

Ботанический журнал, 2019, T. 104, № 3, стр. 377-390
ЖИЗНЕННЫЕ ФОРМЫ НЕКОТОРЫХ ВИДОВ РОДА THYMUS (LAMIACEAE): АРХИТЕКТУРНЫЙ ПОДХОД
В. А. Черемушкина 1, Е. Б. Таловская 1, *
1 Центральный сибирский ботанический сад Сибирского отделения РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: kolegova_e@mail.ru
Поступила в редакцию 20.12.2018
После доработки 09.02.2019
Принята к публикации 12.02.2019
Аннотация
С применением архитектурного подхода изучена структура особей модельных видов Thymus extremus, T. indigirkensis, T. brevipetiolatus и T. pavlovii, характеризующихся разными жизненными формами. В структуре побеговой системы видов рода Thymus выделено 4 архитектурные единицы, которые различаются направлением роста и способом нарастания (ортотропная моно-базисимподиальная, ортотропная моно-акросимподиальная, восходящая моно-базисимподиальная, плагиотропная моно-акросимподиальная). Каждая архитектурная единица состоит из главной (или материнской n-го порядка) составной скелетной оси, составных скелетных осей 1-го (n + 1) порядка, одиночных побегов формирования 1-го порядка, побегов ветвления и обогащения. Показано, что архитектоника особей складывается за счет повторения одной и той же архитектурной единицы или за счет повторения двух архитектурных единиц. Установлено, что в конкретных условиях обитания особенности архитектурных единиц, их комбинация и способ сочленения определяют жизненную форму тимьянов. В работе описано 4 жизненные формы, три из которых впервые приводятся в дополнение к классификации жизненных форм древесных и полудревесных растений И.Г. Серебрякова (Serebryakov, 1964).
Изучение структуры растений разных жизненных форм в настоящее время основывается на анализе архитектурных единиц. Архитектурная единица – это основная структурно-функциональная единица, содержащая полный набор всех иерархически соподчиненных структур и повторяющаяся в общей архитектуре растения (Barthélémy et al., 1989; Barthelemy, Caraglio, 2007). Она характеризуется определенным набором признаков: направление роста, расположение репродуктивных структур, особенности нарастания, положение почек возобновления, число и длина метамеров, элементарных побегов и т.д. Признаки архитектурной единицы у каждого вида растения стабильны и в зависимости от условий произрастания меняются только количественно (Caraglio, Edelin, 1990). В настоящее время выделены архитектурные единицы у деревьев, кустарников (Caraglio, Edelin, 1990; Sabatier, Barthélémy, 1999; Charles-Dominique et al., 2009; Antonova, Gnilovskaya, 2013; Kostina, 2015; Nedoseko, Viktorov, 2016; Gambino et al., 2016; и др.) и частично у трав (Notov, Kuznetzova, 2004; Tumidajowicz, 2005; Bartušková, Klimešová, 2010; Astashenkov, Cheryomushkina, 2016; Cheryomushkina, Guseva, 2017; и др.). Кустарнички и полукустарнички с позиции архитектурного подхода изучены слабо (Navarro et al., 2009, Götmark et al., 2016). Интерес в этом плане представляют виды рода Thymus. В качестве модельных видов выбраны T. extremus Klok., T. indigirkensis Karav., T. brevipetiolatus Čáp и T. pavlovii Serg., произрастающие в разных эколого-ценотических условиях Республики Саха (Якутия). Ранее у этих видов нами были описаны особенности развития и изучена онтогенетическая структура ценопопуляций (Talovskaya, 2018; Talovskaya et al., 2018). Цель работы – изучение архитектоники особей видов рода Thymus для выявления специфических черт жизненных форм и их положения в общей классификации жизненных форм древесных и полудревесных растений.
МАТЕРИАЛЫ И МЕТОДЫ
Изучены модельные виды T. extremus, T. indigirkensis, T. brevipetiolatus и T. pavlovii. Ранее нами установлено, что у видов формируются две жизненные формы (кустарничек и полукустарничек) и три типа биоморф (моноцентрический, неявно- и явнополицентрический) (Talovskaya et al., 2018). Определение жизненных форм видов проведено с использованием эколого-морфологической классификации жизненных форм И.Г. Серебрякова (Serebryakov, 1964). Тип биоморфы модельных видов установлен в соответствии с фитоценотической классификацией О.В. Смирновой (Smirnova, 1987), построенной на особенностях пространственного распределения побегов, почек возобновления и корней растения.
T. extremus – моноцентрический полукустарничек. Собран в окрестностях г. Якутска в разнотравно-типчаковом степном сообществе (Festuca lenensis Drobow, Artemisia pubescens Ledeb., Veronica incana L., Carex duriuscula C.A. Mey., Pulsatilla flavescens (Zucc.) Juz.), расположенном на пологом склоне юго-восточной экспозиции коренного берега р. Лены. Почва лугово-черноземная.
T. indigirkensis – неявнополицентрический полукустарничек. Собран в окрестностях пос. Сангар (Кобяйский р-н), в закустаренном Spiraea dahurica (Rupr.) Maxim. и Rosa acicularis Lindl. разнотравно-злаковом степном сообществе (Agropyron villosum (L.) Link, Festuca lenensis, Carex pediformis C.A. Mey., Dracocephalum palmatum Steph. ex Willd., T. indigirkensis) на склоне южной экспозиции коренного берега р. Лены. T. indigirkensis произрастал на мелкоземистой почве, покрытой щебнем.
T. pavlovii – явнополицентрический полукустарничек. Собран в Томпонском р-не, на склоне южной экспозиции хребта Сете-Дабаш, правого борта долины ручья Тый-Сынога. Склон представляет собой скально-осыпные обнажения, покрытые травяной растительностью (Bromopsis pumpelliana (Scribn.) Holub, Elymus confusus (Roshev.) Tzvelev, T. pavlovii, Dracocephalum stellerianum F. Hiltebr., Rhytidium rugosum (Hedw.) Kindb.); окружен смешанным лесом с участием Betula lanata (Regel) V.N. Vassil., Salix phylicifolia L., Spiraea dahurica, Dryas grandis Juz.
T. brevipetiolatus – неявнополицентрический кустарничек. Собран в окр. с. Смородичный (Кобяйский р-н) в прострело-типчаково-пырейной степи (Festuca lenensis, Elymus turuchanensis (Reverd.) Czerep., Pulsatilla patens (L.) Mill., Koeleria cristata (L.) Pers., Veronica incana) на склоне южной экспозиции. Почва – суглинок с небольшим количеством щебня.
На основе классификации М.Т. Мазуренко и А.П. Хохрякова (Mazurenko, Khokhryakov, 1977) при характеристике побеговой системы выделены разные типы побегов: обогащения, ветвления и формирования, а также составная скелетная ось. Составная скелетная ось – это совокупность многолетних побегов формирования, образующихся в результате моноподиально-симподиального нарастания. В зависимости от происхождения, выделены главная составная скелетная ось, которая сформировалась на основе первичного побега и имеет связь с главным корнем, и материнская составная скелетная ось – это любая ось n-го порядка. На главной и материнской составной скелетной оси развиваются оси 1-го (или n + 1), 2-го (или n + 2) и т.д. порядков. В работе использованы термины: розеточный, безрозеточный, полурозеточный побег (Serebryakov, 1959); верхнерозеточный побег (Nukhimovskii, 1997); среднерозеточный побег (Bobrov, 2009), розеточно-верхнерозеточный (Kolegova, Cheryomushkina, 2012).
Выделение архитектурных единиц проведено согласно представлениям D. Barthélémy, Y. Caraglio (2007). Архитектурная единица тимьянов состоит из главной (или материнской n-го порядка) составной скелетной оси, составной скелетной оси 1-го (или n + 1) порядка, одиночных побегов формирования 1-го порядка, побегов ветвления и обогащения.
Для сравнительного анализа у каждого вида было взято по 25 образцов разных архитектурных единиц. Учитывали число и длину побегов формирования в структуре составной скелетной оси; число боковых побегов и осей 1-го (или n + 1) порядка на главной (или материнской n-го порядка) составной скелетной оси. Проведена статистическая обработка данных: рассчитано среднее арифметическое и его ошибка, минимальное и максимальное значение признаков. Использованы компьютерные программы Excel, Statistica.
РЕЗУЛЬТАТЫ
Анализ взрослых особей T. extremus, T. indigirkensis, T. brevipetiolatus и T. pavlovii показал, что их структура складывалась в ходе развития из разнообразных архитектурных единиц. Выделено 4 архитектурные единицы, различающиеся по способу нарастания составной скелетной оси и положению в пространстве.
Архитектурная единица I
Архитектурная единица I выделена в структуре особей T. extremus и T. indigirkensis. Ее основой являются ортотропные моноподиально-базисимподиально нарастающие главная или материнская составная скелетная ось n-го порядка (рис. 1). Составная скелетная ось построена 4 побегами формирования, разнообразными по структуре, при этом моноподиальное нарастание каждого побега до трех лет. Также в состав архитектурной единицы I входят составные скелетные оси 1-го (или n + 1) порядка, побеги ветвления и обогащения. Сравнительный анализ показал, что большая часть боковых побегов развивается на двух последних побегах формирования составной скелетной оси (рис. 2). В основном это побеги ветвления, которые впоследствии отмирают вместе с частью побега формирования. Также на годичном приросте составной скелетной оси развиваются вегетативные или генеративные побеги обогащения. Составные скелетные оси 1-го порядка по структуре соответствуют материнской оси и сконцентрированы в основном в ее базальной части (первый по порядку побег формирования).
Рис. 1.
Развитие архитектурных единиц (АЕ) I и II (a–e – этапы развития архитектурной единицы, 1 – побег формирования, 2 – отмершая часть побега формирования, 3 – главная составная скелетная ось (или составная скелетная ось n-го порядка), 4 – составная скелетная ось 1-го (или n + 1) порядка, 5 – побег ветвления, 6 – побег формирования в структуре составной скелетной оси, 7 – побег обогащения).
Fig. 1. Development of architectural units (АЕ) I and II (а–e – stages of development of architectural unit, 1 – formation shoot, 2 – dead part of the formation shoot, 3 – the main compound skeletal axis (or compound skeletal axis of n-order), 4 – compound skeletal axis of the 1st (or of n + 1) order, 5 – branching shoot, 6 – formation shoot in compound skeletal axis structure, 7 – enrichment shoot).
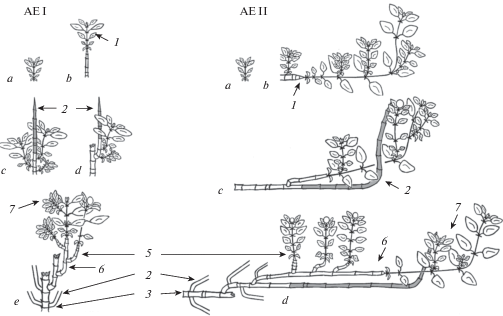
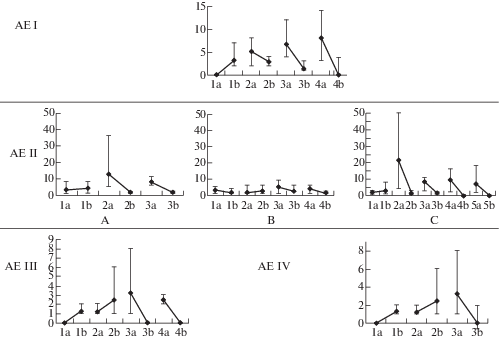
Рис. 2.
Число боковых побегов (одиночные побеги формирования n-го порядка, побеги ветвления и обогащения) (a) и составная скелетная ось n + 1 порядка (b) на каждом побеге формирования составной скелетной оси (ось X – порядковый номер побега формирования в структуре составной скелетной оси; ось Y – число побегов; для каждого побега формирования представлен диапазон (min, max) числа боковых побегов и осей, а также среднее значение (◆); A, B, C – варианты числа побегов формирования в составной скелетной оси АЕ II).
Fig. 2. Number of side shoots (single formation shoots of n-order, branching shoot, enrichment shoot) (а) and compound skeletal axis of n + 1 order (b) on each formation shoot of the compound skeletal axis (X-axis – serial number of a formation shoot in the compound skeletal axis structure; Y-axis – number of shoots: for each formation shoot there is a range (min, max) of the number of side shoots and axes, as well and an average value (◆); А, B, C – variants of formation shoot number in the compound skeletal axis of АЕ II).
Развитие архитектурной единицы I у T. extremus и T. indigirkensis сходно. Архитектурная единица может строиться на основе главной или материнской n-го порядка составной скелетной оси. Так, большая часть первичного побега, состоящего из двух (T. extremus)–трех (T. indigirkensis) годичных приростов, в зимний период отмирает, сохраняется лишь 2–3 базальных междоузлия или первый годичный прирост. Замещающий побег формирования развивается из верхней почки перезимовавшего участка и по типу развития повторяет предыдущий. Формируется моноподиально-базисимподиально нарастающая главная составная скелетная ось. В базальной части главной составной скелетной оси развиваются составные скелетные оси 1-го порядка, которые в дальнейшем станут материнскими осями n-го порядка. В средней части главной составной скелетной оси развертываются вегетативные побеги ветвления, а в апикальной – вегетативные побеги обогащения, отмирающие вместе с частью замещающего побега формирования. На 5 год развития у T. extremus и на 7 год у T. indigirkensis в структуре главной составной скелетной оси развиваются боковые генеративные побеги ветвления. Нарастание главной составной скелетной оси продолжается до 12 лет, с последующим отмиранием в течение 2–3 лет, исключая базальный участок, который сохраняется в течение жизни особи.
Развитие архитектурной единицы I на основе материнской составной скелетной оси n-го порядка происходит аналогично. Отличие заключается в том, что материнская составная скелетная ось построена вегетативными и генеративными побегами формирования. Ее отмирание происходит быстрее.
Архитектурная единица II
Архитектурная единица II описана у T. extremus, T. indigirkensis и T. pavlovii. Ее структура, в целом, соответствует структуре архитектурной единицы I. Она также состоит из главной (или материнской n-го порядка) составной скелетной оси, составной скелетной оси 1-го (или n + 1) порядка и побегов, различных по функциональной значимости. Главная (или материнская n-го порядка) составная скелетная ось отличается восходящим положением (рис. 1). У каждого вида она может быть построена разным числом побегов формирования: 2–3 у T. indigirkensis, 3–4 у T. extremus, 2–5 у T. pavlovii. Побеги формирования моноподиально нарастают два–пять лет. Как показал статистический анализ, определенной приуроченности боковых побегов к какой-либо части составной скелетной оси нет. Независимо от того, сколькими побегами формирования построена ось, боковые побеги развиваются на всем ее протяжении (рис. 2 А, B, C). В целом, их дифференциация соответствует боковым побегам архитектурной единицы I. В зависимости от длительности нарастания составной скелетной оси, составные скелетные оси 1-го порядка могут развиваться в ее базальной и средней частях.
Развитие архитектурной единицы II так же, как и развитие архитектурной единицы I, происходит на основе главной составной скелетной оси или на основе материнской составной скелетной оси n-го порядка. Первый способ выявлен у T. extremus и T. pavlovii. Первичный побег полегает, его моноподиальное нарастание продолжается 3–5 лет. Одновременно с ростом первичного побега развиваются и боковые побеги, большая часть из которых станет основой для формирования составной скелетной оси 1-го порядка. Смена моноподиального нарастания на базисимподиальное происходит одновременно у первичного и боковых побегов. В дальнейшем, каждый новый побег замещения в структуре главной составной скелетной оси или оси 1-го порядка будет повторять развитие предыдущего. Боковые генеративные побеги в структуре осей появляются на 4–5 год. Нарастание главной составной скелетной оси у T. extremus – 12 лет, у T. pavlovii – до 25. В дальнейшем ось постепенно отмирает от апикальной к базальной части.
Второй способ развития архитектурной единицы II отмечен у T. extremus, T. indigirkensis и T. pavlovii. Материнская составная скелетная ось n-го порядка развивается на основе восходящих побегов формирования n-го порядка ветвления. При этом у T. extremus и T. indigirkensis побеги формирования вегетативные или генеративные, у T. pavlovii – только вегетативные. Дальнейшее развитие архитектурной единицы II на основе материнской составной скелетной оси n-го порядка, в целом, соответствует развитию архитектурной единицы на основе главной составной скелетной оси.
Архитектурная единица III
Архитектурная единица III выделена в структуре T. brevipetiolatus. Ее основа – это главная (или материнская n-го порядка) ортотропная моноподиально-акросимподиально нарастающая составная скелетная ось, состоящая из 3–4 побегов формирования, моноподиальное нарастание каждого до пяти лет. Также в состав архитектурной единицы III входит составная скелетная ось 1-го (или n + 1) порядка, одиночные побеги формирования 1 порядка и побеги ветвления (рис. 3). Одиночные побеги формирования 1 порядка отличаются от побегов формирования в структуре главной составной скелетной оси сокращением моноподиального нарастания до 3–4 (5) лет. Они развиваются из спящих почек в средней части оси (второй-третий по порядку побег формирования). На первом по порядку побеге формирования главной (или материнской n-го порядка) составной скелетной оси одновременно с ее ростом развиваются составные скелетные оси 1-го (или n + 1) порядка, на последнем замещающем побеге формирования – развиваются побеги ветвления (рис. 2). Большинство осей и побегов переходят к цветению.
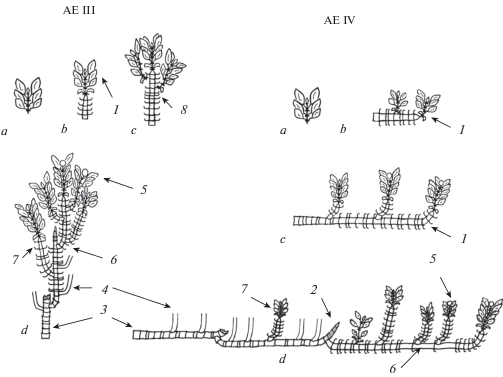
Особенность развития архитектурной единицы III: 1) первичный побег на пятый год переходит к цветению; 2) после плодоношения отмирает только соцветие, остальная часть первичного побега сохраняется и является основой для формирования моноподиально-акросимподиально нарастающей главной составной скелетной оси; 3) длительность нарастания главной составной скелетной оси до 20 лет; 4) отмирание главной составной скелетной оси происходит на протяжении 8–10 лет от апикальной части к базальной. Базальный участок оси сохраняется до отмирания всей особи; 5) развитие архитектурной единицы III на основе материнской составной скелетной оси n-го порядка происходит быстрее, в течение 9–16 лет.
Архитектурная единица IV
Архитектурная единица IV выделена в структуре T. brevipetiolatus. Отличие единицы заключается в том, что она строится на основе материнской плагиотропной моноподиально-акросимподиально нарастающей составной скелетной оси n-го порядка, состоящей из трех побегов формирования, моноподиальное нарастание каждого до десяти лет (рис. 3). Расположение боковых побегов и осей на составной скелетной оси, их структура сходны с архитектурной единицей III (рис. 2). При этом для большинства боковых побегов характерно ортотропное положение.
Особенности развития архитектурной единицы IV: 1) на 4 (5) год развития побега формирования n-го порядка он полегает и ветвится – его моноподиальное нарастание продолжается около 10 лет; 2) структура побегов формирования образована розеточными (редко верхнерозеточными) годичными побегами; 3) в конце своего развития побег формирования n-го порядка переходит к цветению и впоследствии становится основой для построения материнской плагиотропной моноподиально-акросимподиально нарастающей составной скелетной оси n-го порядка; 4) длительность нарастания оси может достигать 30 лет.
ОБСУЖДЕНИЕ
Изучение структуры модельных видов рода Thymus позволило выявить 4 варианта архитектурных единиц, которые отличаются по положению в пространстве и способу нарастания составных скелетных осей. По нашим данным подобные архитектурные единицы также встречаются в структуре Thymus marschallianus Willd. (Kolegova, 2013), T. altaicus Klok. et Schost., T. jenisseensis Iljin, T. minussinensis Serg. (Kolegova, Cheryomushkina, 2012), T. baicalensis Serg. (Talovskaia (Kolegova), 2015) и T. roseus Schipcz. (Talovskaya et al., 2017). Нами установлено, что структура особей может быть образована как за счет повторения одной и той же архитектурной единицы, так и за счет повторения двух архитектурных единиц. У модельных видов тимьянов выделено 5 комбинаций архитектурных единиц и 3 способа их сочленения (рис. 4).
Рис. 4.
Типы сочленения архитектурных единиц в структуре особей видов рода Thymus.
Fig. 4. Types of joining architectural units in individual structures of Thymus species.
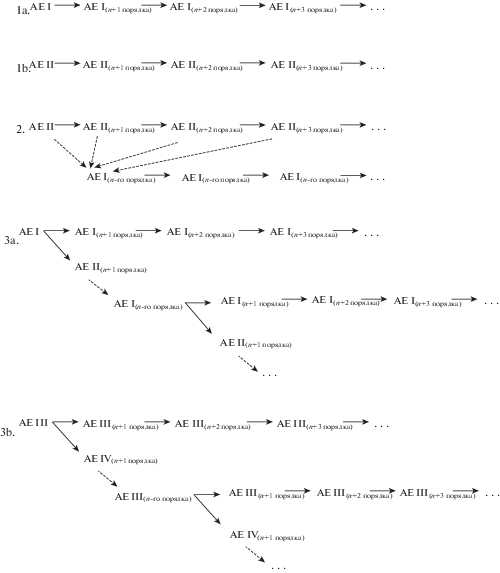
Однообразие архитектурных единиц и способа сочленения в структуре особей характерно для T. extremus и T. pavlovii. Структура первого складывается за счет архитектурной единицы I, на основе которой развиваются аналогичные архитектурные единицы следующих порядков (рис. 4, 1a). Такой способ построения приводит к формированию плотного куста. Иногда структура особей T. extremus может быть построена двумя вариантами – архитектурной единицей I и II. При этом архитектурная единица II является главной, а архитектурная единица I формируется на ее основе (рис. 4, 2). Это способствует формированию рыхлого куста. Структура особей T. pavlovii сложена архитектурными единицами II, сочлененными подобно архитектурным единицам I (рис. 4, 1b), что способствует формированию систем парциальных кустов и побегов.
Структура T. indigirkensis и T. brevipetiolatus устроена иначе: в построении особи принимают участие два варианта архитектурных единиц; способ их сочленения сложнее. Главной архитектурной единицей, как у T. indigirkensis, так и у T. brevipetiolatus является ортотропная единица I и III соответственно. На основе главной архитектурной единицы развиваются единицы двух вариантов: идентичные ортотропным архитектурные единицы n-го порядка и восходящие архитектурные единицы II у T. indigirkensis или плагиотропные архитектурные единицы IV у T. brevipetiolatus. Восходящая и плагиотропная архитектурные единицы являются основой для повторения соответствующей предыдущей структуры (рис. 4, 3a,b). Такой способ сложения архитектуры приводит к образованию куртины.
Анализ структуры особей модельных видов тимьянов в конкретных условиях обитания, показал, что особенности архитектурных единиц, их комбинация и способ сочленения определяют жизненную форму. Полученные результаты позволяют нам дополнить классификацию жизненных форм кустарничков и полукустарничков И.Г. Серебрякова (Serebryakov, 1964).
Так, в классе розеточных кустарничков выделен подкласс вегетативно-подвижные, ползучие; группа с ортотропными и плагиотропными надземными побегами; подгруппа полунеподвижные кустарнички. Среди изученных видов сюда относится T. brevipetiolatus – кустарничек с многолетними розеточными ортотропными и укореняющимися плагиотропными побегами и сохраняющейся в течение всей жизни особи системой главного корня.
Анализ литературы и собственных данных по розеточным кустарничкам показал, что к подгруппе полунеподвижных розеточных кустарничков, вероятно, следует отнести и Dracophyllum menziesii Hook. (Serebryakov, 1962), первичный корень, у которого сохраняется на протяжении всей жизни особи, а побеги ползут по поверхности субстрата. Кроме этого, в класс следует добавить еще один подкласс – вегетативно-неподвижные, прямостоячие; группу аэроксильные кустарнички. Представителем этой группы является Limonium chodshamumynense Lincz. et Czuk., произрастающий в условиях Южного Таджикистана на склонах карстовых скал горы Ходжа-Мумин (Cheryomushkina et al., 2012). Взрослая особь представляет собой прямостоячий первичный куст с симподиально нарастающими скелетными осями. Главный корень сохраняется в течение всей жизни особи.
У T. extremus, T. indigirkensis и T. pavlovii в условиях Якутии формируется жизненная форма полукустарничка. Куст взрослых особей T. extremus, произрастающих при высоком задернении, образован за счет многократного повторения ортотропной архитектурной единицы I. Жизненная форма вида, по классификации И.Г. Серебрякова (Serebryakov, 1964), соответствует прямостоячему полукустарничку. На свободных участках, где микрорельеф имеет ступенчатые понижения, в результате микросолифлюкции, структура особей образована сочетанием ортотропных и укореняющихся восходящих архитектурных единиц (I и II соответственно). Способ сочленения единиц, при котором восходящая архитектурная единица является основой для формирования ортотропных архитектурных единиц, не приводит к смене жизненной формы, куст взрослых особей становится лишь более рыхлым. Такая же рыхлая структура куста описана у прямостоячего полукустарничка Scutellaria supina, произрастающего в петрофитных вариантах настоящих и луговых степей (Cheryomushkina, Guseva, 2015).
Данные, полученные при изучении жизненных форм T. indigirkensis и T. pavlovii, позволяют дополнить классификацию полудревесных растений, включив их в подкласс стелющиеся полукустарники с удлиненными несуккулентными побегами.
Жизненная форма T. indigirkensis развивается в условиях горного склона. Куст взрослой особи так же, как и у полунеподвижного розеточного кустарничка T. brevipetiolatus, построен за счет многократного повторения двух архитектурных единиц I и II. Такая структура и особенности ее построения приводят к формированию новой жизненной формы среди полудревесных форм растений – стелющегося полунеподвижного полукустарничка с ортотропными и плагиотропными надземными побегами. Еще одной новой жизненной формой является форма ползучего полукустарничка, которая описана нами у T. pavlovii, произрастающего среди мха в расщелинах скальных обнажений. Структура особей устроена проще, за счет повторения восходящей архитектурной единицы (II). Составные скелетные оси, на основе которых строятся архитектурные единицы, интенсивно укореняются на всем протяжении. Вместе с этим, у особей T. pavlovii в зрелом генеративном состоянии происходит отмирание главного корня, что сближает его с настоящими ползучими растениями, а наличие ортотропных слаборазветвленных генеративных побегов – подтверждает его соответствие подгруппе.
ЗАКЛЮЧЕНИЕ
Использование архитектурного подхода при изучении жизненных форм модельных видов тимьянов позволило детально охарактеризовать их структуру. Выделено 4 архитектурные единицы, отличающиеся по способу нарастания составных скелетных осей и положению в пространстве. Показано, что архитектоника особей может быть образована как за счет повторения одной и той же архитектурной единицы, так и за счет повторения двух архитектурных единиц. Выделено 5 комбинаций архитектурных единиц и 3 способа их сочленения.
Установлено, что в конкретных условиях обитания особенности архитектурных единиц, их комбинация и способ сочленения определяют жизненную форму тимьянов. У T. indigirkensis и T. brevipetiolatus, произрастающих в сходных условиях – на горном склоне, структура особей складывается за счет повторения ортотропной и восходящей (или плагиотропной) архитектурной единицы. T. indigirkensis и T. brevipetiolatus представляют собой полунеподвижный полукустарничек и полунеподвижный розеточный кустарничек соответственно.
Также новая жизненная форма описана для T. pavlovii, произрастающего среди мха в расщелинах скальных обнажений. Структура особей устроена за счет повторения восходящей архитектурной единицы. Особенности архитектуры и развития особей позволяют охарактеризовать жизненную форму вида как ползучий полукустарничек. У T. extremus, произрастающего в условии высокого задернения, структура особей построена за счет многократного повторения ортотропной архитектурной единицы, что приводит к формированию жизненной формы прямостоячего полукустарничка.
Из 4 описанных жизненных форм три впервые приводятся нами в дополнение к классификации жизненных форм древесных (кустарничков) и полудревесных растений И.Г. Серебрякова (Serebryakov, 1964).
Список литературы
[Antonova, Gnilovskaya] Антонова И.С., Гниловская А.А. 2013. Побеговые системы Acer negundo (Acereceae) в разных возрастных состояниях. – Бот. журн. 98 (1): 53–68.
[Astashenkov, Cheryomushkina] Асташенков А.Ю., Черемушкина В.А. 2016. Архитектурная модель Nepeta mariae (Lamiaceae) – Растительный мир Азиатской России. 4 (24): 22–29. https://doi.org/10.21782/RMAR1995-2449-2016-4(22-29)
[Bobrov] Бобров Ю.А. 2009. Грушанковые России. Киров. 130 с.
Barthélémy D., Edelin C., Halle F. 1989. Architectural concepts for tropical trees. – In: Tropical forests: botanical dynamics, speciation and diversity. London. P. 89–100.
Barthélémy D., Caraglio Y. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. – Annals of botany. 99 (3): 375–407. https://doi.org/https://doi.org/10.1093/aob/mcl260
Caraglio Y., Edelin C. 1990. Architecture et dynamique de la croissance du platane. Platanus hybrida Brot. (Platanaceae) (syn. Platanus acerifolia (Aiton) Willd.). – Bulletin de la Société Botanique de France, Lettres botaniques. 137 (4-5): 279–291. https://doi.org/https://doi.org/10.1080/01811797.1990.10824889
Bartušková A., Klimešová J. 2010 Reiteration in the short lived root-sprouting herb Rorippa palustris: does the origin of buds matter? – Botany. 88 (7): 630–638. https://doi.org/https://doi.org/10.1139/B10-044
Charles-Dominique T., Edelin C., Bouchard A. 2010. Architectural strategies of Cornus sericea, a native but invasive shrub of Southern Quebec, Canada, under an open or a closed canopy. – Annals of Botany. 105 (2): 205–220. https://doi.org/ https://doi.org/10.1093/aob/mcp273
[Cheryomushkina et al.] Черемушкина В.А., Асташенков А.Ю., Бобоев М.Т. 2012. Морфогенез и состояние ценопопуляций Limonium chodshamumynense Lincz. et Czuk. в Южном Таджикистане. – Известия АН РТ. Отделение биологических и медицинских наук. 1: 7–12.
Cheryomushkina V.A., Guseva A.A. 2015. Life form of Scutellaria supina L. (Lamiaceae). – Contemporary Problems of Ecology. 8 (5): 756–769. https://doi.org/.https://doi.org/10.1134/S1995425515050054
[Cheryomushkina, Guseva] Черёмушкина В.А., Гусева А.А. 2017. Морфогенез Scutellaria grandiflora (Lamiaceae) и онтогенетическая структура его ценопопуляций. – Растительные ресурсы. 53 (3): 380–393.
Gambino S., Ratto F., Bartoli A. 2016. Architecture of the genus Gutierrezia (Asteraceae: Astereae, Solidagininae). – Boletin de Sociedad Argentina de Botanica. 51(4): 657–663.
Götmark F., Götmark E., Jensen A.M. 2016. Why be a shrub? A basic modeland hypotheses for the adaptive valuesofa common growth form. – Front. Plant Sci. https://doi.org/https://doi.org/10.3389/fpls.2016.01095
[Kolegova, Cheryomushkina] Колегова Е. Б., Черемушкина В.А. 2012. Структура побеговых систем видов рода Thymus (Lamiaceae) в Хакасии. – Бот. журн. 97 (2): 173–183.
[Kolegova] Колегова Е.Б. 2013. Структурные единицы побеговой системы Thymus marschallianus Willd. (Lamiaceae). – В сб.: Modern Phytomorphology 2nd Internatioonal Scientific Conference on Plant Morphology. Vol. 4. Lviv. P. 125–126.
Kostina M.V., Barabanshikova N.S., Bityugova G.V., Yasinskaya O.I., Dubach A.M. 2015. Structural modifications of birch (Betula pendula Roth.) crown in relation to environmental conditions. – Contemporary Problems of Ecology. 8 (5): 584–597. https://doi.org/https://doi.org/10.1134/S1995425515050091
[Mazurenko, Khokhryakov] Мазуренко М.Т., Хохряков А.П. 1977. Структура и морфогенез кустарников. М. 158 с.
Navarro T., Pascual V., Cabezudo B., Alados C. 2009. Architecture and functional traits of semi-arid shrub species in Cabo de Gata Natural Park, SE Spain. – Candollea. 64: 69–84. https://doi.org/https://doi.org/10.1016/j.jaridenv.2008.09.009
[Nedoseko, Viktorov] Недосеко О.И., Викторов В.П. 2016. Архитектурные модели Salix traindra L. и Salix fragilis L. – Социально-экологические технологии. 2: 39–50.
Notov A., Kusnetzova T. 2004. Architectural units, axiality and their taxonomic implications in Alchemillinae. – Wulfenia. 11: 85–130.
[Nukhimovskii] Нухимовский Е.Л. 1997. Основы биоморфологии семенных растений. Т. 1. М. 630 с.
Sabatier S. Br., Barthélémy D. 1999. Growth dynamics and morphology of annual shoots according to their architectural position in young Cedrus atlantica (Endl.) Manetti ex Carrie′re (Pinaceae). – Annals of botany. 84 (2): 387–392. https://doi.org/https://doi.org/10.1006/anbo.1999.0939
[Serebryakov] Серебряков И.Г. 1959. Типы развития побегов у травянистых многолетников и факторы их формирования. – Ученые записки МГПИ им. В.П. Потёмкина. Вопросы биологии растений. 100 (5): 3–38.
[Serebryakov] Серебряков И.Г. 1962. Экологическая морфология растений. М. 378 с.
[Serebryakov] Серебряков И.Г. 1964. Жизненные формы высших растений и их изучение. – Полевая геоботаника. 3: 146–208.
[Smirnova] Смирнова О.В. 1987. Структура травяного покрова широколиственных лесов. М. 205 с.
Talovskaya (Kolegova) E.B. 2015. Thymus baicalensis (Lamiaceae) morphological transformation under different environmental conditions. – Contemporary Problems of Ecology. 6: 736–749. https://doi.org/https://doi.org/10.1134/S1995425515050133
Talovskaya E.B. 2018. Ontogenetic structure of Thymus L. (Lamiaceae) coenopopulations in Yakutia. – In: BIO Web of Conferences “Prospects of Development and Challenges of Modern Botany. Vol. 11. Novosibirsk. https://doi.org/https://doi.org/10.1051/bioconf/20181100039
[Talovskaya et al.] Таловская Е.Б. Черёмушкина В.А., Асташенков А.Ю., Гусева А.А. 2017. Mорфологическая адаптация Thymus roseus (Lamiaceae) в Восточном Казахстане. – Бот. журн. 102(9): 1232–1248.
Talovskaya E.B., Cheryomushkina V.A., Denisova G.R. 2018. Morphological Adaptation of Species of the Genus Thymus (Lamiaceae) in Yakutia. – Contemporary Problems of Ecology. 8 (5): 607–613.
Tumidajowicz D. 2005. Architecture of the growth and development of the two alpine rhododendron species Rhododendron hirsutum and Rhododendron ferrugineum as a basis for the verification of the assessed relative and absolute age of aboveground shoots. – Oecologia Montana. 14:1–10.
Дополнительные материалы отсутствуют.
Инструменты
Ботанический журнал