территории характеризуются прерывистым распространением многолетней мерзлоты и (или) обширными таликовыми зонами. Мощность деятельного слоя, состоящего из сезонно-талого и сезонно-мерзлого слоев на территории Якутии в основных местах произрастания всех трех таксонов от 1.5–2 до 3 м (Федоров и др., 2019).
На территории СВА только S. hyperboreum встречается повсеместно, в том числе и за пределами крупных речных долин, он достаточно равномерно распределен по районам водоразделов и межгорных равнин и произрастает в водоемах почти на всех доступных уровнях высот. При этом большая часть находок S. emersum s. str. и S. emersum “rothertii” приходится на долинные местообитания и террасы речных долин.
Проведенный анализ данных по долготе, широте и высоте демонстрирует пространственную диверсификацию ежеголовников на СВА, связанную с неоднородностью современной топографии, для которой характерно чередование локальных барьеров – широких речных долин и горных систем в мелком масштабе территории и обилие однородных местообитаний и водных объектов – рек, ручьев и озер – в крупном. В этом аспекте полученные данные согласуются с наблюдаемыми региональными (Stuckey, 1993; Santamaría, 2002; Sun et al., 2022) и глобальными тенденциями (Murphy et al., 2019).
S. emersum произрастает по озерам и старицам, в мочажинах на болотах и в тундрах, реже в реках и протоках. Имеет широкую экологическую амплитуду, может встречаться и в олиготрофных, и в мезотрофных водоемах с рН 6–8.5. На СВА качество воды не имеет большого значения для этого вида, хотя по данным Cook, Nichols (1986) чаще он встречается в мезотрофных водах, реже в олиготрофных или сильно эвтрофных. На СВА S. emersum может произрастать в водоемах с часто меняющимся уровнем воды и формировать наземную форму, а также в глубоких озерах и реках, формируя погруженную, относительно узколистную и редко цветущую глубоководную форму. Именно широкая экологическая амплитуда и разнообразие экотопов обусловливают высокий внутривидовой полиморфизм.
S. hyperboreum встречается по тундровым, горным и долинным озерам, мочажинам, временным водоемам, реже – по медленно текущим неглубоким рекам; фоновый вид – по мелким термокарстовым озерам в горных и равнинных тундрах на СВА; для настоящей работы образцы были собраны в водоемах с рН 5.4–8.1 и минерализацией 2–65 ppm, что указывает на предпочтение вида – слабокислые неглубокие водоемы с торфянистым дном. По мнению Cook, Nicholls (1986), S. hyperboreum не растет ни в ультраолиготрофных, ни в эвтрофных водоемах, однако на СВА его неоднократно находили в ультраолиготрофных водоемах с очень чистой почти дистиллированной водой. Нередко вид встречается в мелких озерках и мочажинах, полностью перемерзающих зимой.
Биоклиматические факторы. Распространение водных растений во многом определяется влиянием климатических факторов, которые изменяют среду обитания (Woodward, Williams, 1987; Dudgeon et al., 2006; Corlett, Westcott, 2013), тем самым обусловливают миграцию в более благоприятные местообитания, нередко на значительные расстояния (Chen et al., 2011; Viana, 2017). Santamaría (2002), напротив, считает, что климатические факторы имеют ограниченное влияние на водные растения, особенно на широко распространенные виды.
Полученные значения параметров биоклиматических факторов, на основе данных о распространении S. emersum s. str., S. emersum “rothertii” и S. hyperboreum на СВА, варьируют для этих таксонов, но по ряду показателей близки между собой (табл. 1). Они составляют биоклиматические характеристики этих видов ежеголовников для СВА.
S. emersum “rothertii” предпочитает местообитания менее контрастные по температуре и с большим обилием осадков, что свойственно территориям с климатом, находящимся под океаническим влиянием. Отчасти это позволяет S. emersum “rothertii” занимать сходные местообитания и произрастать совместно с S. hyperboreum в низкогорных условиях и с S. emersum s. str. в долинах крупных рек арктических районов СВА. По режиму увлажнения S. emersum s. str., сравнительно с S. emersum “rothertii” и S. hyperboreum, предпочитает более сухие местообитания (табл. 1).
Итак, расчеты показали, что дифференциацию местообитаний Sparganium на СВА обусловливают в первую очередь семь факторов: средний дневной диапазон температуры (bio2), температурный годовой диапазон (bio7), средняя температура самого засушливого квартала (bio9), годовое количество осадков (bio12), сезонность осадков (bio15), осадки самого влажного квартала (bio16) и осадки самого засушливого квартала (bio17). Эти биоклиматические переменные могут быть использованы в качестве основных факторов, определяющих распространения ежеголовников на СВА. Неинформативными биоклиматическими переменными для S. emersum s. str., S. emersum “rothertii” и S. hyperboreum на СВА оказались только три из 19: изотермичность (bio3), средняя температура самого влажного квартала (bio8) и осадки самого засушливого месяца (bio14), т.е. подавляющее большинство переменных можно успешно использовать в качестве предикторов.
Известно, что водная среда оказывает нивелирующие действие на температуру, однако, наблюдается дифференциация распространения ежеголовников на СВА в градиенте биоклиматических температурных переменных. Полученные данные подтверждают важную детерминирующую роль температуры как фактора распространения водных растений (Barko et al., 1982; Sun et al., 2022). Однако отметим, что S. emersum и S. hyperboreum чаще произрастают по мелководьям, где “нивелирование” водой температуры воздуха наиболее слабо.
Изменчивость вероятностей находок видов ежеголовников на СВА в градиентах геопространственных и биоклиматических переменных обычно имеет нелинейный вид (рис. 3). По всей видимости, это обусловлено неравномерным распространением трех таксонов вдоль экологических градиентов, редко учитываемыми важными для водных растений локальными факторами (гидрохимия воды, мерзлотный режим и колебания уровня воды в водоемах), а также историческими причинами. Геопространственная дифференциация, в свою очередь, определена экологическими характеристиками Sparganium. Так, S. hyperboreum характеризуется наиболее широким спектром местообитаний. Мощность деятельного слоя в местах его произрастания начинается от 1 м. S. hyperboreum может произрастать в горных озерах и мелких термокарстовых озерках, которые на крайнем северо-востоке нередко перемерзают до дна. S. emersum “rothertii” по большинству биоклиматических показателей ближе к S. hyperboreum (например, по изотермичности, сезонности температуры, максимальным температурам самого теплого месяца, средним температурам самого засушливого квартала и др.). S. emersum s. str. отличается от S. hyperboreum сильнее по температурному годовому диапазону, средним температурам самого влажного и самого теплого кварталов, а также осадкам самого засушливого и самого холодного кварталов.
Надо сказать, что особенности распространения ежеголовников на СВА, подтверждают результаты общего анализа воздействия окружающей среды на водные растения (Lacoul, Freedman, 2006). Авторами показано, что некоторые водные растения, с плюризональным распространением, характеризуются высокими уровнями полиморфизма и фенотипической пластичности в ответ на изменения факторов окружающей среды, и это позволяет им существовать в широком диапазоне условий среды.
Выводы. Среди ежеголовников наиболее широко распространен Sparganium hyperboreum – произрастает на всем пространстве СВА, проникает высоко в горы и далеко в Заполярье, занимает наиболее широкий спектр местообитаний. Реже встречаются S. emersum s. str., и S. emersum “rothertii”, локализованные в западной и восточной частях региона соответственно. Оба тяготеют к низинам, равнинам и речным долинам. При этом S. emersum s. str. более требователен к теплообеспеченности и сухости климата по сравнению с другими. Расчеты показывают, что S. emersum s. str. и S. hyperboreum в пределах СВА располагаются на терминальных участках геопространственных и биоклиматических градиентов. В свою очередь, S. emersum “rothertii” занимает некоторое промежуточное положение, смещаясь по многим показателям в сторону S. hyperboreum. Перекрытие пространственных данных и близость значений отдельных биоклиматических переменных у разных таксонов Sparganium на СВА могут быть обусловлены локальными сложно учитываемыми для водных растений факторами – размер и тип водоема, кислотность и минерализация, глубина промерзания и др. Вся территория северо-востока Азии, с большим разнообразием водоемов и водотоков, по геопространственным и биоклиматическим параметрам пригодна для произрастания S. emersum и S. hyperboreum, а их отсутствие на каких-либо участках связано или с многофакторной комбинацией природно-климатических условий, или же с историческими причинами. Полученные количественные оценки биоклиматических параметров дают возможность прогнозировать изменение границ их ареалов на СВА при различных сценариях изменения климата и ожидать расширения ареала S. emersum при потеплении климата.
Список литературы
Алисов Б.П. 1956. Климат СССР. М.: Изд-во Мос. ун-та.
Андриянова Е.А., Мочалова О.А., Мовергоз Е.А. 2019. Экология и сезонное развитие Ranunculus nipponicus Nakai (Batrachium, Ranunculaceae) в незамерзающих водотоках северного побережья Охотского моря // Биология внутр. вод. № 4. Вып. 1. С. 33.https://doi.org/10.1134/S0320965219040211
Бобров А.А., Волкова П.А., Иванова М.О., Тихомиров Н.П. 2021. Дополнения к списку водных сосудистых растений острова Cахалин // Бот. журн. Т. 106. № 9. С. 902. https://doi.org/10.31857/S0006813621090039
Бобров А.А., Мочалова О.А. 2014. Заметки о водных сосудистых растениях Якутии по материалам якутских гербариев // Новости систем. высш. раст. Т. 45. С. 122.
Бобров А.А., Мочалова О.А. 2017. Водные сосудистые растения долины Колымы: разнообразие, распространение, условия обитания // Бот. журн. Т. 102. № 10. С. 1347. https://doi.org/10.1134/S0006813617100015
Вехов Н.В. 1993. Высшие растения водоемов в низовьях рек Утавээм и Чегитун (Чукотский полуостров) // Бот. журн. Т. 78. № 7. С. 45.
Клюкин Н.К. 1970. Климат // Север Дальнего Востока. Москва: Наука. С. 101.
Определитель высших растений Якутии. 2020. Москва: Тов-во науч. изд. КМК; Новосибирск: Наука. С. 72.
Пармузин Ю.П. 1964. Средняя Сибирь. Москва: Мысль.
Тимохина С.А. 1988. Sparganium L. – Ежеголовник // Флора Сибири. Т. 1. Lycopodiaceae–Hydrocharitaceae. Новосибирск: Наука. С. 88, 160.
Толмачев А.И. 1960. Род Sparganium L. – Ежеголовник // Арктическая флора СССР. Вып. 1. Москва, Ленинград: Изд-во АН СССР. С. 76.
Труфанова Е.Р. 1972. Растительность озер долины среднего течения реки Колымы // Почвенные и ботанические исследования в Якутии. Якутск: Кн. изд-во. С. 110.
Федоров А.Н., Шестакова А.А., Торговкин Я.И. и др. 2019. Цифровое тематическое картирование современного состояния мерзлотных ландшафтов в Якутии // Вест. Сев.-вост. фед. ун-та им. М.К. Аммосова. Сер. Науки о земле. № 2 (14). С. 36. https://doi.org/10.25587/SVFU.2019.14.35445
Цвелев Н.Н. 1984. Заметки о некоторых гидрофильных растениях флоры СССР // Новости систем. высших растений. Т. 21. Ленинград: Наука. С. 232.
Цвелев Н.Н. 1996. Род 1. Ежеголовник – Sparganium L. // Сосудистые растения советского Дальнего Востока. Т. 8. Санкт-Петербург: Наука. С. 346.
Юзепчук С.В. 1934. Сем. Ежеголовниковые – Sparganiaceae Engl. // Флора СССР. Т. 1. Ленинград: Изд-во АН СССР. С. 216.
Alahuhta J., Heino J., Luoto M. 2011. Climate change and the future distributions of aquatic macrophytes across boreal catchments // J. Biogeography. V. 38(2). P. 383. https://doi.org/10.1111/j.1365-2699.2010.02412.x
Alahuhta J., Kosten S., Munemitsu A. et al. 2017. Global variation in the beta diversity of lake macrophytes is driven by environmental heterogeneity rather than latitude // Journal Biogeography. V. 88(8). P. 1758. https://doi.org/10.1111/jbi.12978
Banerjee A.K., Harms N.E., Mukherjee A. et al. 2020. Niche dynamics and potential distribution of Butomus umbellatus under current and future climate scenarios in North America // Hydrobiologia. V. 847(10). P. 1505. https://doi.org/10.1007/s10750-020-04205-1
Barko J.W., Hardin D.G., Matthews M.S. 1982. Growth and morphology of submersed freshwater macrophytes in relation to light and temperature // Can. J. Bot. V. 60(6). P. 877. https://doi.org/10.1139/b82-113
Bornette G., Puijalon S. 2011. Response of aquatic plants to abiotic factors: a review // Aquat. Sci. V. 73. P. 1. https://doi.org/10.1007/s00027-010-0162-7
Chemeris E.V., Romanov R.E., Kopyrina L.I. et al. 2020. How charophytes (Streptophyta, Charales) survive in severe conditions of the permafrost area in Far North-East Asia // Limnologica. V. 83. P. 125784. https://doi.org/10.1016/j.limno.2020.125784
Chen I.-C., Hill J.K., Ohlemüller R. et al. 2011. Rapid range shifts of species associated with high levels of climate warming // Science. V. 333(6045). P. 1024. https://doi.org/10.1126/science.1206432
Cook C.D.K., Nicholls M.S. 1986. A monographic study of the genus Sparganium (Sparganiaceae). Part 1. Subgenus Xanthosparganium Holmberg // Bot. Helv. V. 96(2). P. 213.
Corlett R.T., Westcott D.A. 2013. Will plant movements keep up with climate change? // Trends Ecol. Evol. V. 28(8). P. 482. https://doi.org/10.1016/j.tree.2013.04.003
Dudgeon D., Arthington A.H., Gessner M.O. et al. 2006. Freshwater biodiversity: importance, threats, status and conservation challenges // Biol. Rev. V. 81(2). P. 163. https://doi.org/10.1017/S1464793105006950
Engel J. 2008. Polytomous logistic regression // Statistica Neerlandica. V. 42(4). P. 233. https://doi.org/10.1111/j.1467-9574.1988.tb01238.x
Fick S.E., Hijmans R.J. 2017. WorldClim 2: new 1 km spatial resolution climate surfaces for global land areas // Int. J. Climatology. V. 37(12). P. 4302. https://www.worldclim.org/data/worldclGarcнa-Girynim21. html
García-Girón J., Heino J., Iversen L. et al. 2021. Rarity in freshwater vascular plants across Europe and North America: Patterns, mechanisms and future scenarios // Science Total Env. V. 786. 147491. https://doi.org/10.1016/j.scitotenv.2021.147491
Gillard M.B., Thiébaut G., Deleu C., Leroy B. 2017. Present and future distribution of three aquatic plants taxa across the world: decrease in native and increase in invasive ranges // Biol. Invasions. V. 19. P. 2159. https://doi.org/10.1007/s10530-017-1428-y
Hossain K., Yadav S., Quaik S. et al. 2017. Vulnerabilities of macrophytes distribution due to climate change // Theor. Appl. Climatol. V. 129. P. 1123. https://doi.org/10.1007/s00704-016-1837-3
Lacoul P., Freedman B. 2006. Environmental influences on aquatic plants in freshwater ecosystems // Env. Rev. V. 14(2). P. 89–136. https://doi.org/10.1139/A06-001
McDonald J.H. 2009. Handbook of biological statistics. Baltimore: Sparky House Publ.
Murphy K., Efremov A., Davidson T.A. et al. 2019. World distribution, diversity and endemism of aquatic macrophytes // Aquat. Bot. V. 158. 103127. https://doi.org/10.1016/j.aquabot.2019.06.006
Santamaría L. 2002. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment // Acta Oecol. V. 23(3). P. 137. https://doi.org/10.1016/S1146-609X(02)01146-3
Stuckey R.L. 1993. Phytogeographical outline of aquatic and wetland angiosperms in continental eastern North America // Aquat. Bot. V. 44. № 2–3. P. 259. https://doi.org/10.1016/0304-3770(93)90073-6
Sun J., Doeser A., Cao Y. et al. 2022. Regional macrophyte diversity is shaped by accumulative effects across waterbody types in southern China // Aquat. Bot. V. 176. 103468. https://doi.org/10.1016/j.aquabot.2021.103468
Takahashi H., Sato T., Solomonov N.G., Ivanov B.I. 1994. Phytogeographic notes on some aquatic plants in Yakutia, Eastern Siberia // Acta Phytotax. Geobot. V. 45(2). P. 111.
Takahashi H., Volotovskyi K.A., Sato T. 2001. A quantitative comparison of distribution patterns in four common Sparganium species in Yakutia, Eastern Siberia // Acta Phytotax. Geobot. V. 51(2). P. 155.
Viana D.S. 2017. Can Aquatic Plants Keep Pace with Climate Change // Front. Plant Sci. V. 8. P. 1906. https://doi.org/10.3389/fpls.2017.01906
Woodward F.I., Williams B.G. 1987. Climate and plant distribution at global and local scales // Vegetatio. V. 69. P. 189. https://doi.org/10.1007/BF00038700
Wu H., Carrillo J., Ding J. 2016. Invasion by alligator weed, Alternanthera philoxeroides, is associated with decreased species diversity across the latitudinal gradient in China // J. Plant Ecol. V. 9 (3). P. 311. https://doi.org/10.1093/jpe/rtv060
Дополнительные материалы
- скачать ESM_1.xlsx
- Приложение 1.
Table S 1. Primary dataset for analysis
- скачать ESM_2.docx
- Приложение 2.
Table S2. Descriptive statistical characteristics of bioclimatic variables by Sparganium species (Sr – S. emersum “rothertii”, Se – S. emersum, S. hyperboreum) in NEA
Table S3. Polynomial regression coefficients of bioclimatic variables by Sparganium species (Se – S. emersum, Sr – S. emersum “rothertii”, S. hyperboreum) in NEA.
- скачать ESM_3.R
- Приложение 3.
Инструменты
Биология внутренних вод