

Ботанический журнал, 2019, T. 104, № 11-12, стр. 1712-1726
CТРУКТУРА ЦЕНОПОПУЛЯЦИЙ DRACOCEPHALUM RUYSCHIANA (LAMIACEAE) НА ЮГЕ СИБИРИ
Г. Р. Денисова 1, *, Н. И. Гордеева 1, **, Е. К. Комаревцева 1, А. А. Гусева 1
1 Центральный сибирский ботанический сад СО РАН
630090 г. Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: gulnoria@mail.ru
** E-mail: nataly.gordeeva@gmail.com
Поступила в редакцию 27.06.2019
После доработки 27.08.2019
Принята к публикации 10.09.2019
Аннотация
Изучена структура ценопопуляций симподиально нарастающего короткокорневищного многолетника Dracocephalum ruyschiana L. в разных эколого-фитоценотических условиях юга Сибири. Выявлены два типа онтогенетических спектров ценопопуляций: левосторонний и центрированный. Онтогенетическая структура определяется биологией вида (семенным способом размножения, быстрыми темпами развития в начале и в конце онтогенеза, продолжительностью зрелого генеративного состояния) и условиями произрастания (наличием свободного субстрата и ветоши, степенью задернованности ценоза, влажностью почвы и влиянием антропогенной нагрузки). Для половой структуры ценопопуляций вида в местообитаниях юга Сибири характерна невысокая доля женских особей (3.2%–6.3% от общего числа генеративных особей). Комплексная оценка состояния ценопопуляций показала стабильность популяционных параметров при значительной изменчивости морфологических признаков особей. В оптимальном состоянии находятся ценопопуляции на злаково-разнотравном лугу и в разреженном березняке, где отмечены максимальные и средние значения организменных и популяционных признаков.
В настоящее время популяционные исследования стали незаменимы при решении многих задач. Существенными диагностическими характеристиками популяций являются их онтогенетическая и половая структуры. Изучение онтогенетической структуры позволяет определить способы самоподдержания популяционной системы, дает представление об ее устойчивости и дальнейшей судьбе (Zaugol‘nova, Smirnova, 1990). Наличие полового полиморфизма у видов обеспечивает перекрестное опыление и аутбридинг особей в популяциях (Melikyan, 2000; Starshova, 2000). Исследование половой структуры видов, которая определяется количественным соотношением разных половых форм в популяции, способствует пониманию механизмов адаптации вида и успешности произрастания растений в фитоценозах (Dem‘yanova, Ponomarev, 1979).
Dracocephalum ruyschiana L. – короткокорневищный симподиально нарастающий травянистый многолетник, образующий рыхлый куст. Побеги возобновления удлиненные, формируются ежегодно из почек базальных частей побегов предыдущих лет. В конце вегетационного периода побеги отмирают до нижних коротких метамеров с чешуевидными листьями, которые со временем укореняются и встраиваются в состав гипогеогенного симподиально нарастающего корневища. Побеги возобновления ветвятся, образуя розеточные, верхнерозеточные или удлиненные вегетативные побеги обогащения. Придаточные корни формируются как в узлах, так и на междоузлиях корневища. Главный корень отмирает в зрелом генеративном состоянии. Соцветие – фрондозный, открытый колосовидный тирс, состоящий из супротивно расположенных многочленных дихазиев.
D. ruyschiana имеет голарктический ареал, протянувшийся от Скандинавии и Средней Европы до Приамурья (Shishkin, 1954). Северная граница пролегает по территориям Скандинавии (Швеции, Норвегии, Финляндии) и России (Архангельской и Тюменской областей, Красноярского края и Якутии). Проходя через Западную и Восточную Сибирь, вид едва заходит на территорию Дальнего Востока, где единственное местонахождение описано в бассейне реки Ту в 10 км к северу от деревни Беловеж (Probatova, Krestovskaya, 1995).
Эколого-ценотические условия обитания D. ruyschiana разнообразны. Изучение гербарных коллекций D. ruyschiana Центрального сибирского ботанического сада CO PAH (NS и NSK), Томского государственного университета (TK), Ботанического института им. В.Л. Комарова РАН (LE) и наблюдения за видом в природе на юге Сибири позволили объединить его местообитания в группы. Наиболее характерна для вида луговостепная группа. К этой группе относятся луговые и степные сообщества. Следующая по частоте встречаемости – это группа лесных формаций. Вид встречается в хвойных, березовых и смешанных лесах. На залежах, вырубках, неглубоких болотах, каменистых склонах и в зарослях вдоль ручьев D. ruyschiana встречается редко.
Для D. ruyschiana характерен половой полиморфизм: в популяции произрастают совместно гермафродитные особи с обоеполыми цветками и женские особи с пестичными цветками (гинодиэция), а также особи с обоеполыми и пестичными цветками на одном генеративном побеге (гиномоноэция) (Dem‘yanova, Ponomarev, 1979).
Онтогенетическая структура ценопопуляций (ЦП) этого вида в природе изучалась лишь на территории Якутии (Danilova et al., 2012): была исследована структура двух ЦП D. ruyschiana. Сведения о половой структуре ЦП также малочисленны и приведены в основном для европейской части России.
Цель работы: изучить онтогенетическую и половую структуру ценопопуляций D. ruyschiana и дать оценку их состояния в разных эколого-ценотических условиях юга Сибири.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили в Горном Алтае, в Хакасии и Туве с 2004 по 2018 г. Онтогенез и онтогенетическую структуру ЦП D. ruyschiana изучали с применением принятых в популяционной биологии растений методов и подходов, разработанных Т.А. Работновым (Rabotnov, 1950), А.А. Урановым (Uranov, 1975) и его учениками (Tsenopopulyatsii, 1976, 1988). Онтогенетическая структура ценопопуляций определялась как соотношение разных онтогенетических групп в ценопопуляции и изображалась в виде спектров. За счетную единицу в течение всего онтогенеза D. ruyschiana принималась особь. При характеристике популяционной структуры мы опирались на представления о характерном и базовом спектрах (Zaugol’nova, 1994). Для характеристики ЦП также использовалась экологическая плотность (Odum, 1986). Тип ЦП определялся по классификации А.А. Уранова и О.В. Смирновой (Uranov, Smirnova, 1969) и классификации “дельта–омега” (Δ–ω) Л.А. Животовского (Zhivotovskii, 2001). Состояние ЦП оценивалось по организменным и популяционным признакам (Zaugol’nova, 1994). В качестве организменных признаков нами выбраны число и высота генеративных побегов, потенциальная семенная продуктивность и фитомасса. В качестве популяционных – плотность особей на 1 м2, доля подроста (j–$v$), доля g1–g2, доля g3–s (в процентах), индекс восстановления ЦП (Zhukova, 1995). Потенциальная семенная продуктивность (ПСП) определялась с использованием методики И.В. Вайнагий (Vainagii, 1974). Диапазон каждого признака делили на пять классов с одинаковым объемом; потом каждому классу присваивался балл, наименьший балл соответствовал наименьшим показателям. Для сравнения брались средние значения каждого из организменных признаков у 25 зрелых генеративных особей. Оценку достоверности средних значений организменных признаков определяли по t-критерию Стьюдента при 5%-м уровне значимости (Zaitsev, 1991).
Онтогенетическая структура ценопопуляций D. ruyschiana изучалась в различных вариантах луговых (ЦП 1-8) и лесных сообществах (ЦП 9,10) Сибири (табл. 1).
Таблица 1.
Краткая характеристика фитоценозов Dracocephalum ruyschiana L. Table 1. Brief description of phytocenoses of Dracocephalum ruyschiana L.
| Номер ЦП/ Number of CP | Название сообщества (высота над уровнем моря) Community name (altitude) | Место произрастания сообщества/Locality and habitat of community | ОПП,%General projective cover,% | Доминирующие виды Dominant species |
|---|---|---|---|---|
| 1 | Овсецово-ковыльно-разнотравный остепненный луг (598 м над ур. м.)/ Grass-forb steppe meadow (598 m a. s. l.) | Хакасия. Долина р. Сон. Крутизна склона 5–6°. Максимальная высота травостоя 120 см/Khakassia. Son River valley. Slope 5–6°. Maximum herbage height 120 cm | 80 | Stipa pennata L., Helictotrichon desertorum (Less.) Nevski, Poa angustifolia L., Fragaria viridis Weston, Aconitum barbatum Pers., Iris ruthenica Ker-Gawl. |
| 2 | Стоповидноосоково-вейниково-разнотравный остепненный луг (1100 м над ур. м.)/Sedge-grass-forb steppe meadow (1100 m a. s. l.) | Горный Алтай, Шебалинский район, окр. пос. Верх-Кукуя. Крутизна склона 20‒25°. Максимальная высота травостоя 95 см/Mountain Altai, Shebalinsky Distr., environs of Verkh-Kukuya village. Slope 20–25°. Maximum herbage height 95 cm | 85 | Calamagrostis arundinacea (L.) Roth, Carex pediformis C.A. Mey., Hieracium umbellatum L., Trifolium pratense L., Ligularia glauca (L.) O. Hoffm., Saussurea controversa DC., Artemisia laciniata Willd. |
| 3 | Разнотравно-стоповидноосоковый остепненный луг (641 м над ур. м.)/ Forb-sedge steppe meadow (641 m a. s. l.) | Горный Алтай, Шебалинский район, Чергинский хребет, склоны по левому берегу р. Могута. Крутизна склона 5°. Максимальная высота травостоя 90 см/Mountain Altai, Shebalinsky Distr., Chergin Ridge, slopes on the left bank of the Moguta River. Slope 5°. Maximum herbage height 90 cm | 75 | Carex pediformis, Fragaria viridis, Achillea millefolium L., Aconitum barbatum, Elytrigia repens (L.) Nevski, Filipendula vulgaris Moench, Rubus saxatilis L. |
| 4 | Гераниево-василистниково-разнотравный луг (1410 м над ур. м.)/Forb meadow (1410 m a. s. l.) | Горный Алтай, Усть-Канский район, окр. с. Мендун–Саккон. Крутизна склона 30°. Максимальная высота травостоя 130 см/Mountain Altai, Ust-Kansky Distr., environs of Mendun-Saqcon village. Slope 30°. Maximum herbage height 130 cm | 60 | Thalictrum minus L., Potentilla chrysantha Trevir., Geranium pratense L., Dracocephalum ruyschia-na L., Crepis sibirica L., Phlomis tuberosa (L.) Moench |
| 5 | Овсецово-ежово-разнотравный луг/Grass-forb meadow | Горный Алтай. Усть-Коксинский район, oкр. с. Усть-Кокса. На опушке лиственничного леса. Максимальная высота травостоя 80 см. Сенокос/Mountain Altai. Ust-Koksinsky Distr., environs of Ust-Koksa village. Fringe of larch forest. Maximum herbage height 80 cm. Haymaking | 100 | Dactylis glomerata L., Helictotrichon pubescens (Huds.) Dumort., Rubus saxatilis, Ptarmica impatiens (L.) DC., Dracocephalum ruyschiana, Vicia cracca L., Vicia unijuga A. Br. |
| 6 | Мятликово-овсецово-разнотравный остепненный луг/Grass-forb steppe meadow | Горный Алтай. Усть-Канский район. Окр. с. Усть-Кан. На опушке лиственничного леса. Максимальная высота травостоя 70 см. Выпас/Mountain Altai. Ust-Kansky Distr., environs of Ust-Kan village. Fringe of larch forest Maximum herbage height 70 cm. Grazing | 90 | Helictotrichon pubescens, Poa sibirica Roshev., Galatella biflora (L.) Nees, Dracocephalum ruyschiana, Vicia cracca, Potentilla chrysantha, Galium verum L., Achillea millefolium |
| 7 | Ирисово-злаково-разнотравный луг (626 м над ур. м.)/Iris-grass-forb meadow (626 m a. s. l.) | Хакасия. Усть-Абаканский район. Долина р. Неня. На опушке соснового леса. Максимальная высота травостоя 120 см/Khakassia. Ust-Abakansky Distr., Nenya River valley. Fringe of pine forest. Maximum herbage height 120 cm | 95 | Iris ruthenica, Elytrigia repens, Poa angustifolia, Geranium pratense, Aconitum barbatum, Festuca pratensis Huds. |
| 8 | Стоповидноосоково-разнотравный луг (992 м над ур. м.)/Sedge-forb meadow (992 m a. s. l.) | Тува. Долина р. Арзак, надпойменная терраса, ложбинообразное понижение. Максимальная высота травостоя 110 см/Tyva. Arzak River valley, terrace above the floodplain, hollow-like slide. Maximum herbage height 110 cm | 90 | Carex pediformis, Phleum phleoides (L.) Karst., Fragaria viridis, Geranium pratense, Tanacetum vulgare L., Iris ruthenica |
| 9 | Лиственнично-березовый разнотравный лес (875 м над ур. м.)/Larch-birch forb forest (875 m a. s. l.) | Хакасия. Аскизкий район, окр. оз. Баланкуль. Максимальная высота травостоя 90 см/Khakassia. Askisky Distr., environs of Balankul Lake. Maximum herbage height 90 cm | 85 | Thalictrum minus, Betula pendula Roth, Larix sibirica Ledeb., Phlomis tuberosa, Aconitum barbatum, Viola uniflora L. |
| 10 | Разреженный березовый лес (489 м над ур. м.)/ Thinned birch forest (489 m a. s. l.) | Хакасия. Ширинский район, горный массив Чабалдан 5 км от пос. Джирим. Крутизна склона 3°. Максимальная высота травостоя 115 см/Khakassia. Shirinsky Distr., Chabaldan mountain range, 5 km from Jirim village. Slope 3°. Maximum herbage height 115 cm | 100 | Poa attenuata Trin., Betula pendula, Phleum phleoides, Thalictrum minus, Aconitum barbatum, Geranium pratense, Calamagrostis epigeios (L.) Roth |
Половая структура исследовалась в условиях овсецово-ежово-разнотравного луга (ЦП 5) и мятликово-овсецово-разнотравного остепненного луга (ЦП 6), расположенных в Республике Алтай, где учитывались все генеративные особи на трансектах площадью 20 м2 (табл. 1).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Онтогенез особей D. ruyschiana был исследован ранее в долине реки Сон в Хакасии на овсецово-ковыльно-разнотравном остепненном лугу (Denisova, Kovalyova, 2007). Установлено, что развитие особей D. ruyschiana в природных ЦП длится 13–18 лет, самоподдержание ЦП осуществляется исключительно семенным путем.
Проростки появляются весной и характеризуются формированием верхнерозеточного побега. К середине лета у особей удлиняются все междоузлия и побег становится удлиненным. Наступает ювенильное состояние. Первичный побег ювенильных особей состоит из гипокотиля и 5 удлиненных метамеров с настоящими листьями. Гипокотиль и эпикотиль побега втягиваются в почву, за счет контрактильной деятельности главного корня и становятся первым звеном корневища. Осенью побег отмирает до эпикотильного узла. На следующий год особи переходят в имматурное состояние, которое характеризуется сменой нарастания с моноподиального на симподиальное. Трогается в рост почка, заложенная в пазухе первого настоящего листа. Нереализованные почки становятся спящими. Побег возобновления состоит из одного укороченного метамера с чешуевидными листьями и 6–8 удлиненных метамеров с ассимилирующими листьями, из пазушных почек которых развертываются розеточные вегетативные побеги обогащения. Корневая система смешанная и представлена главным и 1–2 придаточными корнями, которые развиваются в семядольном узле. После вегетации побег отмирает до метамера с чешуевидными листьями. Длительность состояния один год. Первичный куст виргинильных растений формируется за счет раскрытия 2–3 почек, расположенных в пазухах чешуевидных и отмерших семядольных листьев. В структуре побегов возобновления виргинильных особей число укороченных метамеров с чешуевидными листьями увеличивается до двух, удлиненных метамеров с настоящими листьями – до 8–10. Меняется и тип побегов обогащения: с розеточного на верхнерозеточный. Главный и придаточные корни принимают участие в процессе погружения базальных частей растений. Виргинильное состояние длится 1–2 года. Куст молодых генеративных растений состоит из 1–2 генеративных и 1–2 вегетативных удлиненных моноциклических побегов. Строение вегетативных побегов остается неизменным. Базальная часть генеративных побегов представлена 2–3 метамерами с чешуевидными листьями. На генеративных побегах развертываются вегетативные побеги обогащения первого и второго порядков, на вегетативных – только первого порядка. Побеги обогащения первого порядка на генеративных побегах верхнерозеточные и удлиненные, на вегетативных – верхнерозеточные. Побеги обогащения второго порядка розеточные. Корневая система смешанная. Длительность молодого генеративного состояния не превышает двух-трех лет. В начале средневозрастного генеративного состояния у особей отмирает главный корень и образуется рыхлый куст, состоящий из 5–9 (иногда до 21) генеративных и 1–4 вегетативных удлиненных моноциклических побегов. Большое число побегов формируется за счет развертывания почек возобновления и спящих почек. Подземная сфера таких особей представлена разветвленным гипогеогенным симподиальным корневищем, нарастающим укороченными (2–3 метамера) базальными участками побегов. В конце зрелого генеративного состояния начинает отмирать корневище с проксимального конца. У особей в подземной сфере хорошо развита придаточная корневая система. Длительность зрелого генеративного состояния 5–7 лет. Рыхлый куст старых генеративных особей состоит из 1–3 генеративных и 1–3 вегетативных побегов. Субсенильные особи характеризуются развертыванием из спящих почек и почек возобновления 2–4 вегетативных удлиненных побегов имматурного типа. Корневище сильно разрушено. У сенильных особей побеги возобновления развиваются только из спящих почек на живом участке корневища, состоящем из одного, реже двух годичных приростов. Побеги обогащения у особей постгенеративного периода розеточные.
Все изученные ЦП D. ruyschiana, по классификации А.А. Уранова и О.В. Смирновой (Uranov, Smirnova, 1969), нормальные (рис. 1). Большая часть исследованных ЦП неполночленные: отсутствуют ювенильные (ЦП 1–5, 10), имматурные (10) или сенильные особи (ЦП 6, 7). Пополнение ЦП 1–5 и 10 молодыми особями затруднено, что подтверждается и долей виргинильных особей, которая не превышает в них 10%. В ЦП 5 и 6 отмечен резкий спад на особях старого генеративного состояния (1.66%; 7.69% соответственно) и отсутствуют сенильные особи. Неполночленность исследованных ЦП обусловлена такими факторами, как антропогенная нагрузка (выпас и сенокос), высокое проективное покрытие травостоя и задернованность ценоза, наличие ветоши, а также крутизна склонов (до 30°). Произрастая в таких условиях, часть молодых особей D. ruyschiana, не имея развитой корневой системы, погибает.
Большое значение в появлении подроста в ЦП D. ruyschiana имеет семенная продуктивность. Анализ ПСП в разных ЦП установил довольно широкий диапазон ее изменчивости от 293.3 до 937.81 эремов на особь (табл. 2) и прямую зависимость ПСП от размеров соцветия.
Таблица 2.
Организменные и популяционные признаки Dracocephalum ruyschiana L. Table 2. Characteristics of individuals and populations of Dracocephalum ruyschiana L.
| Признак Character |
Ценопопуляция Coenopopulation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Организменные признаки особей g2 Characters of organisms g2 | ||||||||||
| Число генеративных побегов Number of generative shoots |
9.55 ± 1.4 | 6.97 ± 1.03 | 5.35 ± 0.86 | 8.27 ± 1.7 | 6.3 ± 0.69 | 4.95 ± 0.35 | 8.07 ± 0.62 | 7.42 ± 0.86 | 5.73 ± 0.37 | 8.25 ± 2.1 |
| Высота генеративных побегов, см Height of generative shoots, cm |
38.6 ± 1.1 | 44.02 ± 2.1 | 38.29 ± 1.34 | 51.2 ± 2.01 | 39.45 ± 0.63 | 34.65 ± 0.64 | 47.26 ± 1.08 | 41.12 ± 1.26 | 47.57 ± 2.02 | 50.43 ± 1.9 |
| Потенциальная семенная продуктивность, шт./особь Potential seed productivity, pcs/ plant |
937.81 ± 78.19 |
536.69 ± 43.21 |
468.66 ± 51.95 |
548.3 ± 48.79 |
504 ± 31.63 |
515.79 ± 29.9 |
522.9 ± 42.77 |
707 ± 60.33 |
293.3 ± 22.92 |
650.1 ± 68.31 |
| Надземная фитомасса, г Phytomass, g |
9.43 ± 1.57 | 6.16 ± 0.87 | 3.9 ± 0.37 | 11.53 ± 2.1 | 5.38 ± 0.5 | 4.53 ± 0.42 | 10.43 ± 1.99 | 8.09 ± 1.29 | 5.4 ± 0.59 | 15.78 ± 3.49 |
| Популяционные признаки Characters of population | ||||||||||
| Доля подроста (j–v),% Share of undergrowth,% |
8.34 | 5.08 | 7.95 | 4.06 | 13.33 | 11.53 | 43.51 | 82.8 | 19.32 | 7.09 |
| Доля g1–g2,% Share of g1–g2,% |
64.16 | 46.83 | 63.64 | 44.94 | 66.66 | 75.63 | 24.07 | 11.17 | 58.68 | 76.1 |
| Доля g3–s,% Share of g3–s,% |
27.5 | 48.09 | 28.41 | 51 | 20.01 | 12.84 | 32.42 | 6.03 | 22 | 16.81 |
| Плотность, экз./м2 Density, individuals/m2 |
2 | 6.7 | 1 | 3.95 | 6 | 7.8 | 4.32 | 8.64 | 1.2 | 3.4 |
| Индекс восстановления (Iвост) Index of recovery (Iвост) |
0.09 | 0.06 | 0.09 | 0.05 | 0.14 | 0.12 | 0.54 | 0.85 | 0.20 | 0.08 |
По данным P. Milberg и A. Bertilsson (1997), на территории Швеции большая часть эремов у особей D. ruyschiana образуется, в основном, в результате перекрестного опыления, осуществляемого шмелями. Авторами установлено, что семенная продуктивность D. ruyschiana зависит от степени изолированности ЦП, видового богатства сообщества (ценозы с богатым видовым разнообразием больше привлекают шмелей для опыления, чем маловидовые сообщества) и от числа цветков в соцветии, но не зависит ни от размеров особей, ни от длины соцветия.
Одним из механизмов адаптации D. ruyschiana является растянутый период прорастания семян (эремов). По неопубликованным данным И.Н. Гуськовой, в результате исследования лабораторной всхожести семян D. ruyschiana из природных местообитаний Сибири было установлено, что вид характеризуется очень растянутым периодом прорастания: через 30 дней всхожесть составляла 1.5%–2.0% от общего числа семян, а через 360 дней колебалась от 29.0% до 46.5%. Длительный период прорастания семян D. ruyschiana можно рассматривать как приспособление к условиям произрастания в сомкнутых ценозах с высоким уровнем видовой конкуренции. Исходя из данных P. Milberg и A. Bertilsson (1997) и И.Н. Гуськовой, несмотря на высокую ПСП D. ruyschiana, вид имеет слабое и нерегулярное семенное размножение, что также, помимо неблагоприятных условий среды, отрицательно влияет на число прегенеративных растений в ЦП.
Характерный онтогенетический спектр ЦП D. ruyschiana центрированный, он определяется биологией вида: семенным способом размножения, быстрыми темпами развития особи в начале (прегенеративный период длится 3–4 года) и в конце онтогенеза (длительность g3–s – 3–4 года), продолжительностью зрелого генеративного состояния (5–7 лет). Базовый спектр, полученный по нашим данным, также центрированный. Анализ онтогенетической структуры изученных ЦП D. ruyschiana показал, что по доминированию разных онтогенетических групп спектры разделились на два типа: центрированный и левосторонний (рис. 1). Преобладающий тип спектра ЦП – центрированный (ЦП 1-7, 9, 10) и только ЦП 8 имеет левосторонний спектр.
Рис. 1.
Онтогенетические спектры ценопопуляций Dracocephalum ruyschiana L.
По горизонтали – онтогенетические состояния; по вертикали – количество особей на единицу площади, %.
Онтогенетические состояния: j – ювенильное, im – имматурное, v – виргинильное, g1 – молодое генеративное, g2 – зрелое генеративное, g3 – старое генеративное, ss – субсенильное, s – сенильное. ЦП – ценопопуляция.
Fig. 1. Ontogenetic spectra of coenopopulations of Dracocephalum ruyschiana L.
X-axis – ontogenetic states, Y-axis – percentage.
Ontogenetic states: j – juvenile, im – immature, v – virginal, g1 – young generative, g2 – mature generative, g3 – old generative, ss – subsenile, s – senile. CP – coenopopulation.
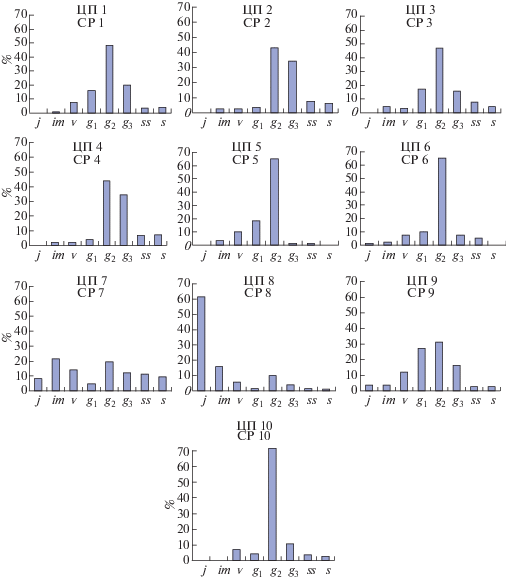
В большинстве ценопопуляций, где формируется центрированный тип спектра, отмечено накопление зрелых генеративных особей за счет их длительного существования в этом состоянии. Такой тип спектра также связан со слабым семенным размножением и быстрым переходом особей из одного онтогенетического состояния в другое в прегенеративном периоде. Абсолютный максимум в спектрах ЦП 1–6, 9, 10 приходится на зрелые генеративные растения (31.2–71.68%). По классификации ∆–ω (Zhivotovskii, 2001) ЦП 1, 3, 5, 6, 9, 10 относятся к зрелым (∆ = 0.41–0.51; ω = 0.73–0.88), ЦП 2, 4 – к стареющим (∆ = 0.61; ω = 0.78–0.79).
Среди изученных нами ЦП по онтогенетическому спектру выделяются две. В спектре ЦП 7 на ирисово-злаково-разнотравном лугу наблюдаются два пика: первый связан с накоплением имматурных (21.29%), второй – зрелых генеративных особей (19.44%). Эта ЦП находится в сукцессивном состоянии. В ней кривая распределения особей по онтогенетическим состояниям перестраивается волнообразно, с перемещением максимума в сторону зрелого генеративного состояния, поэтому данный спектр мы рассматриваем как временный вариант центрированного спектра. По классификации ∆–ω Л.А. Животовского (Zhivotovskii, 2001), ЦП 7 – переходная (∆ = 0.41; ω = = 0.49). На стоповидноосоково-разнотравном лугу (ЦП 8) также складываются благоприятные условия для прорастания семян и накопления молодых особей в ценопопуляции (j + im = 77.24%). В ней больше половины особей находятся в ювенильном состоянии (61.44%). По классификации ∆-ω ЦП 8 молодая (∆ = 0.13; ω = 0.24).
По нашим данным, плотность ЦП D. ruyschiana варьирует от 1 до 8.64 экз./м2. Факторами, влияющими на плотность особей, являются ОПП фитоценоза, его задернованность, наличие ветоши и крутизна склона. Так, в ЦП 3, исследованной на задернованном склоне, где ОПП травостоя 75%, отмечена минимальная плотность особей вида. Дерновины и ветошь Carex pediformis не дают возможности прорасти семенам, и большинство молодых особей появляется на участках, свободных от них. Наибольшая плотность (8.64 экз./м2) установлена на стоповидноосоково-разнотравном лугу (ЦП 8). Такая плотность особей в ценопопуляции обеспечивается высокой долей подроста, успешно развивающегося в условиях повышенной влажности.
Дополнительным фактором адаптации растений D. ruyschiana к условиям окружающей среды является наличие у вида гинодиэции, которая влияет на перекрестное опыление и аутбридинг особей в популяциях. Установлено, что в природных популяциях D. ruyschiana лесостепного Зауралья (Троицкий лесостепной заповедник, Челябинская область) встречаются крупноцветковые гермафродитные особи вида и мелкоцветковые женские особи. При этом встречаемость женских особей изменялась по годам от 10.4% до 23.4%, в среднем 14.82% (Dem‘yanova, Ponomarev, 1979). В лесостепной зоне Центрально-Черноземного заповедника (Курская область), где вид сравнительно обилен по опушкам разреженных лесов и на остепненных лугах, также указывалось на изменение численности женских особей по годам от 9.02% до 21.01%, что в среднем составляет 16.24%, (Dem‘yanova, 2013). В Курской области, кроме того, были обнаружены гиномоноэцичные особи (до 8% от их общего числа) и переходные особи с неполной стерильностью андроцея (до 3%). Для лесостепной зоны Новосибирской области в условиях интродукции указывалось, что на одной особи часто вместе формируются обоеполые и пестичные цветки в различном соотношении (гиномоноэция) (Gus‘kova, 1987).
Изучение половой структуры ЦП 5 и ЦП 6 в луговых сообществах Горного Алтая выявило только две половые формы: гермафродитную и женскую. Установлено, что встречаемость женских особей в этих ЦП низкая и составляет от 3.2% до 6.3% от общего числа генеративных особей (табл. 3). Проективное покрытие D. ruyschiana в исследованных сообществах составляет около 5% от ОПП травостоя, что свидетельствует о достаточно благоприятных условиях произрастания растений вида. Эти ЦП имеют центрированный спектр со значительным преобладанием генеративных особей (85.0% и 83.3% соответственно) (рис. 1). Наличие большого числа генеративных особей позволяет обеспечить хорошее семенное размножение растений и способствовать увеличению числа женских особей, однако, доля женских особей в половой структуре популяций невелика.
Таблица 3.
Половая структура ценопопуляций Dracocephalum ruyschiana L. разнотравно-злаковых лугов Горного Алтая Table 3. Sexual structure of coenopopulations of Dracocephalum ruyschiana L. in meadows of the Mountain Altai
| Ценопопуляции Coenopopulation |
Гермафродитные особи, экз. Hermaphroditic individuals, pcs |
Женские особи, экз. Females, pcs |
Всего, экз. All individuals, pcs |
|---|---|---|---|
| Ценопопуляция 5 Coenopopulation 5 |
74 (93.7%) | 5 (6.3%) | 79 (100%) |
| Ценопопуляция 6 Coenopopulation 6 |
90 (96.8%) | 3 (3.2%) | 93 (100%) |
Таким образом, учитывая результаты наших исследований и литературные данные, можно предположить, что относительно низкая встречаемость женских особей в популяциях является видоспецифическим признаком D. ruyschiana.
Оценка состояния ЦП представлена в табл. 2 и 4.
Таблица 4.
Оценка состояния ценопопуляций Dracocephalum ruyschiana L. (в баллах) Table 4. Assessment of Dracocephalum ruyschiana L. coenopopulations status (points)
| Признак/Character | Ценопопуляции/Coenopopulations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Баллы организменных признаков/Points of individual characters | ||||||||||
| Число генеративных побегов/Number of generative shoots | 5 | 3 | 2 | 4 | 2 | 1 | 4 | 4 | 2 | 4 |
| Высота генеративных побегов/Height of generative shoots | 2 | 3 | 2 | 5 | 3 | 1 | 5 | 3 | 5 | 5 |
| Потенциальная семенная продуктивность/Potential seed productivity | 5 | 3 | 3 | 3 | 3 | 3 | 3 | 4 | 1 | 4 |
| Надземная фитомасса/Phytomass of mature generative individual | 3 | 2 | 1 | 4 | 2 | 2 | 4 | 3 | 2 | 5 |
| Сумма баллов/Total points | 15 | 11 | 8 | 16 | 10 | 7 | 16 | 14 | 10 | 18 |
| Баллы популяционных признаков/Points of population characters | ||||||||||
| Доля подроста/Share of undergrowth | 2 | 2 | 2 | 1 | 2 | 2 | 3 | 5 | 2 | 2 |
| Доля g1–g2/Share of g1–g2 | 5 | 4 | 5 | 4 | 5 | 5 | 2 | 1 | 4 | 5 |
| Доля g3–s/Share of g3–s | 3 | 5 | 4 | 5 | 3 | 2 | 4 | 1 | 3 | 2 |
| Плотность/Density | 2 | 4 | 1 | 3 | 4 | 5 | 3 | 5 | 2 | 3 |
| Индекс восстановления/Index of recovery | 2 | 2 | 2 | 1 | 2 | 2 | 4 | 5 | 2 | 2 |
| Сумма баллов/Total points | 14 | 17 | 14 | 14 | 16 | 16 | 16 | 17 | 13 | 14 |
Нами установлена зависимость высоты генеративного побега особи от высоты травостоя ценоза. Так, в ЦП 4, 7, 9, 10, где высота травостоя в ценозе достигает 110–130 см, отмечен максимальный балл высоты генеративных побегов D. ruyschiana (47.26–51.2 см). В ЦП 6, расположенной на мятликово-овсецово-разнотравном остепненном лугу с высотой травостоя до 70 см, высота D. ruyschiana минимальная – 34.65 см (1 балл). В остальных ЦП этот параметр имеет среднюю балльную оценку – 2 или 3 балла (38.29–44.02 см).
Надземная фитомасса является одним из самых информативных показателей жизненности растения, которая отражает условия произрастания растений. Ее величина определяется числом генеративных побегов в кусте, а также высотой этих побегов (табл. 4). Так, в ЦП 10 генеративный куст имел 8.25 генеративных побегов на особь, до 50.43 см дл., что обусловливает высокую величину фитомассы (15.78 г). Подобное сочетание параметров генеративной особи отмечается в ЦП 4 и 7. Небольшая высота побегов в ЦП 1 и 8 (38.6 см и 41.12 см соответственно), несмотря на большое число побегов (7.42–9.55) в кусте, отрицательно сказывается на величине фитомассы, также как развитые побеги при небольшом их числе в кусте не способствуют накоплению фитомассы особи (ЦП 9) (табл. 2, табл. 4).
В большинстве изученных ЦП отмечены довольно невысокие значения потенциальной семенной продуктивности (ПСП) у генеративных особей вида от 468.66 до 707 шт./особь. Исключение составляют ЦП 1, где у особей отмечается максимальная ПСП – 937.81 шт./особь и ЦП 9, где этот показатель минимален – 293.3 шт./особь. Высокая ПСП в ЦП 1 обусловлена высокой долей генеративных побегов, низкая ПСП связана с небольшим числом цветков на побеге цветущей особи в ЦП 9.
Анализ организменных признаков показал, что в половине исследованных ЦП (1, 4, 7, 8, 10) генеративные особи достигают высокого и среднего развития. В ЦП 2, 3, 5, 6 и 9 произрастают менее развитые особи. Наиболее мощные экземпляры обнаружены в ЦП 10. Среднего уровня развития достигают особи в ЦП 1, 4, 7, 8.
В ЦП 10 в условиях слабого задернения произрастают кусты, состоящие из значительного числа цветущих побегов, максимальной длины, что обеспечивает им высокую фитомассу. Значения ПСП у особей в ЦП 10 немного выше, по сравнению с особями большинства других ЦП. Приближаются к ним по своему развитию и растения ЦП 4 и 7. Особи этих ЦП характеризуются меньшими значениями ПСП и фитомассы. Наименьшие значения параметров особей ЦП 3 и 6 обусловлены сильным задернением и выпасом.
Анализ значений популяционных признаков также позволил выявить несколько закономерностей. Большинство ценопопуляций (ЦП 1–6, 9, 10) характеризуются низкой долей прегенеративных особей от 4.06 до 19.32% при высокой численности молодых и зрелых генеративных растений – 44.94–76.1%. Лимитирующими факторами в появлении подроста являются задернение ценоза, наличие ветоши и высокая крутизна склона (30°). Малочисленность подроста способствует уменьшению индекса восстановления ЦП – 0.05–0.20. Исключение составляет ЦП 8, где при незначительном участии генеративных особей 11.17% отмечаются максимальные значения доли подроста 82.8%, что увеличивает индекс восстановления до 0.85. Доля старых генеративных и постгенеративных растений в составе ценопопуляций подвержена большим колебаниям численности, что связано с уязвимостью особей в этот период. Так, антропогенная нагрузка (ЦП 5,6) ускоряет темпы развития особей старых растений, что способствует сокращению их онтогенеза и уменьшению доли g3–s (табл. 2, 4).
Плотность ЦП – параметр, достаточно изменчивый и определяется как долей подроста, так и долей генеративных растений в ее составе. Для большинства ЦП оценка этого показателя колеблется от 2 до 4 баллов (1.2–6.7 экз./м2). Исключение составляют ЦП 8 на стоповидноосоково-разнотравном лугу в Туве, в которой установлена максимальная плотность особей вида 8.64 экз./м2 (5 баллов) за счет высокой доли прегенеративных растений (82.8%), а также в ЦП 6 на мятликово-овсецово-разнотравном остепненном лугу в Горном Алтае – 7.8 экз./м2 при высокой численности молодых и зрелых генеративных растений (75.63%). Наименьшая плотность особей 1 экз./м2 (1 балл) оказалась в ЦП 3 на овсецово-ковыльно-разнотравном остепненном лугу из-за дерновин и ветоши Carex pediformis.
Таким образом, по популяционным параметрам все изученные ЦП имеют хороший и средний уровень развития. Хорошо развиты ЦП 2, 5–8, к среднеразвитым относятся все остальные ценопопуляции (ЦП 1, 3, 4, 9, 10). Ценопопуляции с низкими значениями популяционных параметров не выявлены.
Комплексная оценка состояния изученных ЦП D. ruyschiana показала несовпадение максимальных показателей организменного уровня (ЦП 10, 18 баллов) с высокими значениями популяционных параметров (ЦП 2 и 8, 17 баллов). Для ЦП 1, 4, 7, 8 и 10 характерно сочетание средних или максимальных оценок популяционных и организменных признаков. Это свидетельствует о приближении этих ценопопуляций к оптимальному состоянию. Они расположены на разнотравных лугах и в разреженном березовом лесу (ЦП 10) с хорошо развитым травостоем высотой 110–130 см. Однако сопоставление балловых оценок всех признаков и онтогенетической структуры показало, что ЦП 4, 7, 8, имеющие высокие и средние значения популяционных и организменных признаков, не находятся в оптимальном состоянии, т.к. в ЦП 4 преобладают старые особи (g3–s – 51%), что приведет ее к быстрому старению, в ЦП 8 вид в основном представлен прегенеративной фракцией (j–v = 82.8%), а ЦП 7 – сукцесивная.
Остальные ЦП, произрастающие на остепненных лугах и в лиственнично-березовом разнотравном лесу с высоким ОПП и невысоким травостоем (70–90 см), характеризуются высоким (ЦП 2, 5, 6) или средним (ЦП 3, 9) уровнем развития популяционных параметров при низких значениях организменного уровня в этих ценопопуляциях, относящихся к зрелым (ЦП 3, 5, 6, 9) и стареющим (ЦП 2), удержание видом территории происходит за счет преобладания в составе ЦП генеративных особей в зрелом состоянии, характеризующимся наибольшей продолжительностью.
ЗАКЛЮЧЕНИЕ
Особи D. ruyschiana, произрастающие на юге Сибири, формируют короткокорневищную биоморфу, растения развиваются по симподиальной длиннопобеговой модели побегообразования. Онтогенез простой. Размножение вида осуществляется исключительно семенным путем. По классификации А.А. Уранова и О.В. Смирновой (Uranov, Smirnova, 1969), большинство исследованных ценопопуляций нормальные, неполночленные (ЦП 1–6, 10) и полночленные (ЦП 7–9). Неполночленность спектров связана с отсутствием ювенильных, имматурных и сенильных особей. Выявлены два типа онтогенетических спектров: центрированный и левосторонний. Центрированный спектр соответствует характерному и базовому спектрам и формируется в большинстве ЦП (1–7, 9, 10), левосторонний спектр – только в увлажненных местообитаниях (ЦП 8). По классификации дельта-омега Л.А. Животовского (Zhivotovskii, 2001) исследованные ЦП разделились на четыре группы: молодые (ЦП 8), переходные (ЦП 7), зрелые (ЦП 1, 3, 5, 6, 9, 10) и стареющие (ЦП 2, 4). Характер онтогенетической структуры и состояние ЦП определяются конкретными экологическими условиями (крутизна склона, ветошь, влажность субстрата, задернованность) и влиянием антропогенного фактора. Наличие свободного субстрата, влажность почвы, отсутствие ветоши и антропогенной нагрузки способствуют появлению молодых растений и увеличению численности особей в ЦП. Исследование половой структуры ценопопуляций D. ruyshiana в луговых сообществах юга Сибири показало низкий процент женских половых форм (3.2%-6.3%) от общего числа генеративных особей, что позволяет рассматривать довольно низкую встречаемость женских особей в популяциях как видоспецифический признак этого вида.
Комплексная оценка состояния ценопопуляций показала стабильность популяционных параметров при значительной изменчивости морфологических признаков особей. В оптимальном состоянии находятся ценопопуляции на злаково-разнотравном лугу (ЦП 1) и в разреженном березняке (ЦП 10). Ценопопуляции в разнотравных сообществах (ЦП 4, 7, 8) приближаются к этому состоянию, но значительная доля в их составе прегенеративных (ЦП 8) или старых (ЦП 4) особей, а также сукцессивное состояние ценопопуляции (ЦП 7) уменьшают их устойчивость. Это не позволяет характеризовать состояние этих ценопопуляций как оптимальное. В остальных ценопопуляциях, произрастающих на остепненных лугах (ЦП 2, 5, 6) и в лиственнично-березовом разнотравном лесу (ЦП 9), отмечается несовпадение организменного и популяционного оптимума. В этих ценопопуляциях, относящихся к зрелым (ЦП 3, 5, 6, 9) и стареющим (ЦП 2), удержание территории видом происходит за счет преобладания в составе средневозрастных генеративных особей с невысокими значениями морфологических параметров.
Список литературы
[Danilova et al.] Данилова Н.С., Иванова Н.С., Борисова С.З. 2012. Эколого-биологические особенности и структура ценопопуляций Dracocephalum ruyschiana в Центральной Якутии. – Наука и образование. 3: 33–38.
[Dem‘yanova] Демьянова Е.И. 2013. Половой полиморфизм некоторых степных растений Центрально-черноземного заповедника. – Вестн. Пермск. ун-та. Сер. Биология. 2: 11–18.
[Dem‘yanova, Ponomarev] Демьянова Е.И., Пономарев А.Н. 1979. Половая структура природных популяций гинодиэцичных и двудомных растений лесостепи Зауралья. – Бот. журн. 64(7): 1017–1024.
[Denisova, Kovalyova] Денисова Г.Р., Ковалева Ю.М. 2007. Онтогенез змееголовника Руйша (Dracocephalum ruyschiana L.). – В кн.: Онтогенетический атлас растений. Т. 5. Йошкар-Ола. С. 175–178.
[Gus‘kova] Гуськова И.Н. 1987. Гинодиэция видов семейства Яснотковых в Сибири. – В сб.: Проблемы размножения цветковых: Тез. докл. совещания по цветению, опылению и семенной продуктивности растений. Пермь. С. 80.
[Melikyan] Меликян А.П. 2000. Половой полиморфизм. – В кн.: Эмбриология цветковых растений. Терминология и концепции. Системы репродукции. Т. 3. СПб. С. 73–75.
Milberg P., Bertilsson A. 1997. What determines seed set in Dracocephalum ryuschiana L. an endangered grassland plant? – Flora. 192: 361–367.
[Odum] Одум Ю. 1986. Экология. Т. 2. М. 376 с.
[Probatova, Krestovskaya] Пробатова Н.С., Крестовская Т.В. 1995. Яснотковые – Lamiaceae. – В кн.: Сосудистые растения советского Дальнего Востока. Т. 7. СПб. С. 284–380.
[Rabotnov] Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Тр. БИН АН СССР. Сер. Геоботаника. 3(6): 179–196.
[Shishkin] Шишкин Б.К. 1954. Род Змееголовник – Dracocephalum L. – В кн.: Флора СССР. Т. 20. М.; Л. С. 439–474.
[Starshova] Старшова Н.П. 2000. Популяционные аспекты детерминации пола. – В кн.: Эмбриология цветковых растений. Терминология и концепции. Системы репродукции. Т. 3. СПб. С. 88–93.
[Tsenopopulyatsii] Ценопопуляции растений: (основные понятия и структура). 1976. М. 217 с.
[Tsenopopulyatsii] Ценопопуляции растений: (очерки популяционной биологии). 1988. М. 182 с.
[Uranov] Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Биол. науки. 2: 7–34.
[Uranov, Smirnova] Уранов А.А., Смирнова О.В. 1969. Классификация и основные черты развития популяций многолетних растений. – Бюл. МОИП. Отд. биол. 74(2): 119–134.
[Vainagii] Вайнагий И.В. 1974. О методике изучения семенной продуктивности растений. – Бот. журн. 59 (6): 826–831.
[Zaitsev] Зайцев Г.Н. 1991. Математический анализ биологических данных. М. 184 с.
[Zaugol’nova] Заугольнова Л.Б. 1994. Структура популяций семенных растений и проблемы их мониторинга: Автореф. дис. … докт. биол. наук. СПб. 70 с.
[Zaugol’nova, Smirnova] Заугольнова Л.Б., Смирнова О.В. 1990. Организация популяций растений в зависимости от условий среды и антропогенных воздействий. – В сб.: Структурно-функциональная организация и устойчивость биологических систем. Днепропетровск. С. 6–17.
[Zhivotovskii] Животовский Л.А. 2001. Онтогенетическое состояние, эффективная плотность и классификация популяций. – Экология. 1: 3–7.
[Zhukova] Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 223 с.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал