

Ботанический журнал, 2019, T. 104, № 10, стр. 1588-1597
БИОЛОГИЯ РАЗВИТИЯ И РАСПРОСТРАНЕНИЕ ADENOPHORA TAURICA (CAMPANULACEAE) В КРЫМУ
С. В. Шевченко 1, *, Е. С. Крайнюк 1, **
1 ФГБУН “Никитский ботанический сад – Национальный научный центр РАН”
298648 г. Ялта, пгт. Никита, Россия
* E-mail: shevchenko_nbs@mail.ru
** E-mail: krainuk54@mail.ru
Поступила в редакцию 14.05.2019
После доработки 31.07.2019
Принята к публикации 02.08.2019
Аннотация
Представлены результаты изучения ценопопуляции Adenophora taurica в Горном Крыму. Описаны цветение, строение генеративных структур, опыление, начальные этапы эндоспермо- и эмбриогенеза, а также возрастная структура ценопопуляции. Показана необходимость разработки дополнительных мер по сохранению данного вида.
Проблема сохранения биоразнообразия в настоящее время является одной из наиболее актуальных, в связи с чем особое внимание приобретают редкие и нуждающиеся в охране виды растений. К таким видам относится бубенчик крымский –Adenophora taurica (Sukacz.) Juz. из сем. Campanulaceae. В чеклисте “Природная флора Крымского полуострова” (Yenа, 2012) и Красной книге Республики Крым (Коrzhenevsky, 2015) вид приводится как бубенчик лилиелистный (Adenophora liliifolia (L.) Ledeb.exA.DC.).
В Крыму A. taurica известен только из нескольких изолированных местонахождений в верхнем поясе Главной гряды Крымских гор. В Красной книге Республики Крым указывается всего три локалитета вида (Коrzhenevsky, 2015), в т. ч. на луговых яйлинских склонах гор Большая и Малая Чучель в Крымском природном заповеднике (Коstina, Bagrikova, 2010). Вид указывается как эндем Крыма (Golubev, 1996; Govaerts, 2019). Основными лимитирующими факторами освоения новых территорий в Крыму A. taurica можно считать глобальную трансформацию климата и сужение экологической ниши вида (Коrzhenevsky, 2015), а также природно-историческую редкость вида, низкую конкурентную способность на границе ареала, пространственную разобщенность популяций, антропогенные нарушения среды обитания. Целью наших исследований является выявление особенностей воспроизведения A. taurica в изучаемой ценопопуляции в связи с необходимостью разработки приемов сохранения данного вида.
МАТЕРИАЛ И МЕТОДИКА
Наблюдения проводили в условиях естественного произрастания Adenophora taurica в Горном Крыму в районе Гурзуфского седла. Растительный материал фиксировали фиксатором Карнуа. Парафиновые срезы толщиной 10–12 мкм готовили с использованием ротационного полуавтоматического микротома RMD-3000 (Россия). Постоянные препараты приготовлены и окрашены по общепринятым методикам (Romeis, 1954; Pausheva, 1990; Shevchenko et al., 1986; Shevchenko, Chebotaru, 1992). Анализ препаратов проводили с помощью микроскопов AxioScopeA.1 и Enamed 2 (“CarlZeiss”, Германия). Микрофотографии получены системой анализа изображений AxioCamERc5s (“CarlZeiss”, Германия) и цифровыми фотокамерами Canon IXUS 256HS и CanonA 550.
Возрастную структуру ценопопуляции изучали в природных условиях согласно общепринятым методикам (Rabotnov, 1969; Uranov, 1975; Coenopopulation…, 1976; 1977). Полевые учеты численности особей по возрастным состояниям проводили на площадках в 0.25 м2, а затем производили пересчет на 1 м2. При этом были использованы следующие условные обозначения возрастных групп: p – всходы, j – ювенильные, im – имматурные, $v$ – виргинильные, g1 – молодые генеративные, g2 – средневозрастные генеративные.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Изучаемая ценопопуляция Adenophora taurica находится на территории Ялтинского горно-лесного природного заповедника (Гурзуфское седло – Бабуган-яйла) на прияйлинских склонах южного макросклона Главной гряды Крымских гор на высоте примерно 1200 м н.у.м. в ущелье Кумбасан ниже перевала Гурзуфское Седло на крутом склоне в сосновом лесу (рис. 1).
Рис. 1.
Карта места нахождения ценопопуляции Adenophora taurica.
Fig. 1. Map of the location of the Adenophora taurica coenopopulation.
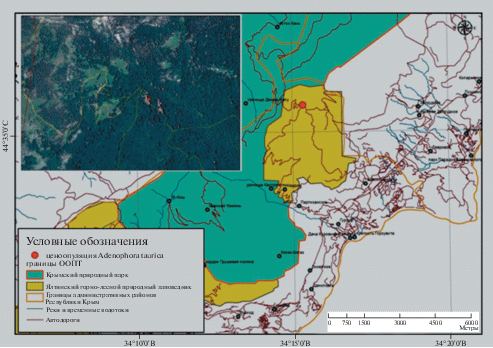
Ценопопуляция в настоящее время является единственным подтвержденным местонахождением данного вида в Крыму, обнаруженным в 2017 году С.А. Свириным. Гербарный образец из данного места сбора хранится в гербарии YALT.
Тип растительности – сосновый лес из Pinus kochiana (Ass. Pimpinello-Pinetum kochianae Korzh. 1986). Травостой злаково-разнотравный, общее проективное покрытие травостоя 50–60%, видовая насыщенность фитоценоза – невысокая.
Ценопопуляция занимает площадь около 100 м2 на крутом (30–40°) склоне балки. Проективное покрытие вида в фитоценозе составляет 5–10%.
A. taurica – травянистый многолетник высотой 30–50 см, с толстым веретеновидным или редьковидным корнем и цилиндрическим, продольно-полосатым, голым или редковолосистым, густо облиственным стеблем. Стеблевые листья очередные, эллиптические, островатые, по краям неравномерно крупно-пильчатозубчатые, черепитчато перекрывающие друг друга в средней части стебля, верхние листья ланцетные, сидячие (рис. 2а). Розеточные прикорневые листья округло-сердцевидные, длинночерешковые, длиной 4–10 см, быстро засыхающие (рис. 2б).
Рис. 2.
Общий вид генеративной (а) и виргинильной (б) особей Adenophora taurica.
Fig. 2. General view of the generative (а) and virginal (b) plants of Adenophora taurica.

Цветет A. taurica в июле–августе, цветение акропетальное, цветки собраны в кистевидное соцветие. Венчик колокольчатый, голубой или светло-синий, неглубоко раздельный на 5 лопастей, длиной до 1.5 см, цветки пониклые, спайнолепестные (рис. 3). Чашечка голая, ее зубцы острые, ланцетовидно-треугольные, равные по длине трубке. Андроцей представлен пятью свободными, плотно прилегающими друг к другу тычинками с прямыми перепончатыми и сильно реснитчатыми тычиночными нитями. У основания тычиночные нити плотно сомкнуты и образуют купол с небольшими отверстиями, через которые насекомые-опылители хоботком могут достигать нектарный диск, который расположен у основания столбика. Стенка микроспорангия, развивающаяся центробежно, состоит из эпидермы, эндотеция, одного среднего слоя и секреторного тапетума. Спорогенная ткань в микроспорангии представлена двумя-тремя слоями клеток. В зрелом пыльнике стенка состоит из сплющенных клеток эпидермы, эндотеция с фиброзными утолщениями и тапетальной пленки с орбикулами (рис. 4). Зрелые пыльцевые зерна 2-клеточные, 3-борозднопоровые.
Рис. 3.
Общий вид растений Adenophora taurica.
Fig. 3. General view of the Adenophora taurica plants.

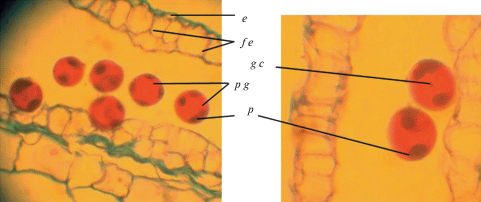
Рис. 4.
Фрагменты зрелых пыльников с пыльцевыми зернами: e – эпидерма, f e – фиброзный эндотеций, g c – генеративная клетка, p g – пыльцевые зерна, p – поры.
Fig. 4. Fragments of mature anthers with pollen grains: e – epidermis, f e – fibrous endothecium, g c – generative cell, p g – pollen grains, p – pores.
Гинецей A. taurica образован тремя плодолистиками. Пестик открытый, столбик имеет канал с сосочковидными волосками эпидермального происхождения. Во время цветения пестик сильно выдается за пределы венчика (рис. 3).
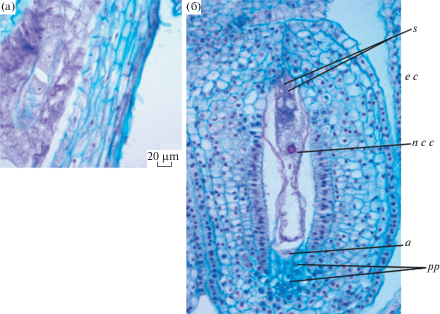
Завязь трехгнездная. Семязачаток, согласно классификации И.И. Шамрова (Shamrov, 2008; 2017), анатропный, унитегмальный, медионуцеллятный, мезохалазальный. Развивается интегументальный тапетум, представленный радиально вытянутыми крупными клетками и достигающий уровня яйцевого аппарата зародышевого мешка. Имеются постаменто-подиум и гипостаза. Зародышевый мешок развивается из халазальной мегаспоры по Polygonum-типу (рис. 5а), содержит крахмальные зерна (рис. 5б).
Рис. 5.
Тетрада мегаспор (а) и зрелый зародышевый мешок (б) Adenophora taurica: s – синергиды, e c – яйцеклетка, n c c – ядро центральной клетки, a – антиподы, pp – поcтаментоподиум.
Fig. 5. Megaspore tetrad (а) and mature embryo sac (b) of Adenophora taurica: s – synergids, e c – egg cell, n c c – nucleus of central cell, a – antipods, pp – postamentopodium.
Опыление происходит с помощью насекомых и волосков на столбике пестика. У A. taurica четко выражена протандрия. При этом в пыльниках наблюдается зрелая пыльца, а женская сфера еще не готова к принятию пыльцы. Рыльца пестика сомкнуты, и зародышевый мешок еще не дифференцирован. Пыльники вскрываются интрорзно, и пыльца из них извлекается волосками растущего столбика. Насекомые-опылители, проникая к нектарному диску и обратно, снимают пыльцу и переносят ее на другие цветки, осуществляя таким образом перекрестное опыление (рис. 6). После извлечения пыльцы из пыльников и снятия ее насекомыми со столбика, покрывающие его волоски втягиваются в поверхностную ткань столбика, расширяя основание волоска. Остаются только кончики волосков, которые незначительно возвышаются над эпидермой столбика, то есть наблюдается явление инвагинации, или ретракции.
Рис. 6.
Фрагмент генеративной особи с насекомым-опылителем (а) и часть столбика с эпидермальными волосками (б).
Fig. 6. Fragment of a generative plant with an insect pollinator (a) and a part of a column with epidermal hairs (b).

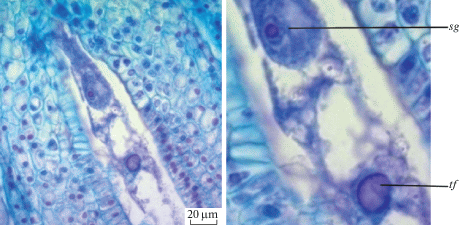
Попав на рыльце пестика, пыльцевые зерна прорастают. Пыльцевая трубка проходит по каналу, в ней происходит спермиогенез с образованием двух спермиев. Достигнув зародышевого мешка, пыльцевая трубка лопается и изливает свое содержимое в полость между яйцеклеткой и центральной клеткой. Один спермий сливается с яйцеклеткой, а второй – с ядром центральной клетки (рис. 7).
Рис. 7.
Двойное оплодотворение у Adenophora taurica: sg – сингамия, t f – тройное слияние.
Fig. 7. Double fertilization in Adenophora taurica: sg – syngamy, t f – triple fusion.
Первичное ядро эндосперма сначала находится в середине центральной клетки, затем постепенно опускается к халазальному концу, где и происходит его первое деление. Оно поперечное с образованием двух неравных клеток – халазальная клетка значительно меньше микропилярной. Ядро халазальной клетки делится один или два раза, несколько увеличивается и образуется небольшой гаусторий. Микропилярная клетка дважды делится поперечно. Из апикальной клетки формируется собственно эндосперм, а базальная либо значительно увеличивается в размерах либо делится продольно и образует двуклеточный гаусторий. Клетки эндосперма крупные, их ядра активны, с ядрышками. Зигота некоторое время находится в покое, затем активно растет и значительно вытягивается внутрь эндосперма. Ядро находится в апикальном конце зиготы и первые его деления поперечные, с образованием ряда клеток (рис. 8). Последующие продольные и поперечные деления позволяют предположить Solanad-тип эмбриогенеза, свойственный большинству представителей сем. Campanulaceae (Korobova, Zhinkina, 1987; Zhinkina,1995; Kamelina, 2009).
Рис. 8.
Некоторые этапы формирования зародыша Adenophora taurica: h – гаусторий, pr – проэмбрио.
Fig. 8. Some stages of the embryo formation in Adenophora taurica: h – haustorium, pr – proembryo.

Плодоносит A. taurica в августе–сентябре. Плод – пониклая, яйцевидной формы коробочка, которая в нижней части открывается тремя створками или порами. Семена мелкие 1–2 мм, сплющенные, рыжеватые. Следует отметить, что почти половина цветков соцветия опадает неопыленной и семян не образует.
Возрастная структура и численность ценопопуляций данного вида в Крыму ранее не изучались. Изучаемая ценопопуляция представлена в виде двух ценопопуляционных локусов – в верхней и нижней части склона ущелья.
Согласно нашим наблюдениям в 2017–2018 гг., численность вида составляет около 100 разновозрастных особей. Вид произрастает одиночно или группами, состоящими из нескольких особей. Численность побегов у генеративных особей составляет 1–5 (Krainyuk, Shevchenko, 2018).
Плотность особей в ценопопуляции составляет: в нижнем локусе – 3.7 особей на 1 м2, в верхнем локусе – 16.0 особей на 1 м2. По типу возрастной структуры изученная ценопопуляция является нормальной, неполночленной, молодой, с преобладанием виргинильных особей, о чем свидетельствуют составленные возрастные спектры (рис. 9). В верхнем локусе (исследования 2017 г.) установлено следующее соотношение возрастных групп: j – 16.7%, im – 23.3%, $v$ – 45.0%, g1 – 3.3%, g2 –11.7%. В нижнем локусе (исследования 2018 г.) соотношение возрастных групп следующее: p – 2.7%, j – 8.2%, im – 37.8%, $v$ –29.7%, g2 – 21.6%. Особи других возрастных состояний при полевых учетах не были обнаружены.
Рис. 9.
Возрастная структура ценопопуляции Adenophora taurica (прямая линия – верхний локус, линия пунктиром – нижний локус).
Fig. 9. Age structure of the Adenophora taurica coenopopulation (straight line – upper locus, dotted line – lower locus).
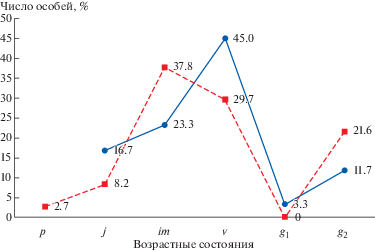
Анализ возрастной структуры изученной ценопопуляции позволяет сделать вывод о том, что естественное возобновление вида осуществляется в достаточной мере, поскольку преобладающей группой являются особи виргинильного периода, составляющие 85% в верхнем локусе и 78.4% – в нижнем. Группа генеративных особей составляет 15% в верхнем локусе и 21.6% – в нижнем локусе, что свидетельствует о достаточных возможностях вида для перехода виргинильных особей в генеративное состояние. Отсутствие в ценопопуляции особей сенильного периода, видимо, можно объяснить особенностями биологии и экологии данного вида, что требует дальнейшего изучения.
ЗАКЛЮЧЕНИЕ
Проведенное изучение формирования генеративных структур, процессов цветения, опыления и семяобразования позволяет заключить, что по основным чертам (центробежный тип развития стенки микроспорангия, Polygonum-тип развития зародышевого мешка, особенности опыления, формирование терминальных гаусториев, Solanad-тип эмбриогенеза и др.) A. taurica сходна с таковыми у других представителей сем. Campanulaceae. Основной причиной редкости данного вида в Крыму является, очевидно, очень узкая экологическая ниша вида, ограниченная высотной поясностью, крутизной склонов и степенью увлажнения экотопов. К лимитирующим факторам, кроме указанных выше, можно отнести также гидротермические условия и отсутствие насекомых-опылителей в период цветения, что приводит в конечном итоге к снижению семенной продуктивности. Необходимыми мерами охраны этого редкого вида должны быть поиск новых локалитетов, изучение динамики возрастной структуры и численности вида в локалитете, дополнительный анализ особенностей биологии, экологии, онтогенеза растений, а также введение вида в культуру с последующей репатриацией в природные условия.
Список литературы
[Cenopopulatsii] Ценопопуляции растений (основные понятия и структура). 1976. М. 215 с.
[Cenopopulatsii] Ценопопуляции растений (развитие и взаимоотношения). 1977. М. 183 с.
[Fedyaeva] Федяева В.В. 2014. Бубенчик лилиелистный. – В кн: Красная книга Ростовской области. Т. 2. Растения и грибы. Минприроды Ростовской области. Ростов-на-Дону. С. 150.
[Golubev] Голубев В.Н. 1996. Биологическая флора Крыма. Издание второе. Ялта. 86 с.
Govaerts R. (ed). For a full list of reviewers see: http://apps.kew.org/wcsp/compilers Reviewers.do (2019).
[Kamelina] Камелина О.П. 2009. Систематическая эмбриология цветковых растений. Двудольные. Барнаул. 501 с.
[Kostina, Bagrikova] Костина В.П., Багрикова Н.А. 2010. Аннотированный список высших растений Крымского природного заповедника. – Научные записки природного заповедника “Мыс Мартьян”. 1: 61–142.
[Korobova, Zhinkina] Коробова С.Н., Жинкина Н.А. 1987. Семейство Campanulaceae. – В кн: Сравнительная эмбриология цветковых растений. Т. 4. Л.: Наука. С. 300–308.
[Korzhenevsky] Корженевский В.В. 2015. Бубенчик лилиелистный. В кн.: Красная книга Республики Крым. Растения, водоросли. Грибы. Симферополь. С. 175.
[Krainyuk, Shevchenko] Крайнюк Е.С., Шевченко С.В. 2018. Adenophora liliifolia – редкий вид флоры Крыма. – В сб. Ботаника в современном мире. Труды XIV съезда Русского ботанического общества и конференции. Т. 1. Махачкала. С. 272–273.
[Pausheva] Паушева З.П. 1990. Практикум по цитологии растений. М. 283 с.
[Rabotnov] Работнов Т.А. 1969. Некоторые вопросы изучения ценотических популяций. – Бюлл. Моск. общ-ва испытателей природы, отд. биол. 74(1): 147–149.
[Romeys] Ромейс Б. 1954. Микроскопическая техника. М. 718 с.
[Shamrov] Шамров И.И. 2008. Семязачаток цветковых растений: строение, функции, происхождение. М. 350 с.
[Shamrov] Шамров И.И. 2017. Морфологические типы семязачатков цветковых растений. – Бот. журн. 102(2): 129–146.
[Shevchenko, Rugusov, Efremova] Шевченко С.В., Ругузов И.А., Ефремова Л.М. 1986. Методика окраски постоянных препаратов метиловым зеленым и пиронином. – Бюлл. Гос. Никит. ботан. сада. 60: 99–101.
[Shevchenko, Chebotaru] Шевченко С.В., Чеботарь А.А. 1992. Особенности эмбриологии маслины европейской (Olea europaea). – В сб. научн. трудов Гос. Никит. ботан. сада. 113: 52–61.
[Uranov] Уранов А.А. 1975. Возрастной спектр ценопопуляций как функция времени и энергетических волновых процессов. – Научные доклады высшей школы. Биол. науки. 2: 7–34.
[Yena] Ена А.В. 2012. Природная флора Крымского полуострова. Симферополь. 232 с.
[Zhinkina] Жинкина Н.А. 1995. Сравнительная эмбриология представителей семейства Campanulaceae: Автореф. дис… кaнд. биол. наук. СПб. 17 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал