

Ботанический журнал, 2019, T. 104, № 10, стр. 1610-1621
МОРФОЛОГИЯ МНОГОБОРОЗДНЫХ ПЫЛЬЦЕВЫХ ЗЕРЕН НЕКОТОРЫХ ВИДОВ PRIMULA (PRIMULACEAE)
Д. А. Брицкий 1, *, А. Н. Иванова 1, 2, **, В. В. Григорьева 1, ***
1 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
2 Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7–9, Россия
* E-mail: dmibri@mail.ru
** E-mail: alyx@bk.ru
*** E-mail: mikhailov_val@mail.ru
Поступила в редакцию 01.08.2019
После доработки 29.09.2019
Принята к публикации 08.10.2019
Аннотация
С помощью светового и сканирующего электронного микроскопов изучена морфология многобороздных пыльцевых зерен 16 видов рода Primula. Скульптура (размеры и очертания просветов ячей, ширина и рельеф стенок ячей), а также форма пыльцевых зерен могут служить дополнительными характеристиками отдельных видов или групп видов. На ультратонких срезах оболочек пыльцевых зерен P. elatior и P. veris показано наличие тонкого прерывистого подстилающего слоя эктэкзины.
Семейство Primulaceae объединяет от 13 до 30 родов и около 600–1000 видов травянистых растений (Cronquist, 1981; Anderberg, 2004). Палиноморфологически оно неоднородно. По числу и строению апертур традиционно выделяют 3 основных палинотипа: сплющенно-3-бороздные, 3-бороздно-оровые и многобороздные. Некоторые исследователи выделяют в семействе до 20 дополнительных палинотипов (Punt et al., 1974). Виды с многобороздной пыльцой в сем. Primulaceae встречаются только в двух родах: Primula L. и Dionysia Fenzl.
В роде Primula многобороздные пыльцевые зерна описаны для небольшого числа видов из 6 секций рода (Wendelbo, 1961; Spanowsky, 1962; Anderberg, El-Ghazaly, 2000; Morozowska, Idzikowska, 2004; Grigoryeva et al., 2018). Род Dionysia считается в палиноморфологическом отношении однородным и характеризуется одним многобороздным типом пыльцы (Wendelbo, 1961; Spanowsky, 1962; Al Wadi, Richards, 1992).
Ультраструктура оболочки многобороздных пыльцевых зерен изучена только у 3 видов рода Primula: P. officinalis (L.) Jacquinh, P. veris L. и P. sibirica Jacq. При анализе ультраструктуры оболочки ацетолизированных пыльцевых зерен P. officinalis и P. veris отмечено отсутствие подстилающего слоя в эктэкзине многобороздной пыльцы (Nowicke, Skvarla, 1977). Однако исследование ультратонких срезов пыльцевых зерен P. sibirica показало наличие тонкого, фрагментарного подстилающего слоя в экзине пыльцевых зерен этого вида (Grigoryeva et al., 2018).
Цель данной работы – оценка таксономической значимости морфологических признаков и ультраструктурных особенностей спородермы многобороздных пыльцевых зерен некоторых видов рода Primula.
МАТЕРИАЛЫ И МЕТОДИКА
Материалом для исследования послужили зрелые пыльцевые зерна, полученные с гербарных образцов, хранящихся в Гербарии Ботанического института им. В Л. Комарова (LE). Микроспоры и пыльцевые зерна P. veris также были взяты с живых растений из коллекции открытого грунта ботанического сада Петра Великого Ботанического института им. В Л. Комарова РАН.
В работе использовали методы световой (СМ), сканирующей (СЭМ) и трансмиссионной (ТЭМ) электронной микроскопии.
Для светооптического исследования применяли классический ацетолизный метод (Erdtman, 1952). Световые микрофотографии получены на микроскопе Carl Zeiss Jenaval, с помощью цифровой фотокамеры Canon EOS 20D.
Детали строения поверхности пыльцевых зерен уточняли на сканирующем электронном микроскопе JEOL JSM-6390 в центре коллективного пользования Ботанического института им. В.Л. Комарова. Для исследования необработанную пыльцу извлекали из пыльников и закрепляли на специальном столике с помощью двусторонней липкой ленты, затем напыляли сплавом золота и палладия в вакуумной установке.
Описания проводили по общепринятой схеме. Учитывали тип и число апертур, форму и очертания пыльцевых зерен, размеры полярной оси и экваториального диаметра, особенности строения борозд и межапертурных участков, толщину экзины и характер скульптуры (Kupriyanova, Aleshina, 1967, 1972). Измерения проводили с помощью окулярмикрометра. В каждом образце измеряли не менее 15 пыльцевых зерен.
Ультраструктуру оболочки изучали на ультратонких срезах с помощью трансмиссионного микроскопа Hitachi-600 и Libra 120 в центре коллективного пользования Ботанического института им. В.Л. Комарова и Jeol JEM 1400 Ресурсного центра “Развитие молекулярных и клеточных технологий” Санкт-Петербургского государственного университета. Пыльники, содержащие пыльцевые зерна, фиксировали в 2.5%-ном растворе глутарового альдегида на фосфатном буфере pH 6.8–7.2 при +4°С в течение 16–10 часов, затем в 1%-ном растворе OsO4 на том же буфере в течение 1 часа при комнатной температуре. Материал обезвоживали ацетоном с нарастающей концентрацией и заключали в эпоксидную смолу эпон EmBed 812 (EMS, США) с помощью автоматического процессора AMW (Leica, Австрия). Ультратонкие срезы изготавливали на ультратомах LKB и Leica EM UC7. Срезы на сеточках контрастировали уранилацетатом (20 мин) и цитратом свинца (10 мин).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сравнительный палиноморфологический анализ показал, что пыльцевые зерна всех изученных видов радиально симметричные, равнополярные, многобороздные.
По общепринятой классификации размеров пыльцевые зерна изученных видов Primula можно отнести к мелким (до 25 мкм) и средним (25–50 мкм) (Erdtman, 1952). Самые мелкие пыльцевые зерна характерны для длинностолбиковой формы P. nutans (полярная ось 11.6–12.5 мкм, экваториальный диаметр 12.7–13.5 мкм), самые крупные – у короткостолбиковой формы P. cordifolia (полярная ось 28.1–29.2 мкм, экваториальный диаметр 28.7–30.1 мкм). Вариабельность размеров в одном образце незначительна и не превышает 2 мкм (табл. 1).
Таблица 1.
Количественные характеристики пыльцы исследованных видов рода Primula Table 1. The quantitative characteristics of the pollen of the studied Primula species
| Исследованный образец Sample |
Число борозд Aperture number |
Длина
полярной оси (мкм) Polar axis length (μm) |
Длина экваториального
диаметра (мкм) Equatorial axis length (μm) |
Толщина экзины (мкм) Exine thickness (μm) |
|---|---|---|---|---|
| Sect. Carolinella | ||||
| P. megaseifolia Boiss et Bal. ex Boiss. Transcaucasia occidentalis. 05.02.1907. A. Fomin (LE) | 8 | 26.2–27.8 | 24.4–25.7 | 0.7–0.8 |
| Sect. Cortusoides | ||||
| P. paxiana Kuntze, короткостолбиковая форма. Ботанический сад БИН РАН. 25.05.1996. Д. Брицкий, В. Попов / P. paxiana Kuntze, short-styled flowers. Botanical Garden of BIN RAS. 25.05.1996. D. Britski, V. Popov | 8 | 22.1–23.4 | 25.7–27.3 | 0.6–0.8 |
| Sect. Primula | ||||
| P. acaulis (L.) L., длинностолбиковая форма. Окрестности г. Туапсе. 27.05.1949. Т.А. Луговая / P. acaulis (L.) L., long-styled flowers. Near Tuapse town. 27.05.1949. Т.А. Lugovaya | 6 | 18.6–20.4 | 13.7–14.3 | 1.1–1.3 |
| P. amoena Bieb., длинностолбиковая форма. Западное Закавказье. 06.07.1978. С. Курганская, А. Андреев / P. amoena Bieb., long-styled flowers. West Transcaucasia. 06.07.1978. S. Kurganskaya, A. Andreev | 7 | 19.3–20.9 | 17.9–18.5 | 1.0–1.1 |
| P. cordifolia Rupr., короткостолбиковая форма. Тифлисская губ. 01.06.1914. В.Л. Козловский / P. cordifolia Rupr., short-styled flowers. Tiflis Guberniya. 01.06.1914. V.L. Kozlovskii | 8–9 | 28.1–29.2 | 28.7–30.1 | 0.7 |
| P. elatior (L.) Hill, короткостолбиковая форма. Ленинградская обл. 20.05.1972. А.О. Хааре / P. elatior (L.) Hill, short-styled flowers. Leningrad Region. 20.05.1972. A.O. Khaare | 6 | 25.5–26.7 | 22.8–23.8 | 0.7–0.9 |
| P. heterochroma Stapf, длинностолбиковая форма. Азербайджанская ССР. 07.05.1968. Н.Ю. Абдуллаева / P. heterochroma Stapf, long-styled flowers. Azerbaidjan SSR. 07.05.1968. N.Yu. Abdullaeva | 8 | 13.9–14.8 | 13.1–14.1 | 07.–0.9 |
| P. macrocalyx Bunge, длинностолбиковая форма. Окрестности г. Красноярска. 05.05.1908. В. Тугаринов / P. macrocalyx Bunge, long-styled flowers. Near Krasnoyarsk. 05.05.1908. V. Tugarinov | 6 | 15.8–16.8 | 15.3–17.4 | 0.8 |
| P. macrocalyx Bunge, короткостолбиковая форма. Ленинградская область / P. macrocalyx Bunge, short-styled flowers. Leningrad Region | 9 | 13.0–14.1 | 12.2–13.7 | 0.8 |
| P. nutans Georgi, длинностолбиковая форма. Тувинская АССР. Тандинский р-н. 28.06.1971. В. Ханминчун / P. nutans Georgi, long-styled flowers. Tuva ASSR. Tandinskiy distr. 28.06.1971. V. Khanminchun | 5 | 11.6–12.5 | 12.7–13.5 | 0.8 |
| P. nutans Georgi, короткостолбиковая форма. Алтай. 20.06.1963. Э. Гонтарь / P. nutans Georgi, short-styled flowers. Altai. 20.06.1963. E. Gontar | 5 | 14.8–15.8 | 15.2–16.4 | 0.8–0.9 |
| P. pallasii Lehm. длинностолбиковая форма. Западная Сибирь. Гора Шаман. 24.06.1897. Ю. Вагнер / P. pallasii Lehm., long-styled flowers. West Siberia. Shaman Mt. 24.06.1897. Yu. Vagner | 6 | 18.9–20.3 | 19.1–20.6 | 1.2–1.3 |
| P. pallasii Lehm., короткостолбиковая форма. Западная Сибирь. Гора Шаман. 24.06.1897. Ю. Вагнер / P. pallasii Lehm., short-styled flowers West Siberia. Shaman Mt. 24.06.1897. Yu. Vagner | 6 | 20.7–22.8 | 21.3–22.7 | 1.2–1.3 |
| P. pseudoelatior Kusn., длинностолбиковая форма. Западное Закавказье. 06.07.1973. Беатус / P. pseudoelatior Kusn., long-styled flowers. West Transcaucasia. 06.07.1973. Beatus | 6 | 19.5–20.8 | 19.5–20.7 | 1.2–1.3 |
| P. ruprechtii Kusn., длинностолбиковая форма. СО АССР. Орджоникидзевский р-н. 25.06. Н.Е. Варгина / P. ruprechtii Kusn., long-styled flowers. North–Osetinskaya ASSR. Ordzhonikidzevskiy distr. 25.06. N.E. Vargina | 6–7 | 12.9–14.1 | 13.9–15.3 | 0.7–0.8 |
| P. sibthorpii Hoffmg., длинностолбиковая форма. Грузия. 02.05.1912. В. Козловский / P. sibthorpii Hoffmg., long-styled flowers. Georgia. 02.05.1912. V. Kozlovskii | 7 | 15.1–16.2 | 15.9–17.6 | 1.1–1.2 |
| P. veris L., короткостолбиковая форма. Ленинградская обл. 09.06.1955. Григорьева / P. veris L., short-styled flowers. Leningrad Region. 09.06.1955. Grigor’eva | 7–8 | 25.5–27.1 | 24.2–25.6 | 0.7 |
| Sect. Juliae | ||||
| P. juliae Kusn., короткостолбиковая форма P. juliae Kusn., short-styled flowers |
7–8 | 16.6–17.7 | 17.6–18.8 | 0.6–0.7 |
| Sect. Sikkimensis | ||||
| P. pseudosikkimensis Forrest. Главный Ботанический сад. 07.1921. В. Комаров / P. pseudosikkimensis Forrest. Main Botanical Garden. 07.1921. V. Komarov | 7–8 | 13.5–14.5 | 14.3–15.7 | 0.6–0.7 |
Для гетеростильных видов сем. Primulaceae характерен диморфизм пыльцы, выраженный в размерах пыльцевых зерен. При этом короткостолбиковые растения продуцируют пыльцевые зерна заметно крупнее, по сравнению с длинностолбиковыми формами того же вида. Размеры пыльцы у разных форм гетеростильного вида примулы могут различаться в 1.5 раза (Morozowska, Idzikowska, 2004; Grigoryeva et al., 2018). Однако у изученных нами видов с многобороздной пыльцой эта разница в размерах пыльцы короткостолбиковых и длинностолбиковых форм не столь значительна. Так у P. nutans разница в размерах между самыми мелкими пыльцевыми зернами у длинностолбиковой формы и самыми крупными зернами у короткостолбиковой формы составляет всего 4.2 мкм, у P. pallasii – 4 мкм, у P. macrocalyx – 3.8 мкм.
Форма пыльцевых зерен эллипсоидальная (табл. I , 2) или почти сфероидальная (табл. I , 11). Очертания с полюса почти округлые, округлоугловатые или слабо многолопастные, с экватора почти округлые или эллиптические.
Таблица I . Морфология пыльцы видов рода Primula (СЭМ).
1–3 – P. heterochroma (длинностолбиковая форма); 4, 7 – P. macrocalyx (короткостолбиковая форма); 5, 6 – P. ruprechtii (длинностолбиковая форма); 8, 9 – P. amoena (длинностолбиковая форма); 10–2 – P. sibthorpii (длинностолбиковая форма). 1, 4, 8, 10 – общий вид пыльцевого зерна с полюса; 2, 5, 7, 11 – общий вид пыльцевого зерна с экватора; 3, 6, 9, 12 – скульптура поверхности.
Масштабные линейки, мкм: 1–12 – 1.
Plate I . Pollen morphology of Primula species (SEM).
1–3 – P. heterochroma (long-styled morph); 4, 7 –P. macrocalyx (short-styled morph); 5, 6 – P. ruprechtii (long-styled morph); 8, 9 – P. amoena (long-styled morph); 10–12 – P. sibthorpii (long-styled morph). 1, 4, 8, 10 – polar view of pollen grain; 2, 5, 7, 11 – equatorial view of pollen grain; 3, 6, 9, 12 – detail of ornamentation.
Scale bars, μm: 1–12 – 1.
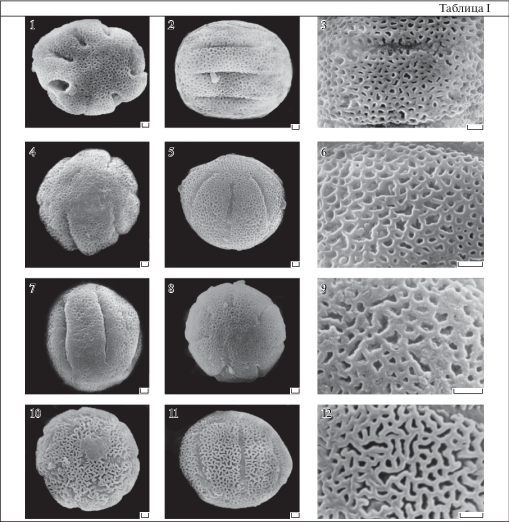
Борозды неширокие, недлинные, часто слегка извилистые, с заостренными или слегка закругленными концами и мембраной с редкими некрупными гранулами. Число борозд у разных видов варьирует от 6 до 9. Пыльцевые зерна с максимальным числом борозд (9) обнаружены у короткостолбиковых форм P. macrocalyx. В литературе имеются сведения о том, что для многобороздных пыльцевых зерен некоторых примул кроме разницы в размерах существуют различия в размерах скульптурных элементов и числе борозд (Morozowska, Idzikowska, 2004; Mačukanović Jocić et al., 2010). Считается, что более мелкие пыльцевые зерна длинностолбиковых цветков имеют меньшее число борозд по сравнению с пыльцой короткостолбиковых форм одного и того же вида. Так у P. macrocalyx в препаратах, полученных из длинностолбиковых цветков, преобладают 6-бороздные пыльцевые зерна, тогда как у короткостолбиковых цветков преобладает 9-бороздная пыльца. Следует отметить, что число борозд у многобороздной пыльцы примул часто сильно варьирует в пределах одного цветка от 5 до 8, поэтому трудно установить закономерность между числом борозд и формой цветка (Grigoryeva et al., 2018). У гетеростильных P. pallasii и P. nutans диморфизм пыльцы по числу борозд не обнаружен. Для обеих изученных форм цветков P. pallasii характерна 6-бороздная пыльца, а для P. nutans 5-бороздная пыльца.
Скульптура у пыльцы всех изученных видов сетчатая. Можно выделить 2 варианта сетчатой скульптуры:
1. Скульптура мелкосетчатая (табл. II , 3, 6, 9, 12).
Таблица II . Морфология пыльцы видов рода Primula (СЭМ).
1, 2 – P. elatior (короткостолбиковая форма); 3, 4 – P. juliae (короткостолбиковая форма); 5, 6 – P. veris (короткостолбиковая форма); 1, 5 – общий вид пыльцевого зерна с экватора; 3 – общий вид пыльцевого зерна с полюса. 2, 4, 6 – скульптура поверхности.
Масштабные линейки, мкм: 1, 3, 5 – 10; 2, 4, 6 – 1.
Plate II . Pollen morphology of Primula species (SEM).
1, 2 – P. elatior (short-styled morph); 3, 4 – P. juliae (short-styled morph); 5, 6 – P. veris (short-styled morph). 1, 5 – equatorial view of pollen grain; 3 – polar view of pollen grain; 2, 4, 6 – detail of ornamentation.
Scale bars, μm: 1, 3, 5 – 10; 2, 4, 6 – 1.
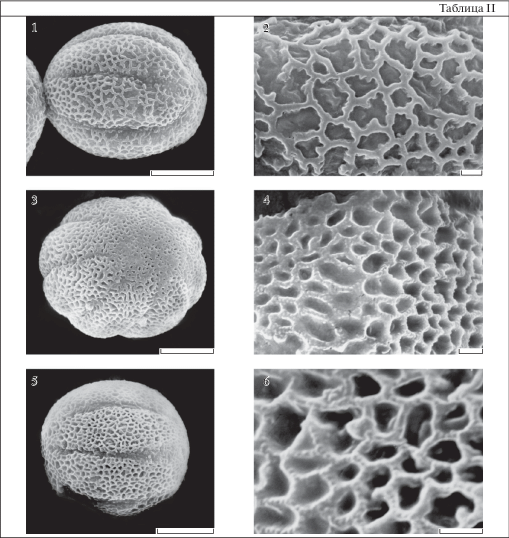
При исследовании на светооптическом уровне не просматривается. Данные СЭМ показывают, что просветы ячей меньше или равны расстоянию между ними. Просветы менее 0.5 мкм по максимальной оси, округло-угловатые или узкие вытянутые. Этот вариант встречается у P. sibthorpii (табл. I , 3), P. heterochroma (табл. I , 12) и у P. ruprechtii (табл. I , 6).
2. Скульптура крупносетчатая (табл. II , 2, 4, 6).
Ячеи довольно крупные, скульптура хорошо различима при исследовании с помощью СМ. Просветы ячей заметно больше ширины стенок. Исследование с помощью СЭМ показало, что очертания просветов ячей округлые, округло-угловатые или извитые, довольно крупные от 0.5 до 1.6 мкм по максимальной оси, с преобладанием крупных просветов. Размер ячей на мезокольпиумах несколько больше чем на полюсах.
Стенки ячей слегка изогнутые, неширокие, 0.3–0.5 мкм шириной.
В эту группу можно отнести пыльцу P. cordifolia, P. elatior, P. juliae, P. nutans и P. veris.
Самая крупная сетка характерна для пыльцевых зерен P. cordifolia.
У пыльцевых зерен P. cordifolia, P. elatior и P. nutans стенки ячей гладкие. На поверхности стенок ячей пыльцы P. juliae, P. paxiana и P. veris есть дополнительная орнаментация в виде мелких довольно частых гранул (табл. II , 4, 6). У пыльцевых зерен P. paxiana и P. veris гранулы настолько мелкие, что их возможно рассмотреть только при больших увеличениях СЭМ (не менее 20 000), тогда как обычно для изучения скульптуры пользуются увеличением ×10 000.
Следует отметить, что граница между выделенными группами условная, поскольку некоторые виды имеют переходный характер скульптуры.
Некоторые исследователи отмечают, что у гетеростильного вида P. vulgaris Huds. пыльцевые зерна с разных типов цветков заметно различаются размерами ячей сетчатой скульптуры. Так, ячеи на поверхности пыльцы короткостолбиковых форм заметно крупнее таковых у длинностолбиковых форм (Mačukanović Jocić et al., 2010). Нам удалось исследовать пыльцу трех гетеростильных примул (P. macrocalyx, P. nutans и P. pallasii) с обеих форм цветков. У них не было обнаружено существенных различий в скульптуре, размеры просветов и толщина стенок ячей у пыльцевых зерен длинностолбиковых и короткостолбиковых форм также не различаются. Видимо, диморфизм скульптур пыльцы у разных форм гетеростильных видов этого рода видоспецифичен и не является общей чертой всех гетеростильных представителей с многобороздной пыльцой. Как известно, для большинства гетеростильных видов рода Linum (Linaceae) характерен диморфизм пыльцы, выраженный в разных скульптурах у пыльцевых зерен длинно- и короткостолбиковых цветков. Однако среди гетеростильных льнов есть виды, у которых диморфизм пыльцы отсутствует и пыльцевые зерна короткостолбиковых и длинностолбиковых цветков имеют одинаковую скульптуру (Grigoryeva, 1989).
Ультраструктура спородермы многобороздной пыльцы нами изучена с помощью трансмиссионного электронного микроскопа у P. elatior и P. veris (табл. III , 7, 8). Для них характерна очень тонкая оболочка, около 1 мкм толщиной. Спородерма имеет сходное строение. На ультратонких срезах отчетливо видны экзина и интина. В экзине по электронной плотности различаются эктэкзина (более светлая) и эндэкзина (более темная). Эктэкзина тектатно-колумеллятного типа. Колумеллы (столбики) часто неровные, около 0.2 мкм высотой и около 0.1 мкм в диаметре, с расширенными головками. Столбики располагаются плотно друг к другу, расстояние между ними не более 0.1 мкм. Головки колумелл сливаются, образуя довольно толстый наружный слой тектум (покров), около 0.2–0.3 мкм толщиной. Тектум прерывистый, характерный для пыльцы с сетчатой скульптурой поверхности. Подстилающий слой очень тонкий, не более 0.04 мкм толщиной, прерывистый. Он различим только в случаях, когда срез проходит строго перпендикулярно оболочке. На “косых” срезах подстилающий слой не виден, что, по-видимому, и позволило сделать ошибочный вывод об отсутствии подстилающего слоя в оболочках многобороздной пыльцы среди видов Primula (Nowicke, Skvarla, 1977). Прерывистый подстилающий слой был обнаружен у многобороздной пыльцы P. sibirica из секции Farinosae (Grigoryeva et al., 2018). Расположенная под подстилающим слоем эндэкзина отличается от эктэкзины большей электронной плотностью, и на срезах выглядит более темной по сравнению с эктэкзиной. Представлена редкими, часто плохо различимыми ламеллами, плотно прилегающими к эктэкзине, и гомогенным прерывистым слоем, с максимальной толщиной не более 0.2 мкм. Интина мелко фибриллярная, довольно толстая, 0.4 мкм толщиной, без утолщений в области апертур. Мембрана борозд образована за счет интины и очень тонкого плохо различимого слоя спорополленина эктэкзины.
Таблица III . Вариабельность пыльцевых зерен Primula paxiana. Ультраструктура спородермы P. veris.
1–6 – P. paxiana. 7, 8 – P. veris. 1 – типичное пыльцевое зерно; 2–6 – атипичные пыльцевые зерна; 7, 8 – ультратонкие срезы оболочки; 7 – межапертурный участок; 8 – предапертурный участок.
Масштабные линейки, мкм: 1–8 – 1.
Plate III . Variability of pollen grains in Primula paxiana. Sporoderm ornamentation in P. veris.
1–6 – P. paxiana. 7, 8 – P. veris. 1 – typical pollen grain; 2–6 – atypical pollen grains; 7, 8 – ultra-thin sections of sporoderm; 7 – interaperture part; 8 – pre-aperture part.
Scale bars, μm: 1–8 – 1.
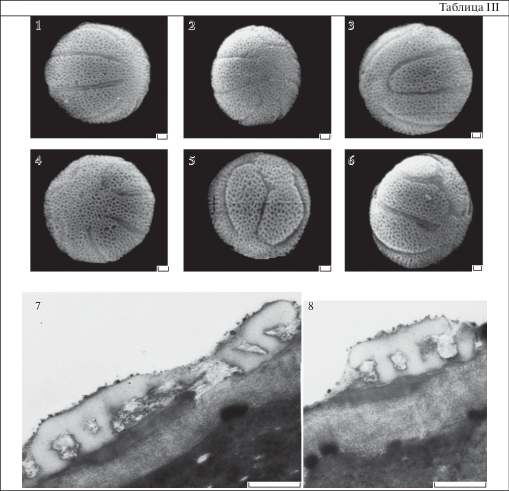
Внутривидовая изменчивость. Размеры и форма пыльцевого зерна внутри видов изменяются незначительно. Также консервативными признаками являются тип апертур (борозды у изученных видов) и скульптура, тогда как признаки борозд не стабильны. Даже в одном цветке встречаются пыльцевые зерна, которые различаются числом, расположением, характером слияния борозд. У типичных зерен все борозды б. м. одинаковой длины и не сливаются концами. У атипичных зерен длина борозд может варьировать. Встречаются пыльцевые зерна, у которых сливаются концы соседних борозд (табл. III , 3, 5), могут сливаться только 2 борозды или несколько. У некоторых зерен процесс слияния концов борозд лишь намечается (табл. III , 2, 4). У типичных зерен борозды ровные или слегка извилистые, тогда как у нетипичных зерен борозды имеют разнообразные изгибы и различаются длиной. У большинства изученных видов атипичные зерна встречаются редко и составляют менее 1% от общего числа в образце. Однако у P. macrocalyx, P. megaseifolia и P. paxiana доля пыльцевых зерен с разнообразными отклонениями по числу и расположению апертур может достигать 30–40%. Показано, что наличие нетипичных фертильных пыльцевых зерен и их количество характеризует отдельный образец и не является видоспецифичным признаком (Pozhidaev, 1993, 1995).
Причины появления в одном образце (а часто и в одном цветке) пыльцевых зерен с другим числом и расположением апертур по сравнению с основным типом пока не ясны, хотя в настоящее время ведутся работы по выявлению генов, влияющих на число апертур у разных мутантов Arabidopsis thaliana (L.) Heynh. (Dobritsa, Coerper, 2012; Prieu et al., 2016).
Для некоторых видов рода Viola (Violaceae) характерно наличие в одном цветке пыльцы с разным числом апертур. Было показано, что преобладание тех или других зерен у одного и того же вида зависит от места произрастания (в горах или на равнине). Пыльца фиалок с большим числом апертур прорастает быстрее, но при этом и быстрее теряет жизнеспособность. Пыльца с меньшим числом апертур прорастает медленнее, но дольше сохраняет способность к прорастанию (Dajoz et al, 1991, 1993; Mignot et al., 1994; Till-Bottraud et al., 1999). Вероятно, разное число апертур у пыльцевых зерен одного растения расширяет его репродуктивные возможности.
Таким образом, многобороздные пыльцевые зерна исследованных видов из секций Carolinella, Cortusoides, Primula, Juliae и Sikkimensis обладают сходными чертами, при этом виды из одной и той же секции могут различаться особенностями скульптуры, числом борозд, размерами пыльцевых зерен и толщиной экзины. Такие признаки, как форма пыльцевого зерна, размеры, число борозд и характер скульптуры могут быть важными дополнительными характеристиками отдельных видов Primula с многобороздной пыльцой, но не секций этого рода.
Список литературы
Al Wadi H., Richards A.J. 1992. Palynological variation in Primula L. subgenus Sphondylia (Duby) Rupr., and its relationship with Dionysia Fenzl. – New Phytol. 121(2): 303–310.
Anderberg A.A. 2004. Primulaceae. – In: The families and genera of vascular plant. Vol. 6. Springer. Berlin, Heidelberg. P. 313–319.
Anderberg A.A., El-Ghazaly G. 2000. Pollen morphology in Primula sect. Carolinella (Primulaceae) and its taxonomic implication. – Nordic J. Bot. 20(1): 5–14. https://doi.org/10.1111/j.1756-1051.2000.tb00726.x
Cronquist A. 1981. An integrated system of classification of flowering plants. New York. 1262 p.
Dajoz I., Till-Bottraud I., Gouyon P.H. 1991. Evolution of pollen morphology. – Science. 253(5015): 66–68. https://doi.org/10.1126/science.253.5015.66
Dajoz I., Till-Bottraud I., Gouyon P.H. 1993. Pollen aperture polymorphism and gametophyte performance in Viola diversifolia. – Evolution. 47(4): 1080–1093. https://doi.org/10.1111/j.1558-5646.1993.tb02137.x
Dobritsa A., Coerper D. 2012. The novel plant protein inaperturate pollen 1 marks distinct cellular domains and controls formation of apertures in the Arabidopsis pollen exine. – Plant Cell. 24(11): 4452–4464. https://doi.org/10.1105/tpc.112.101220
Erdtman G. 1952. Pollen morphology and taxonomy. Angiosperms. Stockholm. 539 p.
[Grigoryeva] Григорьева В.В. 1989. О диморфизме пыльцы некоторых видов рода Linum, Linaceae. – Бот. журн. 74(1): 65–72.
[Grigoryeva et al.] Григорьева В.В., Брицкий Д.А., Иванова А.Н., Коробков А.А. 2018. Морфология пыльцевых зерен некоторых видов секции Farinosae рода Primula (Primulaceae). – Бот. журн. 103(5): 645–654.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1967. Палинологическая терминология покрытосеменных растений. Л. 84 с.
[Kupriyanova, Aleshina] Куприянова Л.А., Алешина Л.А. 1972. Пыльца и споры растений флоры европейской части СССР. Т. 1. Л. 184 с.
Mačukanović Jocić M.P., Rančić D.V., Dajić Stevanović Z. P. 2010. Palynomorphological study of primrose (Primula vulgaris Huds.) grown in natural reserve obedska bara (Serbia). – J. Agricult. Sci. 55(3): 227–234. https://doi.org/10.2298/JAS1003227M
Mignot A., Hoss C., Dajoz I., Leuret C., Henry J.-P., Dreuillaux J.-M., Heberle-Bors E., Till-Bottraud I. 1994. Pollen aperture polymorphism in the Angiosperms: importance, possible causes and consequences. – Acta Bot. Gallica. 14(2): 109–122. https://doi.org/10.1080/12538078.1994.10515144
Morozowska M., Idzikowska K. 2004. Morphological differentiation of Primula veris L. pollen from natural and cultivated populations. – Acta Soc. Bot. Pol. 73(3): 229–232. https://doi.org/10.5586/asbp.2004.029
Nowicke J.W., Skvarla J.J. 1977. Pollen morphology and the relationship of the Plumbaginaceae, Polygonaceae and Primulaceae to the order Centrospermae. – Smithsonian Contrib. Bot. 37: 1–64.
Pozhidaev A.E. 1993. Polymorphism of pollen in the genus Acer (Aceraceae). Isomorphism of deviant forms of Angiosperm pollen. – Grana. 32: 79–85. https://doi.org/10.1080/00173139309429457
Pozhidaev A.E. 1995. Pollen morphology of the genus Aesculus (Hippocastanaceae). Patterns in the variety of morphological characteristics. – Grana. 34: 10–20. https://doi.org/10.1080/00173139509429028
Prieu C., Matamoro-Vidal A., Raquin C., Dobritsa A., Mercier R., Gouyon R., Albert B. 2016. Aperture number influences pollen survival in Arabidopsis mutants. – Am. J. Bot. 103(3): 452–459. https://doi.org/10.3732/ajb.1500301
Punt W., Weenen D.L.V., Van Dostrum W.A.P. 1974. The Northwest European Polen Flora. 3. Primulaceae. – Rev. Paleobot. Palynol. 17(3/4): 31–70.
Spanowsky W. 1962. Die Bedeutung der Pollenmorphologie für die Taxonomie der Primulaceae-Primuloideae. – Feddes Repert. Spec. Nov. Reg. Veg. 65(3): 149–214.
Till-Bottraud I., Vincent M., Dajoz I., Mignot A. 1999. Pollen aperture heteromorphism: Variation in pollen type proportions along altitudinal transects in Viola calcarata (Violaceae). – Comptes Rendus de l’Académie des Sciences Paris. Life Sciences. 322(7): 579–589.
Wendelbo P. 1961. Studies in Primulaceae III. On the genera related to Primula with special reference to their pollen morphology. – Arb. Univ. Bergen Mat. Nat. 19: 1–31.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал